

Abstract
Bacteria that cause cytoplasmic incompatibility (CI) are perhaps the most widespread parasites of arthropods. CI symbionts cause reproductive failure when infected males mate with females that are either uninfected or infected with a different, incompatible strain. Until recently, CI was known to be caused only by the α-proteobacterium Wolbachia. Here we present the first study of the population biology of Cardinium, a recently discovered symbiont in the Bacteroidetes that causes CI in the parasitic wasp Encarsia pergandiella (Hymenoptera: Aphelinidae). Cardinium occurs at high frequency (∼92%) in the field. Using wasps that were recently collected in the field, we measured parameters that are crucial for understanding how CI spreads and is maintained in its host. CI Cardinium exhibits near-perfect rates of maternal transmission, causes a strong reduction in viable offspring in incompatible crosses, and induces a high fecundity cost, with infected females producing 18% fewer offspring in the first 4 days of reproduction. We found no evidence for paternal transmission or horizontal transmission of CI Cardinium through parasitism of an infected conspecific. No evidence for cryptic parthenogenesis in infected females was found, nor was sex allocation influenced by infection. We incorporated our laboratory estimates into a model of CI dynamics. The model predicts a high stable equilibrium, similar to what we observed in the field. Interestingly, our model also predicts a high threshold frequency of CI invasion (20% for males and 24% for females), below which the infection is expected to be lost. We consider how this threshold may be overcome, focusing in particular on the sensitivity of CI models to fecundity costs. Overall our results suggest that the factors governing the dynamics of CI Wolbachia and Cardinium are strikingly similar.
MICROBES that cause reproductive incompatibility in their hosts, commonly known as cytoplasmic incompatibility (CI) symbionts, are among the most prevalent parasites of arthropods and have thus far been reported in seven orders, including beetles, mosquitoes and other flies, wasps, planthoppers, moths, mites, and isopods (Breeuwer 1997; Hoffmann and Turelli 1997; Moret et al. 2001; Hunter et al. 2003; Gotoh et al. 2007). Cytoplasmic incompatibility symbionts are generally strictly maternally inherited (as are mitochondria). At its simplest, CI occurs when infected male hosts mate with uninfected females, producing few or no viable offspring. Infected females are at a reproductive advantage relative to uninfected females because they produce viable offspring when mated to both infected and uninfected males. CI symbionts thus decrease the fitness of uninfected females instead of increasing that of infected ones (Werren and O'Neill 1997). While the mechanism is not known, the CI phenotype fits a “modification-rescue” model (Werren 1997), in which sperm from infected males are modified such that the proper union of paternal and maternal chromosomes can occur only when both parents harbor compatible CI strains. In incompatible eggs, the embryo generally dies before hatching.
CI infections can spread rapidly, with infected individuals quickly replacing uninfected ones. In one dramatic example of the transformational power and speed of a CI invasion, Turelli and Hoffmann (1991) captured the spread of Wolbachia-infected Drosophila simulans across California in just a few years time. The spread of CI infections are of interest because they may cause significant evolutionary change in host populations, including reductions in mitochondrial diversity, and life history changes associated with infection (e.g., a lowered fecundity or shortened life span) (Hoffmann and Turelli 1997). Understanding CI symbiont spread is also important for their application in controlling medically and economically important pest species (Sinkins et al. 1997; Xi et al. 2005). Potential control approaches include using CI to drive a beneficial gene into a host population, such as a gene that makes a mosquito refractory to malaria transmission, or using a CI strain that causes early death (and reduces the parasite transmission potential) of its host. Mass release of incompatible males has also been proposed as a means of population suppression or eradication (Laven 1967; Zabalou et al. 2004).
Assuming strict maternal transmission, theory predicts that the main factors determining whether a CI symbiont will spread within a population are the (1) fitness cost of the infection, (2) severity or strength of incompatibility (i.e., relative progeny production of incompatible vs. compatible crosses), and (3) fidelity of vertical transmission (Caspari and Watson 1959; Hoffmann et al. 1990; Turelli 1994; Turelli and Hoffmann 1995; Rasgon and Scott 2003). With perfect maternal transmission and no fitness costs to the infection, CI infections will sweep through the population and become fixed. In the more realistic case of imperfect maternal transmission and/or fitness costs to the infection, there are three possible equilibria; the first is zero (i.e., loss of infection), and the second is a stable equilibrium, with infections often close to fixation. The third is an unstable intermediate equilibrium; this is the “threshold frequency” below which the infection will be lost and above which it will reach the stable polymorphism. The threshold frequency is, at its simplest, determined by the ratio of the selection against infected individuals due to fecundity costs and selection against uninfected individuals due to CI-induced reduced egg hatch (Hoffmann and Turelli 1997). Given that there are often fitness costs to having the infection, it is important and sometimes difficult to understand how this threshold is overcome and how infections reach high frequencies in nature.
All of the experimental and theoretical work on the population biology of CI has been done on the α-proteobacterium Wolbachia, which was previously thought to be the only causal agent of CI. However, an unrelated bacterial symbiont, Candidatus Cardinium (henceforth called Cardinium), was recently shown to cause CI in the parasitoid wasp Encarsia pergandiella (Hymenoptera: Aphelinidae) (Hunter et al. 2003; Zchori-Fein et al. 2004) and in the spider mite Eotetranychus suginamensis (Gotoh et al. 2007). Cardinium is in the Bacteroidetes group and thus in a different bacterial phylum than Wolbachia. Like Wolbachia, Cardinium is widely distributed in arthropods (Weeks et al. 2003; Zchori-Fein and Perlman 2004) and is associated with a number of phenotypes involved in the manipulation of host reproduction, including host feminization and induction of parthenogenesis (Weeks et al. 2001; Zchori-Fein et al. 2001). The discovery of a novel lineage of symbionts that causes CI offers the possibility of comparative analysis of the mechanisms and consequences of reproductive manipulation.
