

Abstract
The capacity of 20 healthy adult subjects for detecting differences in the amplitude of two simultaneously delivered 25 Hz vibrotactile stimuli was assessed both in the absence and presence of prior exposure to different conditions of adapting stimulation. Results obtained from this study demonstrate that increasing durations of adapting stimulation at one of the two skin sites, in the range of 0.2 to 2.0 seconds, leads to a systematic and progressive decrease in a subject’s ability to accurately discriminate between the two different amplitudes. Delivery of adapting stimuli to both of the sites of skin stimulation prior to simultaneous delivery of the test and standard stimuli, however, leads to an improvement in amplitude discrimination performance – a finding which is consistent with prior published psychophysical studies that demonstrate improvements in discriminatory capacity with much longer durations of adaptation. Striking parallels between the results obtained in this study and those reported in a prior study of the effects of vibrotactile adaptation on the optical response of squirrel monkey contralateral SI cortex to vibrotactile stimulation (Simons et al., 2007; Simons et al., 2005) suggest that the perceptual effects detected in this study could be attributable to adaptation-induced alterations of SI response.
1. Introduction
It is established that prolonged pre-exposure to sensory stimulation modifies discriminative capacity and alters the ability of both peripheral and CNS neurons to process sensory information. It is less widely appreciated, however, that primary sensory cortical mechanisms undergo transient, but significant alteration in response to even a brief exposure to adequate sensory stimulation. For example, both visual and somatosensory cortical pyramidal neurons undergo prominent use-dependent modifications of their receptive fields and response properties. Such modifications attain full development within a few tens of milliseconds of stimulus onset, and disappear within seconds after stimulus termination (visual cortical neurons: (Bredfeldt and Ringach, 2002; Celebrini et al., 1993; Das and Gilbert, 1995; DeAngelis et al., 1995; Dinse and Kruger, 1990; Pack and Born, 2001; Pettet and Gilbert, 1992; Ringach et al., 1997; Shevelev et al., 1998; Shevelev et al., 1992; Sugase et al., 1999); for recent review of short-term primary somatosensory cortical neuron dynamics see (Kohn and Whitsel, 2002)).
Recent optical intrinsic signal (OIS) imaging studies have characterized the short-term dynamics of the population-level response of squirrel monkey contralateral primary somatosensory (SI) cortex using different amplitudes and durations of vibrotactile stimulation (Chiu et al., 2005; Simons et al., 2007; Simons et al., 2005). Simons et al. (Simons et al., 2005) demonstrated that 25 Hz vibrotactile (flutter) skin stimulation at different amplitudes evoked an OIS response confined to the same local region of SI, but the magnitude of response varied proportionally with the stimulus amplitude. Additionally, Chiu et al. (Chiu et al., 2005) observed that the spatial pattern of response in SI on a sub-macrocolumnar scale changed systematically with increases in the amplitude of the stimulus. In subsequent studies, it was demonstrated that increasing stimulus duration (from 0.5 to 5.0 secs) led to differences not only in the peak magnitude of the evoked cortical response, but also in the relative rates of rise and decay of the magnitude of the evoked intrinsic signal (Chiu, 2006; Simons et al., 2007). Of particular note was the fact that relatively short duration stimuli (≤5 sec) evoked transient OIS responses which were persistent for several seconds (e.g., a 2 second stimulus duration evokes a response that persists above baseline for 5–6 seconds). This prolonged persistence of the OIS evoked in the contralateral SI cortex by increasing durations of skin flutter stimulation, as well as the observation that development of the spatial pattern of the evoked OIS response in SI is much more prominent and significant after 1–2 seconds than 0.5 seconds of stimulation (Chiu, 2006), led the authors to consider the possibility that the percept evoked in a conscious human subject by skin flutter stimulation might be significantly altered following a pre-exposure of the stimulus site to adapting stimulation for varying durations of stimulation less than or equal to 2 seconds.
The evidence obtained in this study (i) indicates that 25 Hz adapting stimulation modifies the ability of subjects to distinguish a difference in the amplitudes of two simultaneously-applied and otherwise identical skin flutter stimuli, and (ii) demonstrates that delivery of adapting stimulation to one or to both of the sites contacted by the dual-site test stimulus determines the sign of the effect of adaptation on subject performance (i.e., performance deteriorates when only one of the 2 sites receives the adapting stimulation, but is enhanced when adapting stimulation is delivered to both sites).
2. Results
A two alternative forced-choice (2AFC) tracking protocol was used to determine subjects’ capacities to discriminate between the amplitudes of two simultaneously delivered vibrotactile stimuli and to directly compare subjects’ discriminative capacities under different conditions of adapting stimulation.
Average across-subject (n = 20) performance during both blocks of the protocol is summarized by the “tracking plots” in Figure 1. The pattern of response for all subjects during Block #1 was highly stereotyped – i.e., in the initial trials each of the subjects easily identified the site that received the most intense stimulus and, as a result, the difference between the amplitudes of the test and standard stimuli was reduced on the subsequent trial. The decreasing slope of the average across-subject plot between trials 5–10 reflects the fact that the task became increasingly more difficult during these trials, and the leveling of the plot after trial 12 indicates that discrepancy between the test and standard stimuli at this point in the protocol was the smallest that each subject could detect (i.e., the minimally detectable amplitude difference). The average across-subject value of the minimally detectable amplitude difference determined in this way was approximately 23 μm (123 μmtest vs. 100 μm standard).
Figure 1.
