

Abstract
Soluble cytokine receptors modulate the activity of their cognate ligands. Interleukin (IL)-6 in association with the soluble IL-6 receptor (sIL-6R) can activate cells expressing the gp130 signal transducer lacking the specific IL-6R. To investigate the function of the IL-6–sIL-6R complex in vivo and to discriminate the function of the IL-6–sIL-6R complex from the function of IL-6 alone, we have established a transgenic mouse model. Double-transgenic mice coexpressing IL-6 and sIL-6R were generated and compared with IL-6 and sIL-6R single-transgenic mice. The main phenotype found in IL-6–sIL-6R mice was a dramatic increase of extramedullary hematopoietic progenitor cells in liver and spleen but not in the bone marrow. In IL-6 single-transgenic mice and sIL-6R single-transgenic mice no such effects were observed. The high numbers of hematopoietic progenitor cells were reflected by a strong increase of peripheral blood cell numbers. Therefore, activators of the gp130 signal transducer like the IL-6–IL-6R complex may represent most powerful stimulators for extramedullary hematopoietic progenitor cells. gp130 activators may become important for the expansion of hematopoietic progenitor cells in vivo and in vitro.
IL-6 is a mediator of hematopoietic cell growth and differentiation acting on B cells, T cells, keratinocytes, neuronal cells, osteoclasts, and endothelial cells (for reviews, see references 1 and 2). In the liver, IL-6 modulates the transcription of acute phase response genes during acute and chronic inflammatory states (3).
The IL-6–type cytokine family comprises IL-6, IL-11, ciliary neurotrophic factor (CNTF)1, leukemia inhibitory factor (LIF), oncostatin M (OSM), and cardiotrophin-1 (CT1). The biological action of these cytokines is mediated by multisubunit cell surface receptors that share a common signaling subunit, the gp130 signal-transducing molecule (4, 5). The subunits of these receptors are members of the cytokine/growth hormone receptor family that have conserved cysteine and tryptophane residues in the extracellular domain and that signal via the JAK–STAT pathway (6–8). IL-6 and IL-11 use a gp130 homodimer and OSM, CNTF, LIF, and CT-1 use a heterodimer of gp130 and the 190 kD LIF receptor for the activation of intracellular signaling cascades (4). The receptor signaling complexes for CNTF, IL-6, IL-11, and probably CT-1 (9) contain an additional ligand specific subunit that is not required for LIF and OSM binding. Recently, a separate OSM-specific receptor has been identified that also forms a heterodimer with gp130 (10).
Many soluble receptors (sR), like TNF-R, IL-1R, and IL-4R, act antagonistically, competing with membrane receptors for the binding of the cytokines (11–13). However, some agonistic cytokine–sR complexes stimulate signal-transducing receptors, rendering cells sensitive to the respective cytokines that lack specific cytokine receptors on their surface. Such cells on their own would not be able to respond to the respective cytokine alone. This pathway, recently termed transignaling, is used by the cytokines of the IL-6 family (14, 15).
On target cells, IL-6 first binds to a specific IL-6 receptor (IL-6Rα) (16). This IL-6–IL-6R complex induces the homodimerization of two gp130 molecules (17, 18), leading to intracellular signaling events. Soluble forms of the IL-6R complex are generated by limited proteolysis from the cell surface (19, 20). Elevated concentrations can be found in inflammatory diseases (21–24). In vivo, the sIL-6R acts as a serum-binding protein for IL-6 and prolongs the plasma half life of IL-6 (25). However, the biologic function of the IL-6–IL-6R complex in vivo is unknown.
To discriminate between biological functions of IL-6 alone and those of the IL-6–IL-6R complex, double-transgenic mice coexpressing human IL-6 and the human soluble IL-6R (IL-6–sIL-6R mice) were generated and compared with sIL-6R (25) and IL-6 (26) single-transgenic mice and nontransgenic littermates. There were striking differences regarding the phenotypes of single and doubletransgenic mice. IL-6–sIL-6R mice were smaller and displayed a marked hepatosplenomegaly caused by an extreme expansion of extramedullary hematopoietic progenitor cells. No such phenotype was observed in single-transgenic and nontransgenic littermates. This study identifies the gp130 signal-transducing protein to be a most powerful mediator of hematopoietic cell growth and differentiation in vivo.
Materials and Methods
Generation of Transgenic Mice.
The generation of human sIL-6R and human IL-6 transgenic mice has been described in detail elsewhere (25, 26). By crossing homozygous sIL-6R and IL-6 mice, hemizygous double transgenic mice (IL-6–sIL-6R mice) were generated that coexpressed both transgenes. Expression of the sIL-6R transgene was driven by the neonatal phosphoenolpyruvate carboxykinase (PEPCK) promoter, which is transcriptionally activated at day 3 after birth and does not produce intrauterine developmental effects (25). The IL-6 transgene was driven by the murine metallothionein-1 promoter (26). Serum concentration of IL-6 ranged between 10 and 20 ng/ml in IL-6 and IL-6–sIL-6R mice; concentrations of sIL-6R ranged between 4 and 8 μg/ml in sIL-6R and IL-6–sIL-6R mice (data not shown).
Antibodies.
The following antibodies were used: anti-Gr-1 (RB6-8C5), Mac-1 (M1/70), B220 (RA3-6B2), CD4 (H129.19), CD8a (53-6.7) (all PharMingen, San Diego, CA). mAbs were used as lineage markers. Anti-c-kit (2B8) and Sca-1 (E13-161.7) (both PharMingen) were used as hematopoietic stem cell markers. FITC- or PE-conjugated antibodies were used.
Cell Sorting and Flow Cytometric Analysis.
Freshly isolated spleen, liver, and bone marrow cells were obtained according to methods previously described (27). Cells were washed three times with RPMI 1640 medium and suspended at a concentration of 2 × 105 cells/ml of PBS containing 2.5% FCS and 0.05% sodium azide before use. Double staining using anti-CD45 (30F11.1) (PharMingen) mAbs and various lineage and hematopoietic stem cell–specific mAbs was performed. The cell suspensions were incubated for 30 min on ice with mAbs against cell surface molecules in PBS containing 2.5% FCS and washed three times with the same buffer. The fluorescence of the cells were examined using a FACScan® (Becton Dickinson, Mountain View, CA).
Organ Weight Recording and Histomorphological Analysis.
At ages indicated in the figures, three mice each of single-transgenic and nontransgenic littermates and IL-6–sIL-6R mice were sacrificed, and liver, spleen, and total body weights were recorded. Mean values of the ratio of organ weight:body weight are shown. Organs were fixed in 4% formaldehyde/PBS and embedded in paraffin. 5-μm sections were stained with haematoxylin–eosin.
Immunohistochemistry.