In this article, we report the first study of the population biology and ecology of CI Cardinium. Our goal is to understand whether the factors governing the maintenance and spread of CI differ between the two unrelated symbionts, Cardinium and Wolbachia. In addition to determining Cardinium infection frequency in E. pergandiella in the wild, we measure factors that are crucial in CI symbiont maintenance and spread, including effects on host fitness and sex allocation and efficiency of vertical transmission. We test whether Cardinium can be transmitted via paternal transmission or horizontally through parasitism of an infected conspecific (see materials and methods) and whether there are low levels of parthenogenesis associated with infection. We then incorporate these factors into a model of CI Cardinium dynamics to predict the invasion threshold and stable equilibrium frequencies.
MATERIALS AND METHODS
E. pergandiella and autoparasitism:
E. pergandiella wasps are minute (∼1 mm) parasitoids of whiteflies. Adult females lay eggs singly at immature stages of their hosts, and their progeny develop, eventually killing and consuming their host entirely before pupation and emergence. In addition, E. pergandiella, like almost all Encarsia species, has an unusual life history that is characterized by obligate sex-specific differences in host use (Hunter and Woolley 2001). Whereas female E. pergandiella eggs are laid and develop in whitefly immatures, male E. pergandiella eggs are laid and develop in late larval–pupal stages of other parasitoid wasps developing within the whitefly. Hosts for male E. pergandiella eggs include female conspecifics. This unusual life history is termed autoparasitism (Hunter and Woolley 2001). Sex-specific host use is enabled by haplodiploid sex determination in wasps, where females determine the sex of their offspring by selective fertilization: males develop from unfertilized (haploid) eggs, and females from fertilized (diploid) eggs. E. pergandiella females thus lay female eggs when they encounter whitefly hosts and male eggs when they encounter developing wasps.
E. pergandiella attacks many species of whitefly and has been used in the biological control of the sweet potato whitefly Bemisia tabaci (Hemiptera: Aleyrodidae), a worldwide pest of crops (Goolsby et al. 1998). Male E. pergandiella develop as parasitoids of many species of wasps. E. pergandiella consists of a number of well-characterized reproductively isolated lineages that differ in reproductive mode—sexual vs. strictly parthenogenetic reproduction—and in the orientation of the pupa within the whitefly (Liu and Stansly 1996). One lineage of E. pergandiella, collected in Brazil, is strictly parthenogenetic and infected with a strain of Cardinium (Zchori-Fein et al. 2001). Our study concerns the sexual lineage that pupates with the ventral side facing up and attacks B. tabaci in the Rio Grande Valley of Texas. This lineage is infected with a different strain of Cardinium that causes CI (Hunter et al. 2003). In incompatible crosses, wasps lay (fertilized) eggs in whiteflies, but these eggs do not hatch; this is the predominant type of CI found in haplodiploid species and is termed the “female mortality” type (Breeuwer 1997; Vavre et al. 2000, 2003). In the second, less common type of CI in haplodiploids, termed the “male development” type, incompatible offspring develop as males after the paternal set of chromosomes is completely destroyed, leaving a haploid (i.e., male) embryo (Breeuwer and Werren 1990).
Infection frequency, PCR, and wasp culturing:
To determine the frequency of Cardinium infection in the field, we collected wasp pupae in the lower Rio Grande Valley in Texas in October 2003. We collected wasps at two sites separated by ∼30 miles in agricultural fields (lettuce and melon) in the vicinity of Weslaco, Texas, in October 2003. At this time, E. pergandiella was uncommon relative to native and exotic parasitoids in the genus Eretmocerus. A trip to resample infection frequencies in autumn 2006 was not successful, as E. pergandiella had become extremely rare relative to two exotic species, Eretmocerus mundus and Encarsia sophia, both of which were introduced to control the exotic biotype “B” of B. tabaci.
Pupae from the 2003 collection were returned to the lab, and after emergence, adults were killed and PCR was used to assess infection. The DNA extraction protocol is described in Hunter et al. (2003). We used the Cardinium-specific 16S rDNA diagnostic primers CLOf and CLOr (Weeks et al. 2003). Our reaction mixture consisted of 0.8 μl 10 mm dNTPs, 0.5 μl 25 mm MgCl2, 0.5 μl 5 pmol primers, 0.25 units Taq, 2 μl 10× buffer, and 2 μl template in a 10-μl reaction. PCR reaction conditions consisted of 40 cycles of 60 sec at 95°, 60 sec at 50°, and 90 sec at 72°.
Initially, PCR using insect mitochondrial primers (mtd10 and mtd12; Kenyon and Hunter 2007) was used as a positive control for our DNA extractions. However, we also developed a single-copy nuclear gene PCR assay as a more conservative test of the sensitivity of our DNA extraction method. We therefore designed primers to amplify an ∼400-bp fragment of EF1-α (f1 copy) in E. pergandiella. EF1-α occurs as a duplication in many Hymenoptera (Danforth and Ji 1998). We first used the degenerate primers, EF1-For3 and Cho10 (Danforth and Ji 1998), and produced two bands. Bands were cloned and sequenced, and primers amplifying a portion of only the f1 copy were designed (EpergEF-F CGTCAAGGAGCTCCGTCGCGGTTACGTCACCGGCG; EpergEF-R CGCATGTCGCGCACGGCGAATCTACCGAGAGGCGG). The sequence corresponding to this amplified product has been deposited in GenBank (accession no. EU186066). Reaction conditions were as with the Cardinium-specific primers. Both the mitochondrial and Ef1-α primers consistently amplified products from our single-wasp DNA extractions. Finally, uninfected wasps (from a lab culture) and no-DNA negative controls were run as well, to guard against contamination (false positives).