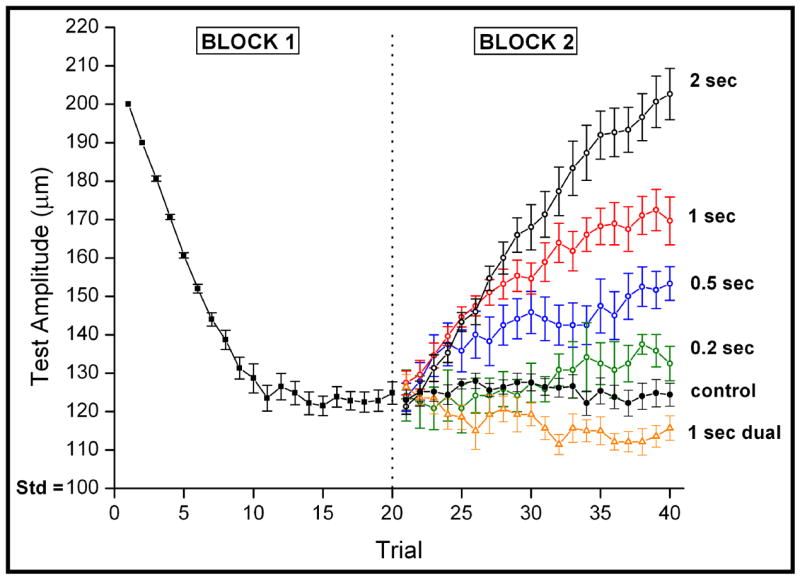
Comparison of averaged tracking response (with s.e. bars) to different conditions of adaptation. No adapting stimulus was applied to either stimulus site in Block 1 or for the control condition in Block 2. All other test conditions in Block 2 were preceded by either an adapting stimulus at the site of the test stimulus (ranging from 0.2 to 2 sec in duration) or by two 1 sec adapting stimuli delivered to both stimulus sites. Note that although dual site adapting stimulation leads to an improvement in performance, single site adapting stimulation leads to a progressive and systematic decrease in performance.
Four conditions in which adapting stimulation was presented only to the site of the test stimulus were studied in Block #2 of the protocol (the adapting stimulus duration implemented under these conditions was 2.0, 1.0, 0.5 or 0.2 sec, respectively). The impact of such single-site adaptation is made apparent by the tracking plots in Figure 1. For all durations of single-site adaptation, the subjects’ ability to detect a discrepancy between the amplitudes of the simultaneously applied skin flutter stimuli was prominently degraded. Furthermore, increasing the duration of single-site adapting stimulation was accompanied by a progressive decrease in the ability to detect a discrepancy in the amplitudes of the test and standard stimuli. Notably, however, the no-adaptation “control” condition was accompanied by no change in subject performance. It should be noted that the discrepancy between the 2.0 sec condition and the control condition is perhaps slightly larger than recorded, as the tracking of that condition has only begun to reach the initial stages of a plateau. Additionally, the effects of identical “dual-site” adapting stimulation (100 μm, 25 Hz adapting stimulation for 1 sec at both sites) are shown in Figure 1. In marked contrast to the effects of single-site adaptation, dual-site adaptation improves the ability to detect a discrepancy in the amplitudes of 2 otherwise identical, simultaneously-applied skin flutter stimuli.
To more directly compare the responses measured under the different conditions of adapting stimulation, the tracking values obtained from the last five trials of Block #2 across all subjects were averaged; the results are shown in Figure 2. In the control condition, no adapting stimulation was delivered and, as anticipated, the minimally detectable difference in amplitude did not change from the value determined in Block #1: i.e., it remained at ~23 μm. In contrast, (1) performance on the task improved in Block #2 under the condition where the standard and test stimuli were preceded by dual-site adapting stimulation: i.e., under this condition the amplitude difference was tracked to ~13 μm; and (2) when adapting stimulation was delivered only at the test location (single-site adaptation), performance worsened, and became progressively worse as the duration of single-site adapting stimulation was increased from 0.2 to 2 sec (the minimally detectable difference in amplitude was approximately 37, 49, 71, and 104 μm for the 0.2, 0.5, 1.0, and 2.0 sec durations of single-site adaptation, respectively).
Figure 2.
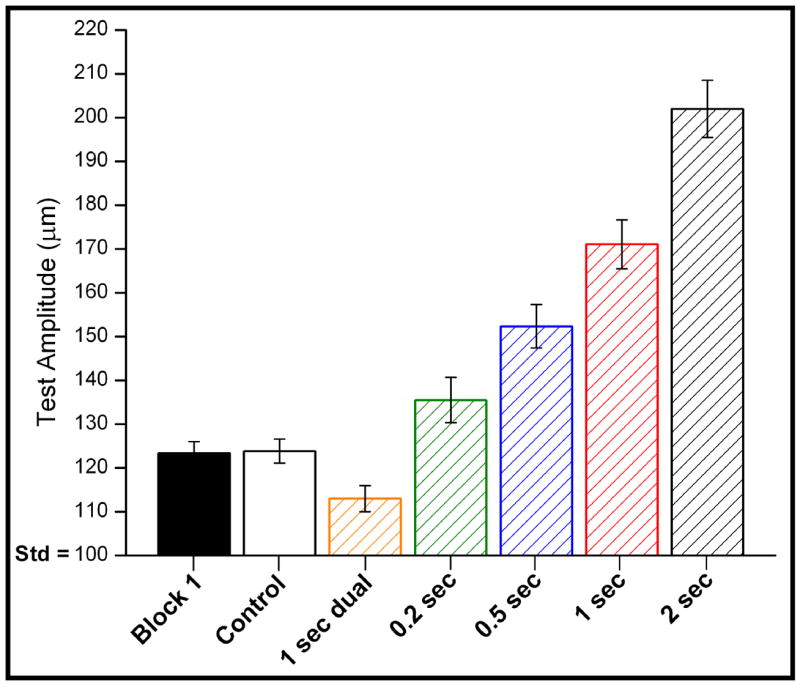
Averaged response of the different conditions (with s.e. bars). Averaging the outcome of the last 5 trials shows significant differences between each condition (see text for details).
