

Abstract
Different strains of mice have varying susceptibilities to ultraviolet radiation (UV) of wavelength 280–320 nm (UVB) for 50% suppression of systemic contact hypersensitivity (CHS) responses. Prevalence of histamine-staining dermal mast cells in different strains of mice (C57BL/ 6J, DBA/2, BALB/c) correlated directly with their susceptibility to UVB-induced systemic immunosuppression. BALB/c mice carrying Uvs1, a major locus for susceptibility to UV-induced immunosuppression, contained greater numbers of dermal mast cells than BALB/c mice of the same parental origin. Strains of mice that were differentiated on their susceptibility to UVB-induced downregulation of systemic CHS responses were similar in their susceptibility to histamine-induced immunomodulation. Histamine, but not UVB irradiation, decreased systemic CHS responses in mast cell–depleted mice (W f/W f). Reconstitution of the dorsal skin of W f/W f mice with bone marrow–derived mast cell precursors from nonmutant mice rendered the mice susceptible to UVB irradiation for systemic suppression of CHS responses. UVB irradiation did not suppress delayed type hypersensitivity responses to allogeneic spleen cells in W f/W f mice. In contrast, UV irradiation suppressed CHS responses in W f/W f mice when hapten was applied to the irradiated site. This study demonstrates that dermal mast cells are necessary for the induction of systemic suppression of CHS responses by UVB radiation, and suggests that mast cell– derived histamine is one component of this UVB-induced systemic immunosuppression.
Keywords: rodent, skin, mast cells, ultraviolet B radiation, systemic immunosuppression
The acute effects of UV irradiation of skin may include edema and erythema. However, UV irradiation also suppresses the immune system and provides a mechanism by which UV-induced tumors escape immune destruction (1). UV-induced immunosuppression has been investigated for potential use in a transplantation setting (2) and in treatment of inflammatory diseases of the skin (3). However, mice of different strains differ in their susceptibility to UV radiation of wavelength 280–320 nm (UVB)1-induced immunosuppression (4, 5) and there is evidence that humans with increased incidence of nonmelanoma skin tumors have decreased contact hypersensitivity (CHS) responses to haptens after UVB exposure (6). The mechanisms by which UV irradiation, in particular UVB of 280–320 nm, can regulate immune responses have been the focus of many studies.
Recognizing C57BL/6 and BALB/c mice as relatively susceptible and resistant, respectively, to UVB-induced immunosuppression, a genetic basis for these differences in susceptibility of different mouse strains to UVB has been sought. In a comprehensive study of 18 strains of inbred mice, three phenotypes with significantly different susceptibilities to UV suppression were identified, namely high susceptibility with 50% suppression at 0.7–2.3 kJ/m2 UVB, intermediate susceptibility with 50% suppression at 4.7–6.9 kJ/m2 UVB, and low susceptibility (but not resistance) with 50% suppression at 9.6–12.3 kJ/m2 UVB. Representative strains of mice from these three groups were C57BL/6, DBA/2, and BALB/c, respectively (5). In these studies of UV-induced “systemic” immunosuppression in which the sensitizing hapten was applied to unirradiated sites 3–5 d after irradiation, MHC haplotype did not correlate with UV suppression in four MHC congenic strains (5). In contrast, polymorphisms at a locus within the murine MHC and at the Lps locus, in turn controlling transcription and translation of the Tnf-α gene, were shown to correlate with susceptibility to the effects of UVB for “local” suppression, i.e., of CHS responses to haptens or antigens applied to UV-irradiated sites (4, 7).
Further studies in mice of UVB-induced systemic immunosuppression have identified the locus, Uvs1, as a major locus for susceptibility to UV suppression (8). BALB/c- Uvs1 mice were derived by nine generations of successive backcrossing of F1 hybrid BALB/c × C57BL/6 (CB6F1) mice to the BALB/c parent with selection for high susceptibility to UV suppression (8), followed by three generations of intercrossing. Under these conditions, high susceptibility segregates as an autosomal dominant trait. Both BALB/c and BALB/c-Uvs1 mice are albino. With a dose of UV of 4.5 kJ/m2, C57BL/6 mice (n = 39) show a mean percent suppression of 84 ± 3, BALB/c mice (n = 55) of 30 ± 3 and BALB/c-Uvs1 (n = 123) of 53 ± 2 (8).
Dermal mast cells are activated and degranulate after UVB irradiation (9). Findings of reduced UV-induced swelling in the ears of mast cell–deficient mice (W/Wv; reference 10) support the conclusion that mast cells play an important role in UV-induced inflammation. Histamine is the major biogenic amine of skin mast cells and is stored bound to proteoglycans in cytoplasmic granules (11). As histamine provides an important signal to keratinocytes to produce prostanoids (12, 13), and in vivo studies by us and others with histamine receptor antagonists (14, 15) and indomethacin (13, 16) have highlighted directly or indirectly histamine and prostanoid involvement in UVB-induced immunosuppression, we investigated the distribution of histamine-staining mast cells by immunohistochemistry in the dermis and subcutaneous fatty tissue (SCFT, also known as the hypodermis, the hypodermal fat layer or the stratum adiposum) of C57BL/6J, DBA/2, BALB/c, and BALB/c-Uvs1 female mice and the correlation of these with their susceptibility to UVB-induced systemic suppression of CHS responses. Further, we investigated the susceptibility of mast cell–depleted mice to the immunomodulatory effects of both UV irradiation and administration of histamine. In these latter studies, the involvement of mast cells in the ability of UVB radiation to modulate local as well as systemic CHS responses was investigated, and also in a delayed type hypersensitivity (DTH) response to allogeneic spleen cells. The effect of reconstitution of the dorsal skin of mast cell–depleted mice with dermal mast cells on susceptibility to systemic immunomodulation by UVB was also examined.
Materials and Methods
Mice.
Pathogen-free, female BALB/c, DBA/2, and C57BL/6J mice were obtained (Animal Resource Center of the South Australian Department of Agriculture, Adelaide, Australia). Mice carrying the W f mutation on both the C57BL/6 and DBA/2 background were obtained originally from the Walter and Eliza Hall Institute of Medical Research (Melbourne, Australia; reference 17). The homozygous mast cell–depleted mice (W f/W f), the heterozygous mice (W f/+), and the wild-type mice (+/+) were the progeny of the mating of (C57BL/6W f × DBA/2W f) F1 hybrids, with the W f/W f, W f/+, and +/+ mice identified by coat color. The parental strains for the Uvs1 mice were obtained from the Frederick Cancer Research Facility (Frederick, MD). All mice were of equivalent age (69–80 d) and were not in hair growth as determined by histology and increased skin fold thickness. All experiments were performed according to the ethical guidelines of the National Health and Medical Research Council of Australia.