We also established a laboratory culture from infected wasps collected on this trip to be used in fitness, sex allocation, and horizontal transmission experiments. This culture appeared fixed for Cardinium infection. We also reconfirmed the absence of Wolbachia in E. pergandiella by PCR, using Wolbachia-specific primers (Hunter et al. 2003). Rearing conditions for the culture were as described in Hunter et al. (2003). Cured laboratory cultures were derived from infected cultures, following three generations of antibiotic treatment, and at least one additional generation to remove potential maternal effects. Cured and infected cultures therefore had similar genetic backgrounds. Wasps were cured of their infection by feeding adults 50 mg/ml rifampicin in honey solution for 48 hr. We used PCR from single-wasp extractions to monitor infection status.
Maternal transmission efficiency:
Our laboratory cultures appeared fixed for CI Cardinium, suggesting that transmission in the laboratory is close to perfect. To assess vertical transmission efficiency in conditions that might more accurately reflect those in the field, we assessed the infection status of the offspring of 20 females that were collected in the field as pupae. Upon emergence as adults, these females were mated to uninfected males from our laboratory culture. Adult females were then allowed to oviposit in arenas consisting of leaf disks bearing whitefly nymphs, all set on agar in a 35-mm petri dish (Hunter et al. 2003). The female offspring were reared until adulthood, and both mothers and offspring were tested for infection using PCR.
Testing for paternal transmission:
To test for paternal transmission of Cardinium, infected males were mated with uninfected females and set on two whitefly-infested plants. All emerging daughters were collected (i.e., the few offspring resulting from an incompatible cross) and tested for Cardinium infection via PCR.
Testing for horizontal transmission via autoparasitism:
Horizontal transmission of symbionts among immature parasitoids developing within the same insect host has been previously demonstrated, including transfer of Wolbachia among conspecific Trichogramma in moth eggs (Werren et al. 1986; Huigens et al. 2000). Rates of horizontal transmission have not been reported for Cardinium. We tested whether Encarsia males can pick up Cardinium from infected female conspecifics when developing on them. We expected that males that became infected in this way would not transmit the symbiont (i.e., no paternal transmission), but frequent horizontal transfer from infected females to males could raise the frequency of infected males and the frequency of CI and assist spread of a CI symbiont. Our experimental protocol ensured that all emerging males were the sons of uninfected females but had developed on infected females. First, we set Cardinium-infected female wasps on whitefly hosts. All adult females were removed 3 days later by exposing plants to a dichlorvos-impregnated pesticide strip for 2 hr (Hot Shot No Pest Strip, St. Louis). Fumigation kills adult wasps but does not affect whiteflies or immature parasitoids. Six days later, when developing wasps were young larvae, we removed all leaves from plants and set leaf disks in agar in petri dishes. Nine days later, we set uninfected virgin females on these leaf disks with female pupae for 24 hr, so that they would lay (uninfected) male eggs. Leaves were examined daily and emerging males were isolated and scored for infection via PCR.
Cytoplasmic incompatibility:
We confirmed the presence and expression of CI Cardinium in the wasps from the 2003 field collections, repeating the CI experiments described in Hunter et al. (2003). The experiment was conducted on individuals that were nine generations removed from the field. We conducted single-pair crosses in all four possible combinations (infected/cured, male/female) and set mated females for 6 hr on leaf-disk arenas, as described above. In this experiment, each leaf disk contained 30 late third or early fourth instar whitefly hosts and each treatment was replicated 12 times. In the previous study, we found that the offspring of an incompatible cross die in the egg stage and cause developmental arrest of the whitefly host (Hunter et al. 2003). Arrested whiteflies can remain alive for days to weeks and never complete development (M. S. Hunter, S. J. Perlman and S. E. Kelly, unpublished results). We recorded all emerged whiteflies (i.e., unparasitized), arrested whiteflies, and developing wasp pupae and used logistic regression to compare the proportion of developing vs. dead wasps (i.e., arrested whiteflies) between the four different crosses. Due to the obligate sex-specific host use of E. pergandiella, only female (i.e., fertilized) eggs are laid in whitefly hosts.
Cryptic parthenogenesis:
As mentioned previously, a parthenogenetic population of E. pergandiella is infected with a closely related strain of Cardinium. We were interested in determining whether the CI-Cardinium strain might also occasionally cause parthenogenesis, as this could potentially affect symbiont dynamics. We placed 26 infected virgin females (isolated as pupae from field collections) individually in leaf-disk arenas bearing whitefly nymphs for at least 12 hr. To minimize the possibility that the experimental females were siblings, we did not include more than one female collected from the same plant leaf. Adult females were removed, and the arenas incubated and later examined for progeny production. Because sex-specific host use is strictly obligate, virgin sexual females do not ordinarily lay eggs in whiteflies, nor are male eggs able to develop in these hosts. Thus, sexual females would not be expected to produce any offspring, while parthenogens would be expected to produce female progeny.
Effect of CI Cardinium on sex allocation:
The spread of a CI symbiont can be enhanced by a greater female bias in the offspring of infected females relative to uninfected individuals (Egas et al. 2002). We tested whether CI Cardinium causes a female sex-ratio bias by inducing infected wasps to lay more eggs in whiteflies (female offspring) than in wasps (male offspring). This possibility was suggested to us by evidence that host preference in the parthenogenetic E. pergandiella is influenced by infection status (Kenyon and Hunter 2007). Arenas consisted of 35-mm petri dishes containing a grid of alternating whitefly and wasp hosts on leaf disks on moistened filter paper, with 16 hosts of each type. In this experiment, wasp hosts consisted of Eretmocerus eremicus (Hymenoptera: Aphelinidae) pupae, which are one of many possible hosts for E. pergandiella males in nature (Gerling 1966). Whiteflies were late third–early fourth instar nymphs. Experimental E. pergandiella females were mass mated to males of similar infection status and placed individually in experimental arenas for 5 hr. Hosts were incubated at 25° for 24 hr, after which they were refrigerated until they could be dissected and the number of eggs within each host determined. Any females that did not oviposit in whitefly hosts were removed from the analysis, as it was possible that these were unmated and would not lay female eggs. We compared the proportion of female eggs laid by infected vs. uninfected wasps using a t-test, following arcsine-square-root transformation.