Differences between subjects’ performance on the discrimination task in Block #1 and multiple conditions in Block #2 were analyzed for statistical significance using a one-way repeated measures ANOVA test. No statistically significant differences were found between subjects’ performance in Block #1 and Block #2 under conditions with no adaptation (control, p = 0.87). Conversely, statistically significant differences were confirmed between the non-adapting condition and condition with dual-site 1 sec adapting stimuli (p < 0.01). Furthermore, statistically significant differences were found across all Block #2 conditions of adaptation, including the non-adaptation condition (i.e., the effect with no adaptation was significantly less than 0.2 sec; the effect of 0.2 sec adaptation was significantly less than 0.5 sec; 0.5 sec had less effect than 1 sec; 2 sec adaptation had greater effect than 1 sec; p < 0.01 for all comparisons). Alternatively, a regression analysis of adaptor duration values versus amplitude difference limen values demonstrated a near-linear (R2 = 0.9870) correspondence.
3. Discussion
In this study, we have described the impact of adaptation on the ability of a subject to discriminate between the amplitude of two flutter stimuli, presented simultaneously to the dorsal surface of the hand. Our results demonstrate that subjects were able to discriminate stimuli which differed by ~23 μm in the absence of an adapting stimulus. When adapting stimuli were delivered at both sites, subjects demonstrated an improved ability to discriminate between the amplitudes of two simultaneously delivered stimuli. All subjects tested demonstrated a decreased ability in the amplitude discrimination task when the adapting stimulus was delivered to only one skin site prior to the presentation of the pair of test stimuli, presumably due to a decrease in the perceived intensity that was evoked at the skin site where the adapting stimulus was delivered. Additionally, each subject’s ability to correctly discriminate between the two amplitudes delivered to the skin was systematically and progressively degraded as the duration of the single-site adapting stimulus was increased. We view these findings to be in strong agreement with a large body of literature detailing not only the perceptual but also the neurophysiologic effects of adaptation. Moreover, the methodology used in this study, because of its ability to efficiently assess the impact of a conditioning stimulus, demonstrates a potential utility for the measurement of perceptual metrics of cortical function in a number of settings.
The effects of delivering an adapting stimulus on the perception of subsequent test stimuli – particularly the reduction in sensation - has been characterized in some detail (Delemos and Hollins, 1996; Gescheider et al., 1995; Goble and Hollins, 1993; Laskin and Spencer, 1979; Tommerdahl et al., 2005a; Verrillo and Gescheider, 1977). Many psychophysical studies have reported that the presentation of an adapting stimulus causes an increase in the detection threshold, and thus a reduction in the perceived intensity, of a subsequent stimulus (for review see (Gescheider et al., 1995; Goble and Hollins, 1993; Verrillo, 1985; Verrillo and Gescheider, 1977)). More specifically, Gescheider et al. showed that the threshold shift which occurred after the presentation of an adapting stimulus increased systematically with adapting stimulus duration (Gescheider et al., 1995). Fewer studies, however, have examined the impact of an adapting stimulus on the perceptual task of amplitude discrimination. Goble and Hollins reported that while an adapting stimulus increased the threshold for detection of a subsequent stimulus, it also enhanced a subject’s ability to discriminate subtle differences in the intensity of two subsequent (suprathreshold) stimuli which were presented serially to the same site (Goble and Hollins, 1993). One important difference of note between the protocol implemented in this study versus that in the Goble and Hollins study, in addition to simultaneously delivered stimuli, is that the duration of adaptation in that study was much longer (15 sec adapter followed by two 1 sec test stimuli). In this study, we demonstrated that much shorter stimulus durations were adequate to evoke robust changes in discriminative capacity. Additionally, this study demonstrates a systematic change in performance in which the impact of different durations of adapting stimulation shows a statistically significant change on a subject’s amplitude discriminative capacity, and it would be difficult to ascribe these differences to changes that occur in the periphery. The decrease in overall mean firing rate of RA afferents at the stimulus durations used in this study (≤ 2 secs) evoked changes that cannot account for the decrease in sensory performance. Most notably, Whitsel and colleagues (Whitsel et al., 2000) demonstrated that after 2 seconds of continuous stimulation, mean firing rate of RA afferents had decreased less than 10% - much less than could be accounted for by the changes observed in SI RA overall mean firing rate under the same conditions (for discussion, see Whitsel et al., 2002).
