

Abstract
I consider a vesicle with an open pore of small radius rp, exposed to a DNA solution. The crucial moment is the entry, when a chain end faces the pore and enters it. I discuss qualitatively the following three characteristic times: (i) the duration of the entry of one chain end (defining the minimum lifetime of the pore) τe ∼ 10−4 sec, (ii) the transfection time τt (the time required to be sure that one chain has gone in) τt ∼ hours, and (iii) the sliding time τS (the time between entry of one end and entry of the other end) ∼ 1 sec. A fortunate feature is that sliding may proceed even if the pore tends to close itself after entry.
General Aims
For various purposes, it is important to insert a gene into the cytoplasm of a cell. The DNA may be presented inside a viral capside (1) or (possibly) in a special lipid package (2, 3). The DNA also may be bare, because vesicles and living cells can be opened by electroporation, under voltages of order 1 V between the two sides of one membrane (4, 5). I focus my attention here on the latter case. Very often the resulting pore will be open only during a small time (microseconds to milliseconds) and will be small. Is it possible to have a reasonable amount of DNA entry in these conditions?
I propose some estimates here, choosing the simplest situation: a single pore of radius rp opened in a vesicle. I assume that, as soon as the pore is opened, the short circuit is complete: no macroscopic electric field is present, and the DNA chains drift only because of their concentration gradients. I also assume that the DNA is in a perfect double-helix form, excluding the possibility of some denatured regions that would generate relatively large loops able to block the entry.
The process of sliding through a pore has been studied experimentally (6) and theoretically (7), by using an ion channel and a voltage drop as the driving force. The ion channel corresponds to a very different situation: (i) the DNA must be single stranded and is thus flexible, (ii) strong interactions exist between the bases and the channel, leading to a potential energy dependent on the sequence of base pairs, and (iii) the process of entry is not probed directly.
For my problem, as shall be seen, entry is probably the crucial moment. I discuss this below, at the (naive) level of scaling laws. Later, I compare entry and sliding.
The Entry Process
The Driving Force.
My starting point is an outer solution of DNA with a chain length L and a number n = L/a of base pairs per chain. The number of base pairs per unit volume is c; on the other hand, the concentration inside the vesicle is 0, which leads to a difference in chemical potentials:
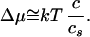 |
1 |
Here cs is the salt concentration (assumed to be larger than c). The origin of the scaling law (1) can be explained in simples terms as follows. Any unit (base pair) sees a repulsive electrostatic potential from units that belong to other chains, lying at a distance r, and of magnitude (ignoring correlations):
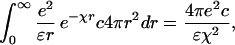 |
2 |
where k−1 is the Debye screening length, e the unit charge, ɛ the dielectric constant of water. By using the standard formula for κ:
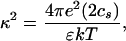 |
3 |
(for monovalent salts) I arrive at Eq. 1.
Penetration.
As usual, this process should be dominated by the chain ends. I start with a pore of radius rp, and a chain end that is near the pore, i.e., within a distance ∼rp, from its center (Fig. 1a). What I want is that the chain enters by a length of order rp. Each monomer on entry then will gain the energy Δμ, which corresponds to a force:
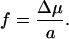 |
4 |
If the whole DNA chain was totally rigid, its friction coefficient in water would be ζtot ∼ ηL, where η is the viscosity. However, the whole chain does not have to be moved to achieve penetration. As shown in Fig. 1b, all that is required is that a chain portion comparable to the persistence length lp move in. This sliding gives a local friction coefficient ζl = ηlp, and the forward velocity of this chain is of order:
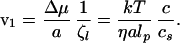 |
5 |
The time required for the entry process, if this initial chain end was in the right region, is:
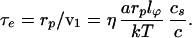 |
6 |
Taking a = 3 Ȧ, rp =10 nm, lp = 100 nm, and cs/c = 1,000, then τt ≅ 10−4 sec.
Figure 1.