Immunohistochemical Staining for, and Quantification of, Histamine-containing Mast Cells.
Samples of dorsal skin (2–4/mouse) were rolled, fixed in 10% buffered formalin, and paraffin embedded ensuring vertical orientation of the roll, and 4-μm sections were cut and immunohistochemically stained with a 1/400 dilution of rabbit polyclonal antihistamine antibody (Chemicon, Temecula, CA). After incubation with a biotin-conjugated sheep anti–rabbit immunoglobulin (Silenus, Hawthorn, Australia) and peroxidase-conjugated streptavidin (Dako, Carpinteria, CA), the reaction product was developed with 3,3′-diaminobenzidine tetrahydrochloride (Sigma Chemical Co., St. Louis, MO). Sections were counterstained with hematoxylin solution. Control sections were incubated with diluted rabbit serum (1/400) or no primary antibody. The staining for histamine was specific as preincubation of the antibody with histamine significantly reduced the staining detected.
A video image analysis system (Video Pro 32; Leading Edge Pty Ltd., Adelaide, South Australia) was used to quantify histamine-staining cells in 15 consecutive fixed fields in each roll of skin. The epidermis was orientated horizontally and using a ×10 objective and a ×2.5 camera eyepiece; fields covering the epidermis through to the SCFT were selected. Mast cells (i.e., histamine-staining cells) and area of dermis and SCFT per fixed field of 800-μm width were measured (but expressed per mm); the coefficient of variation for measurement of mean dermal area for 15 fields was <14%. Dermal mast cell prevalence (see Figs. 1 A and 4 A) was determined by dividing the mast cell numbers per mm horizontal field length (Figs. 1 B and 4 B) by the dermal area for that field (Figs. 1 C and 4 C). In all experiments, the identity of the sections for examination by video image analysis was coded except for the positive control that was a section from skin rolls prepared from a C57BL/6J mouse. This positive control demonstrated minimal difference in staining intensity for mast cells from different staining runs. For measurement of the image background in all measurements of optical density, readings were standardized using a bright field image of the background illumination. This allowed correction of nonuniform illumination over a field of view, which would otherwise affect the discrimination of features by their gray value.
Figure 1.
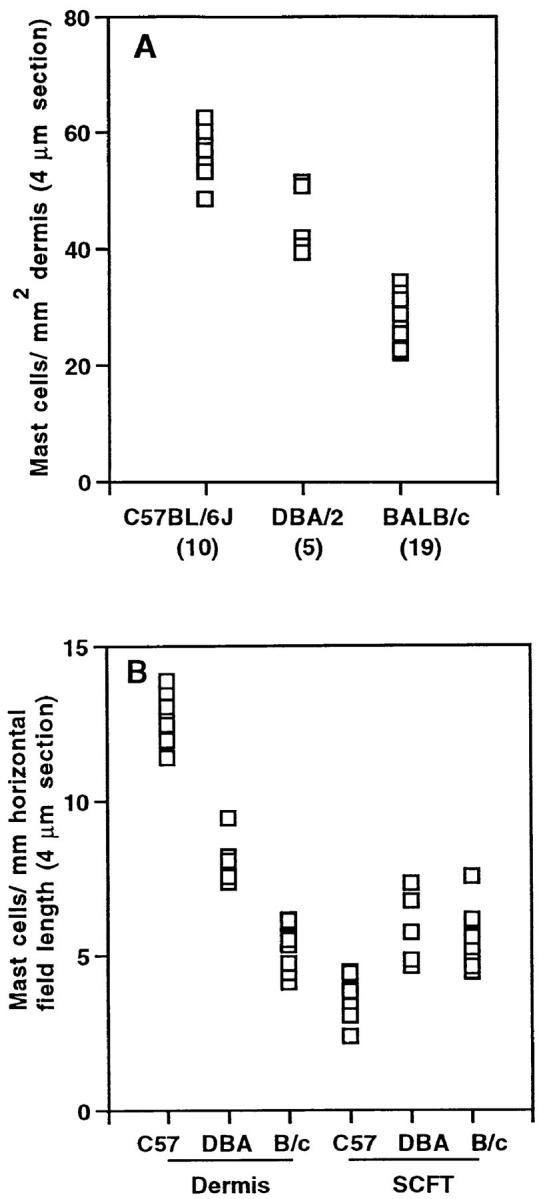
Mast cells in the dorsal skin of C57BL/6J, DBA/2, and BALB/c mice. (A) Mast cells per mm2 dermal area. (B) Mast cells per mm horizontal field length in the dermis and SCFT of C57BL/6J (C57; n = 10), DBA/2 (DBA; n = 5), and BALB/c (B/c; n = 11) mice. (C) The area (mm2) of the dermis and SCFT per mm of fixed field length for the same mice as shown in B. (D) The correlation between mast cell prevalence in the dermis (A) and susceptibility to UVB radiation for systemic suppression of CHS responses. Each point in A and B represents the mean mast cell number (expressed per area and fixed field length, respectively), and in C, area per fixed field length for 30–60 measurements/mouse. In A and D, the number of mice per group is shown in brackets. In D, the mean mast cell number per mm2 dermis for 4-μm sections (±SD) was plotted against the log of the UV dose (as documented by others [5]) required for 50% suppression of CHS responses.
Figure 4.