The effects of short-duration adaptation have been shown to change the performance of other aspects of a subject’s tactile discriminative capacity. For example, Tannan et al. (Tannan et al., 2006) recently demonstrated that the performance of neurologically healthy human subjects on a spatial localization task undergoes a prominent change with pre-task exposure to an adapting stimulus. In that study, it was determined that adaptation with a 5 sec stimulus resulted in an approximately 2-fold improvement in spatial localization performance over that achieved with a 0.5 sec adapting stimulus. It was proposed that this observed improvement in spatial discrimination was due to the enhanced spatial funneling of the population-level response of contralateral primary somatosensory cortex (SI) – a robust phenomenon that is at least in part due to GABAergic inhibitory neurotransmission and has been demonstrated using comparable stimulus conditions in neuroimaging studies of anesthetized non-human primates (Juliano et al., 1989). A subsequent report demonstrated that neurologically compromised subjects (adults with autism) showed no improvement at the spatial localization task with 5.0 sec adaptation although they outperformed healthy controls under the shorter duration 0.5 sec adaptation condition (Tommerdahl et al., 2007). One possible reason for this outcome could be related to the relatively recent demonstration that autism is associated with a mutation in regions centered around the GABAA-β3 receptor subunit gene, which has led some researchers (Belmonte et al., 2004; Polleux and Lauder, 2004) to suggest that the neocortical dysfunction in this disorder may be attributable to a deficiency during early development in GABA-mediated synaptic neurotransmission (see (Tommerdahl et al., 2007) for full discussion).
The fact that prolonged stimulation leads to a reduction in the response of neurons to subsequent stimuli at both the peripheral and central levels of neural processing is well documented (Bensmaia et al., 2005; Chung et al., 2002; O’Mara et al., 1988). Leung et al. demonstrated changes in the firing rate in the periphery at much longer stimulus durations (Leung et al., 2005) and showed that extended suprathreshold vibratory stimulation applied to the skin results in a desensitization of cutaneous mechanoreceptive afferents. Cannestra et al. reported that stimulus duration exerted a profound effect on the interval during which periods of reduced responsivity in the rat cortex could be observed during optical imaging studies (Cannestra et al., 1998). This reduction in cortical response would predictably lead to a reduction in the perceived magnitude of a subsequent stimulus, as a number of findings suggest that the cortical representation of the amplitude of a vibrotactile stimulus may be proportional to the magnitude of the population response in somatosensory cortex. For example, several studies have reported that the responses evoked – measured both via individual SI neurons as well as SI population measures – are systematically increased with an increase in the intensity of stimulation (Jousmaki and Forss, 1998; LaMotte and Mountcastle, 1975; Nelson et al., 2004; Simons et al., 2005; Tommerdahl et al., 2002; Torquati et al., 2002). Thus, the decreases in responsivity evoked by periods of prolonged stimulation (such as the adaptation intervals used in this study) result in a reduced response in the same neurons to a subsequent stimulus and may account for the decreases in a subject’s capacity for amplitude discrimination that were observed under the different conditions of single-site adapting stimuli.
Changes in the responsivity of neurons have been proposed to underlie the cortical mechanisms for stimulus feature extraction and may be important in the improvements observed in amplitude discrimination such as that demonstrated for the dual-site adaptation (for review see (Kohn et al., 2002)). This enhancement of discrimination capacity could be due at least in part to the moment-to-moment changes that occur in the spatio-temporal patterns of response with repetitive vibrotactile stimulation. For example, repetitive vibrotactile stimulation of the skin leads not only to a more funneled SI cortical response (Chen et al., 2003; Juliano et al., 1989; LaMotte and Mountcastle, 1975; Tommerdahl et al., 2002; Tommerdahl et al., 1993; Tommerdahl et al., 1996; Whitsel et al., 1991) but to changes in the temporal (Lee and Whitsel, 1992; Lee et al., 1992; Whitsel et al., 2003) as well as the spatial pattern of cortical activity evoked (Simons et al., 2007; Simons et al., 2005; Tommerdahl et al., 2002). Thus, the patterns of evoked SI cortical response maintain a number of spatial features that could provide the context in which subsequent stimuli are interpreted. Just as reduced responsivity in SI evoked by repetitive stimulation parallels a reduction in perceived intensity, increases in discriminative capacity could be paralleled by changes in the spatiotemporal pattern and the persistence of that response. Observations of the spatial patterns of SI cortical response within an activated cortical region, such as those evoked by flutter stimulation of the skin, suggest that evoked cortical activity within such a territory is not evenly distributed (Chiu et al., 2005; Kohn et al., 2000; McCasland and Woolsey, 1988; Tommerdahl et al., 1993; Tommerdahl et al., 2005b). It is important to note that while the magnitude of the cortical response increases with stimulus amplitude the spatial extent of the stimulus remains constant (Simons et al., 2005) and the spatial patterns within the activated cortical field change systematically and progressively with stimulus amplitude and duration (Chiu, 2006; Chiu et al., 2005). These changes in the spatial patterns of the evoked response – which are persistent (Simons et al., 2007) – could contribute to the improvements in discriminative performance that were observed with dual-site conditioning in this study. Consideration of the fact that adapting and/or repetitive vibrotactile stimuli lead to a reduction in the magnitude of the evoked cortical response (for review, see (McLaughlin and Kelly, 1993)) elevates the role that the spatial patterns of cortical response evoked by different amplitudes of vibrotactile stimulation could play in sensory perception.