Immunohistochemistry was performed using the indirect peroxidase staining method. Rat anti–mouse Ly-6, rat anti–mouse CD3, rat anti–mouse B220, (PharMingen), and rat anti–mouse Ia (Boehringer Mannheim, Germany) were used as primary antibodies. As a secondary antibody, peroxidaseconjugated goat anti–rat IgG (H+L) (Dianova, Hamburg, Germany) was used.
In Vitro Colony-forming Assays.
The number of hematopoietic progenitor cells was determined in clonogenic assays as described previously (28). Cell samples were obtained from femurs and suspensions of spleen and liver derived from one individual animal at the age of 8–12 wk. The culture mixture for the assays consisted of 0.8% methyl cellulose, 1% BSA, 10% FCS, 5% human plasma and growth factors. Cells were seeded in agar cultures at a concentration of 104/well, and CFU-GM and BFU-E were stimulated with G-CSF (50 ng/ml) plus murine IL-3 (300 U/ml) or stem cell factor (150 ng/ml) plus IL-3 plus erythropoietin (2 U/ml), respectively. The total number of colonies per organ was calculated using the cellularity obtained for each organ and for the bone marrow, assuming that each femur represents 6% of the total marrow.
Peripheral Blood Counts.
In general anesthesia, blood was obtained by cardiac puncture and analyzed in a CellDyn 3500 hemocounter (Abbott, Delkenheim, Germany).
Results
IL-6–sIL-6R Mice Are Smaller and Have a Reduced Body Weight.
The IL-6 transgene expression was regulated by the mouse metallothionein-1 promoter. The sIL-6R transgene expression was driven by the rat PEPCK promoter, which acts neonatally and permits evaluation of the influence of gene products after birth without intrauterine developmental effects. A distinct phenotype was observed in IL-6– sIL-6R mice compared with single-transgenic mice: IL-6 and sIL-6R single-transgenic and nontransgenic littermates (Fig. 1 A, left; data not shown) developed normally with respect to weight gain, food and water intake, appearance, and behavior. However, IL-6–sIL-6R mice (Fig. 1 A, right) were remarkably smaller compared with single-transgenic and nontransgenic littermates. When body weights were measured over a period of 20 wk, it became evident that the IL-6–sIL-6R mice developed reduced body weights as soon as 6 wk after birth (Fig. 1 B). At the age of 20 wk, mean values obtained in IL-6–sIL-6R mice were around 20–25 g compared with 40 g in single-transgenic and nontransgenic littermates (Fig. 1 B). Moreover, at autopsy and at histopathological analysis, IL-6–sIL-6R mice were found to have markedly reduced body fat in all regions of the organism compared with single-transgenic littermates (data not shown).
Figure 1.


Weight abnormalities in IL-6–sIL-6R mice. (A) An IL-6– sIL-6R double-transgenic mouse on the right side and a sIL-6R singletransgenic mouse on the left side at the age of 8 wk. (B) Total body weight recorded of IL-6–sIL-6R mice, of single-transgenic mice, and of nontransgenic littermates.
Massive Increase of Liver and Spleen Weight in IL-6–sIL-6R Mice.
Most remarkably, at the age of 4–6 wk, IL-6–sIL6R mice displayed distended abdominal regions (Fig. 1 A). Single-transgenic and nontransgenic littermates were normal in this respect. The abdominal distension was caused by a marked increase of liver and spleen weights relative to the total body weight. Time course experiments (Fig. 2, A and B) revealed steadily increasing relative liver and spleen weights in IL-6–sIL-6R mice, while the respective relative organ weights in single-transgenic and nontransgenic littermates were unchanged. At wk 20, the relative liver weight (Fig. 2 A) had duplicated and the relative spleen weight (Fig. 2 B) had increased by a factor of 5 when compared with single-transgenic and nontransgenic littermates.
Figure 2.



Organ weight and gross morphology of liver and spleen. Liver (A) and spleen (B) weight of IL-6–sIL-6R, IL-6, and sIL-6R mice and nontransgenic littermates. Organ weight is presented as liver weight/total body weight and spleen weight/total body weight, respectively. Gross morphology of livers and spleens of sIL-6R mice (C, left) and IL-6–sIL-6R mice (C, right) at 12 wk of age.
The gross morphology of livers and spleens were examined in mice at 8 wk of age (Fig. 2 C). Livers and spleens of 8-wk-old IL-6R mice (Fig. 2 C, left) and IL-6 mice and nontransgenic littermates (data not shown) were completely normal. Livers of 8-wk-old IL-6–sIL-6R mice (Fig. 2 C, right), however, appeared dark in color and had a rugged and humpy surface. This appearance was caused by round, whitish, and with respect to the liver surface elevated foci. The spleens of IL-6–sIL-6R mice were clearly enlarged in size (Fig. 2 C, right). The enlargement of the spleen is even more remarkable in light of the fact that IL-6–IL-6R mice had half the body weight of single-transgenic and nontransgenic littermates (see Fig. 1 B).
Increase of Liver and Spleen Weight in IL-6–sIL-6R Mice Is Caused by Extramedullary Hematopoiesis.
The increase of liver and spleen weight in IL-6–sIL-6R mice was caused by a marked extramedullary proliferation of hematopoietic cells in both organs (Fig. 3, A–D), which was not detected in any other parenchymal organ. No extramedullary hematopoiesis was found in single-transgenic and nontransgenic littermates (Fig. 3, A′–D′; data not shown). In the livers and spleens of IL-6–sIL-6R mice, multiple hematopoietic foci were present, which increased in size with time. At 8 wk, there were occasional foci (Figs. 3, A and B) present in both organs. However, at 16 wk, complete involvement of liver and spleen with multiple and large confluent hematopoietic foci accompanied by loss of the parenchymal organ architecture was demonstrated (Fig. 3, C and D). Determination of enzymatic values in the serum (LDH, alkaline phosphatase, and transaminases) did not show any alteration either in IL-6–sIL-6R mice or in single-transgenic and nontransgenic littermates (data not shown), indicating that the liver function of the mice was not compromised.
Figure 3.
