

Abstract
The external environment influences stem cells, but this process is poorly understood. Our previous work showed that germline stem cells (GSCs) respond to diet via neural insulin-like peptides (DILPs) that act directly on the germ line to upregulate stem cell division and cyst growth under a protein-rich diet in Drosophila. Here, we report that DILPs specifically control the G2 phase of the GSC cell cycle via phosphoinositide-3 kinase (PI3K) and dFOXO, and that a separate diet mediator regulates the G1 phase. Furthermore, GSC tumors, which escape the normal stem cell regulatory microenvironment, or niche, still respond to diet via both mechanisms, indicating that niche signals are not required for GSCs to sense or respond to diet. Our results document the effect of diet and insulin-like signals on the cell cycle of stem cells within an intact organism, and demonstrate that the response to diet requires multiple signals. Moreover, the retained ability of GSC tumors to respond to diet parallels the long known connections between diet, insulin signaling, and cancer risk in humans.
Keywords: diet, insulin pathway, germline stem cell, oogenesis, tumor, proliferation, cell cycle, clonal analysis, Drosophila
INTRODUCTION
Stem cells self-renew and give rise to various differentiated cell types within many adult tissues (Potten and Loeffler, 1990; Weissman, 2000). The maintenance of stem cell properties and the precise regulation of their proliferation are, therefore, crucial to maintain tissue integrity and function. Stem cells reside in a specialized microenvironment, or niche, where they receive local signals, such as bone morphogenetic proteins (BMPs), Hedgehogs, and Wnts that regulate their maintenance and proliferation (Li and Xie, 2005; Spradling et al., 2001). Despite the unquestionable importance of local signals in regulating stem cells, stem cell activity is also influenced by stimuli originating outside of the tissues in which they reside, such as diet, hormones, or physical insults (Drummond-Barbosa, 2005; Narbonne and Roy, 2006b). By sensing and responding to external signals, stem cells can tailor the rate of cell production to the ever-changing demands imposed on living organisms by their environment.
The ease of identification and manipulation of germline stem cells (GSCs) in the Drosophila melanogaster ovary provides an ideal model system for studying GSC behavior in vivo (Wong et al., 2005). Each Drosophila ovary is comprised of ovarioles, or strings of progressively more developed egg chambers (Fig. 1A) (Spradling, 1993). The production of egg chambers is maintained by small populations of stem cells located within the germarium, the anterior-most region of the ovariole (Fig. 1B) (Kirilly and Xie, 2007). The division of a GSC produces a cystoblast that divides four more times to form a 2-, 4-, 8-, and 16-cell cyst (Fig. 1C). One cell becomes the oocyte; the others become nurse cells. Follicle cells derived from somatic stem cells surround the cyst, generating an egg chamber that goes through fourteen developmental stages to form a mature oocyte.
Figure 1. Stem cells support oogenesis throughout adult life in the Drosophila ovary and the insulin pathway is required for their response to diet.

(A) Each Drosophila ovariole contains a string of egg chambers formed in the germarium as 16-cell germline cysts are enveloped by follicle cells. One germline cell becomes the oocyte, while the others become nurse cells. Each egg chamber goes through fourteen developmental stages to form a mature oocyte, with vitellogenesis beginning at stage 8. (B) In the germarium, GSCs occupy a niche composed of somatic cap cells, terminal filament cells, and escort stem cells (not shown). Cystoblasts are GSC daughters that give rise to germline cysts. After the cysts are surrounded by somatic follicle cells, they leave the germarium as newly formed egg chambers. (C) Each cystoblast or cyst within the germarium corresponds to a single GSC division. Thus, the number of germline products from a given GSC serves as a direct measure of their division rate. (D) The insulin pathway, which is required for the ovarian response to diet, has known roles in cell growth and proliferation in Drosophila.
The effect of diet on stem cells and their descendents has been well documented in the Drosophila ovary (Drummond-Barbosa and Spradling, 2001). On a protein-rich diet, germline and somatic stem cells have high division rates, and their progeny also divide and grow fast. On a protein-poor diet, these rates are reduced and vitellogenesis is blocked. The response to diet is rapid and reversible, and it requires insulin signaling. Specifically, insulin-like peptides (DILPs) produced in two clusters of neurosecretory cells in the brain directly regulate GSC division, cyst growth and vitellogenesis, while follicle cells receive a secondary signal from the germ line (LaFever and Drummond-Barbosa, 2005). Several questions regarding the direct role of DILPs in mediating the effects of diet on GSC proliferation remain. It is unclear how DILPs impinge on the GSC division cycle, whether DILPs alone mediate the effect of diet on GSCs, and whether GSCs require proximity to niche cells to respond directly to neural DILPs.
The insulin/insulin-like growth factor (IGF) pathway is evolutionarily conserved and controls many essential processes linked to nutrient sensing, such as metabolism, reproduction, longevity and cell growth and proliferation (Goberdhan and Wilson, 2003; Hafen, 2004). Drosophila has one homolog for each component of the insulin/IGF pathway, including one receptor (Drosophila insulin receptor, or dinr); one exception, however, is the presence of seven dilp genes. Stimulation of cells by insulin-like signals results in activation of the insulin receptor substrate (encoded by the chico gene, in Drosophila) downstream of the receptor, and activation of the Ras/MAPK and phosphoinositide-3 kinase (PI3K) branches of the insulin pathway (Oldham and Hafen, 2003). Activation of PI3K increases the production of phosphatidylinositol (3,4,5)-trisphosphate, which recruits Akt to the plasma membrane, where it becomes activated and phosphorylates several downstream targets. In mammals, the Ras/MAPK pathway is required for cell proliferation in response to IGF-1 (Lu and Campisi, 1992; Tanaka et al., 1996). In contrast, the PI3K pathway is necessary and sufficient to promote DILP-induced growth and proliferation downstream of the insulin receptor substrate-like gene chico during Drosophila development (Goberdhan and Wilson, 2003; Oldham and Hafen, 2003) (Fig. 1D). The requirement for specific branches of the insulin pathway during GSC proliferation, however, has not been previously examined.