What are the advantages of a technique such as the one described in this report? One overriding goal that we have in the development of all of our psychophysical protocols is the ultimate utility of providing not only the accurate assessment of sensory cortical function, but an efficient process that could be useful in a clinical and/or clinical research environment. The inherent advantage that comes with monitoring the percepts evoked by comparison of two simultaneously presented stimuli rather than single site could be significant. For example, the temporal confounds normally presented by stimuli delivered sequentially in a single skin site protocol could be prominent. Given that even the shorter duration adapting stimuli reported in this study had a significant impact on amplitude discriminative capacity, it is evident that the use of single-site protocols should, at the very least, require the significantly longer inter-stimulus intervals than the ones used in this study. It should be noted that the simultaneous amplitude discrimination task described in this report takes approximately 3–4 minutes per subject to complete, and this duration contrasts sharply with many traditional single-site stimulation psychophysical techniques which usually require significantly longer periods of time and are difficult, if not impossible, to implement in a clinical setting. Demonstration of measures – such as the one described in this report that are sensitive to changing conditions of adaptation – could prove useful for the study of subject populations that have systemic alterations of the cerebral cortex that compromise factors/mechanisms which contribute to the process of adaptation to a short duration repetitive stimulus. Current ongoing studies are investigating CNS compromised subject populations with such measures, and those results will be reported in subsequent papers.
4. Experimental procedures
Twenty subjects (22–31 years in age) were studied who were naïve both to the study design and issue under investigation. The subjects consisted of 13 males and 7 females, all right-hand dominant. All procedures were reviewed and approved in advance by an institutional review board.
A two-alternative forced-choice (2AFC) tracking protocol was used to evaluate the amplitude discriminative capacity of each subject. The subject was seated comfortably in a chair with the right arm resting on a table surface. The subject’s right hand was placed under a dual-site portable vibrotactile stimulator (CM-1; for full description, see (Tannan et al., 2007)). Two probe tips (5 mm diameter) were positioned 30 mm apart along a transversally oriented linear axis along the hand dorsum. The hand dorsum was selected to receive the stimulation because: 1) innervation density across this skin region remains relatively constant, 2) the surface is easily accessible and permits convenient stimulator placement, 3) the surface is relatively flat, reducing confounds of skin curvature present at other potential sites of stimulation, 4) it permits positioning of the subject’s arm and hand in a comfortable and stable position for the full duration of an experimental session and, perhaps most importantly, there is very little, if any between-subject use-dependent changes in sensitivity at this particular site. Previous studies have demonstrated that human subjects demonstrate very consistent performance with similar amplitude discrimination tasks on the hand dorsum (Tannan et al., 2007; Tannan et al., 2006; Tommerdahl et al, 2007). One significant aspect of those previous studies was that consistent results were obtained although stimulus positions were randomly located on a trial-by-trial basis, and thus, the relatively large size of the probe tip apparently compensates for the differential distribution of bone vs. muscle across the hand dorsum.
Visual cueing was provided with a computer monitor during the experimental run. Specifically, an array of LEDs was used to indicate to the subject when the stimulus was on and when the subject was to respond. The subject was not given performance feedback or knowledge of the results during the data acquisition until all sessions were completed. At the start of each run, the two probe tips were driven towards the skin until each tip registered a force of 0.1 g, as determined by a closed-loop algorithm in the CM-1 stimulator feedback system. The tips were then further indented into the skin by 500 μm to insure good contact with the skin. An audiometer was used to insure that no auditory cues were emitted from the stimulator during delivery of the range of stimuli used in this study.
The tracking protocol in each experimental run consisted of 2 sequential blocks. The first block of all experimental runs, in this report, was performed identically. In the first block, a vibrotactile test stimulus (25 Hz, amplitude between 105–200 μm) was delivered to one skin site at the same time that a standard stimulus (25 Hz, amplitude fixed at 100 μm) was applied to the other skin site. Previous studies have demonstrated that, for 25 Hz flutter stimuli, (i) the distance at which the two stimuli were positioned apart on the hand dorsum (30 mm) is well outside a subject’s two point limen (Tannan et al., 2005a; Tannan et al., 2005b; Tannan et al., 2006) and (ii) at a 30 mm probe separation there is no difference in the ability of a subject to detect a difference in the amplitudes of flutter stimulation applied simultaneously or sequentially to the 2 skin sites (Tannan et al., 2007). The loci of the test and standard stimuli were randomly selected on a trial-by-trial basis. Stimulus duration was 0.5 sec, followed by subject response (the subject was queried to select, using a two-button switchbox, the skin site that received the most intense stimulus) and a 5 sec delay before onset of the next trial. At the beginning of the experimental run, the test stimulus was 200 μm (peak-to-peak amplitude) and the standard was 100 μm. In the initial 10 trials the amplitude of the test stimulus was modified based on the subject’s response to the preceding trial – accomplished using a 1-up/1-down forced-choice tracking protocol. This approach was selected because it enabled rapid determination (“tracking”) of each subject’s minimally detectable difference in the amplitudes of two-site skin flutter stimulation (Tannan et al., 2007). The difference in the amplitudes of the test and standard stimuli delivered on each of these initial 10 trials was adjusted on the basis of the subject’s response in the preceding trial (the discrepancy in amplitude was decreased if the subject’s response in the preceding trial was correct; it was increased if the response was incorrect). After the initial 10 trials were completed, test stimulus amplitude was modified using a 2-up/1-down protocol – in these trials two correct/one incorrect subject response(s) resulted in a decrement/increment, respectively, in the amplitude difference between the test and standard stimuli. The step size was held constant throughout all experimental runs at 10 μm.