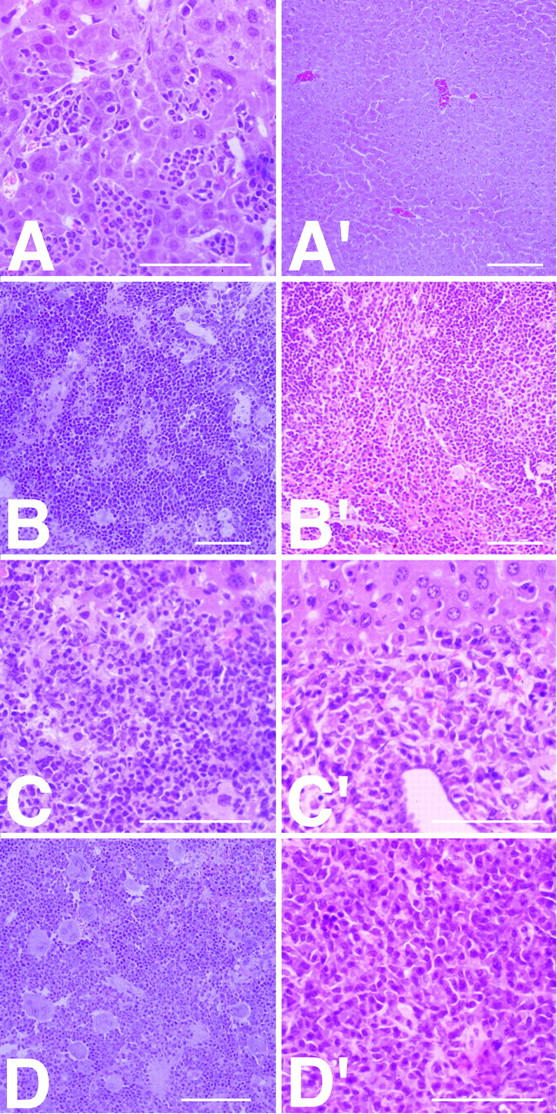
Histomorphological analysis of livers and spleens in IL-6–sIL-6R (A–D), and IL-6 (A′–D′) mice. (A) Liver tissue with several hematopoietic foci containing predominantly granulopoietic precursor cells and terminally differentiated segmental granulocytes (IL-6–sIL-6R, 8 wk). (A′) Normal liver tissue with absence of hematopoietic foci in an 8-wk-old IL-6 transgenic mouse. (B) Spleen tissue with activated extramedullary hematopoiesis showing enlarged granulopoietic and erythropoietic areas as well as increased number of megakaryocytes (IL-6–sIL-6R, 8 wk). (B′) Spleen tissue from an IL-6 transgenic mouse at 8 wk showing regular distribution of red and white pulp and little hematopoiesis. (C) Liver tissue with large hematopoietic focus displaying blastic precursor cells and terminally differentiated granulocytes (IL-6–sIL6R, 16 wk). (C′) Liver tissue from an IL-6 transgenic mouse at 16 wk with periportal plasmacytoma infiltrate. (D) Spleen tissue with destroyed architecture and complete involvement by hematopoiesis (IL-6–sIL-6R, 16 wk). (D′) Spleen tissue completely infiltrated by plasmacytoma (IL-6, 16 wk). The scale bars indicate 100 μm.
Also, a large number of megakaryocytes was detected in IL-6–sIL-6R mice (Fig. 3, B and D) but not in single-transgenic and nontransgenic littermates (Fig. 3, A′–D′). Remarkably, megakaryocytes were present only in the spleen (Fig. 3, B and D) and not in the liver (Fig. 3, A and C). Megakaryocytes were found as soon as the age of 8 wk (Fig. 3 B) and further increased in number at the age of 16 wk (Fig. 3 D).
Plasmacellular infiltrates in the liver and spleen with consecutive involvement of all parenchymal organs were observed in 50% of both IL-6 (Fig. 3, C′ and D′) and IL-6– sIL-6R mice (data not shown) around the age of 16 wk. In IL-6 single-transgenic animals, plasmacellular infiltrates were the only histopathological abnormalities detected. However, in IL-6–sIL-6R mice, extramedullary hematopoiesis (Fig. 3, A–D) was the most dominant histopathological finding observed in all mice examined.
These results clearly demonstrate that the presence of the IL-6–sIL-6R complex leads to extramedullary accumulation of hematopoietic progenitor cells, which are unresponsive to IL-6 alone.
Characterization of Hematopoietic Cells by Immunohistochemistry and FACS® Analysis.
To characterize further the nature of the hematopoietic cells in the liver, immunohistochemistry was performed with antibodies to surface antigens of neutrophils (Ly-6), monocytes/macrophages (Ia), B cells (B220), and T cells (CD3). As early as 4 wk after birth, IL-6– sIL-6R mice showed single positive neutrophils (Fig. 4 A) and monocytes/macrophages (Fig. 4 A′) in the liver. At the age of 6 wk, there were increased numbers of scattered neutrophilic granulocytes (Fig. 4 B) and small clusters of monocytic cells (Fig. 4 B′) present in the liver. Large confluent foci of both cell types were detected at the age of 16 wk (Figs. 4, C and C′). Immunohistochemistry using antibodies directed against B and T cell epitopes revealed that B cells were also present, but to a lesser extent, and that T cells were absent (data not shown). In single-transgenic and nontransgenic littermates, immunohistochemistry using the above mentioned antibodies gave negative results (data not shown).
Figure 4.
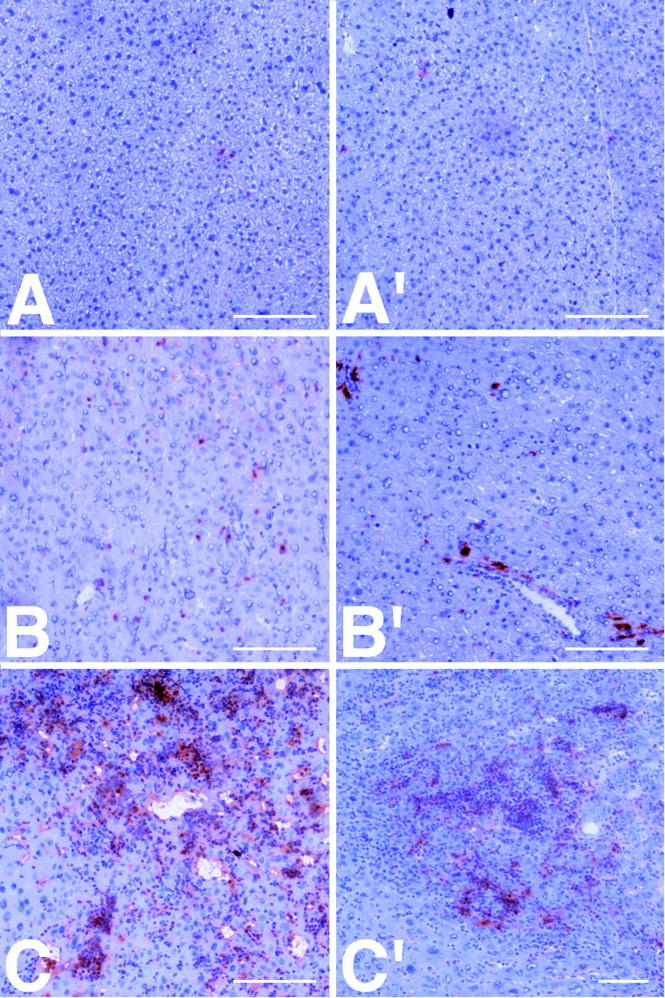
Immunohistochemical analysis of the Ly-6 (A–C), and Ia (A′–C′) surface protein expression on hematopoietic liver cells in IL-6–sIL-6R double-transgenic mice. Granulocytic (A) and monocytic/macrophage (A′) cells are almost completely absent in the liver at the age of 4 wk. At 6 wk, scattered granulocytic (B) and small clusters of monocytic/ macrophage cells (B′) appear in the liver, representing early hepatic extramedullary hematopoiesis. At the age of 12 wk, there are large confluent hepatic foci showing predominantly granulopoietic differentiation (C), but also significant monopoietic cells (C′). The scale bars indicate 100 μm.