Effect of CI-Cardinium infection on host female fecundity:
One- to 2-day-old cured and infected females were paired for 24 hr in small vials with a male of the same infection status. Females were then placed individually on whitefly-infested leaf disks. We recorded any developing wasp larvae and compared fecundity at 2, 4, 6, and 8 days of adult age, after which most of the wasps had died. This experiment was performed in two blocks. In the first, wasps were transferred to new whitefly-infested leaves every 2 days, and in the second, they were transferred daily. We used repeated-measures ANOVA to compare fecundity of infected and uninfected females. Only female eggs were laid in this experiment, as only whitefly hosts were available for oviposition.
RESULTS
Infection frequency:
Of 52 field-collected E. pergandiella, 48 (92.3%; 95% binomial confidence interval of 81.4–97.9%) were infected with Cardinium. If we include only one wasp per plant leaf in our survey to minimize the possibility that individuals collected on the same leaf may have been siblings, 27 of 29 (93.1%; 95% binomial confidence interval of 77.2–99.2%) wasps were infected. One leaf yielded a mixture of infected and uninfected individuals.
Maternal transmission efficiency:
All but 2 of the 510 offspring that we tested (from a total of 20 different field-collected females) were infected (99.6%; 95% binomial confidence interval of 98.6–100%). These uninfected females were from two different mothers.
Paternal and horizontal transmission:
We did not find any evidence for paternal transmission (0/86 individuals tested were infected). Nor did we find evidence for horizontal transmission via autoparasitism. None of the 207 males that developed on infected females was infected.
Cytoplasmic incompatibility:
Matings from the predicted incompatible cross (uninfected females with infected males) resulted in fewer offspring (mean number of wasp pupae produced: 1.54 ± 0.65 SE, n = 11; Figure 1A) and more arrested whiteflies (16.18 ± 0.92; Figure 1B), compared to the other three crosses (pupae: 21.11 ± 0.63, n = 35; arrested whiteflies: 0.43 ± 0.08). This difference was highly significant (effect of the incompatible cross: 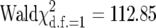 , P < 0.0001).
, P < 0.0001).
Figure 1.—

(A) Mean number of viable E. pergandiella pupae (±SE) produced in single-pair crosses of infected (I) and uninfected (U) individuals, with males listed first in the cross designations. Numbers above bars indicate sample sizes. (B) Mean number of developmentally arrested whiteflies, corresponding to whiteflies that were parasitized but wasp eggs did not hatch. Developmentally arrested whiteflies occurred at significantly higher frequencies in the predicted incompatible cross. Matings from the predicted incompatible cross (uninfected females with infected males) resulted in a significantly higher proportion of inviable offspring (arrested whiteflies) (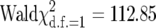 , P < 0.0001).
, P < 0.0001).
Cryptic parthenogenesis:
None of the virgin females produced offspring on whitefly hosts (0/26). Although we cannot rule out the possibility that parthenogenesis occurs at very low frequencies (e.g., Lawson et al. 2001), parthenogenesis is not likely an ecologically important force in the maintenance of CI Cardinium.
Sex allocation of cured vs. infected wasps:
Cured females laid slightly, but not significantly, more eggs in total (20.06 ± 1.64 vs. 16.82 ± 2.19) than infected females (td.f.=33 = 1.19, P = 0.24). There was also no significant difference in the rate of parasitism of whiteflies relative to wasps, with whiteflies composing 59% of the hosts parasitized by cured females and 53% of the hosts parasitized by infected females (td.f.=33 = 0.44, P = 0.66 for arcsine-transformed data). These data suggest a relatively even allocation of males and females by both infected and uninfected females.
Effect of CI-Cardinium infection on host female fecundity:
There was no significant effect of block on fecundity so both blocks were pooled into a single data set. Uninfected females produced significantly more offspring than infected females (F1,34 = 7.39, P = 0.0101, Figure 2). After 4 days, cured females had produced 18% more offspring than infected females (92.62 ± 4.98 vs. 78.52 ± 4.74) and this difference increased to 35% after 8 days of reproduction (143.56 ± 10.27 vs. 106.46 ± 9.07).
Figure 2.—

Mean number of viable E. pergandiella pupae (±SE) produced by infected and uninfected females after 2, 4, 6, and 8 days, with unlimited whitefly hosts. Numbers above and below means indicate sample sizes. Uninfected females had significantly greater fecundity, producing 18 and 35% more offspring after 4 and 8 days, respectively.
DISCUSSION
Here we report the first population biology study of a cytoplasmic incompatibility microbe other than Wolbachia. We show that CI Cardinium infecting E. pergandiella displays a syndrome of traits that is strikingly similar to strains of Wolbachia that cause strong CI, including perfect or near-perfect vertical transmission, extremely low or absent horizontal transmission, and high prevalence. The CI described here is strong, with incompatible crosses producing only 7% of the number of female offspring as compatible ones.
Much variation in Wolbachia CI strength has been documented, ranging from nearly complete to weak incompatibility (Hoffmann and Turelli 1988; Hoffmann et al. 1998). It is not clear, however, how levels of incompatibility may differ in the field vs. the laboratory. The few studies that have measured CI in near-field conditions have found that CI expression tends to be weaker in the field (Hoffmann et al. 1990, 1998; Turelli and Hoffmann 1995). CI may be weaker in the field due to factors that decrease bacterial densities and/or CI expression, including naturally occurring antibiotics (Stevens and Wicklow 1992), temperature variation (Hoffmann and Turelli 1988), and stress (Sinkins et al. 1995). Also, older males have been shown to express weaker CI due to lower Wolbachia titers in some (e.g., Turelli and Hoffmann 1995; Jamnongluk et al. 2000), but not all (e.g., Rasgon and Scott 2003) host species, although the effect of male age on CI strength has not yet been tested for Cardinium in Encarsia.