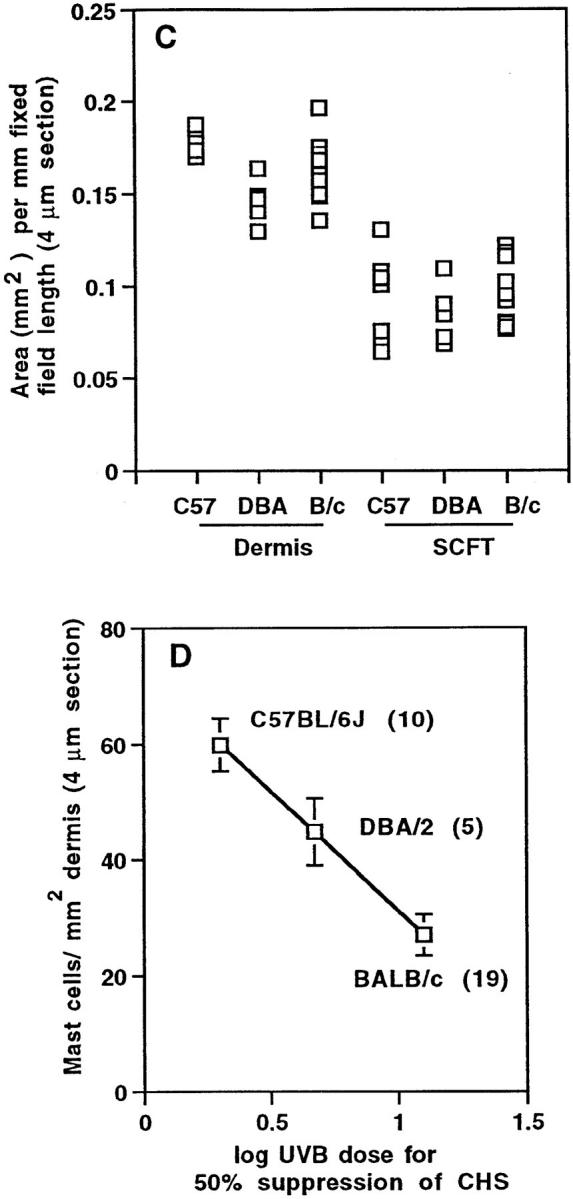
Mast cells in the dorsal skin of BALB/c and BALB/c-Uvs1 mice. (A) Mast cells per mm2 dermal area. (B) Mast cells per mm horizontal field length in the dermis and SCFT of BALB/c (–, n = 9) and BALB/c-Uvs1 (Uvs1, n = 11) mice. (C) The area (mm2) of the dermis per mm of fixed field length for the same mice as shown in B. Each point in A and B represents the mean mast cell number (expressed per area and fixed field length, respectively), and in C, area per fixed field length, for 30–60 measurements/mouse. In A and C, the number of mice per group is shown in brackets.
For quantification of the histamine content in each cell, the integrated optical density (representing the total amount of histamine staining in a given cell) was measured, as well as the area of histamine staining in each cell. For two rolls from each of six mice (three C57BL/6J and three BALB/c) all processed and stained within the same staining run, 200 mast cells were captured systematically in consecutive fields using a ×40 objective. The cells were categorized according to distribution in the upper or lower dermis (equal halves) or SCFT.
UV Irradiation.
The UV source was a bank of FS40 sunlamps (Westinghouse Corp., Pittsburgh, PA) emitting a broad band of UV, 250–360 nm, with 65% of the output in the UVB range (280–320 nm). A polyvinylchloride plastic film was used to screen out wavelengths of <290 nm. The dose rate was monitored using a UVX radiometer with a UVX-31 sensor (Ultraviolet Products Inc., San Gabriel, CA). For irradiation of the mice, a uniform dorsal area (8 cm2) was clean shaven, the ears protected with black adhesive insulation tape, and the mice housed in individual compartments of perspex cages. The sunlamps were held 20 cm above the cages.
Histamine Administration.
Histamine (0.2–200 μg/mouse; Sigma Chemical Co.) was dissolved in mouse osmolality PBS (330 mOsm/kg H2O) and injected subcutaneously into the backs of mice 5 d before sensitization with hapten. All mice received an equal volume of diluent, with or without histamine.
Assay of CHS.
For assay of systemic CHS responses, 5 d after UVB irradiation or histamine administration, mice (five per group) were sensitized on the shaved ventral skin with 100 μl of freshly prepared 5% (wt/vol) 2,4,6-trinitrochlorobenzene (TNCB; Tokyo Kasei Kogyo Ltd., Tokyo, Japan) in acetone. 5 d later and after coding the identities of the mouse groups, a CHS response was elicited by applying 10 μl freshly prepared 1% TNCB in acetone to each of the ventral and dorsal surfaces of both ears. 24 h after challenge, the ear thickness was measured with a micrometer (Mitutoyo Corp., Tokyo, Japan) and the extent of ear swelling for each mouse was calculated by subtracting the ear thickness before challenge. The mean swelling measured in mice that were challenged, but not sensitized, with TNCB (∼0.03 mm) was subtracted from this value. For local CHS responses, mice were sensitized 3 d after irradiation on the same dorsal site as UVB-irradiated with 100 μl freshly prepared 1% TNCB in acetone. The challenge of ears with TNCB was as described above.
Assay of DTH.
4 d after UVB irradiation, W f/W f and W f/+ mice were immunized by subcutaneous injection of 2.5 × 107 allogeneic spleen cells (from BALB/c mice) in 50 μl PBS into each flank of each mouse. 7 d later, the thickness of the hind footpads of the animals was measured with a micrometer before challenge with a subcutaneous injection of 107 allogeneic spleen cells in a volume of 40 μl PBS. After 24 h, the footpad thickness was measured again and the extent of swelling for each footpad was calculated by subtracting the footpad thickness before challenge.
Reconstitution of the Dorsal Skin of W f/W f Mice with Bone Marrow–derived Mast Cell Precursors.
Bone marrow cells from wild-type (+/+) mice were cultured at 106/ml in Eagles's MEM, alpha modification (alpha MEM; Sigma Chemical Co.) supplemented with 10−4 M 2-mercaptoethanol, 20% horse serum (Gilles Plains Animal Resource Centre, Adelaide, Australia), and 10% pokeweed-mitogen–stimulated spleen cell–conditioned medium (PWM-SCM). PWM-SCM was prepared as previously published (18) with 2 × 106 cells/ml incubated for 5 d in alpha MEM supplemented with 10% fetal calf serum, 10−5 M 2-mercaptoethanol, and 1:300 dilution of pokeweed mitogen (Sigma Chemical Co.). Culture flasks were incubated at 37°C in 5% CO2 with half the medium replaced every 7 d. After 31 d, the cells were washed and suspended in alpha MEM (2.5 × 106/ml) for injection in vivo. Each W f/W f mouse received 106 cells in 8 × 50 μl subcutaneous injections in two rows within a 1-cm width × 2-cm length of shaved dorsal skin. From 3.00 × 108 bone marrow cells harvested from 16 +/+ mice, 2.3 × 107 cells (7.6% yield) were harvested at the end of the 31-d culture period. The cultured cells (>85%) stained blue after staining with alcian blue–safranin (Csaba's mast cell stain; reference 19) indicating immature mast cells.