Cells harvested from liver, spleen, and bone marrow from single-transgenic and nontransgenic littermates and from IL-6–sIL-6R mice were subjected to FACS® analysis to determine the percentage of macrophages, granulocytes, progenitor cells, and B and T lymphocytes present in these organs. In the bone marrow, there were no significant differences detected between IL-6–sIL-6R mice and singletransgenic and nontransgenic littermates with respect to the various surface antigens tested (data not shown). In the spleen of IL-6–sIL-6R mice (Table 1, upper panel), the number of granulocytes, macrophages, and Sca-1+ progenitor cells increased by a factor of 2.3, 1.5, and 2, respectively, compared with single-transgenic or nontransgenic littermates. B cells were increased 2.3-fold in both IL-6 singletransgenic and in IL-6–sIL-6R mice compared with nontransgenic littermates. CD4+ and CD8+ lymphocytes were not different among the groups of mice tested.
Table 1.
Distribution of Phenotypes of Hematopoietic Cells Isolated from the Spleen and Liver
| Organ | Mice | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Marker | Nontransgenic | sIL-6R | IL-6 | IL-6–sIL-6R | ||||||
| Spleen | GR1 | 10.0 | 10.5 | 10.7 | 23.6 | |||||
| MAC-1 | 12 | 14 | 6.7 | 21.6 | ||||||
| SCA-1 | 20 | 21 | 23.4 | 42 | ||||||
| B220 | 14 | 15 | 35 | 36 | ||||||
| CD8 | 2.15 | 2.11 | 2.34 | 3.2 | ||||||
| CD4 | 6.8 | 8.8 | 3.8 | 5.8 | ||||||
| Liver | GR1 | 0.003 | 0.004 | − | 1.14 | |||||
| MAC-1 | − | 0.006 | 0.08 | 1.04 | ||||||
| SCA-1 | − | − | − | 0.932 | ||||||
| B220 | 0.001 | 0.002 | 0.09 | 0.665 | ||||||
Freshly isolated cells obtained from organs as indicated in the table were subjected to FACS® analysis. Values are expressed as total numbers of cells × 106 per organ. One representative experiment out of three is shown.
The most prominent findings were seen in the liver of IL-6–sIL-6R mice (Table 1, lower panel). Granulocytes, macrophages, Sca-1+ progenitor cells, and B cells were absent or present only scarcely in single-transgenic sIL-6R mice and nontransgenic littermates. Macrophages and B cells were increased 15-fold and 45-fold in IL-6 single-transgenic mice, respectively, compared with sIL-6R single-transgenic or nontransgenic littermates. All cell lines (granulocytes, macrophages, progenitor cells, and B cells) were increased by a factor of 200–300 in IL-6–sIL-6R mice compared with IL-6R single-transgenic or nontransgenic littermates.
These results show that hematopoietic cells in the liver and spleen of IL-6–sIL-6R mice were mainly granulocytes and monocytes, to a lesser extent B cells, and not T cells.
Massive Hematopoietic Progenitor Cell Proliferation in the Liver and Spleen.
In view of the extramedullary hematopoiesis observed in liver and spleen, we investigated whether the continuous stimulation of the gp130 signal transducer affected committed hematopoietic progenitor cells of the granulocytic–monocytic and erythocytic lineages in different hematopoietic organs. The frequency of hematopoietic progenitor cells in the bone marrow, spleen, and liver of transgenic and nontransgenic mice was determined using in vitro clonogenic assays. The IL-6–sIL-6R-mediated gp130 stimulation had distinct effects on different hematopoietic lineages in extramedullary organs: the total number of CFU-GM and BFU-E in spleen and liver derived from IL-6–sIL-6R mice was increased ten times when compared with IL-6–sIL-6R single-transgenic mice and nontransgenic littermates (Fig. 5, A and B). Most interestingly, in the bone marrow, the relative and absolute number of clonogenic hematopoietic cells was not elevated in IL-6–sIL6R mice compared with IL-6 and sIL-6R single-transgenic and nontransgenic mice (Fig. 5 C).
Figure 5.
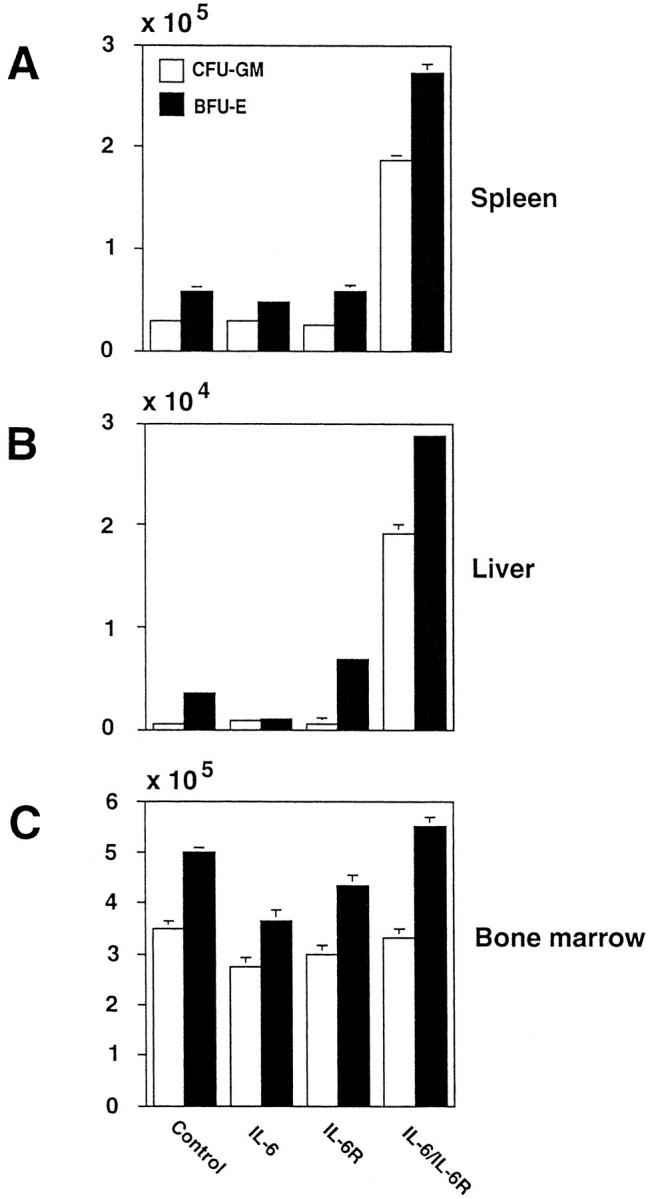
Evaluation of hematopoietic progenitor cells in the bone marrow, spleen, and liver of transgenic mice and nontransgenic littermates. Standard CFU-GM (open bars) and BFU-E (closed bars) assays were performed using spleen (A), liver (B), or bone marrow (C) cells from either IL-6–sIL-6R, IL-6, and sIL-6R mice or nontransgenic littermates, as indicated in the figure. Triplicate cultures were scored after staining with May-Grünwald-Giemsa (28) and are presented as mean values and standard deviation of one representative experiment out of three.