Insulin/IGF-mediated growth is negatively regulated by the transcriptional factor FOXO (Forkhead box, sub-group “O”) (Barthel et al., 2005; Puig and Tjian, 2006). Under high insulin signaling, FOXO is phosphorylated by Akt and retained in the cytoplasm. Under low insulin signaling, FOXO translocates from the cytoplasm to the nucleus and activates transcription of its target genes, which have roles in the regulation of cell cycle, protein synthesis and metabolism. In Drosophila, dFOXO mediates the decrease in imaginal disc cell proliferation resulting from reduced insulin signaling, and overexpression of dFOXO results in a starvation-like phenotype (Junger et al., 2003; Kramer et al., 2003; Puig et al., 2003).
In this study, we find that at least two separate mechanisms mediate the effect of diet on the proliferation of GSCs. DILPs regulate the G2 phase of the GSC division cycle via PI3K and dFOXO, while a separate, DILP-independent mediator of diet regulates the G1 phase. Intriguingly, GSC tumors, which escape the niche, still respond to diet via both mechanisms, demonstrating that nutritional inputs modify stem cell activity via multiple signals acting independently of niche controls.
MATERIALS AND METHODS
Drosophila strains and culture
Drosophila stocks were maintained at 22-25°C. yw was used as a wildtype control. chico1, dinrE19, dinr339, dFOXO21, dFOXO25, and bam86 alleles have been described (Drummond-Barbosa and Spradling, 2001; Junger et al., 2003; LaFever and Drummond-Barbosa, 2005; McKearin and Ohlstein, 1995). chico1; dFOXO21/dFOXO25 double mutants were generated by standard crosses. Genomic rescue constructs P{chicoWT4.2}, P{chicoDrk2.1} and P{chicoPI3K9} have been described and are not expected to affect Chico protein stability (Oldham et al., 2002). We also confirmed by RT-PCR that these transgenes are expressed at similar levels. Germline-specific expression of P{UAS.p-TkvAct}, encoding a constitutively active form of the Thickveins receptor, was used to induce GSC tumors (Casanueva and Ferguson, 2004) but, due to technical difficulties with the penetration of antibodies, cell cycle analyses was not conducted. Instead, only bam86 tumors were analyzed. Other genetic elements are described in Flybase (http://flybase.bio.indiana.edu). Flies were cultured in standard medium with wet yeast paste (protein-rich diet) or in an empty vial containing a Kimwipe soaked in 5% molasses (protein-poor diet).
Generation and analysis of mosaic ovarioles
Genetic mosaics were generated as described (LaFever and Drummond-Barbosa, 2005). For chico mosaic analyses, females of the genotype hs-FLP/+; FRT40A chico1/FRT40A arm-lacZ or hs-FLP/; FRT40A chico1/FRT40A arm-lacZ; P{chico*}/+ were generated. (P{chico*} represents genomic rescue constructs.) To induce FLP-mediated recombination, 0- to 3-day old females were heat shocked for 1 h at 37°C twice a day for 3 days, and subsequently transferred to fresh food with dry yeast daily for 10 days before dissection. chico1 homozygous clones were identified by the absence of β-galactosidase (β-gal). GSC division and cyst growth rates were determined as described (LaFever and Drummond-Barbosa, 2005). For dFOXO mosaic analysis, hs-FLP/; FRT 82B dFOXO*/FRT82B arm-lacZ females were generated, heat shocked, and cultured for 10 days as above. (dFOXO* represents wild-type or dFOXO25 alleles.) The dFOXO and control mosaics were then transferred to either rich or poor diets for 0, 2, 5, and 10 days before dissection and analysis as above. Ten days was chosen as the last time point because by then we would expect a nearly complete turnover of the population of cystoblasts and cysts within the germarium on either diet (Drummond-Barbosa and Spradling, 2001; LaFever and Drummond-Barbosa, 2005), which would be important for the detection of any potential changes in the relative division rate of the GSCs (measured as the relative number of β-gal negative and β-gal positive cystoblasts and cysts inside the germaria). Please note that although wild-type GSC division rates are lower on a poor diet (Drummond-Barbosa and Spradling, 2001), the relative division rate remains unchanged if both GSCs within each germarium equally modulate their proliferation upon a dietary switch (see Fig. 2G). Results were subjected to Chi-square analysis (for GSC relative division rates) or Student’s t-test (for cyst growth rates).
Figure 2. DILPs control the rates of GSC division and cyst growth via the PI3K pathway.

(A) FLP/FRT technique used to generate chico1 null clones recognized by the absence of β-gal and expressing chico transgenes. (B) A mosaic germarium used for analysis of GSCs (dashed ovals). Arrowhead, β-gal negative GSC. (C) A mosaic ovariole showing delayed chicoPI3K- mutant cysts (arrowheads). (D) Example of chicoDrk- mutant cysts (arrowheads). (E) The relative division rate of chicoPI3K- GSCs is significantly lower than that of chicoWT GSCs. (F) chicoPI3K- cysts are severely delayed while chicoDrk- cysts are comparable to chicoWT cysts. (G) The relative division rates of dFOXO25 GSCs are not significantly different from those of wild-type GSCs on rich or poor diets. α-spectrin (green) highlights cell membranes and fusomes, whereas β-gal (red) labels control cysts. Ovarioles in C, D are shown at the same magnification. n, number of cystoblasts and cysts counted (E,G) or of β-gal-negative cysts analyzed (F). Scale bars, 10 μm. Asterisks, P<0.001.