We found very efficient vertical transmission, as virtually all offspring of field-collected infected females that we tested were infected. The fact that we collected a few uninfected individuals in the field is best explained by the occurrence of some incomplete vertical transmission (Hoffmann and Turelli 1997). A small number of studies have measured vertical transmission of CI Wolbachia in the field and have shown that it is often, but not always, weaker than in the laboratory (Subbarao et al. 1977; Turelli and Hoffmann 1995; Hoffmann et al. 1996), again possibly due to factors affecting density and/or expression.
We found no evidence for paternal transmission of CI Cardinium or for horizontal transmission via autoparasitism. In this way, too, Cardinium appears to be similar to Wolbachia. Horizontal transmission of Wolbachia over ecological timescales is extremely rare. Low rates of paternal transmission (∼1–2%) of CI Wolbachia have been reported in D. simulans and Leptopilina heterotoma in the laboratory (Hoffmann et al. 1990; Vavre et al. 2000), but in most experimental systems paternal transmission appears to be rare or absent. Transmission of bacterial symbionts between parasitic wasps via a shared host has been reported in a strain of Wolbachia causing parthenogenesis in Trichogramma wasps (Huigens et al. 2000). While both Wolbachia and Cardinium appear to be primarily transmitted vertically over ecological timescales, phylogenetic studies clearly demonstrate frequent horizontal transmission over evolutionary timescales (Werren et al. 1995; Zchori-Fein and Perlman 2004). Initial surveys suggest that Cardinium may show greater host fidelity and therefore presumably lower rates of horizontal transfer than Wolbachia, as it has been found in only three arthropod taxa to date: Hymenoptera, Hemiptera, and Acari (Weeks et al. 2003; Zchori-Fein and Perlman 2004). Also, although the Cardinium phylogeny suggests horizontal transfer within host groups, the clade of Cardinium that infects Hymenoptera is distinct from the one infecting Acari (Zchori-Fein and Perlman 2004).
We found a significant effect of Cardinium infection on female fecundity, with cured females producing 18% more offspring after just 4 days and 35% more offspring after 8 days. The 18% value is likely a more realistic estimate of fitness differences, since minute parasitoids are unlikely to live as long in the field as they do in the laboratory (Mansfield and Mills 2002), and theory suggests that, for organisms such as whitefly parasitoids with overlapping generations, offspring that are produced late have relatively little reproductive value (Stearns 1992). Uninfected individuals were recently descended from curing multiple infected individuals, suggesting that the fitness cost is unlikely to be due to differing host genetic backgrounds. Other studies have reported variable estimates of the fecundity cost of infection with CI Wolbachia, including lower (Hoffmann et al. 1990), higher (Dobson et al. 2002; Weeks et al. 2007), and no differences in fecundity (Bordenstein and Werren 2000).
Since CI-Cardinium infections are not horizontally transmitted, do not induce parthenogenesis, or affect host sex-allocation effects, we can use a version of the standard Turelli–Hoffmann models of CI maintenance and spread (Turelli 1994; Turelli and Hoffmann 1995; Hoffmann and Turelli 1997), modified for haplodiploid hosts with the “female mortality” type of CI by Vavre et al. (2000), as a framework for understanding CI invasion and maintenance. The basic model for the frequency of infected males (m) and females (f) at time t + 1 is
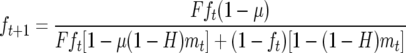 |
and
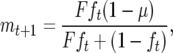 |
where F is the fecundity of infected females relative to uninfected, H is the proportion of eggs that escape CI in the incompatible cross, and μ is the proportion of uninfected eggs produced by infected females. To generate a set of predictions for both stable and unstable (threshold) equilibria for males and females, we used a set of core parameters determined by our experiments with E. pergandiella as “best estimates” and then varied each of the three parameters in turn across a range of values of interest. Equilibria were calculated using the appendix formulae in Vavre et al. (2000). We initially set F as 0.82, the estimate of the fecundity cost after 4 days of reproduction, and varied F between 0.65 (fecundity cost after 8 days of reproduction) and one (if fitness differences are not realized in the field: see discussion below). We initially set H as 0.07, the proportion of eggs that escaped CI, and varied this parameter between 0.07 and 0.55. The latter represents weak CI and allows the examination of the effect of dramatically reduced CI on equilibria predictions. Finally, we set μ at 0.01, a more conservative estimate than the nearly perfect maternal transmission indicated by our experiments and near the lower limit of our 95% confidence interval, and varied μ between 0.01 and 0.18. We chose a range that would allow the investigation of the effects of potentially much lower field rates of vertical transmission on the stable equilibria.
With the core parameters, the model predicted a stable equilibrium for males and females of 0.99 (Figure 3, b, d, and f). This is higher than our estimate of the field infection frequency of females (92%), but close to the upper bounds of the confidence interval of 81–98% [and within the confidence interval (77.2–99.2%) of our more conservative estimate of infection frequency based on sampling one wasp/leaf]. In general, both data and model predictions conform to the common pattern of strong CI infections found at high frequencies (Hoffmann et al. 1990; Rasgon and Scott 2003; but see Tagami et al. 2006 for an exception). The discrepancy between the observed infection frequency and model predictions may be explained by a combination of sampling error and lower maternal transmission in the field. For example, transmission efficiency <86% (μ = 0.15) yields a stable equilibrium of 92% (Figure 3b). It is also possible that the infection was not at equibilibrium when sampled. This does not seem likely, however, since the model predicts that only two generations would be required for the infection frequency to increase from 92% to the stable equilibrium of 99%. Clearly, if E. pergandiella was still abundant, more field samples would be valuable for testing the robustness of the observed infection frequency. In contrast to the potentially strong influence of μ, stable equilibria appear to be relatively insensitive to changes in H and F, so it is less likely that changes in these estimates would change the predicted value (Figure 3, d and f).