6 wk after reconstitution, an 8-cm2 dorsal area was shaved on the 18 W f/W f mice injected with bone marrow–derived mast cell precursors (10 male, 8 female). On the next day, 13 mice were irradiated with 12 kJ/m2 UVB and 5 served as controls. 5 d later, the mice were assessed for a systemic CHS response as described above. At the end of the systemic CHS assay, the mice were killed and dorsal skin sampled for evaluation of mast cell numbers. Mast cells in the dorsal skin of the bone marrow–reconstituted W f/W f mice were confirmed to be mature connective tissue mast cells by staining red with the alcian blue–safranin staining procedure (data not shown).
Expression of Results and Statistical Analysis.
For mast cell quantification and measurement of area of the dermis and SCFT, a mean value for each mouse was determined from 30–60 measurements. For measurement of integrated optical density and stained area per cell, the mean result from individual mice was used to calculate the mean value ± SD for three mice. A multiple comparison procedure using a one-way analysis of variance and Fisher's least significant difference test was used to determine the statistical significance (P <0.05) of differences between groups of mice from different strains. This form of analysis was also used for a test of significance between groups of mice within a CHS experiment. If the results from only two strains were compared (e.g., BALB/c versus BALB/c-Uvs1, C57BL/6J versus BALB/c for measurements of integrated optical density), an unpaired Student's t test was used. Strain-dependent effects of UVB or histamine on CHS to TNCB was examined by a multifactorial analysis of variance (MANOVA). Probabilities of <0.05 were considered significant.
Results
Dermal Mast Cells.
With the use of a video image analysis system, the prevalence of histamine-staining dermal mast cells (mast cells per mm2) was found to be significantly greater in C57BL/6J mice than in DBA/2 mice (P <0.05), which in turn had a significantly greater dermal mast cell prevalence than measured in BALB/c mice (P <0.05; Fig. 1 A). When mast cell numbers and tissue areas were considered separately in the different mouse strains, similar significant differences (P <0.05) were detected in dermal mast cell numbers per mm horizontal field length (Fig. 1 B). In the SCFT, there was no significant difference in the number of mast cells per mm horizontal field length of DBA/2 and BALB/c mice (Fig. 1 B). However, there was a significant decrease in mast cell prevalence in the SCFT of C57BL/6J mice (P <0.05; Fig. 1 B). The area of the dermis or SCFT per unit horizontal length of tissue for the three strains of mice examined did not vary significantly (Fig. 1 C). There was a linear relationship between the prevalence of histamine-staining dermal mast cells and the log of the dose of UVB demonstrated by others (5) to cause 50% suppression of CHS responses in the different mouse strains (Fig. 1 D). Representative histologic sections from BALB/c and C57BL/6J mice are shown in Fig. 2, A and B, respectively.
Figure 2.

Histologic sections of dorsal skin from representative mice. (A) BALB/c, (B) C57BL/6J. Arrows, Mast cells.
Differences in the histamine content of mast cells from C57BL/6J and BALB/c mice were investigated by measurement of the integrated optical density for histamine staining in 400 skin mast cells/mouse. The relative distribution of the 400 mast cells assessed varied for the different mouse strains (Fig. 3 A), as well as the tissue area required to be scanned in order to measure 400 mast cells. The mean area for histamine staining in mast cells from all mice was similar, the variation in stained areas approximating a normal distribution. The mean integrated optical density for histamine staining per cell was very similar for C57BL/6J and BALB/c mice (Fig. 3 B).
Figure 3.

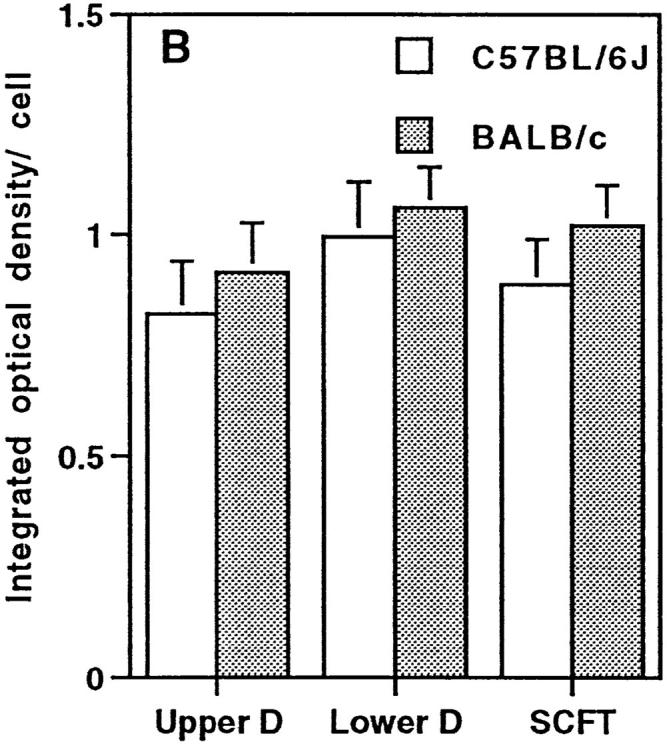
(A) Location of 400 mast cells from the dorsal skins of each of three C57BL/6J and three BALB/c mice examined for histamine content. The proportion of mast cells (mean + SD) in the upper dermis (D), lower dermis, and SCFT is shown. (B) Histamine content of these cells as assessed by measuring the integrated optical density (mean + SD) for individual histamine-stained cells.
BALB/c-Uvs1 Mice.
Mast cell numbers in the dermis and SCFT of BALB/c-Uvs1 mice and BALB/c mice of the same parental origin were investigated (Fig. 4). Within each of the three experiments (five BALB/c-Uvs1 and five BALB/c in experiment 1, three BALB/c-Uvs1 and two BALB/c in experiment 2, and three BALB/c-Uvs1 and two BALB/c in experiment 3), all BALB/c-Uvs1 animals were littermates. Dermal mast cell numbers were significantly higher in the BALB/c-Uvs1 mice (P = 0.0001; Fig. 4 A), although there were no differences in the numbers of mast cells in the SCFT (Fig. 4 B). It should be noted that variation in dermal mast cell numbers within the BALB/c- Uvs1 group occurred in all three experiments (three high and two low in experiment 1, two high and one low in experiment 2, and two high and one low in experiment 3). Dermal areas for the BALB/c-Uvs1 and BALB/c mice were not significantly different (Fig. 4 C).
