

Abstract
Calcium ion transiently blocks Na+ channels, and it shortens the time course for closing of their activation gates. We examined the relation between block and closing kinetics by using the Na+ channels natively expressed in GH3 cells, a clonal line of rat pituitary cells. To simplify analysis, inactivation of the Na+ channels was destroyed by including papain in the internal medium. All divalent cations tested, and trivalent La3+, blocked a progressively larger fraction of the channels as their concentration increased, and they accelerated the closing of the Na+ channel activation gate. For calcium, the most extensively studied cation, there is an approximately linear relation between the fraction of the channels that are calcium-blocked and the closing rate. Extrapolation of the data to very low calcium suggests that closing rate is near zero when there is no block. Analysis shows that, almost with certainty, the channels can close when occupied by calcium. The analysis further suggests that the channels close preferentially or exclusively from the calcium-blocked state.
Voltage-gated Na+ channels have positively charged transmembrane helices, the S4 segments, that move in response to changes of membrane voltage, driving conformational changes within each channel that open or close its activation gate. According to current views (1, 2), conformational changes in each of the four domains of a Na+ channel, or each subunit of a K+ channel, are followed by a “concerted” step, a single event, that opens the gate, allowing flux through the pore. The S4 movements associated with the early conformational changes have been detected with labeling experiments (3, 4), but the nature of the concerted step is unknown. In this paper and the following one, the possibility is explored that the concerted step for a Na+ channel consists of liberating a calcium ion bound in the channel lumen.
Calcium and other divalent cations have well-documented effects on the properties of Na+ channels (5). These effects were conveniently summarized by Frankenhaueser and Hodgkin (6), who said that increasing external calcium concentration 10-fold has about the same effect as hyperpolarizing the membrane by 15 mV, thus taking nerve fibers further from threshold and making them harder to excite. Stated another way, calcium appears to stabilize the resting state, making the channels less likely to open. Two other experimental findings regarding calcium were subsequently added. First, it was found that calcium permeates measurably through Na+ channels, about 1/100th as well as Na+ (7). Second, Woodhull (8) showed that high calcium concentrations have a distinct blocking action on sodium current through open Na+ channels. Putting these last two phenomena together, it appears that calcium enters Na+ channels, but permeates slowly, thus interfering with Na+ flux through the channels.
The effects of calcium on gating kinetics and open probability, as described by Frankenhaeuser and Hodgkin (6), are usually explained by the surface charge hypothesis. This hypothesis holds that calcium alters gating by neutralizing negative charge at the membrane surface, thus changing the local field near the voltage-sensing parts of the Na+ or K+ channels (5). The permeation-blocking effects are regarded as separate phenomena, having no effect on kinetics and open probability. An obvious alternative not ruled out by existing data is that calcium’s effects on gating are associated with its ability to occupy and block Na+ channels, and that it is calcium occupancy rather than a surface charge mechanism that stabilizes the closed state when external calcium concentration is increased. In support of this idea, we show here that calcium block and closing rate of Na+ channels are closely related, and that Na+ channels close freely, and perhaps preferentially, when calcium occupied. The following paper (9) shows that calcium has large effects on gating only in cases where it is free to enter and leave the channel, and that calcium seems to be essential for channel closing.
METHODS
Experiments were performed on GH3 cells, a rat pituitary cell line, using methods described previously (10). The cells were cultured to maximize Na+ channel expression and were whole-cell patch-clamped. The internal medium contained 1 mg/ml papain to destroy fast inactivation of Na+ channels. Current through other channels was suppressed by removal of K+ and by spontaneous decay of calcium current. The remaining current was almost exclusively through Na+ channels, and current resistant to 1 μM tetrodotoxin was very small.
To improve time resolution, we used very low resistance pipettes (0.4 to 0.7 MΩ) and “supercharging” (10). The calculated series resistance error was <5 mV. The holding potential was −80 mV in all experiments, and the temperature was 15°C. Linear leak and capacitive current have been subtracted from all records. Na+ conductance, gNa, was computed by dividing Na+ current (INa) by the driving force, Vm − VNa.
All records presented were bracketed by averaging a control measurement taken before and after exposure to a test solution and accepting only experiments where the brackets matched closely. External solution names and contents (in mM) are in Table 1. All solutions had pH of 7.3. The internal solution contained 135 mM Na+, 32 mM Cl−, 100 mM F−, 10 mM EGTA, 1 mM Ca2+, and 10 mM Hepes.