Figure 3.—

Predictions of the unstable equilibria (i.e., the threshold frequency: a, c, e) and stable equilibria (b, d, and f), of CI-Cardinium infection rates in E. pergandiella. The equilibria differ slightly for females (solid line), and males (dashed line). Equilibria were calculated using the model of Vavre et al. (2000) for a haplodiploid insect with the “female mortality” type of CI. The parameters varied were μ (the proportion of eggs laid by infected females that are not infected), H (the proportion of female offspring produced in the incompatible cross relative to the normal crosses), and 1-F (the proportional reduction in fecundity of infected females relative to uninfected females). The core set of parameters was estimated from experiments and is indicated by asterisks: μ (0.01), H (0.07), and 1-F (0.18). The rationale for the range of values varied for each factor is explained in the text.
One of the most interesting predictions of the model is a high unstable equilibrium of 0.24 for females and 0.20 for males (Figure 3, a, c, and e). This high threshold is predicted in part because the host is haplodiploid. In haplodiploids in which CI causes female mortality, male production by uninfected females is not affected by incompatible matings (since males develop from unfertilized eggs) (Vavre et al. 2000). The threshold is predicted to be even higher in the “male development” type of CI in haplodiploids. Lower vertical transmission rates and higher fecundity costs all serve to increase the unstable equilibrium (Figure 3, a, c, and e). Within the ranges of parameters that we examined, however, only lower fecundity costs—below the levels that we observed in the laboratory—could lower the predicted unstable equilibrium (Figure 3e).
The high predicted threshold in this study system begs explanation: one would not predict that a CI infection could invade under these circumstances, yet CI Cardinium has clearly invaded. Indeed, there are many examples of CI symbionts at high frequency in haplodiploid wasps and mites, in spite of the expectation that these symbionts should have difficulty invading these hosts (Breeuwer and Werren 1990; Breeuwer 1997; Vavre et al. 2002; Gotoh et al. 2007). The ways in which CI infections overcome predicted invasion thresholds remain largely unknown, and a number of possible mechanisms have been invoked, including genetic drift, population structure, conditional fitness benefits, low rates of horizontal transmission or parthenogenesis, and avoidance of matings with incompatible males (Hoffmann and Turelli 1997; Egas et al. 2002).
For E. pergandiella, we can rule out the role of horizontal transmission, parthenogenesis, and sex-allocation effects in facilitating invasion of CI Cardinium. The role of population structure of E. pergandiella is difficult to evaluate, as the current environment may not be the one in which the infection invaded. Whiteflies and whitefly parasitoids in general, however, do not show highly structured populations (Hunter 1993). We suggest that a more important factor may be that insect parasitoids often experience very strong resource limitation in the field due to the relative scarcity of hosts and that the realized fecundity cost of infection may be low. In our study, uninfected females parasitized on average 93 hosts in the first 4 days of life, 14 more than infected females did, but this cost is unlikely to be relevant if a female in the field encounters only a small proportion of that number of hosts. Fecundity costs have been shown to be more pronounced in the laboratory than in the field (Turelli and Hoffmann 1995). While the life-time survival and fecundity of E. pergandiella in the field are not known, E. pergandiella sex ratios in the field suggest that they are more host limited than egg limited (Hunter 1993; West and Rivero 2000). While host limitation in the field may be one factor permitting invasion of CI Cardinium, clearly many other possibilities exist, such as an as-yet-unknown fitness benefit to its hosts. Finally, conditions may have been different at the time of initial invasion. As a striking example, a recent study documented rapid evolution of fecundity effects of CI Wolbachia in natural populations of D. simulans (Weeks et al. 2007). Thus it would be very informative to assess infection frequencies and mitochondrial sequence diversity in other E. pergandiella populations to estimate the age of the CI-Cardinium infection in this insect.
The laboratory estimates of CI-Cardinium parameters clearly indicate the overall ecological similarity of CI Cardinium to CI Wolbachia and are an important first step toward understanding CI-Cardinium dynamics. However, population-level testing of model predictions is a critical future step. A recent elegant study of CI-Wolbachia invasion in an artificially transfected line of the mosquito Aedes aegypti illustrates the importance of reconciling CI theory with population processes (Xi et al. 2005). This study used population cages with varying frequencies of infected mosquitoes to show that the threshold for invasion was ∼20%, a level that theory predicts would correspond with a 15% fecundity cost. Yet comparisons of infected and uninfected mosquitoes in the laboratory did not show any difference in fecundity, suggesting that subtle but as-yet-elusive fitness differences may be important (Xi et al. 2005). Clearly, the understanding of CI spread in nature, as well as the application of CI symbionts for management of insect pests and disease vectors, depends on a detailed grasp of both individual and population processes.
Acknowledgments
We thank John Goolsby, T.-X. Liu, and Bill Warfield for invaluable help in the field; Cara Gibson, Leanne Harris, Sarah Kenyon, and Laurie McCormick for technical assistance; and Larry Harshman, Kerry Oliver, Fabrice Vavre, Jen White, and two anonymous reviewers for helpful comments. This project was supported in part by the National Research Initiative of the U. S. Department of Agriculture Cooperative State Research Education and Extension Service (grant no. 2001-35302-10986 to M.S.H.) and by the National Science Foundation (grant no. DEB-0542961 to M.S.H. and S.J.P.).