Immunostaining and fluorescence microscopy
Ovaries were dissected in Grace’s insect medium (Cambrex), fixed for 13 min at room temperature in Grace’s medium plus 5% formaldehyde (Ted Pella), washed and stained as described (de Cuevas et al., 1996). The following antibodies were used: mouse monoclonal 1B1 (1:10) (Developmental Studies Hybridoma Bank, DSHB), rabbit polyclonal α-spectrin (de Cuevas et al., 1996) (1:100), mouse monoclonal anti-β-gal (1:500) (Promega), rabbit polyclonal anti-β-gal (1:1000) (Cappel), mouse monoclonal anti-CycE 8B10 (Richardson et al., 1995) (1:10), guinea pig polyclonal anti-CycE (1:1000) (T. Orr-Weaver), mouse monoclonal anti-CycB E2F4 (1:20) (DSHB), and rabbit polyclonal anti-PHH3 (1: 250) (Upstate Biotechnology). Alexa 488-, Alexa 568- or Alexa 633-conjugated goat anti-mouse, anti-rabbit and anti-guinea pig secondary antibodies (1:400) (Molecular Probes) were used. Samples were incubated in 1 μg/ml DAPI (Sigma) for 8 min. Ovaries were mounted in Vectashield (Vector Laboratories). BrdU incorporation was performed as described (Lilly and Spradling, 1996). Briefly, dissected ovaries were incubated with 10 μM of BrdU (Sigma) in Grace’s medium for 1 h at room temperature, washed, and fixed with 8% formaldehyde in Grace’s medium for 5 min. After staining with primary antibodies as above, ovaries were fixed again with 12% formaldehyde for 20 min, denatured in 2N HCl for 30 min, and neutralized in 100 mM borax for 2 min. The ovaries were then immunostained using mouse anti-BrdU (1:20) (Becton-Dickinson). Samples were examined using a Zeiss Axioplan 2 (images shown in Figs. 3D,F-I and 4B), or a Zeiss LSM 510 confocal microscope.
Figure 3. CycE is co-expressed with CycB during G2 and M in GSCs and cystoblasts, but not in dividing cysts or follicle cells.
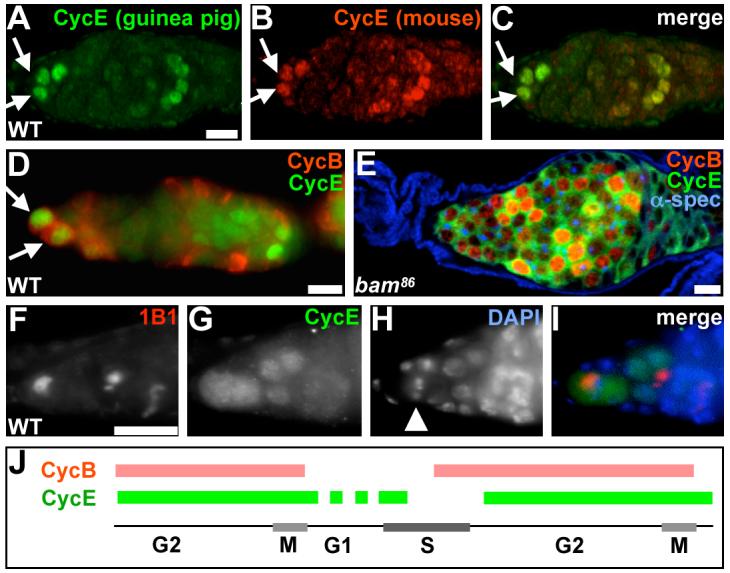
(A-C) Example of a germarium demonstrating the specificity of the anti-CycE antibodies we used. Guinea pig anti-CycE antibodies (green, from T. Orr-Weaver) (A) and mouse anti-CycE antibodies (red, from H. Richardson) (B) show the same staining pattern, as shown in the merged image (C). We found that 76% of 1,660 GSCs analyzed expressed CycE. (D) CycB (red) and CycE (green) are co-expressed in wild-type (WT) GSCs and cystoblasts, but this was never observed in dividing cysts or follicle cells. (E) Similar CycB and CycE co-expression is observed in bam86 GSC tumors. (F-I) Example of mitotic GSC expressing CycE (G). 1B1 (F) labels the fusome, and DAPI (H) is a DNA dye. Arrows in A-D indicate GSCs. Arrowhead in H indicates a GSC mitotic figure. Merged image is shown in I. (J) Diagram summarizing the expression profile of CycB and CycE in GSCs. Expression of CycB (red line) initiates at the end of S phase, and is turned off during M phase. CycE (green line) overlaps with CycB during G2 and M phases. We have also confirmed that CycE is expressed during S phase by double labeling with BrdU (not shown), but we have not directly examined whether or not it is expressed during G1 (dashed green line) in GSCs. Images in A-C are shown at the same magnification. Images in F-I are also shown at the same magnification. Scale bars, 10 μm in A, D and E; 5 μm in F.
Figure 4. Diet controls the rate of GSC division via both the G1 and G2 phases of the cell cycle, but the insulin pathway specifically affects G2.