In the second block, delivery of the test and standard stimuli was preceded by one of 6 different conditions of adapting stimulation. In the first condition, no adapting stimulus was delivered (control condition). In four of the conditions, a 25 Hz 100 μm adapting stimulus at the location of the test stimulus was delivered 1 sec prior to the presentation of the test and standard stimuli. By presenting the adapting stimulus to the same site as the test stimulus, it was possible to quantify the effect of reduced perceived intensity due to varying durations of adapting stimulation. Four different durations of adapting stimuli were delivered: 0.2, 0.5, 1 and 2 sec. The final condition consisted of a 25 Hz 100 μm adapting stimulus being delivered for 1 sec to both skin sites prior to the delivery of the test and standard stimuli. In all six conditions, a 2-up/1-down protocol was used in Block #2 to track the subject’s ability to determine the most intense stimulus. The initial conditions of Block #2 were the final conditions of Block #1. A series of training trials, each consisting of a pair of stimuli differing in amplitude by 100 μm (200 μm vs. 100 μm), were conducted prior to the first session. The subject was provided with feedback only during training trials and was allowed to continue on to the first session after answering correctly 5 times in a row. Each subject participated in a single experimental session that consisted of 6 separate runs of different adapting stimulus conditions (randomized in order). A single session, including actual testing time and short breaks between each run, took 45–60 minutes.
Figure 3.

Schematic of the protocols used for amplitude discrimination. Two blocks of stimulus delivery were employed. In the first block, two 25 Hz vibrotactile stimuli, the standard (S) and test (T), were delivered at the same time for 0.5 sec. A 5 sec delay (excluding subject response interval (RI)) was imposed before onset of the next trial. In the second block, varying conditions of single-site or dual-site adapting stimulation were first delivered, followed by a 1 sec inter-stimulus interval, and then the standard and test stimuli.
Acknowledgments
This work was supported, in part by NIH R01 grant NS043375 (M. Tommerdahl, P.I.) and a grant from the Cure Autism Now Foundation (M. Tommerdahl, P.I.).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bensmaia SJ, Leung YY, Hsiao SS, Johnson KO. Vibratory adaptation of cutaneous mechanoreceptive afferents. J Neurophysiol. 2005;94:3023–36. doi: 10.1152/jn.00002.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bredfeldt CE, Ringach DL. Dynamics of spatial frequency tuning in macaque V1. J Neurosci. 2002;22:1976–84. doi: 10.1523/JNEUROSCI.22-05-01976.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cannestra AF, Pouratian N, Shomer MH, Toga AW. Refractory periods observed by intrinsic signal and fluorescent dye imaging. J Neurophysiol. 1998;80:1522–32. doi: 10.1152/jn.1998.80.3.1522. [DOI] [PubMed] [Google Scholar]
- Celebrini S, Thorpe S, Trotter Y, Imbert M. Dynamics of orientation coding in area V1 of the awake primate. Vis Neurosci. 1993;10:811–25. doi: 10.1017/s0952523800006052. [DOI] [PubMed] [Google Scholar]
- Chen LM, Friedman RM, Roe AW. Optical imaging of a tactile illusion in area 3b of the primary somatosensory cortex. Science. 2003;302:881–5. doi: 10.1126/science.1087846. [DOI] [PubMed] [Google Scholar]
- Chiu J. Biomedical Engineering. University of North Carolina at Chapel Hill; Chapel Hill: 2006. Characterization of Minicolumnar Patterns in SI Cortex. [Google Scholar]
- Chiu JS, Tommerdahl M, Whitsel BL, Favorov OV. Stimulus-dependent spatial patterns of response in SI cortex. BMC Neurosci. 2005;6:47. doi: 10.1186/1471-2202-6-47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung S, Li X, Nelson SB. Short-term depression at thalamocortical synapses contributes to rapid adaptation of cortical sensory responses in vivo. Neuron. 2002;34:437–46. doi: 10.1016/s0896-6273(02)00659-1. [DOI] [PubMed] [Google Scholar]
- Das A, Gilbert CD. Receptive field expansion in adult visual cortex is linked to dynamic changes in strength of cortical connections. J Neurophysiol. 1995;74:779–92. doi: 10.1152/jn.1995.74.2.779. [DOI] [PubMed] [Google Scholar]
- DeAngelis GC, Anzai A, Ohzawa I, Freeman RD. Receptive field structure in the visual cortex: does selective stimulation induce plasticity? Proc Natl Acad Sci U S A. 1995;92:9682–6. doi: 10.1073/pnas.92.21.9682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delemos KA, Hollins M. Adaptation-induced enhancement of vibrotactile amplitude discrimination: the role of adapting frequency. J Acoust Soc Am. 1996;99:508–16. doi: 10.1121/1.414509. [DOI] [PubMed] [Google Scholar]