References
- Bordenstein, S. R., and J. H. Werren, 2000. Do Wolbachia influence fecundity in Nasonia vitripennis? Heredity 84 54–62. [DOI] [PubMed] [Google Scholar]
- Breeuwer, J. A. J., 1997. Wolbachia and cytoplasmic incompatibility in the spider mites Tetranychus urticae and T. turkestani. Heredity 79 41–47. [Google Scholar]
- Breeuwer, J. A. J., and J. H. Werren, 1990. Microorganisms associated with chromosome destruction and reproductive isolation between two insect species. Nature 346 558–560. [DOI] [PubMed] [Google Scholar]
- Caspari, E., and G. S. Watson, 1959. On the evolutionary importance of cytoplasmic sterility in mosquitoes. Evolution 13 568–570. [Google Scholar]
- Danforth, B. N., and S. Q. Ji, 1998. Elongation factor-1 alpha occurs as two copies in bees: implications for phylogenetic analysis of EF-1 alpha sequences in insects. Mol. Biol. Evol. 15 225–235. [DOI] [PubMed] [Google Scholar]
- Dobson, S. L., E. J. Marsland and W. Rattanadechaku, 2002. Mutualistic Wolbachia infection in Aedes albopictus: accelerating cytoplasmic drive. Genetics 160 1087–1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egas, M., F. Vala and J. A. J. Breeuwer, 2002. On the evolution of cytoplasmic incompatibility in haplodiploid species. Evolution 56 1101–1109. [DOI] [PubMed] [Google Scholar]
- Gerling, D., 1966. Studies with whitefly parasites of Southern California. I. Encarsia pergandiella Howard (Hymenoptera-Aphelinidae). Can. Entomol. 98 707. [Google Scholar]
- Goolsby, J. A., M. A. Ciomperlik, B. C. Legaspi, J. C. Legaspi and L. E. Wendel, 1998. Laboratory and field evaluation of exotic parasitoids of Bemisia tabaci (Gennadius) (Biotype “B”) (Homoptera: Aleyrodidae) in the lower Rio Grande valley of Texas. Biol. Control 12 127–135. [Google Scholar]
- Gotoh, T., H. Noda and S. Ito, 2007. Cardinium symbionts cause cytoplasmic incompatibility in spider mites. Heredity 98 13–20. [DOI] [PubMed] [Google Scholar]
- Hoffmann, A. A., and M. Turelli, 1988. Unidirectional incompatibility in Drosophila simulans – inheritance, geographic variation and fitness effects. Genetics 119 435–444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffmann, A. A., and M. Turelli, 1997. Cytoplasmic incompatibility in insects, pp. 42–80 in Influential Passengers: Inherited Microorganisms and Invertebrate Reproduction, edited by S. L. O'Neill, A. A. Hoffmann and J. H. Werren. Oxford University Press, Oxford.
- Hoffmann, A. A., M. Turelli and L. G. Harshman, 1990. Factors affecting the distribution of cytoplasmic incompatibility in Drosophila simulans. Genetics 126 933–948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffmann, A. A., D. J. Clancy and J. Duncan, 1996. Naturally-occurring Wolbachia infection in Drosophila simulans that does not cause cytoplasmic incompatibility. Heredity 76 1–8. [DOI] [PubMed] [Google Scholar]
- Hoffmann, A. A., M. Hercus and H. Dagher, 1998. Population dynamics of the Wolbachia infection causing cytoplasmic incompatibility in Drosophila melanogaster. Genetics 148 221–231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huigens, M. E., R. F. Luck, R. H. Klaassen, M. F. Maas, M. J. Timmermanns et al., 2000. Infectious parthenogenesis. Nature 405 178–179. [DOI] [PubMed] [Google Scholar]
- Hunter, M. S., 1993. Sex allocation in a field population of an autoparasitoid. Oecologia 93 421–428. [DOI] [PubMed] [Google Scholar]
- Hunter, M. S., and J. B. Woolley, 2001. Evolution and behavioral ecology of heteronomous aphelinid parasitoids. Annu. Rev. Entomol. 46 251–290. [DOI] [PubMed] [Google Scholar]
- Hunter, M. S., S. J. Perlman and S. E. Kelly, 2003. A bacterial symbiont in the Bacteroidetes induces cytoplasmic incompatibility in the parasitoid wasp Encarsia pergandiella. Proc. Biol. Sci. 270 2185–2190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jamnongluk, W., P. Kittayapong, K. J. Baisley and S. O'Neill, 2000. Wolbachia infection and expression of cytoplasmic incompatibility in Armigeres subalbatus (Diptera: Culicidae). J. Med. Entomol. 37 53–57. [DOI] [PubMed] [Google Scholar]
- Kenyon, S. G., and M. S. Hunter, 2007. Manipulation of oviposition choice of the parasitoid wasp, Encarsia pergandiella, by the endosymbiotic bacterium Cardinium. J. Evol. Biol. 20 707–716. [DOI] [PubMed] [Google Scholar]
- Laven, H, 1967. Eradication of Culex pipiens fatigans through cytoplasmic incompatibility. Nature 216 383–384. [DOI] [PubMed] [Google Scholar]
- Lawson, E. T., T. A. Mousseau, R. Klaper, M. D. Hunter and J. H. Werren, 2001. Rickettsia associated with male-killing in a buprestid beetle. Heredity 86 497–505. [DOI] [PubMed] [Google Scholar]
- Liu, T.-X., and P. A. Stansly, 1996. Pupal orientation and emergence of some aphelinid parasitoids (Hymenoptera: Aphelinidae) of Bemisia argentifolii (Homoptera: Aleyrodidae). Ann. Entomol. Soc. Am. 89 385–390. [Google Scholar]
- Mansfield, S., and N. J. Mills, 2002. Direct estimation of the survival time of commercially produced adult Trichogramma platneri Nagarkatti (Hymenoptera: Trichogrammatidae) under field conditions. Biol. Control 25 41–48. [Google Scholar]
- Moret, Y., P. Juchault and T. Rigaud, 2001. Wolbachia endosymbiont responsible for cytoplasmic incompatibility in a terrestrial crustacean: effects in natural and foreign hosts. Heredity 86 325–332. [DOI] [PubMed] [Google Scholar]
- Rasgon, J. L., and T. W. Scott, 2003. Wolbachia and cytoplasmic incompatibility in the California Culex pipiens mosquito species complex: parameter estimates and infection dynamics in natural populations. Genetics 165 2029–2038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinkins, S. P., H. R. Braig and S. L. O'Neill, 1995. Wolbachia superinfections and the expression of cytoplasmic incompatibility. Proc. Biol. Sci. 261 325–330. [DOI] [PubMed] [Google Scholar]
- Sinkins, S. P., C. F. Curtis and S. L. O'Neill, 1997. The potential application of inherited symbiont systems to pest control, pp. 155–175 in Influential Passengers: Inherited Microorganisms and Invertebrate Reproduction, edited by S. L. O'Neill, A. A. Hoffmann and J. H. Werren. Oxford University Press, Oxford.