These data show that continuous gp130 activation leads to a massive increase of extramedullary committed hematopoietic progenitor cells and not to an increase of bone marrow–derived progenitor cells.
Extramedullary Hematopoietic Progenitor Cells Are Functionally Normal.
Peripheral blood cell numbers were determined at the age of 8, 16, and 24 wk in IL-6–sIL-6R mice and compared with numbers measured in IL-6 and sIL-6R single-transgenic mice and nontransgenic littermates. Whereas the peripheral blood cell numbers of single-transgenic and nontransgenic littermates were unchanged over the period of time tested, a most dramatic increase of white blood cells consisting mainly of neutrophilic granulocytes was observed in IL-6–sIL-6R mice (Fig. 6, A and B). At the age of 8 wk, there was no difference observed between IL-6–sIL-6R mice and single-transgenic mice and nontransgenic littermates. However, at the age of 16 wk, white blood cells and neutrophils increased by a factor of 10 and 17, respectively, and at the age of 24 wk by a factor of 27 and 48, respectively, in IL-6–sIL-6R mice. Platelets (Fig. 6 C), red blood cells (Fig. 6 D), and hemoglobin values (Fig. 6 E) of IL-6– sIL-6R mice increased by a factor of 2.1, 1.5, and 1.5, respectively, when compared with age-matched nontransgenic littermates and did not change in 16- and 24-wk-old mice. In single-transgenic animals, peripheral blood cell counts and hemoglobin values were not altered compared with nontransgenic mice (Fig. 6, C–E). Thus, the expanded pool of hematopoietic progenitor cells in IL-6–sIL-6R mice appears to retain its capacity to undergo proliferation and differentiation into mature blood cells, resulting in expansion of circulating blood cells.
Figure 6.

Peripheral blood hematological data. White blood cell (A), neutrophil (B), platelet (C), red blood cell (D), and hemoglobin (E) values were analyzed from six transgenic mice and nontransgenic littermates per group at ages indicated in the figure. Mean values with standard deviations are presented.
Discussion
Our study demonstrates that the activation of the gp130 signal transducer by a complex of IL-6 and the sIL-6R represents a major stimulation of growth and differentiation of hematopoietic progenitor cells in vivo. The presence of IL-6 alone is not sufficient to stimulate these cells. As demonstrated by histomorphological and immunohistochemical data and in vitro clonogenic assays, in IL-6–sIL-6R mice an expansion of hematopoietic progenitor cells of the granulocytic–monocytic, megakaryocytic, and erythroid lineages was observed.
We conclude that these hematopoietic progenitor cells express gp130 signal-transducing molecules on their surface and lack the specific IL-6Rα. This notion is underscored by a recently published study using gp130-deficient mice, which were shown to exhibit a greatly diminished number of hematopoietic progenitor cells (29). Thus, gp130 activation is required to stimulate hematopoietic progenitor cells. Because IL-6 and IL-11 act via a gp130 homodimer, loss of IL-6 can be compensated by IL-11, possibly explaining why IL-6–deficient mice are viable (30, 31). In mice overexpressing cytokines like LIF (32) or OSM (33), requiring the gp130/LIF-R heterodimer for signal transduction, hematopoietic effects are scarce, indicating that hematopoietic progenitor cells express little or no LIF-R on their surface. This hypothesis is supported by findings from mice in which the LIF-R has been deleted by targeted disruption (34). These mice show no significant abnormalities of the hematopoietic system.
The fact that increased B cell numbers are detected in spleen and liver of IL-6 and IL-6–sIL-6R transgenic mice supports the notion that B cells express the specific IL-6R on their surface and thus may be activated by IL-6 alone (35). Accordingly, IL-6 induced plasmacytosis in IL-6 and IL-6–sIL-6R mice by stimulating proliferation and maturation of B cells (36, 37).
In IL-6–sIL-6R double transgenic mice a marked reduction of body weight and body size associated with a reduction of body fat was observed compared with single-transgenic and nontransgenic littermates. Currently, we can not explain the reduced body size observed in these animals. However, the reduced body fat that we observed in IL-6– sIL-6R mice could be explained by the fact that the recently identified leptin receptor (OB-R) (38) is a signaltransducing protein similar to gp130, involving the same intracellular signaling molecules (39, 40). The OB-R is located in many tissues, including the hypothalamus, and is involved in the regulation of the size of adipose tissue mass through effects on satiety and energy metabolism. The natural ligand of the OB-R, leptin, a 16-kD protein, is expressed primarily in the white adipose tissue (41). It might be possible that the IL-6–sIL-6R complex activates gp130 molecules on OB-R–expressing cells, thereby activating leptinspecific signaling pathways.
We have provided evidence that the IL-6–sIL-6R complex in vivo induces extramedullary hematopoiesis in liver and spleen but not in bone marrow. It is known that there are resident hematopoietic stem cells in the adult liver (42). These cells might be expanded after stimulation by IL-6– sIL-6R in a concentration-dependent manner. Rather high concentrations of IL-6 (2 nM) and sIL-6R (>20 nM) are required for the activation of gp130-expressing cells in vitro (43). This is somewhat surprising considering that the KD of the IL-6–sIL-6R interaction is in the range of 1 nM (16, 44). A possible explanation could be that the average half-life of the IL-6–IL-6R complex is shorter than the time required to assemble the functionally active IL-6–IL6R/gp130 complex. We have recently demonstrated that the sIL-6R acts predominantly at the site of its generation (M. Peters et al., submitted for publication). Because liverspecific promoters drive the expression of the transgenes used in the present study, one could reason that sufficient concentrations of IL-6 and sIL-6R required for an effective stimulation of hematopoietic progenitor cells are found predominantly in the liver. Recently, transgenic mice coexpressing IL-6 and the membrane-bound IL-6R have been generated (45) in which soluble IL-6R was generated by shedding at levels 100-fold lower compared with IL-6– sIL-6R mice of the present study. No hematopoietic abnormalities were found in these mice, indicating that high levels of the IL-6–sIL-6R complex are required to stimulate hematopoietic progenitor cells.