Table 1.
External solutions
| Name | [Na+], mM | [Ca2+], mm | [Cl−], mM | [Hepes], mM | [Other], mM |
|---|---|---|---|---|---|
| 2Ca 85Na | 85 | 2 | 84 | 10 | 144 sucrose |
| 10Ca 85Na | 85 | 10 | 100 | 10 | 120 sucrose |
| 20Ca 85Na | 85 | 20 | 120 | 10 | 100 sucrose |
| 50Ca 85Na | 85 | 50 | 180 | 10 | |
| 2Ca 155Na | 155 | 2 | 154 | 10 | |
| 20Ca 128Na | 128 | 20 | 163 | 10 | |
| 20Ba 128Na | 128 | 163 | 10 | 20 Ba2+ |
RESULTS
The two major effects of calcium on Na+ channels, blocking and alteration of gating kinetics, are illustrated in Fig. 1. The cell, under voltage-clamp, was depolarized from −80 mV to +60 mV to open the Na+ channels, then was repolarized to −80 mV. The instantaneous value of the conductance, gNa, is given by
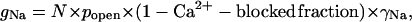 |
where N is the total number of Na+ channels, popen is the probability that the channels have open gates, and γNa is the conductance of a single Na+ channel when not calcium blocked. The conductance of a Ca2+-blocked channel is taken as zero. The step to +60 mV is large enough to increase open probability to its maximum value in both calcium concentrations. At the end of the activating step, gNa is lower in 50 mM Ca2+ than in 2 mM Ca2+ because about one-fourth of the channels are blocked in the higher calcium concentration. Immediately after the step from +60 to −80 mV, gNa in 2 mM Ca2+ drops from 198 to 116 nS (145 to 17 nS in 50 mM Ca2+) as Ca2+ is drawn into blocking position in the pore by increased internal negativity. The equilibration of Ca2+ with the channels is too fast to resolve (for free entry of Ca2+ into a pore, the theoretical equilibration time constant at 2 mM Ca2+ is less than 1 μs), so the decrease appears to be instantaneous. It is followed in both solutions by a decaying tail of conductance as the activation gates of the Na+ channels close. It is clear that the tail in 50 mM Ca2+ is both smaller, reflecting more block, and faster, reflecting accelerated closing kinetics of the gate.
Figure 1.

Calcium blocks Na+ channels and speeds deactivation. Gates of the Na+ channels were activated by stepping to +60 mV and were deactivated by returning to −80 mV. gNa is proportional to the number of Na+ channels that have open (activated) gates and are not Ca2+-blocked. Compared with 2 mM Ca2+, 50 mM Ca2+ blocks weakly at +60 mV, strongly at −80 mV. Deactivation is clearly faster in 50 mM Ca2+. Solutions: 2Ca 85Na and 50Ca 85Na.
The phenomena of block and altered gating kinetics are qualitatively the same for all of the divalent cations we have tested, and for the trivalent cation lanthanum. As an example, data from an experiment comparing calcium with barium ion is shown in Fig. 2. Changing from 2 mM Ca2+ to 20 mM Ba2+ (no added Ca2+) greatly increased the drop in conductance on changing from +60 to −60 mV, and it speeded the subsequent decay of the conductance (decay rate 0.8 ms−1 in 2 mM Ca2+, 1.6 ms−1 in 20 mM Ba2+; in 2 mM Ca2+ the closing rate is slower than in Fig. 1 because the repolarization voltage is −60 rather than −80 mV). As is clear from Fig. 2, 20 mM Ba2+ was similar in effect to 20 mM Ca2+, but block by the latter was a bit more pronounced, and the decay rate was faster (2.2 ms−1). All of the other divalent cations tested (Sr2+, Mg2+, Zn2+) and La3+ also block Na+ channels, and they accelerate closing kinetics when the block is made stronger by increased concentration. This wide range of adequate divalent substitutes may seem surprising, but the crystal radii of the divalent cations (Mg2+, 0.66 Å to Ba2+, 1.34 Å) has the same span as the radii of monovalent cations that easily permeate the Na+ channel (Li+, 0.68 Å to K+, 1.33 Å; see ref. 5).
Figure 2.