(A) Cell cycle markers and fusome morphology used to analyze the cell cycle of GSCs. PHH3 and BrdU incorporation are markers for the M and S phases, respectively. CycB is a G2 and M marker. GSC fusomes are juxtaposed to cap cells and change their morphology during the cell cycle (de Cuevas and Spradling, 1998). During M, the GSC fusome has a “round” morphology. After M, the newly formed cystoblast remains attached to the GSC by a ring canal. During G1 and S phases of the next division cycle, the GSC fusome changes its morphology to “plug”, “fusing” and “bar” as a new fusome structure or plug is assembled, fused to the old fusome and partitioned between the GSC and connected cystoblast (CB). During early G2, the GSC fusome has an “exclamation point” morphology, as the connection between the GSC and the cystoblast is severed. Later in G2, the GSC fusome becomes “round” again. (B) The frequency of GSCs in M, detected by the presence of PHH3 (green), is lower on a poor diet compared to that on a rich diet. PHH3 label co-localizes with DAPI-labeled DNA (blue), and 1B1 (red) labels fusomes. (C) The percentage of GSCs in S phase, which are labeled with BrdU (green), is also reduced under a poor diet or low insulin signaling. In chico; dFOXO double mutants, the frequency of BrdU-positive GSCs is significantly increased relative to that of chico mutants. α-spectrin (red) labels fusomes. (D) The percentage of GSCs with a “G1&S” fusome morphology is similar in wild-type females under rich and poor diets, but it is reduced in dinr and chico mutant flies. This reduction is reversed in chico; dFOXO double mutants. (E) The percentage of GSCs displaying “G2&M” fusome morphology or CycB expression does not change in response to diet, but is higher in dinr and chico mutants. This increase is reversed in chico; dFOXO double mutants. In D, E, CycB (green) and α-spectrin (red) are shown. CycB-negative and -positive GSCs are shown in D and E, respectively. GSCs are outlined by dashed ovals. All images are shown at the same magnification. Scale bar, 5 μm. Bars in graphs show standard deviation. Single asterisks, P<0.05. Double asterisk, P<0.001.
Cell cycle analyses
Two-day-old females of genotypes yw, bam86, dFOXO21/dFOXO25, and dFOXO25/+ were maintained on a rich diet for 3 days, and transferred to rich or poor diets for 2 days. dinrE19/dinr339, chico1, chico1; dFOXO21/dFOXO25, and controls were maintained on a rich diet for 5 days. The fusome morphology, visualized by 1B1 or α-spectrin antibodies, was used to identify cell cycle stages (de Cuevas and Spradling, 1998) (see text). Additional markers used were BrdU incorporation (S phase), CycB (G2 and M phases), and PHH3 (M phase). In tumorous germaria, GSC cells were identified by their round fusome morphology, and scored for the presence of BrdU, PHH3, or CycB. Statistical analyses were performed using the Student’s t-test.
RESULTS
The PI3K pathway mediates the effects of insulin-like signals on GSC proliferation and cyst growth
To disrupt activation of the PI3K or Ras/MAPK branches, we used chico genomic rescue transgenes (Oldham et al., 2002) that carry point mutations in the predicted consensus binding sites for either the p60 (chicoPI3K-) or Grb2/Drk (chicoDrk-) adaptor proteins, respectively. We removed the endogenous chico function in GSCs expressing these point mutants using the flipase (FLP)/FLP-recognition target (FRT) technique (Fig. 2A-D), and determined the ratio of mutant to control cystoblasts and cysts within mosaic germaria as a measure of the relative division rates of chico mutant GSCs (LaFever and Drummond-Barbosa, 2005; Xie and Spradling, 1998) (Fig. 2E). The reduced relative division rate of chico1 mutant GSCs in the absence of a chico transgene (chico1 control) was comparable to that of previously described dinr mutant GSCs (LaFever and Drummond-Barbosa, 2005).
Expression of a wild-type chico transgene (chicoWT) restored the GSC relative division rate to wild-type levels. In contrast, expression of the chicoPI3K- transgene resulted in a GSC relative division rate indistinguishable from that of the chico1 control, indicating that activation of PI3K downstream of chico is absolutely required for the upregulation of GSC proliferation in response to DILPs. The activity of the chicoDrk- transgene was equivalent to that of chicoWT, suggesting that the Ras/MAPK branch is dispensable. Similar requirements were found for cyst growth (Fig. 2C,D,F) and progression through vitellogenesis (0% of fully mutant chico1 control [n=12] or chicoPI3K- [n=7] ovarioles showed progression into vitellogenesis instead of degeneration, whereas 100% of chicoWT [n=7] or chicoDrk- [n=5] ovarioles contained vitellogenic egg chambers). These results show that the direct effect of DILPs on GSC proliferation, cyst growth and vitellogenesis are entirely mediated by PI3K during the response to diet. Thus, they reveal a considerable degree of similarity between the control of proliferation by DILPs in growing larval cells and in adult GSCs.
dFOXO is not required to maintain the repressed state of GSCs under a poor diet
Although mutation of the Drosophila homolog dFOXO does not result in any overt phenotype, dFOXO is required for the inhibitory effects of low insulin signaling on the proliferation of larval tissues (Puig and Tjian, 2006). Thus, we asked if dFOXO is required to maintain the repressed state of GSCs under a poor diet. We generated clones of cells homozygous for the null dFOXO25 mutation (Junger et al., 2003), and measured the relative division rates of dFOXO25 GSCs in mosaic females transferred to a poor diet. We reasoned that, if dFOXO were required to inhibit GSC proliferation on a poor diet (when insulin signaling is low), dFOXO25 GSCs would divide faster than control GSCs within mosaic germaria, resulting in a higher relative division rate on a poor diet relative to that on a rich diet (see Materials and Methods).