- Stearns, S. C, 1992. The Evolution of Life Histories. Oxford University Press, Oxford.
- Stevens, L., and D. T. Wicklow, 1992. Multispecies interactions affect cytoplasmic incompatibility in Tribolium flour beetles. Am. Nat. 140 642–653. [DOI] [PubMed] [Google Scholar]
- Subbarao, S. K., B. S. Krishnamurthy, C. F. Curtis, T. Adak and R. K. Chandrahas, 1977. Segregation of cytoplasmic incompatibility properties in Culex pipiens fatigans. Genetics 87 381–390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tagami, Y., M. Doi, K. Sugiyama, A. Tatara and T. Saito, 2006. Survey of leafminers and their parasitoids to find endosymbionts for improvement of biological control. Biol. Control 38 210–216. [Google Scholar]
- Turelli, M., 1994. Evolution of incompatibility-inducing microbes and their hosts. Evolution 48 1500–1513. [DOI] [PubMed] [Google Scholar]
- Turelli, M., and A. A. Hoffmann, 1991. Rapid spread of an inherited incompatibility factor in California Drosophila. Nature 353 440–442. [DOI] [PubMed] [Google Scholar]
- Turelli, M., and A. A. Hoffmann, 1995. Cytoplasmic incompatibility in Drosophila simulans: dynamics and parameter estimates from natural populations. Genetics 140 1319–1338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vavre, F., F. Fleury, J. Varaldi, P. Fouillet and M. Bouletreau, 2000. Evidence for female mortality in Wolbachia-mediated cytoplasmic incompatibility in haplodiploid insects: epidemiologic and evolutionary consequences. Evolution 54 191–200. [DOI] [PubMed] [Google Scholar]
- Vavre, F., F. Fleury, J. Varaldi, P. Fouillet and M. Bouletreau, 2002. Infection polymorphism and cytoplasmic incompatibility in Hymenoptera-Wolbachia associations. Heredity 88 361–365. [DOI] [PubMed] [Google Scholar]
- Vavre, F., P. Fouillet and F. Fleury, 2003. Between- and within-host species selection on cytoplasmic incompatibility-inducing Wolbachia in haplodiploids. Evolution 57 421–427. [DOI] [PubMed] [Google Scholar]
- Weeks, A. R., F. Marec and J. A. J. Breeuwer, 2001. A mite species that consists entirely of haploid females. Science 292 2479–2482. [DOI] [PubMed] [Google Scholar]
- Weeks, A. R., R. Velten and R. Stouthamer, 2003. Incidence of a new sex-ratio distorting endosymbiotic bacterium among arthropods. Proc. Biol. Sci. 270 1857–1865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weeks, A. R., M. Turelli, W. R. Harcombe, K. T. Reynolds and A. A. Hoffmann, 2007. From parasite to mutualist: rapid evolution of Wolbachia in natural populations of Drosophila. PLoS Biol. 5 997–1005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werren, J. H., 1997. Biology of Wolbachia. Annu. Rev. Entomol. 42 587–609. [DOI] [PubMed] [Google Scholar]
- Werren, J. H., and S. L. O'Neill, 1997. Evolution of heritable symbionts, pp. 1–41 in Influential Passengers: Inherited Microorganisms and Invertebrate Reproduction, edited by S. L. O'Neill, A. A. Hoffmann and J. H. Werren. Oxford University Press, Oxford.
- Werren, J. H., S. W. Skinner and A. M. Huger, 1986. Male-killing bacteria in a parasitic wasp. Science 231 990–992. [DOI] [PubMed] [Google Scholar]
- Werren, J. H., W. Zhang and L. R. Guo, 1995. Evolution and phylogeny of Wolbachia: reproductive parasites of arthropods. Proc. Biol. Sci. 261 55–71. [DOI] [PubMed] [Google Scholar]
- West, S. A., and A. Rivero, 2000. Using sex ratios to estimate what limits reproduction in parasitoids. Ecol. Lett. 3 294–299. [Google Scholar]
- Xi, Z. Y., C. C. H. Khoo and S. L. Dobson, 2005. Wolbachia establishment and invasion in an Aedes aegypti laboratory population. Science 310 326–328. [DOI] [PubMed] [Google Scholar]
- Zabalou, S., M. Riegler, M. Theodorakopoulou, C. Stauffer, C. Savakis et al., 2004. Wolbachia-induced cytoplasmic incompatibility as a means for insect pest population control. Proc. Natl. Acad. Sci. USA 101 15042–15045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zchori-Fein, E., Y. Gottlieb, S. E. Kelly, J. K. Brown, J. M. Wilson et al., 2001. A newly discovered bacterium associated with parthenogenesis and a change in host selection behavior in parasitoid wasps. Proc. Natl. Acad. Sci. USA 98 12555–12560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zchori-Fein, E., and S. J. Perlman, 2004. Distribution of the bacterial symbiont Cardinium in arthropods. Mol. Ecol. 13 2009–2016. [DOI] [PubMed] [Google Scholar]
- Zchori-Fein, E., S. J. Perlman, S. E. Kelly, N. Katzir and M. S. Hunter, 2004. Characterization of a ‘Bacteroidetes’ symbiont in Encarsia wasps (Hymenoptera: Aphelinidase): proposal of ‘Candidatus Cardinium hertigii’. Int. J. Syst. Evol. Microbiol. 54 961–968. [DOI] [PubMed] [Google Scholar]