Ba2+ is an adequate substitute for Ca2+. The traces show activation and deactivation of gNa in calcium at two concentrations, and after complete replacement of calcium by barium. Channel block in 20 mM Ba2+ is slightly less than in 20 mM Ca2+ (as can be seen from the initial value of the conductance tails at −60 mV) and deactivation is slightly slower. Solutions: 2Ca 155Na, 20Ca 128Na, and 20Ba 128Na.
Both Figs. 1 and 2 suggest a correlation between the closing rate of the channel gates and the fraction of channels that are calcium-blocked. This was examined in more detail in Fig. 3. Each trace shows the conductance tail at −80 mV, after a large activating pulse to +60 mV had increased open probability to its maximum value. As Ca2+ is increased from 2 to 10, 20, and 50 mM, the initial value of conductance at −80 mV becomes steadily smaller. This decrease reflects increasing block by Ca2+, and it was completely reversible. As the tails get smaller, they are also quicker in time course, showing that closing of the activation gates is faster.
Figure 3.

Conductance tails in four calcium concentrations. Block, judged from initial tail amplitude, increases with calcium concentration. Deactivation is progressively faster as block increases. Solutions: 2, 10, 20, and 50Ca 85Na.
The correlation between closing rate of the channel gates and the Ca2+-blocked fraction is made clear in Fig. 4. The blocked fraction, which is known to follow a simple saturation curve with Kd of about 7 mM (11), was determined from the initial values of the conductance tails in Fig. 3, by the method used previously (11). The closing rate at −80 mV with inactivation removed is closely approximated from the reciprocal of the time constant of the tail. The plot shows that closing rate increases approximately linearly as a function of the fraction of the channels that are blocked by Ca2+ over the range from 2 to 50 mM Ca2+. Extrapolating the curve to lower fraction blocked (lower calcium concentration) suggests that the closing rate would be near zero when there is no block. Clearly it would be of interest to know the closing rate at lower calcium concentration, but cells were almost always damaged irreversibly by Ca2+ below 2 mM. As reported in the next paper (9), low-calcium experiments were more successful with squid giant axons. The broken line is predicted by the model in Discussion.
Figure 4.

Closing rate is roughly proportional to the fraction of the channels that are calcium-blocked. Using the data of Fig. 3, we estimated the fraction of blocked channels from initial tail amplitude. Closing rate was measured as 1/τ—i.e., the reciprocal of the time constants of the conductance tails at −80 mV. The broken line is the prediction of the model in Discussion.
DISCUSSION
The data presented above make strong suggestions about the ion occupancy of Na+ channels as they are closing and, by extension, in the closed state. The following state diagram is helpful in considering this question:
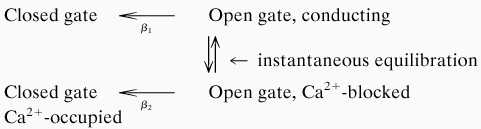 |
In the two states on the left, the activation gate is closed, and the channel is calcium free (upper) or calcium-occupied. The gate is open for the two states on the right, but only the upper state is conducting. According to the diagram, the rate constant for gate-closing of a channel is β2 ms−1 if the channel is Ca2+-blocked, and β1 ms−1 if it is not. The rates for the reverse transitions, closed to open gate, are negligible at −80 mV. The channels with open gates are distributed between conducting and Ca2+-blocked states. In 2 mM Ca2+ about 71% of the open-gated channels are in the conducting state immediately after stepping from +60 to −80 mV, and about 29% are in the Ca2+-blocked state. The rate constants for the transitions from conducting to Ca2+-blocked are so fast that the two states are always in equilibrium.
Consider first the possibility that a channel cannot close when it contains a Ca2+ ion; i.e., rate constant β2 is zero. The closing time constant is then given by
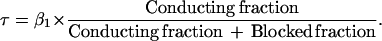 |
When Ca2+ is increased from 2 to 20 mM, the conducting fraction (of the open-gated channels) at −80 mV decreases from 0.71 to 0.28. If blocking is the only effect of Ca2+ (i.e., if β1 is unaffected) the overall closing rate would decrease by a factor of 2.5. Fig. 5 shows the experimental traces in 2 and 20 mM Ca2+, and, in curve a, the predicted closing time course for the case just outlined. This is a very poor fit, and it is clear that if the Ca2+-blocked channels cannot close, then β1 must increase.
Figure 5.

Conductance tail in 20 mM Ca2+, fitted by several models. Curve a shows the predicted time course if Ca2+-blocked channels cannot close, and the closing rate constant (β1) is unaffected by calcium. For curve b, β1 is increased by a factor predicted from surface charge theory. Please see text. Solutions: 2 and 20Ca 85Na.