As expected, we found that on a rich diet (when dFOXO is normally off) the relative division rate of dFOXO25 GSCs was comparable to that of wild-type GSCs (Fig. 2G). Surprisingly, the relative division rates of dFOXO25 GSCs remained statistically unchanged after 2, 5, and 10 days on a poor diet, similarly to those of wild-type GSCs. dFOXO25 cyst growth rates and progression through vitellogenesis were also comparable to those of the wild-type control on both rich and poor diets (Table 1). These results indicate that dFOXO25 GSCs reduce their proliferation rates in response to a poor diet to the same extent as wild-type GSCs and thus, that dFOXO is not required to maintain the poor diet-induced repressed state of GSCs. It is possible that the effects of low insulin signaling are dFOXO-independent. Alternatively, other signals may be required to modulate GSC division in response to diet such that activation of the insulin pathway by removal of dFOXO is not sufficient to compensate for the effects of a poor diet.
Table 1.
dFOXO is not required for the response to a protein-poor diet
| Strain | Diet | Days after switch* | GSC relative division rate† | Cyst growth rate† | Vitellogenesis rate† |
|---|---|---|---|---|---|
| Wild type | Rich | 0 | 0.79 (528)‡ | 100% (54)‡ | - |
| 2 | 0.72 (453) | 100% (52) | 100% (3)ठ| ||
| 5 | 0.73 (630) | 100% (60) | 100% (8) | ||
| 10 | 0.82 (426) | 100% (73) | 100% (6) | ||
| Poor | 2 | 0.73 (541) | 100% (21) | 0% (2) | |
| 5 | 0.82 (462) | 100% (77) | 25% (4) | ||
| 10 | 0.71 (72) | 100% (14) | 0% (1) | ||
| dFOXO25 | Rich | 0 | 0.74 (586) | 100% (62) | - |
| 2 | 0.58 (459) | 100% (14) | - | ||
| 5 | 0.76 (603) | 100% (61) | 100% (9) | ||
| 10 | 0.87 (518) | 100% (98) | 100% (18) | ||
| Poor | 2 | 0.68 (311) | 100% (42) | 0% (3) | |
| 5 | 0.75 (459) | 100% (87) | 0% (4) | ||
| 10 | 0.72 (330) | 100% (38) | 0% (8) | ||
Females were maintained for 10 days on a rich diet after generation of clones, and subsequently transferred to either rich or poor diets for the indicated number of days.
Cyst growth rates are expressed as a percentage of the wild-type development rate, and vitellogenesis rates represent the percentage of cysts within ovarioles containing fully β-gal negative germline that initiated vitellogenesis instead of degenerating. The results obtained from dFOXO25 clonal analyses were not statistically different from those of the wild-type control.
The total number of cystoblasts and cysts, β-gal-negative cysts, and ovarioles containing fully β-gal negative germline analyzed, respectively, are shown in parentheses.
The small number of fully β-gal negative ovarioles analyzed reflects the rarity of double GSC recombination events within a single germarium.
Cyclin E is not a valid G1 cell cycle marker in GSCs because it is highly expressed during the G2 and mitosis (M) phases
To analyze and compare changes in cell cycle parameters of GSCs in response to diet and insulin signaling, it was necessary to identify appropriate cell cycle markers for these analyses. Cyclin (Cyc) E is typically expressed during late G1 and turned off during early S phase (Ekholm et al., 2001). The expression pattern of CycB, a known late S and G2 marker (Bassermann et al., 2005), has been previously analyzed specifically in GSCs and shown to reach its highest level during late G2, and subsequently decrease during early M phase (Wang and Lin, 2005). The CycB pattern that we observed was entirely consistent with this earlier description (see Fig. 4D,E). Surprisingly, however, we found that 76% of GSCs (n=1660) expressed CycE, and that CycE was co-expressed with CycB in 68% of all GSCs (n=763) during G2 and M phases (Fig. 3). CycE was also co-expressed with CycB in cystoblasts, but co-expression was never observed in dividing cysts or follicle cells in 178 ovarioles analyzed. The finding that CycE behaves in an unusual manner in GSCs indicates that CycE is not a valid G1 marker for these cells. Thus, we took advantage instead of the well characterized cycle that the fusome undergoes during GSC division (de Cuevas and Spradling, 1998) (see Fig. 4A).
Diet regulates both the G1 and G2 phases of the GSC division cycle, but insulin signaling mediates only the effect on G2
Several lines of evidence suggest that diet and insulin/IGF signaling control the G1 phase of the cell cycle both in vivo and in culture (Cameron and Cleffmann, 1964; Koga and Kimura, 1980; Lu and Campisi, 1992). Effects on the G2 phase have also been reported (Adesanya et al., 1999; Cameron and Cleffmann, 1964; Frederick and Wood, 2004; Fukuyama et al., 2006). During Drosophila development, however, insulin signaling components appear to specifically regulate the length of G1 (Goberdhan and Wilson, 2003). We thus asked how diet affects GSC division, and whether changes in insulin signaling levels account for all of the effects of diet on their proliferation rates. To compare the cell cycle profiles of GSCs upon manipulation of diet or insulin receptor (dinr) activity, we identified GSCs based on their position relative to cap cells (see Fig.1B) and on the morphology of the fusome, an early germline-specific membranous structure (de Cuevas and Spradling, 1998), and examined the frequency of cell cycle markers (Fig. 4A). Cells in S phase were identified by BrdU incorporation, cells in G2 or M expressed CycB, and those in M were positive for the phosphohistone H3 (PHH3) epitope. The fusome morphology was also used to identify cell cycle stages (de Cuevas and Spradling, 1998). Specifically, cells in G1 or S had a fusome with “plug”, a“fusing”, or a “bar-shaped” fusome (referred to as “G1&S” fusomes), while cells in G2 or M had an “exclamation point” or a “round” fusome abutting the cap cells (referred to as “G2&M” fusomes).