- Dinse HR, Kruger K. Contribution of area 19 to the foreground-background-interaction of the cat: an analysis based on single cell recordings and behavioural experiments. Exp Brain Res. 1990;82:107–22. doi: 10.1007/BF00230843. [DOI] [PubMed] [Google Scholar]
- Gescheider GA, Santoro KE, Makous JC, Bolanowski SJ. Vibrotactile forward masking: effects of the amplitude and duration of the masking stimulus. J Acoust Soc Am. 1995;98:3188–94. doi: 10.1121/1.413808. [DOI] [PubMed] [Google Scholar]
- Goble AK, Hollins M. Vibrotactile adaptation enhances amplitude discrimination. J Acoust Soc Am. 1993;93:418–24. doi: 10.1121/1.405621. [DOI] [PubMed] [Google Scholar]
- Jousmaki V, Forss N. Effects of stimulus intensity on signals from human somatosensory cortices. Neuroreport. 1998;9:3427–31. doi: 10.1097/00001756-199810260-00017. [DOI] [PubMed] [Google Scholar]
- Juliano SL, Dusart I, Peschanski M. Somatic activation of thalamic neurons transplanted into lesioned somatosensory thalamus. Brain Res. 1989;478:356–60. doi: 10.1016/0006-8993(89)91516-3. [DOI] [PubMed] [Google Scholar]
- Kohn A, Metz C, Quibrera M, Tommerdahl MA, Whitsel BL. Functional neocortical microcircuitry demonstrated with intrinsic signal optical imaging in vitro. Neuroscience. 2000;95:51–62. doi: 10.1016/s0306-4522(99)00385-1. [DOI] [PubMed] [Google Scholar]
- Kohn A, Metz C, Tommerdahl MA, Whitsel BL. Stimulus-evoked modulation of sensorimotor pyramidal neuron EPSPs. J Neurophysiol. 2002;88:3331–47. doi: 10.1152/jn.01012.2001. [DOI] [PubMed] [Google Scholar]
- Kohn A, Whitsel BL. Sensory cortical dynamics. Behav Brain Res. 2002;135:119–26. doi: 10.1016/s0166-4328(02)00139-0. [DOI] [PubMed] [Google Scholar]
- LaMotte RH, Mountcastle VB. Capacities of humans and monkeys to discriminate vibratory stimuli of different frequency and amplitude: a correlation between neural events and psychological measurements. J Neurophysiol. 1975;38:539–59. doi: 10.1152/jn.1975.38.3.539. [DOI] [PubMed] [Google Scholar]
- Laskin SE, Spencer WA. Cutaneous masking. I. Psychophysical observations on interactions of multipoint stimuli in man. J Neurophysiol. 1979;42:1048–60. doi: 10.1152/jn.1979.42.4.1048. [DOI] [PubMed] [Google Scholar]
- Lee CJ, Whitsel BL. Mechanisms underlying somatosensory cortical dynamics: I. In vivo studies. Cereb Cortex. 1992;2:811–06. doi: 10.1093/cercor/2.2.81. [DOI] [PubMed] [Google Scholar]
- Lee CJ, Whitsel BL, Tommerdahl M. Mechanisms underlying somatosensory cortical dynamics: II. In vitro studies. Cereb Cortex. 1992;2:107–33. doi: 10.1093/cercor/2.2.107. [DOI] [PubMed] [Google Scholar]
- Leung YY, Bensmaia SJ, Hsiao SS, Johnson KO. Time-course of vibratory adaptation and recovery in cutaneous mechanoreceptive afferents. J Neurophysiol. 2005;94:3037–45. doi: 10.1152/jn.00001.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCasland JS, Woolsey TA. High-resolution 2-deoxyglucose mapping of functional cortical columns in mouse barrel cortex. J Comp Neurol. 1988;278:555–69. doi: 10.1002/cne.902780407. [DOI] [PubMed] [Google Scholar]
- McLaughlin DF, Kelly EF. Evoked potentials as indices of adaptation in the somatosensory system in humans: a review and prospectus. Brain Res Brain Res Rev. 1993;18:151–206. doi: 10.1016/0165-0173(93)90001-g. [DOI] [PubMed] [Google Scholar]
- Nelson AJ, Staines WR, Graham SJ, McIlroy WE. Activation in SI and SII: the influence of vibrotactile amplitude during passive and task-relevant stimulation. Brain Res Cogn Brain Res. 2004;19:174–84. doi: 10.1016/j.cogbrainres.2003.11.013. [DOI] [PubMed] [Google Scholar]
- O’Mara S, Rowe MJ, Tarvin RP. Neural mechanisms in vibrotactile adaptation. J Neurophysiol. 1988;59:607–22. doi: 10.1152/jn.1988.59.2.607. [DOI] [PubMed] [Google Scholar]
- Pack CC, Born RT. Temporal dynamics of a neural solution to the aperture problem in visual area MT of macaque brain. Nature. 2001;409:1040–2. doi: 10.1038/35059085. [DOI] [PubMed] [Google Scholar]
- Pettet MW, Gilbert CD. Dynamic changes in receptive-field size in cat primary visual cortex. Proc Natl Acad Sci U S A. 1992;89:8366–70. doi: 10.1073/pnas.89.17.8366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ringach DL, Hawken MJ, Shapley R. Dynamics of orientation tuning in macaque primary visual cortex. Nature. 1997;387:281–4. doi: 10.1038/387281a0. [DOI] [PubMed] [Google Scholar]
- Shevelev IA, Eysel UT, Lazareva NA, Sharaev GA. The contribution of intracortical inhibition to dynamics of orientation tuning in cat striate cortex neurons. Neuroscience. 1998;84:11–23. doi: 10.1016/s0306-4522(97)00363-1. [DOI] [PubMed] [Google Scholar]
- Shevelev IA, Volgushev MA, Sharaev GA. Dynamics of responses of V1 neurons evoked by stimulation of different zones of receptive field. Neuroscience. 1992;51:445–50. doi: 10.1016/0306-4522(92)90328-y. [DOI] [PubMed] [Google Scholar]