Immunomodulation of Systemic CHS Responses.
To confirm that histamine was a mast cell product that could limit quantitatively UVB-induced immunosuppression in BALB/c (low susceptibility) versus C57BL/6J (high susceptibility) mice, the different mouse strains were UVB irradiated or injected subcutaneously with increasing doses of histamine 5 d before sensitization with the hapten, TNCB, and assay of a CHS response to that hapten (Fig. 5). The strain-dependent effects of UV on CHS to TNCB were significant by multifactorial analysis of variance, with 50% suppression at UVB doses as published (reference 5; Fig. 5 A). However, there was no difference between the strains at a dose of histamine that was immunosuppressive (20 μg/mouse; Fig. 5 B).
Figure 5.


The effect of (A) UVB and (B) histamine on the CHS responses of BALB/c and C57BL/6J mice to the hapten, TNCB. In A, mice were UVB irradiated on their shaved dorsal surface with increasing doses of UVB (290–320 nm) as indicated. In B, mice were injected subcutaneously on their backs with increasing doses of histamine per mouse. The mean change in ear swelling (+SD) for five mice/group is shown; the groups of mice were always coded before challenge with hapten and ear thickness measurements. *Result significantly different (P <0.05) to that of nonirradiated mice in A or control mice in B.
Mast Cell–depleted Mice.
Mast cell–depleted mice (W f/W f; reference 17) were then tested for susceptibility to UVB-induced systemic immunosuppression. The prevalence of dorsal dermal mast cells in the homozygous (W f/W f) mice was ∼12% of that measured in wild-type mice of the same genotype (Fig. 6 A). Systemic CHS responses by W f/W f mice, in contrast to heterozygous (W f/+) and wild-type (+/+) mice, were not suppressed by UVB at the dose of irradiation tested (Fig. 6 B). However, when the mast cell product histamine was exogenously introduced 5 d before hapten sensitization, CHS responses in these animals were reduced (Fig. 6 C).
Figure 6.



(A) Dermal mast cells per mm2 area in dorsal skin of W f/W f, heterozygous W f/+, and wild-type +/+ mice (n = three in each group). Quantification was as per Fig. 1 A. (B) The contact hypersensitivity response to TNCB by W f/W f mice after exposure to 12 kJ/m2 UVB. The mean result (+SD) for mice from three experiments is shown. (C) The CHS response to TNCB by W f/W f mice after UVB irradiation or administration of 200 and 2 μg histamine/ mouse. The mean result (+SD) for five mice/group is shown. In B and C, a result significantly different (*P <0.05) to that of nonirradiated mice of the same type.
In a manner similar to regulation of systemic CHS responses, UVB irradiation was able to suppress a DTH response to allogeneic spleen cells in heterozygous (W f/+), but not in homozygous (W f/W f), mice (representative of two experiments in Table 1). In contrast, mast cell depletion in mice did not affect downregulation of local CHS responses by UVB irradiation, i.e., CHS responses after sensitization with hapten applied to the irradiated site (representative of two experiments in Table 1).
Table 1.
Effect of UVB Irradiation on Immune Responses in Mast Cell–depleted Mice
| UVB (12 kJ/m2) | Nonsensitized | Sensitized | ||||
|---|---|---|---|---|---|---|
| − | − | + | ||||
| DTH responses to allogeneic spleen cells* | ||||||
| W f/+ | 0.01 ± 0.02 | 0.13 ± 0.01 | 0.02 ± 0.02‡ | |||
| W f/W f | 0.01 ± 0.01 | 0.10 ± 0.01 | 0.08 ± 0.01 | |||
| CHS responses to TNCB applied for sensitization at the irradiated site (local CHS)§ | ||||||
| W f/+ | 0.03 ± 0.02 | 0.26 ± 0.01 | 0.14 ± 0.01‡ | |||
| W f/W f | 0.03 ± 0.01 | 0.26 ± 0.01 | 0.13 ± 0.01‡ | |||
Mean swelling (mm) of the hind footpad (±SD, n = 4 mice, 8 footpads) 24 h after challenge.
P <0.05.
Mean swelling (mm) of ears (±SD, n = 5 mice, 10 ears) 24 h after challenge.
Susceptibility of W f/W f Mice, Reconstituted in the Dorsal Skin with Bone Marrow–derived Mast Cell Precursors, to Immunomodulation by UVB Irradiation.
Of the 18 W f/W f mice administered bone marrow–derived mast cell precursors, only one was judged to be poorly reconstituted after 8 wk (6 wk + 2 wk CHS assay; Fig. 7 A). In the reconstituted mice, the mast cells were found nonuniformly distributed in the dorsal skin and the data summarized in Fig. 7 A comprise an average for 15 consecutive fields from the skin of each mouse, some with numerous mast cells and others with very few. There was no significant difference in the dorsal mast cell numbers of reconstituted mice that had been either UVB irradiated or nonirradiated 12 d before skin sampling.
Figure 7.


Reconstitution of the dorsal skin of W f/W f mice with bone marrow–derived mast cell precursors. (A) Mast cells per mm horizontal field length in the dermis and SCFT of wild-type (+/+, n = 7), W f/W f (W f; n = 7), and W f/W f mice reconstituted with bone marrow–derived mast cell precursors (BMWf; n = 18). (B) The effect of UVB (12 kJ/m2) on the systemic CHS response to the hapten, TNCB, by wild-type (+/+), W f/W f, and W f/W f mice reconstituted with bone marrow–derived mast cell precursors (BMW f/W f). The mean change in ear swelling (+SD) for 5 mice/group is shown with the exception of 12 UVB-irradiated BMW f/ W f mice; the groups of mice were coded before challenge with hapten and ear thickness measurements. *A result significantly different (P <0.05) to that of nonirradiated mice.
Susceptibility to UVB (12 kJ/m2) for systemic suppression of CHS responses was evaluated in 13 mast cell–reconstituted W f/W f mice. The CHS response by all mice except one was significantly suppressed by UVB irradiation; this mouse was the one shown in Fig. 7 A to be poorly reconstituted and the results for this mouse have been excluded from the results of Fig. 7 B.