As expected based on the known effect of diet on GSCs (Drummond-Barbosa and Spradling, 2001), the frequencies of GSCs positive for the PHH3 marker and BrdU incorporation under a poor diet were reduced 1.8-fold relative to those on a rich diet (Fig. 4B,C and Table 2), reflecting the lower rates of GSC proliferation under a poor diet. In agreement with the well documented role of DILPs in this process (Drummond-Barbosa and Spradling, 2001; LaFever and Drummond-Barbosa, 2005), the percentages of BrdU-positive GSCs in dinr or chico mutant females were also reduced relative to that of the wild-type. The reduction in GSC proliferation rate in response to a poor diet or low insulin signaling suggests that GSCs must delay cell cycle progression at either G1, G2, or both.
Table 2.
Frequencies of GSCs positive for cell cycle markers
| Strain | Diet* | Percentage of cell cycle marker-positive GSCs† |
||||
|---|---|---|---|---|---|---|
| PHH3 | BrdU | G1&S fusome | CycB | G2&M fusome | ||
| yw control | Rich | 3.7±0.8 (2405)‡ | 11±2.2 (2176) | 28±1.3 (738) | 72±6.2 (2315) | 72±1.2 (738) |
| Poor | 2.1±0.4§ (2476) | 6.5±1.0§ (1985) | 28±3.3 (719) | 77±4.8 (2390) | 72±3.3 (719) | |
| Poor 18d | - | 7.6 (224) | 26 (224) | - | 72 (224) | |
| dinr339/+ | Rich | - | 9.6±2.1 (1729) | 29±3.9 (510) | 76±4.9 (585) | 71±3.9 (510) |
| dinrE19/+ | Rich | - | 11±1.5 (1309) | - | - | - |
| dinrE19/dinr339 | Rich | - | 2.1±1.5∥ (476) | 17±3.2§ (642) | 87±2.7§ (638) | 83±3.2§ (642) |
| chico1/+ | Rich | - | 10±1.6 (588) | 28±5.6 (319) | 76±0.7 (322) | 71±6.5 (319) |
| chico1 | Rich | - | 2.5±1.2∥ (240) | 12±5.2§ (808) | 86±4.7§ (240) | 88±5.1§ (808) |
| chico1; dFOXO21/dFOXO25 | Rich | - | 8.1±0.5¶ (694) | 27±2.3¶ (694) | - | 71±2.5¶ (694) |
| dFOXO25/+ | Rich | - | 9.2±1.5 (996) | 36±4.6 (996) | - | 64±4.0 (996) |
| Poor | - | 5.7±1.2 (807) | 30±7.9 (807) | - | 70±7.6 (807) | |
| dFOXO21/dFOXO25 | Rich | - | 6.1±1.6 (1063) | 27±2.4 (1063) | - | 73±2.7 (1063) |
| Poor | 4.9±1.5 (1142) | 22±3.4 (1142) | - | 78±3.1 (1142) | ||
Females were kept on a protein-rich diet for 5 days (Rich), on a rich diet for 3 days and transferred to a poor diet for 2 days (Poor), or on a rich diet for 3 days and transferred to a poor diet for 18 days (Poor 18d).
All results represent the mean±s.d. of at least three independent experiments, with the exception of yw Poor 18d females, which were analyzed twice (only the mean is shown).
The total number of GSCs analyzed is shown in parentheses.
Values obtained were significantly different from those of yw Rich females (P<0.05).
Values obtained were significantly different from those of yw Rich females (P<0.001).
Values obtained were significantly different from those of chico1 Rich females (P<0.001).
We next measured the relative length of time that GSCs spend in the G1 versus G2 phases of the cell cycle. On a rich diet, approximately 30% of the GSCs exhibited “G1&S” fusome morphology, while the percentage of GSCs displaying “G2&M” fusome morphology or expressing CycB was 70%, indicating that GSCs normally have a relatively long G2 phase (Fig. 4D,E and Table 2). If GSCs delayed cell cycle progression specifically during G1 or G2 in response to a poor diet, we would expect a relatively higher proportion of GSCs exhibiting the corresponding marker relative to that on a rich diet. In contrast, we find that on a poor diet the frequency of GSCs displaying “G1&S” fusomes remained comparable to that on a rich diet, as did the percentage of GSCs with “G2&M” fusomes/CycB expression (Fig. 4D,E and Table 2). When the observed decrease in the frequencies of GSCs specifically in M or S phases (see Fig. 4B,C) is factored in, these results indicate that both G1 and G2 are lengthened in response to a poor diet. We obtained similar results in females kept on a poor diet for 18 days (Table 2), suggesting that progression through both G1 and G2 remain proportionately slow over time under a poor diet.