Entry of a semirigid chain into a pore. (a) View at high magnification, showing the pore of size rp and the volume of interest (∼rp3). (b) View at a lower magnification, showing the initial state (end at E2).
The Transfection Time τt.
The transfection time is the time required to achieve (on the average) the entry of one chain: it is related to τt, but it incorporates two extra features:
(i) The average number n of chain ends that are in the “useful” region near the pore is small:
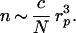 |
7 |
(ii) In most electroporation processes, the field is applied only during a small fraction ψ of the total time.
Thus we may write:
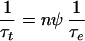 |
8 |
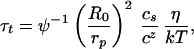 |
9 |
where R02 = Nalp is the end-to-end mean square size of one chain.
Taking n = 104 bp, ψ = 10−3, and c ∼ 2.104 moles/liter, it can be estimated that τt ∼ 2 hr.
The Sliding Process
Once the chain has entered, the force f will push it in, even if the pore closes back. The closing of the pore may induce an extra friction ζ∼, because of local interactions between the double helix and the membrane lipids, and somewhat similar to what is discussed for a specific channel in refs. 1 and 2. But in my case ζ∼ is not large; the overall friction should not be dominated by ζ∼, but rather by the hydrodynamic friction in the water. If entanglements are not important at the concentration c, we should write: ζtot ≅ ηRo. The sliding velocity is thus:
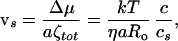 |
10 |
and the overall sliding time is:
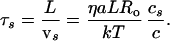 |
11 |
This time is of order 1 sec for this example. It is seen clearly that the bottleneck is at the entry.
Discussion
This model of the entry process is obviously very crude.
If the pore radius rp was smaller than the Debye screening length, electrostatic barriers (between a negatively charged bilayer and the DNA extremity) would play a role.
When the pore is opened, if there is an osmotic pressure difference between inside and outside, caused by small ions or other solutes (sugars, etc.), it shall cause a hydrodynamic flow. The flow may assist or oppose the entry. The present considerations can apply only to the case of exact equilibration.
The transfection time τt is associated with the entry of one chain, which may be the relevant parameter from a biological point of view. From a physical point of view, one would measure a (much larger) time τe, the time required for equilibration of concentrations between outside and inside:
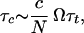 |
12 |
where Ω is the volume of the vesicle.
I restricted my attention to a single pore. However, it may happen that two pores, for instance, are formed, and that one DNA has the end E′ entering through one pore, while the other end, E", enters through the other pore: a “hairpin” process (Fig. 3). There will be a force pushing the DNA inside and decreasing the separation between the two pores(Fig. 3). With a fluid vesicle, this decrease results in fusion of the two pores and should not hinder the process. But with a living cell, the cytoskeleton will block the motion, and a hairpin DNA could be stuck.
Figure 3.

“Hairpins” for a two-pore system. In a vesicle, the two pores (P1, P2) may fuse under the force pushing the chain inside.
However, in spite of all these complications, I believe that my estimate of characteristic times may be useful. The entry time τe tells us what is the minimum allowable lifetime for a pore. The transfection time τt should tell us what is the minimum injection time for a germ in vicinity of an electroporated cell.
Figure 2.

The sliding process.
Acknowledgments
This reflection was greatly stimulated by discussions at a Gordon Conference on complex fluids (Barga, Italy, 1998). I am particularly indebted to A. Ajdari, H. Gruler, A. Parsegian, and E. Sackmann for their comments.
References
- 1.Samulski R J. Development of Human Gene Therapy. Plainview, NY: Cold Spring Harbor Lab. Press; 1999. [Google Scholar]
- 2.Crystal R G. Science. 1995;270:404. doi: 10.1126/science.270.5235.404. [DOI] [PubMed] [Google Scholar]
- 3.Salditt T, Koltover I, Rädler J, Safinya C. Phys Rev Lett. 1997;79:2582–2585. [Google Scholar]
- 4.Neumann E, Schäfer-Ridder M, Yang Y, Hofschneider P. EMBO J. 1982;1:841–859. doi: 10.1002/j.1460-2075.1982.tb01257.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gabriel, B. & Teissié, J. (1999) Biophys. J., in press.
- 6.Kasianowicz J, Brandin E, Branton D, Deamer D. Proc Natl Acad Sci USA. 1996;93:13770–13773. doi: 10.1073/pnas.93.24.13770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lubensky, D. & Nelson, D. (1999) Phys. Rev. E., in press.