Discussion
The evidence supporting the hypothesis that mast cells determine susceptibility to UVB-induced systemic immunosuppression is of two types. (a) In different strains of mice, susceptibility to UVB immunosuppression correlated with dermal mast cell prevalence and the prevalence in W f/W f mice was insufficient to mediate susceptibility. This established a correlation. (b) The ability of reconstitution of the dorsal skin of W f/W f mice with mast cells to confer susceptibility to UVB-induced systemic immunosuppression confirmed the functional role of mast cells in this response, and eliminated the concern that the lack of responsiveness of unreconstituted W f/W f mice to UVB might be the result of other abnormalities (anemia, albino nature) of these mice. The results with the albino BALB/c- Uvs1 mice also argued against the possibility that pigment was contributing in this study to the extent of susceptibility of mice to systemic immunomodulation by UVB. In a previous study, it was confirmed that Uvs1 was not genetically linked to 12 chromosomal markers including the pigment genes b (brown) and c (albino) (8).
By use of a combination of histamine types 1 and 2 receptor antagonists, we have previously shown that histamine is an important regulatory molecule in the pathway(s) by which UVB regulates systemic CHS responses in mice (14). The source of the histamine had not been confirmed. These studies implicate mast cell–derived histamine as an important contributor to a pathway that determines degree of susceptibility of mice to UVB-induced systemic, but not local, suppression of CHS responses. The histamine content of dermal mast cells as estimated using quantitative video image analysis did not vary significantly for different mouse strains. However, there was greater than a twofold increase in mast cell prevalence in the dermis of the C57BL/6J mice, i.e., UV highly susceptible mice, than in the dermis of the poorly susceptible BALB/c mice. In fact, there was a direct correlation between dermal mast cell prevalence and susceptibility to UVB-induced systemic suppression of CHS responses. Dermal mast cells in the BALB/c-Uvs1 mice were also increased to a level similar to that of DBA/2 mice (Fig. 1 A). The level of mast cell prevalence is consistent with the previously reported extent of suppression of CHS responses in BALB/c-Uvs1 mice with 4.5 kJ/m2 UVB (8). Overall, these objective measures of histamine in stained sections suggest a correlation between total histamine that may be released from dermal mast cells after degranulation and the extent of systemic immunomodulation after UVB exposure.
That histamine was a component of a pathway defining degree of susceptibility of mice to UVB-induced systemic suppression of CHS responses was supported by further in vivo experiments. C57BL/6J and BALB/c mice differed in their susceptibilities to UVB for decreased CHS responses, but were similar in their susceptibilities to histamine-induced suppression of CHS responses. In addition, systemic CHS responses by mast cell–depleted mice (W f/W f) were not suppressed after UVB irradiation, but were susceptible to suppression by histamine. If keratinocytes were producing histamine after UV irradiation as recently reported in vitro (20), it was not sufficient to suppress the CHS response in W f/W f mice. We believe that we have administered physiological concentrations of histamine to the mice; we estimated that the dermis of 8 cm2 dorsal skin (∼270 μm deep, ∼40 mast cells/mm2 horizontal section of dermis, and 3–5 pg histamine/mast cell; reference 20) contains ∼8 μg histamine. This must be considered a crude estimation of available histamine, as not all histamine would be released from degranulating mast cells. Whether histamine was the only important mediator from mast cells was not established. A low, but significant, level of UVB-induced immunosuppression was still detected in the mice administered histamine receptor antagonists (14). The potential contribution of mast cell–derived TNF-α and IL-4 to UVB-associated immunomodulation is currently under investigation. However, it may be that there was insufficient histamine receptor antagonist available at histamine's site(s) of action to ensure total inhibition.
UVB was also unable to suppress DTH responses in the W f/W f mice and implicates mast cells and their products in the regulation of this other systemic response. In contrast, there was no evidence for mast cell involvement in the modulation by UVB of CHS responses when the sensitizing hapten was applied to the irradiated site. These results suggest that antigen-presenting cells at a distant, nonirradiated site, but not at the irradiated site, are susceptible to immunomodulation, directly or indirectly, by mast cell products.
It is uncertain whether UV triggers dermal mast cell degranulation directly or indirectly. Signals from sensory nerve fibres originating in the epidermis may be involved (21). Trans-urocanic acid (UCA) is a histidine-derived product in the stratum corneum of the skin (22) and has been implicated as a photoreceptor for UV-induced immunosuppression (23). There has been a report that cis-UCA, an immunosuppressive molecule and the product of UV-induced isomerization of trans-UCA, can stimulate mast cell degranulation (24). Mast cell degranulation induced by cis-UCA would support observations by us assaying systemic CHS responses (our unpublished data) and by others (25) of differences in response to cis-UCA by BALB/c and C57BL/6J mice, the latter being better responders to cis-UCA.
The mechanisms by which histamine alone, or together with other mast cell or keratinocyte products, signals systemic immunosuppression are not known. Histamine stimulates prostaglandin E2 production by cultured human keratinocytes (12, 13); prostaglandin E2 may act locally on epidermal cells (26), in lymph nodes (27), or systemically (28). Further, by indomethacin administration to mice, prostanoids have been shown to be responsible for at least 50% of the systemic suppression of CHS responses by UVB irradiation (13). There is also evidence that DNA is a UVB photoreceptor that initiates immunosuppression (29, 30). Damage to DNA can regulate the transcription in epidermal cells of regulatory cytokines (e.g., IL-10 [31], TNF-α [7, 32]) and adhesion molecules that enable cell–cell interaction (e.g., intercellular adhesion molecule 1 [ICAM-1]; reference 33). However, whether there is a relationship between histamine and other mast cell products, and cyclobutyl pyrimidine dimer formation in DNA of UVB-irradiated cells (30), and the enzymes responsible for dimer repair, is not known. With respect to UV-induced IL-10 production, it has been reported that UVB can suppress systemic CHS responses, but not DTH responses, in IL-10–deficient mice (34) and suggests that IL-10 may not be an important molecule in determining responses to UVB irradiation in W f/ W f mice. Histamine may directly influence the migration of immune cells into the UV-irradiated site, or to nearby lymph nodes, and regulate their proliferation and cytokine profiles (35–38).