In contrast to what we find for wild-type GSCs under a poor diet, the percentage of GSCs with “G1&S” fusome morphology was decreased to approximately 17% in dinr mutant females, while the percentage of GSCs displaying “G2&M” fusome morphology was increased to 83% (Fig. 4D,E and Table 2). Consistent with that, the percentage of CycB-positive GSCs was 87% in dinr mutants. Similar results were obtained in chico mutant females. These data suggest that a reduction in insulin signaling in GSCs predominantly slows down the progression through the G2 phase of the cell cycle. Importantly, these results demonstrate that while DILPs control the G2 phase, a separate diet-dependent signal controls G1 progression of the GSCs.
Reduced insulin signaling lengthens the G2 phase of GSCs via dFOXO
Although our results clearly demonstrated that DILPs are not solely responsible for the regulation of GSCs in response to diet, it remained unclear whether or not dFOXO mediates the effects of low insulin signaling on G2 under a poor diet. We therefore asked whether dFOXO is required for lengthening the G2 phase under low insulin signaling by comparing the cell cycle markers in GSCs of chico versus chico; dFOXO null mutant females (Fig. 4C-E and Table 2). Elimination of dFOXO function suppresses the extended G2 caused by reduced insulin signaling in chico; dFOXO double mutants, as indicated by the reduced frequency of GSCs displaying “G2&M” fusomes relative to that in chico mutants. The reduction in “G2&M” fusome frequency is accompanied by an increase in the frequency of BrdU-positive and “G1&S” fusome GSCs in chico; dFOXO mutants. Intriguingly, dFOXO mutation is not sufficient to eliminate the lengthening of G2 on a poor diet, as the frequency of GSCs in G2 in dFOXO mutant females was not decreased on a poor diet relative to that of control females (Table 2). These results demonstrate that low insulin signaling extends the G2 phase of GSCs in a dFOXO-dependent manner, but that redundant mechanisms lengthen G2 in response to a poor diet.
Interactions with a normal niche are not required for the dietary control of GSC proliferation at the level of either the G1 or G2 phases of the cell cycle
The direct regulation of GSCs by DILPs suggested that it may not depend on their presence within a normal niche (LaFever and Drummond-Barbosa, 2005); however, it was still possible that normal niche architecture might be required for the competence of GSCs to respond to DILPs. In addition, it was unclear whether the signal(s) mediating the effect of diet on G1 was dependent on the niche. Dpp, a transforming growth factor-β signal produced by the niche, is required to maintain the GSC fate via repression of the differentiation factor bam (Li and Xie, 2005). To address whether an intact niche is required for the response of GSCs to diet, we tested this response in GSC tumors induced by the removal of bam function (Fig. 5A,B). We found that GSC-like cells within these tumors still respond to diet (Fig. 5C,D). As in normal GSCs, both G1 and G2 are lengthened on a poor diet, as indicated by the similar frequency of CycB-positive tumor GSCs on rich and poor diets (Fig. 5E). These results suggest that the presence of GSCs within an intact niche is not required for either the DILP-dependent G2 control or the G1 regulation by diet (Fig. 6).
Figure 5. Tumor GSCs can respond to diet similarly to normal GSCs.
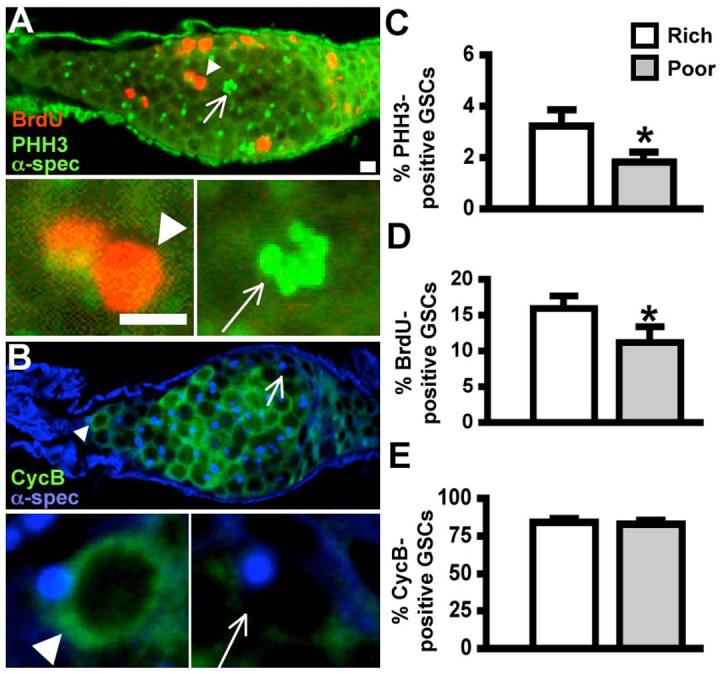
(A) bam86 germarium labeled with BrdU (red), PHH3 (green) and α-spectrin (green, fusome). Examples of BrdU positive (arrowhead) and PHH3-positive (arrow) GSCs are shown below at a higher magnification. (B) bam86 germarium stained for CycB (green) and α-spectrin (blue, fusome). Examples of CycB-positive (arrowhead) and -negative (arrow) GSCs are shown below at a higher magnification. (C,D) GSCs within bam86 tumors downregulate their proliferation under a poor diet, as shown by the decreased percentage of PHH3-positive (C) and BrdU-positive (D) GSCs. (E) The frequency of CycB-positive GSCs remains unchanged during the response of bam86 tumors to diet, indicating the both G1 and G2 are lengthened under a poor diet (see text). Scale bars, 5 μm. Bars in graphs show standard deviation. Asterisks, P<0.05.
Figure 6. A model describing how diet regulates GSC division in parallel to niche signals.