Extramedullary hematopoiesis in the spleen has been recognized after administration of recombinant G-CSF to mice (46). Splenic stimulation of progenitors in IL-6–sIL-6R mice indicates that cytokines of the IL-6 family can contribute to hematopoiesis in the spleen. Recently, it has been demonstrated that LIF overexpression results in moderate splenomegaly and slightly activated extramedullary hematopoiesis (32, 47). LIF-deficient mice show a reduction of hematopoietic progenitors in the spleen and bone marrow. However, hematopoietic spleen and bone marrow progenitor cells from those mice transplanted into lethally irradiated normal mice retained their normal function, indicating that LIF is required in the microenvironment to maintain stem cell numbers, rather than affecting the potential of hematopoietic progenitors (48). In light of these results, one might speculate, that similar to LIF, the IL-6– sIL-6R complex contributes to the microenvironment required by hematopoietic cells in spleen and liver. However, it is of interest that stimulation by IL-6–sIL-6R leads to far stronger hematopoiesis than stimulation by LIF.
It is possible that liver and spleen cells express other important growth factors like stem cell factor (49, 50), chemokines as pre-B cell–stimulating factor (51), or flt-3 (52), either in membrane-bound or soluble form. These cytokines might be involved as cofactors in the growth of hematopoietic progenitor cells in these organs.
Hematopoietic progenitor cells stimulated by the IL-6– sIL-6R complex are functionally normal. This is reflected by the highly elevated numbers of granulocytes, megakaryocytes, and erythrocytes in the peripheral blood. Thus, the expanded pool of hematopoietic progenitor cells appears to retain its capacity to undergo proliferation and differentiation into mature blood cells, resulting in expansion of circulating blood cells.
In summary, there are three major findings of this study. First, we provided evidence that continuous activation of the gp130 signal transducer by molecules like the IL-6–sIL-6R leads to a most effective stimulation of hematopoietic progenitor cells. Progenitor cells of the granulocytic–monocytic, megakaryocytic, and erythroid lines are involved. Second, IL-6–sIL-6R causes a stimulation of predominantly extramedullary and not bone marrow hematopoietic progenitor cells. Third, progenitor cells are functionally normal, retaining their capacity to undergo maturation into mature blood cells. Apart from expanding CD34+ cells obtained from cord blood in vitro (43), continuous activation of the gp130 signal transducer by agents like the hIL-6–hsIL-6R complex could be the most effective mechanism to reconstitute hematopoiesis in vivo or ex vivo in patients with cytopenic conditions.
Acknowledgments
We thank B. Gilberg and K. Petmecky for excellent technical assistance and Dr. M. Blessing, Boehringer Ingelheim Research Group, I. Department of Medicine, University of Mainz, for continuous advice and discussions throughout the study. Dr. B. Lotz is thanked for the determination of the peripheral blood values.
This work was supported by the Deutsche Forschungsgemeinschalt (Bonn, Germany) and the Naturwissenschaftlich-Medizinisches Forschungszentrum (Mainz, Germany) to M. Peters, P. Schirmacher, C. Peschel, and S. Rose-John.
Footnotes
1 Abbreviations used in this paper: CNTF, ciliary neurotrophic factor; CT-1, cardiotrophin-1; LIF, leukemia inhibitory factor; OSM, oncostatin M; PEPCK, phosphoenolpyruvate carboxykinase; sIL-6R, soluble interleukin-6 receptor; TNF-R, tumor necrosis factor receptor.
References
- 1.Akira S, Taga T, Kishimoto T. Interleukin-6 in biology and medicine. Adv Immunol. 1993;54:1–78. doi: 10.1016/s0065-2776(08)60532-5. [DOI] [PubMed] [Google Scholar]
- 2.Bauer J, Herrmann F. Interleukin-6 in clinical medicine. Ann Hematol. 1991;62:203–210. doi: 10.1007/BF01729833. [DOI] [PubMed] [Google Scholar]
- 3.Gauldie J, Richards C, Harnish D, Lansdorp P, Baumann H. Interferon beta 2/B-cell stimulating factor type 2 shares identity with monocyte-derived hepatocytestimulating factor and regulates the major acute phase response in liver cells. Proc Natl Acad Sci USA. 1987;84:7251–7255. doi: 10.1073/pnas.84.20.7251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hibi M, Nakajima K, Hirano T. IL-6 cytokine family and signal transduction: a model of the cytokine system. J Mol Med. 1996;74:1–12. doi: 10.1007/BF00202068. [DOI] [PubMed] [Google Scholar]
- 5.Pennica D, King KL, Shaw KJ, Luis E, Rullamas J, Luoh S-M, Darbonne WC, Knutzon DS, Yen R, Chien KR, et al. Expression cloning of cardiotrophin 1, a cytokine that induces cardiac myocyte hypertrophy. Proc Natl Acad Sci USA. 1995;92:1142–1146. doi: 10.1073/pnas.92.4.1142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kishimoto T, Taga T, Akira S. Cytokine signal transduction. Cell. 1994;76:253–262. doi: 10.1016/0092-8674(94)90333-6. [DOI] [PubMed] [Google Scholar]
- 7.Ihle JN. Cytokine receptor signalling. Nature (Lond) 1995;377:591–594. doi: 10.1038/377591a0. [DOI] [PubMed] [Google Scholar]
- 8.Schindler C, Darnell JE. Transcriptional responses to polypeptide ligands: the JAK–STAT pathway. Annu Rev Biochem. 1995;64:621–651. doi: 10.1146/annurev.bi.64.070195.003201. [DOI] [PubMed] [Google Scholar]
- 9.Pennica D, Arce V, Swanson TA, Vejsada R, Pollock RA, Armanini M, Dudley K, Phillips HS, Rosenthal A, Kato AC, Henderson CE. Cardiotrophin-1, a cytokine present in embryonic muscle, supports long-term survival of spinal motoneurons. Neuron. 1996;17:63–74. doi: 10.1016/s0896-6273(00)80281-0. [DOI] [PubMed] [Google Scholar]
- 10.Mosley B, DeImus C, Friend D, Boiani N, Thoma B, Park LS, Cosman D. Dual Oncostatin M (OSM) receptors. Cloning and characterization of an alternative signaling subunit conferring OSM-specific receptor activation. J Biol Chem. 1996;271:32635–32643. doi: 10.1074/jbc.271.51.32635. [DOI] [PubMed] [Google Scholar]