In conclusion, our results show that mast cells contribute critically to the manifestation of UVB-induced systemic suppression of CHS and DTH responses. Further, they add to our previous suggestion that histamine is one important component of the pathway to immunosuppression (14) and indicates mast cells as the source of the histamine. Downregulation of systemic immune responses in mice after UVB exposure correlated directly with the prevalence of histamine-containing dermal mast cells. In addition, the strains of mice investigated were equivalently susceptible to the immunomodulatory effects of exogenous histamine, further emphasizing the relationship of dermal mast cell prevalence with susceptibility to UV systemic immunomodulation. The mechanism(s) by which histamine, and potentially other mast cell products, can modulate immune reactivity remains an unanswered question. The answers will be very important in our quest to understand and prevent or treat the harmful effects of UVB that are becoming of greater concern due to greater ozone depletion in the earth's stratosphere.
Acknowledgments
We are grateful to Mr. J. Brennan and Ms. K. Clark for performing the Csaba's mast cell stain, and to our colleagues for critical reading of the manuscript.
This work was supported by the Anti-Cancer Foundation of South Australia, the National Health and Medical Research Council of Australia, the Flinders Medical Centre Foundation (to P.H. Hart and J.J. Finlay-Jones) and in part by National Institutes of Health grant RO1 CA-53765 (to F.P. Noonan).
Abbreviations used in this paper
- CHS
contact hypersensitivity
- DTH
delayed type hypersensitivity
- SCFT
subcutaneous fatty tissue
- TNCB
2,4,6-trinitrochlorobenzene
- UCA
urocanic acid
- UVB
UV radiation of wavelength 280–320 nm
References
- 1.Kripke ML. Immunologic mechanisms in UV-radiation carcinogenesis. Adv Cancer Res. 1981;34:69–106. doi: 10.1016/s0065-230x(08)60239-0. [DOI] [PubMed] [Google Scholar]
- 2.Mottram PL, Mirisklavos A, Clunie GJA, Noonan FP. A single dose of UV radiation suppresses delayed type hypersensitivity responses to alloantigens and prolongs heart allograft survival in mice. Immunol Cell Biol. 1988;66:377–385. doi: 10.1038/icb.1988.49. [DOI] [PubMed] [Google Scholar]
- 3.Gilmour JW, Vestey JP, Norval M. The effect of UV therapy on immune function in patients with psoriasis. Br J Dermatol. 1993;129:28–38. doi: 10.1111/j.1365-2133.1993.tb03307.x. [DOI] [PubMed] [Google Scholar]
- 4.Yoshikawa T, Streilein JW. Genetic basis of the effects of ultraviolet light B on cutaneous immunity. Evidence that polymorphism at the Tnfα and Lpsloci governs susceptibility. Immunogenetics. 1990;32:398–405. doi: 10.1007/BF00241633. [DOI] [PubMed] [Google Scholar]
- 5.Noonan FP, Hoffman HA. Susceptibility to immunosuppression by ultraviolet B radiation in the mouse. Immunogenetics. 1994;39:29–39. doi: 10.1007/BF00171794. [DOI] [PubMed] [Google Scholar]
- 6.Streilein JW, Taylor JR, Vincek V, Kurimoto I, Richardson J, Tie C, Medema J-P, Golomb C. Relationship between ultraviolet radiation–induced immunosuppression and carcinogenesis. J Invest Dermatol. 1994;103:107S–111S. doi: 10.1111/1523-1747.ep12399400. [DOI] [PubMed] [Google Scholar]
- 7.Vincek V, Kurimoto I, Medema JP, Prieto E, Streilein JW. Tumor necrosis factor α polymorphism correlates with deleterious effects of ultraviolet B light on cutaneous immunity. Cancer Res. 1993;53:728–732. [PubMed] [Google Scholar]
- 8.Noonan FP, Hoffman HA. Control of UVB immunosuppression in the mouse by autosomal and sex-linked genes. Immunogenetics. 1994;40:247–256. doi: 10.1007/BF00189969. [DOI] [PubMed] [Google Scholar]
- 9.Gilchrest BA, Soter NA, Stoff JS, Mihm MC. The human sunburn reaction: histologic and biochemical studies. J Am Acad Dermatol. 1981;5:411–422. doi: 10.1016/s0190-9622(81)70103-8. [DOI] [PubMed] [Google Scholar]
- 10.Ikai K, Danno K, Horio T, Narumiya S. Effect of ultraviolet irradiation on mast cell–deficient W/Wv mice. J Invest Dermatol. 1985;85:82–84. doi: 10.1111/1523-1747.ep12275365. [DOI] [PubMed] [Google Scholar]
- 11.Galli SJ. New insights into “the riddle of the mast cells”: microenvironmental regulation of mast cell development and phenotypic heterogeneity. Lab Invest. 1990;62:5–33. [PubMed] [Google Scholar]
- 12.Pentland AP, Mahoney M, Jacobs SC, Holtzman MJ. Enhanced prostaglandin synthesis after ultraviolet injury is mediated by endogenous histamine stimulation. A mechanism for irradiation erythema. J Clin Invest. 1990;86:566–574. doi: 10.1172/JCI114746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jaksic A, Finlay-Jones JJ, Watson CJ, Spencer LK, Santucci I, Hart PH. Cis-urocanic acid synergizes with histamine for increased PGE2production by human keratinocytes: link to indomethacin-inhibitable UVB-induced immunosuppression. Photochem Photobiol. 1995;61:303–309. doi: 10.1111/j.1751-1097.1995.tb03976.x. [DOI] [PubMed] [Google Scholar]
- 14.Hart PH, Jaksic A, Swift G, Norval M, El-Ghorr AA, Finlay-Jones JJ. Histamine involvement in UVB– and cis–urocanic acid–induced systemic suppression of contact hypersensitivity responses. Immunology. 1997;91:601–608. doi: 10.1046/j.1365-2567.1997.00284.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Norval M, Gilmour JW, Simpson TJ. The effect of histamine receptor antagonists on immunosuppression induced by the cis-isomer of urocanic acid. Photodermatol Photoimmunol Photomed. 1990;7:243–248. [PubMed] [Google Scholar]
- 16.Chung H-T, Burnham DK, Robertson B, Roberts LK, Daynes RA. Involvement of prostaglandins in the immune alterations caused by exposure of mice to ultraviolet radiation. J Immunol. 1986;137:2478–2484. [PubMed] [Google Scholar]
- 17.Thomas WR, Schrader JW. Delayed hypersensitivity in mast-cell–deficient mice. J Immunol. 1983;130:2565–2567. [PubMed] [Google Scholar]
- 18.Nakano T, Sonoda T, Hayashi C, Yamatodani A, Kanayama Y, Yamamura T, Asai H, Yonezawa T, Kitamura Y, Galli SJ. Fate of bone marrow–derived cultured mast cells after intracutaneous, intraperitoneal, and intravenous transfer into genetically mast cell–deficient W/Wvmice. Evidence that cultured mast cells can give rise to both connective tissue type and mucosal mast cells. J Exp Med. 1985;162:1025–1043. doi: 10.1084/jem.162.3.1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Culling, C.F.A., and W.L. Dunn. 1974. Handbook of Histopathological and Histochemical Techniques (Including Museum Techniques). 3rd ed. Butterworths & Co., London. 419 p.