The surface charge theory of calcium action predicts an increase of β1 as Ca2+ increases and makes the external surface of the membrane more positive. The predicted change however, is not large enough to rescue the idea that only Ca2+-free channels can close, as the following reasoning shows. Changing Ca2+ from 2 to 20 mM in this experiment shifted the g–V curve by +21 mV (data not shown). The same shift would apply to tail kinetics. When combined with the known voltage dependence of the closing rate, a +21-mV shift predicts an increase of β1 somewhere between 2.7 and 3.8×. [The uncertainty arises because the rate constant changes more rapidly with voltage in 2 mM Ca2+ than in 20 mM Ca2+. This fact is examined in the next paper (9).] Even the higher factor results in a poor fit (curve b). A good fit requires that β1 be increased by 7.2×, as shown by curve c, which is almost twice the increase predicted by surface charge theory. We are forced to abandon the idea that Ca2+-blocked channels cannot close.
If β2 is not zero, the values of β1 and β2 cannot be rigorously determined from the experimental information available. An interesting suggestion, however, comes from Fig. 4: when a straight-line extrapolation is used, it appears that the closing rate would be very small or zero when none of the channels are Ca2+-blocked. If so, then β1 is either zero or very small, and closing occurs only or mainly from the Ca2+-blocked state. The effective closing rate would then be proportional to the fraction of the channels that are Ca2+-blocked, as the graph suggests. This idea was tested by using the state diagram given above with β1 equal to zero. Closing in this hypothetical case is purely from the open gate, Ca2+-blocked state. The results are given by the broken line in Fig. 4. β2 had the fixed value of 7 ms−1, and the predicted rate constant of closing is
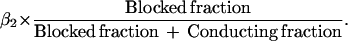 |
The closing rate increases linearly with the blocked fraction, giving a reasonable fit to the experimental points.
In conclusion, the data presented show with near certainty that Na+ channels can close when they are Ca2+-occupied. They suggest but do not prove that Na+ channels close preferentially or exclusively when Ca2+-occupied. This suggestion is strongly supported by the data of the next paper (9). They further suggest, but do not prove, that the closed channel remains Ca2+-occupied, and that Ca2+ is released from the channel at a late stage in the activation chain.
Acknowledgments
We are grateful to Drs. Steve Baylor, Brian Salzberg, Zhe Lu, and W. F. Gilly for reading and commenting on the manuscript. This work was supported by U.S. Public Health Service Grant NS12547.
Footnotes
A Commentary on this article begins on page 3331.
References
- 1.Schoppa N E, Sigworth F J. J Gen Physiol. 1998;111:312–342. doi: 10.1085/jgp.111.2.295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zagotta W N, Hoshi T, Aldrich R W. J Gen Physiol. 1994;103:321–362. doi: 10.1085/jgp.103.2.321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Yang N, George A L, Horn R. Neuron. 1996;16:113–122. doi: 10.1016/s0896-6273(00)80028-8. [DOI] [PubMed] [Google Scholar]
- 4.Larsson H P, Baker O S, Dhillon D S, Isacoff E Y. Neuron. 1996;16:387–397. doi: 10.1016/s0896-6273(00)80056-2. [DOI] [PubMed] [Google Scholar]
- 5.Hille B. Ionic Channels of Excitable Membranes. Sunderland, MA: Sinauer; 1992. [Google Scholar]
- 6.Frankenhaeuser B, Hodgkin A L. J Physiol (London) 1957;137:218–244. doi: 10.1113/jphysiol.1957.sp005808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Baker P F, Hodgkin A L, Ridgeway E B. J Physiol (London) 1971;218:709–755. doi: 10.1113/jphysiol.1971.sp009641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Woodhull A M. J Gen Physiol. 1973;61:687–708. doi: 10.1085/jgp.61.6.687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Armstrong C M. Proc Natl Acad Sci USA. 1999;96:4158–4163. doi: 10.1073/pnas.96.7.4158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cota G, Armstrong C M. J Gen Physiol. 1989;94:213–232. doi: 10.1085/jgp.94.2.213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Armstrong C M, Cota G. Proc Natl Acad Sci USA. 1991;88:6528–6531. doi: 10.1073/pnas.88.15.6528. [DOI] [PMC free article] [PubMed] [Google Scholar]