- 11.Maliszewski CR, Sato TA, Vanden T, Bos, Waugh S, Dower SK, Slack J, Beckmann MP, Grabstein KH. Cytokine receptors and B cell functions. I. Recombinant soluble receptors specifically inhibit IL-1– and IL-4– induced B cell activities in vitro. J Immunol. 1990;144:3028–3033. [PubMed] [Google Scholar]
- 12.Aderka D, Engelmann H, Hornik V, Skornik Y, Levo Y, Wallach D, Kushtai G. Increased serum levels of soluble receptors for tumor necrosis factor in cancer patients. Cancer Res. 1991;51:5602–5607. [PubMed] [Google Scholar]
- 13.Layton MJ, Cross BA, Metcalf D, Ward LD, Simpson RJ, Nicola NA. A major binding protein for leukemia inhibitory factor in normal mouse serum: identification as a soluble form of the cellular receptor. Proc Natl Acad Sci USA. 1992;89:8616–8629. doi: 10.1073/pnas.89.18.8616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rose-John S, Heinrich PC. Soluble receptors for cytokines and growth factors: generation and biological function. Biochem J. 1994;300:281–290. doi: 10.1042/bj3000281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Baumann H, Wang Y, Morella KK, Lai C-F, Dams H, Hilton DJ, Hawley RG, Mackiewicz A. Complex of the soluble IL-11 receptor and IL-11 acts as IL-6–type cytokine in hepatic and nonhepatic cells. J Immunol. 1996;157:284–290. [PubMed] [Google Scholar]
- 16.Yamasaki K, Taga T, Hirata Y, Yawata H, Kawanishi Y, Seed B, Taniguchi T, Hirano T, Kishimoto T. Cloning and expression of the human interleukin-6 (BSF-2/ IFN β2) receptor. Science (Wash DC) 1988;241:825–828. doi: 10.1126/science.3136546. [DOI] [PubMed] [Google Scholar]
- 17.Taga T, Hibi M, Hirata Y, Yamasaki K, Yasukawa K, Matsuda T, Hirano T, Kishimoto T. Interleukin-6 triggers the association of its receptor with a possible signal transducer, gp130. Cell. 1989;58:573–581. doi: 10.1016/0092-8674(89)90438-8. [DOI] [PubMed] [Google Scholar]
- 18.Hibi M, Murakami M, Saito M, Hirano T, Taga T, Kishimoto T. Molecular cloning and expression of an IL-6 signal transducer, gp130. Cell. 1990;63:1149–1157. doi: 10.1016/0092-8674(90)90411-7. [DOI] [PubMed] [Google Scholar]
- 19.Müllberg J, Schooltink H, Stoyan T, Günther M, Graeve L, Buse G, Mackiewicz A, Heinrich PC, Rose-John S. The soluble interleukin-6-receptor is generated by shedding. Eur J Immunol. 1993;23:473–480. doi: 10.1002/eji.1830230226. [DOI] [PubMed] [Google Scholar]
- 20.Müllberg J, Dittrich E, Graeve L, Gerhartz C, Yasukawa K, Taga T, Kishimoto T, Heinrich PC, RoseJohn S. Differential shedding of the two subunits of the interleukin-6 receptor. FEBS Lett. 1993;332:174–178. doi: 10.1016/0014-5793(93)80507-q. [DOI] [PubMed] [Google Scholar]
- 21.Novick D, Engelmann H, Wallach D, Rubinstein M. Soluble cytokine receptors are present in normal human urine. J Exp Med. 1989;170:1409–1414. doi: 10.1084/jem.170.4.1409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Narazaki M, Yasukawa K, Saito K, Ohsug Y, Fukui H, Koishihara Y, Yancopoulos GD, Taga T, Kishimoto T. Soluble forms of the interleukin-6 signal transducing receptor component gp130 in human serum possessing a potential to inhibit signals through membrane-anchored gp130. Blood. 1993;82:1120–1126. [PubMed] [Google Scholar]
- 23.De Benedetti F, Massa M, Pignatti P, Albani S, Novick D, Martini A. Serum soluble interleukin 6 (IL-6) receptor and IL-6/soluble IL-6 receptor complex in systemic juvenile rheumatoid arthritis. J Clin Invest. 1994;93:2114–2119. doi: 10.1172/JCI117206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Walev I, Vollmer P, Palmer M, Bhakdi S, RoseJohn S. Pore-forming toxins trigger shedding of receptors for interleukin 6 and lipopolysaccharide. Proc Natl Acad Sci USA. 1996;93:7882–7887. doi: 10.1073/pnas.93.15.7882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Peters M, Jacobs S, Ehlers M, Vollmer P, Müllberg J, Wolf E, Brem G, Meyer zum Büschenfelde K-H, Rose-John S. The function of the soluble interleukin 6 (IL-6) receptor in vivo: sensitization of human soluble IL-6 receptor transgenic mice towards IL-6 and prolongation of the plasma half-life of IL-6. J Exp Med. 1996;183:1399–1406. doi: 10.1084/jem.183.4.1399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Fattori E, Della C, Rocca, Costa P, Giorgio M, Dente B, Pozzi D, Ciliberto G. Development of progressive kidney damage and myeloma kidney in interleukin-6 transgenic mice. Blood. 1994;83:2570–2579. [PubMed] [Google Scholar]
- 27.Ohteki T, Seki S, Abo T, Kumagai K. Liver is a possible site for the proliferation of abnormal CD3+4−8− double-negative lymphocytes in autoimmune MRL-lpr/lprmice. J Exp Med. 1990;172:7–12. doi: 10.1084/jem.172.1.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Peschel C, Paul WE, Ohara J, Green I. Effects of B cell stimulatory factor-1/interleukin 4 on hematopoietic progenitor cells. Blood. 1987;70:254–263. [PubMed] [Google Scholar]
- 29.Yoshida K, Taga T, Saito M, Suematsu S, Kumanogh A, Tanaka T, Fujiwara H, Hirata M, Yamagami T, Nakahata T, Kishimoto T. Targeted disruption of gp130, a common signal transducer for the interleukin 6 family of cytokines, lead to myocardial and hematological disorders. Proc Natl Acad Sci USA. 1996;93:407–411. doi: 10.1073/pnas.93.1.407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kopf M, Baumann H, Freer G, Freudenberg M, Lamers M, Kishimoto T, Zinkernagel R, Bluethmann H, Köhler G. Impaired immune and acute-phase responses in interleukin-6–deficient mice. Nature (Lond) 1994;368:339–342. doi: 10.1038/368339a0. [DOI] [PubMed] [Google Scholar]
- 31.Bernad A, Kopf M, Kulbacki R, Weich N, Köhler G, Guttierrez-Ramos JC. Interleukin-6 is required in vivo for the regulation of stem cells and committed progenitors of the hematopoietic system. Immunity. 1994;1:725–731. doi: 10.1016/s1074-7613(94)80014-6. [DOI] [PubMed] [Google Scholar]