- Simons SB, Chiu J, Favorov OV, Whitsel BL, Tommerdahl M. Duration-dependent response of SI to vibrotactile stimulation in squirrel monkey. J Neurophysiol. 2007;97:2121–9. doi: 10.1152/jn.00513.2006. [DOI] [PubMed] [Google Scholar]
- Simons SB, Tannan V, Chiu J, Favorov OV, Whitsel BL, Tommerdahl M. Amplitude-dependency of response of SI cortex to flutter stimulation. BMC Neurosci. 2005;6:43. doi: 10.1186/1471-2202-6-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugase Y, Yamane S, Ueno S, Kawano K. Global and fine information coded by single neurons in the temporal visual cortex. Nature. 1999;400:869–73. doi: 10.1038/23703. [DOI] [PubMed] [Google Scholar]
- Tannan V, Dennis R, Tommerdahl M. A novel device for delivering two-site vibrotactile stimuli to the skin. J Neurosci Methods. 2005a;147:75–81. doi: 10.1016/j.jneumeth.2005.05.001. [DOI] [PubMed] [Google Scholar]
- Tannan V, Dennis RG, Tommerdahl M. Stimulus-dependent effects on tactile spatial acuity. Behav Brain Funct. 2005b;1:18. doi: 10.1186/1744-9081-1-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tannan V, Dennis RG, Zhang Z, Tommerdahl M. A portable tactile sensory diagnostic device. J Neurosci Methods. 2007;164:131–8. doi: 10.1016/j.jneumeth.2007.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tannan V, Whitsel BL, Tommerdahl MA. Vibrotactile adaptation enhances spatial localization. Brain Res. 2006;1102:109–16. doi: 10.1016/j.brainres.2006.05.037. [DOI] [PubMed] [Google Scholar]
- Tommerdahl M, Favorov O, Whitsel BL. Optical imaging of intrinsic signals in somatosensory cortex. Behav Brain Res. 2002;135:83–91. doi: 10.1016/s0166-4328(02)00159-6. [DOI] [PubMed] [Google Scholar]
- Tommerdahl M, Favorov O, Whitsel BL, Nakhle B, Gonchar YA. Minicolumnar activation patterns in cat and monkey SI cortex. Cereb Cortex. 1993;3:399–411. doi: 10.1093/cercor/3.5.399. [DOI] [PubMed] [Google Scholar]
- Tommerdahl M, Hester KD, Felix ER, Hollins M, Favorov OV, Quibrera PM, Whitsel BL. Human vibrotactile frequency discriminative capacity after adaptation to 25 Hz or 200 Hz stimulation. Brain Res. 2005a;1057:1–9. doi: 10.1016/j.brainres.2005.04.031. [DOI] [PubMed] [Google Scholar]
- Tommerdahl M, Simons SB, Chiu JS, Favorov O, Whitsel B. Response of SI cortex to ipsilateral, contralateral and bilateral flutter stimulation in the cat. BMC Neurosci. 2005b;6:29. doi: 10.1186/1471-2202-6-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tommerdahl M, Tannan V, Cascio CJ, Baranek GT, Whitsel BL. Vibrotactile adaptation fails to enhance spatial localization in adults with autism. Brain Res. 2007;1154:116–23. doi: 10.1016/j.brainres.2007.04.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tommerdahl M, Whitsel BL, Vierck CJ, Jr, Favorov O, Juliano S, Cooper B, Metz C, Nakhle B. Effects of spinal dorsal column transection on the response of monkey anterior parietal cortex to repetitive skin stimulation. Cereb Cortex. 1996;6:131–55. doi: 10.1093/cercor/6.2.131. [DOI] [PubMed] [Google Scholar]
- Torquati K, Pizzella V, Della Penna S, Franciotti R, Babiloni C, Rossini PM, Romani GL. Comparison between SI and SII responses as a function of stimulus intensity. Neuroreport. 2002;13:813–9. doi: 10.1097/00001756-200205070-00016. [DOI] [PubMed] [Google Scholar]
- Verrillo RT. Psychophysics of vibrotactile stimulation. J Acoust Soc Am. 1985;77:225–32. doi: 10.1121/1.392263. [DOI] [PubMed] [Google Scholar]
- Verrillo RT, Gescheider GA. Effect of prior stimulation on vibrotactile thresholds. Sens Processes. 1977;1:292–300. [PubMed] [Google Scholar]
- Whitsel BL, Favorov O, Kelly DG, Tommerdahl M. Mechanisms of dynamic peri- and intracolumnar interactions in somatosensory cortex: stimulus specific contrast enhancement by NMDA receptor activation. In: Franzen O, Westman J, editors. Information Processing in Somatosensory System. Macmillan Press; London: 1991. pp. 353–369. [Google Scholar]
- Whitsel BL, Kelly EF, Delemos KA, Xu M, Quibrera M. Stability of rapidly adapting afferent entrainment vs responsivity. Somatosens Mot Res. 2000;17:13–31. doi: 10.1080/08990220070265. [DOI] [PubMed] [Google Scholar]
- Whitsel BL, Kelly EF, Quibrera M, Tommerdahl M, Li Y, Favorov OV, Xu M, Metz CB. Time-dependence of SI RA neuron response to cutaneous flutter stimulation. Somatosens Mot Res. 2003;20:45–69. doi: 10.1080/0899022031000083834. [DOI] [PubMed] [Google Scholar]
- Whitsel BL, Kelly EF, Xu M, Tommerdahl M, Quibrera M. Frequency-dependent response of SI RA-class neurons to vibrotactile stimulation of the receptive field. Somatosens Mot Res. 2001;18:263–285. doi: 10.1080/01421590120089659. [DOI] [PubMed] [Google Scholar]