- 20.Malaviya R, Morrison AR, Pentland AP. Histamine in human epidermal cells is induced by ultraviolet light injury. J Invest Dermatol. 1996;106:785–789. doi: 10.1111/1523-1747.ep12346356. [DOI] [PubMed] [Google Scholar]
- 21.Eschenfelder CC, Benrath J, Zimmerman M, Gillardon F. Involvement of substance Pin ultraviolet irradiation–induced inflammation in rat skin. Eur J Neurosci. 1995;7:1520–1526. doi: 10.1111/j.1460-9568.1995.tb01147.x. [DOI] [PubMed] [Google Scholar]
- 22.Noonan FP, De Fabo EC. Immunosuppression by ultraviolet B radiation: initiation by urocanic acid. Immunol Today. 1992;13:250–254. doi: 10.1016/0167-5699(92)90005-R. [DOI] [PubMed] [Google Scholar]
- 23.De Fabo EC, Noonan FP. Mechanism of immune suppression by ultraviolet irradiation in vivo. I. Evidence for the existence of a unique photoreceptor in skin and its role in photoimmunology. J Exp Med. 1983;158:84–98. doi: 10.1084/jem.158.1.84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wille JJ, Kydonieus A. Abrogation of contact hypersensitivity in mice by topically-applied mast cell degranulating agents. J Invest Dermatol. 1995;104:679. . (Abstr.) [Google Scholar]
- 25.Kurimoto I, Streilein JW. Cis-urocanic acid suppression of contact hypersensitivity induction is mediated via tumor necrosis factor–α. J Immunol. 1992;148:3072–3078. [PubMed] [Google Scholar]
- 26.Grewe M, Klammer M, Krutmann J. The role of prostaglandin E2 (PGE2) in ultraviolet B radiation (UVBR)-induced inhibition of ICAM-1 expression in human antigen presenting cells (APC) J Invest Dermatol. 1995;105:498. . (Abstr.) [Google Scholar]
- 27.Van der Pouw Kraan, T.C.T.M., L.C.M. Boeije, R.J.T. Smeenk, J. Wijdenes, and L.A. Aarden. Prostaglandin-E2is a potent inhibitor of human interleukin 12 production. J Exp Med. 1995;181:775–779. doi: 10.1084/jem.181.2.775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Phipps PP, Stein SH, Roper RL. A new view of prostaglandin E regulation of the immune response. Immunol Today. 1991;12:349–351. doi: 10.1016/0167-5699(91)90064-Z. [DOI] [PubMed] [Google Scholar]
- 29.Applegate LA, Ley RD, Alcalay J, Kripke ML. Identification of the molecular target for the suppression of contact hypersensitivity by ultraviolet radiation. J Exp Med. 1989;170:1117–1131. doi: 10.1084/jem.170.4.1117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kripke ML, Cox PA, Alas LG, Yarosh DB. Pyrimidine dimers in DNA initiate systemic immunosuppression in UV-irradiated mice. Proc Natl Acad Sci USA. 1992;89:7516–7520. doi: 10.1073/pnas.89.16.7516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Vink AA, Yarosh DB, Kripke ML. Chromophore for UV-induced immunosuppression: DNA. Photochem Photobiol. 1996;63:383–386. doi: 10.1111/j.1751-1097.1996.tb03050.x. [DOI] [PubMed] [Google Scholar]
- 32.Hejmadi V, Kibitel J, O'Connor A, Sutherland B, Beutler B, Klein J, Yarosh D. DNA damage induced by UV is a signal for increased TNFα expression in human cells. Photochem Photobiol. 1996;63:110S. . (Abstr.) [Google Scholar]
- 33.Krutmann J, Ahrens C, Roza L, Arlett CF. The role of DNA damage and repair in ultraviolet B radiation–induced immunomodulation: relevance to human photocarcinogenesis. Photochem Photobiol. 1996;63:394–396. doi: 10.1111/j.1751-1097.1996.tb03053.x. [DOI] [PubMed] [Google Scholar]
- 34.Beissert S, Hosoi J, Kulm R, Rajewsky K, Muller W, Granstein RD. Impaired immunosuppressive response to ultraviolet radiation in interleukin-10–deficient mice. J Invest Dermatol. 1996;107:553–557. doi: 10.1111/1523-1747.ep12582809. [DOI] [PubMed] [Google Scholar]
- 35.Laberge S, Cruikshank WW, Kornfeld H, Center DM. Histamine-induced secretion of lymphocyte chemoattractant factor from CD8+ T cells is independent of transcription and translation. Evidence for constitutive protein synthesis and storage. J Immunol. 1995;155:2902–2910. [PubMed] [Google Scholar]
- 36.Bury TB, Corhay JL, Radermecker MF. Histamine-induced inhibition of neutrophil chemotaxis and T-lymphocyte proliferation in man. Allergy. 1992;47:624–629. doi: 10.1111/j.1398-9995.1992.tb02385.x. [DOI] [PubMed] [Google Scholar]
- 37.Falus A, Meretey K. Histamine: an early messenger in inflammatory and immune reactions. Immunol Today. 1992;13:154–156. doi: 10.1016/0167-5699(92)90117-p. [DOI] [PubMed] [Google Scholar]
- 38.Lagier B, Lebel B, Bousquet J, Pene J. Different modulation by histamine of IL-4 and interferon-gamma (IFN-γ) release according to the phenotype of human Th0, Th1 and Th2 clones. Clin Exp Immunol. 1997;108:545–551. doi: 10.1046/j.1365-2249.1997.3791276.x. [DOI] [PMC free article] [PubMed] [Google Scholar]