- 32.Shen MM, Skoda RC, Cardiff RD, Campos-Torres J, Leder P, Ornitz DM. Expression of LIF in transgenic mice results in altered thymic epithelium and apparent interconversion of thymic and lymph node morphologies. EMBO (Eur Mol Biol Org) J. 1994;13:1375–1385. doi: 10.1002/j.1460-2075.1994.tb06391.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Malik N, Haugen HS, Modrell B, Shoyab M, Clegg CH. Developmental abnormalities in mice transgenic for bovine Oncostatin M. Mol Cell Biol. 1995;15:2349–2358. doi: 10.1128/mcb.15.5.2349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ware CB, Horowitz MC, Renshaw BR, Hunt JS, Liggit D, Kobler SA, Gliniak BC, McKenna HJ, Papayannopoulou T, Thoma B, et al. Targeted disruption of the low-affinity leukemia inhibitory factor receptor gene causes placental, skeletal, neural and metabolic defects and results in perinatal death. Development. 1995;121:1283–1299. doi: 10.1242/dev.121.5.1283. [DOI] [PubMed] [Google Scholar]
- 35.Taga T, Kawanishi Y, Hardy R, Hirano T, Kishimoto T. Receptors for B cell stimulatory factor 2. Quantification, specificity, distribution and regulation of their expression. J Exp Med. 1987;166:967–981. doi: 10.1084/jem.166.4.967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Suematsu S, Matsuda T, Aozasa K, Akira S, Nakano N, Ohno S, Miyazaki J-I, Yamamura K-I, Hirano T, Kishimoto T. IgG1 plasmacytosis in interleukin 6 transgenic mice. Proc Natl Acad Sci USA. 1989;86:7547–7551. doi: 10.1073/pnas.86.19.7547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Suematsu S, Matsusaka T, Matsuda T, Ohno S, Miyazaki J-I, Yamamura K-I, Hirano T, Kishimoto T. Generation of plasmacytomas with the chromosomal translocation t(12;15) in interleukin 6 transgenic mice. Proc Natl Acad Sci USA. 1992;89:232–235. doi: 10.1073/pnas.89.1.232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tartaglia LA, Dembski M, Weng X, Deng N, Culpepper J, Devos R, Richards GJ, Campfield SA, Clark FT, Deeds J, et al. Identification and expression cloning of a leptin receptor, OB-R. Cell. 1995;83:1263–1271. doi: 10.1016/0092-8674(95)90151-5. [DOI] [PubMed] [Google Scholar]
- 39.Baumann H, Morella KK, White DW, Dembski M, Bailon PS, Kim H, Lai C-F, Tartaglia LA. The full-length leptin receptor has signaling capabilities of interleukin 6-type cytokine receptors. Proc Natl Acad Sci USA. 1996;93:8374–8378. doi: 10.1073/pnas.93.16.8374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Vaisse C, Halaas JL, Horvarth CM, Darnell JE, Stoffel M, Friedman JM. Leptin activation of Stat3 in the hypothalamus of wild-type and ob/ob mice but not db/dbmice. Nat Gen. 1996;5:765–768. doi: 10.1038/ng0996-95. [DOI] [PubMed] [Google Scholar]
- 41.Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nat (Lond) 1994;372:425–431. doi: 10.1038/372425a0. [DOI] [PubMed] [Google Scholar]
- 42.Taniguchi H, Toyoshima T, Fukao K, Nakauchi H. Presence of hematopoietic stem cells in the adult liver. Nature Med. 1996;2:198–203. doi: 10.1038/nm0296-198. [DOI] [PubMed] [Google Scholar]
- 43.Sui X, Kohichiro K, Tanaka R, Tajima S, Muraoka K, Ebihara Y, Ikebuchi K, Yasukawa K, Taga T, Kishimoto T, Nakahata T. gp130 and c-Kit signalings synergize for ex vivo expansion of human primitive hemopoietic progenitor cells. Proc Natl Acad Sci USA. 1995;92:2859–2863. doi: 10.1073/pnas.92.7.2859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Rose-John S, Schooltink H, Len D, Hipp E, Dorfheus G, Schmitz H, Schiel X, Hirano T, Kishimoto T, Heinrich PC. Studies on the structure and regulation of the human hepatic interleukin-6-receptor. Eur J Biochem. 1990;190:79–83. doi: 10.1111/j.1432-1033.1990.tb15548.x. [DOI] [PubMed] [Google Scholar]
- 45.Hirota H, Yoshida K, Kishimoto T, Taga T. Continuous activation of gp130, a signal-transducing receptor component for interleukin-6-related cytokines, causes myocardial hypertrophy in mice. Proc Natl Acad Sci USA. 1995;92:4862–4866. doi: 10.1073/pnas.92.11.4862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Fukushima N, Nishim H, Koishihara Y, Ohkawa H. Enhanced hematopoiesis in vivo and in vitro by splenic stromal cells derived from the mouse with recombinant granulocyte colony-stimulating factor. Blood. 1992;80:1914–1922. [PubMed] [Google Scholar]
- 47.Metcalf D, Gearing DP. Fatal syndrome in mice engrafted with cells producing high levels of the leukemia inhibitory factor. Proc Natl Acad Sci USA. 1989;86:5948–5952. doi: 10.1073/pnas.86.15.5948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Escary JL, Perreau J, Duménil D, Ezine S, Brûlet P. Leukaemia inhibitory factor is necessary for maintenance of haematopoietic stem cells and thymocyte stimulation. Nature (Lond) 1993;363:361–364. doi: 10.1038/363361a0. [DOI] [PubMed] [Google Scholar]
- 49.Flanagan JG, Leder P. The kit ligand: a cell surface molecule altered in steel mutant fibroblasts. Cell. 1990;63:195–194. doi: 10.1016/0092-8674(90)90299-t. [DOI] [PubMed] [Google Scholar]
- 50.Zsebo KM, Williams DA, Geissler EN, Broudy VC, Martin FH, Atkins HL, Hsu RY, Birkett NC, Otkino KH, Murdock DC. Stem cell factor is encoded at the SIlocus of the mouse and is the ligand for the c-kit tyrosine kinase receptor. Cell. 1990;63:213–224. doi: 10.1016/0092-8674(90)90302-u. [DOI] [PubMed] [Google Scholar]
- 51.Nagasawa T, Hirota S, Tachibana K, Takakura N, Nishikawa S-I, Kitamura Y, Yoshida N, Kikitani H, Kishimoto T. Defects of B-cell lymphopoiesis and bonemarrow myelopoiesis in mice lacking the CXC chemokine PBSF/SDF-1. Nature (Lond) 1996;382:635–638. doi: 10.1038/382635a0. [DOI] [PubMed] [Google Scholar]
- 52.Hunte BE, Hudak S, Campbell D, Xu Y, Rennick D. flk2/flt3 ligand is a potent cofactor for the growth of primitive B cell progenitors. J Immunol. 1995;156:489–496. [PubMed] [Google Scholar]