Multiple signals act in parallel and independently of the niche to regulate the response of GSCs to diet. (A) On a rich diet, neural DILPs directly activate the insulin receptor on GSCs to stimulate PI3K activity, repression of dFOXO, and faster progression through G2, while another diet-controlled signal(s) regulates G1 progression. (B) On a poor diet, reduced insulin signaling results in dFOXO activation, which in turn represses progression through the G2 phase. In addition, regulation by redundant diet-dependent signals ensures that cell cycle progression through both G1 and G2 remains slow when nutritional inputs are limited.
DISCUSSION
Multiple signals mediate the response of GSCs to diet
The environment exerts tremendous pressure during the evolution of biological processes to such a degree that many of these processes themselves can adjust to frequent, short-term external changes. Stem cells often lie at the root of these processes due to their critical role in maintaining the function and integrity of many adult tissues. It is, therefore, very likely that stem cells in other systems will exhibit responses to environmental stimuli comparable to the response of ovarian stem cells to diet in the Drosophila ovary. In fact, parallels can be drawn between the effects of DILPs on GSCs and those of insulin/IGFs in other stem cell systems. For example, reduced insulin signaling results in the decreased proliferation of germline precursors in Caenorhabditis elegans during preparation for dauer diapause (Narbonne and Roy, 2006a), while in adults it inhibits gamete production (Gems et al., 1998). In addition, focal cerebral ischemia results in increased proliferation of adult mammalian neural progenitor cells in the subgranular zone of the dentate gyrus and in the subventricular zone of the lateral ventricles, and this response was shown to require IGF-1 activity in rats (Yan et al., 2006). Similarly, IGF-2 is required for fetal liver cells to support the proliferation of hematopoietic stem cells in culture (Zhang and Lodish, 2004). Finally, insulin normalizes delayed corneal wound healing in diabetic rats (Zagon et al., 2007), although it is unclear whether or not the activity of corneal epithelial stem cells is affected.
Fast and effective responses of stem cells to complex stimuli such as diet would be expected to result from multiple signals. Our data provide evidence that in the case of Drosophila GSCs, in addition to DILPs regulating G2, at least one other signal mediates the effect of diet on GSCs via G1 and possibly G2 as well (Fig. 6). It is possible that the regulation of G1 involves the Target of Rapamycin (TOR) kinase. TOR integrates many stimuli such as amino acid levels, metabolic status, or signaling inputs, and it has a known role in growth control (Oldham and Hafen, 2003). In addition, it has been shown to regulate the cell cycle via the G1 phase (Chan, 2004) and appears to be required for normal ovarian function in Drosophila (Zhang et al., 2006). Alternatively, the effects of diet on G1 may be mediated by microRNAs, which have been reported to regulate GSC division in Drosophila (Hatfield et al., 2005). Although Dicer-1 (Dcr-1), the gene encoding the ribonuclease III required for microRNA biogenesis, was proposed to regulate the G1/S transition in GSCs, this conclusion was based on the increased frequency of Dcr-1 mutant GSCs expressing CycE, also accompanied by higher numbers of GSCs positive for the cyclin-dependent kinase inhibitor Dacapo (Hatfield et al., 2005). Our results demonstrate that CycE is normally present during G2 and M in GSCs and, therefore, that changes in the frequencies of CycE-positive GSCs are difficult to interpret. It is likely that Dacapo also behaves differently in GSCs, given that CycE regulates its expression (de Nooij et al., 2000). Thus, it remains unclear what phase of the division cycle is lengthened in Dcr-1 mutant GSCs, and whether or not Dcr-1 may mediate the G1 effect of diet.
The control of proliferation by DILPs shows similarities and differences between GSCs and larval somatic cells
The G2-specific effect of insulin signaling in GSCs is in contrast to other findings in Drosophila. For example, in haemocyte-derived Drosophila cell lines, insulin independently stimulates G1/S progression and inhibits G2/M progression (Wu et al., 2007). Similarly, overexpression of the catalytic subunit of PI3K increases the rate of progression through G1 in wing imaginal discs (Weinkove et al., 1999). Loss of the PTEN tumor suppressor, a negative regulator of PI3K, also shortens the G1 phase of the division cycle of wing imaginal disc cells (Gao et al., 2000). It is possible that the G2 regulation of GSCs by DILPs may reflect specific properties of these stem cells (see below). Interestingly, germline precursors undergo G2 arrest in C. elegans insulin receptor mutants or hatchlings deprived of food, and this arrest is bypassed by loss of PTEN (Fukuyama et al., 2006; Narbonne and Roy, 2006a). IGF-1 is also required for G2/M progression in oligodendrocyte progenitor cells in culture (Frederick and Wood, 2004).
The apparently exclusive role that we find for the PI3K branch of the insulin pathway in mediating the effects of DILPs on GSC proliferation and cyst growth has also been demonstrated in proliferating larval cells (Goberdhan and Wilson, 2003; Oldham and Hafen, 2003). The fact that disruption in the insulin receptor substrate Chico of the predicted consensus binding site for Grb2/Drk, the adaptor protein in the Ras/MAPK branch, results in no obvious phenotype in this or other studies (Oldham et al., 2002) raises the concern that perhaps it does not completely abolish Ras/MAPK activation downstream of insulin receptor activation. Indeed, the cytoplasmic region of the Drosophila insulin receptor can induce both PI3K and MAPK activation in the absence of the insulin receptor substrate in cultured 32D cells, although it still requires the insulin receptor substrate for mitogenesis (Yenush et al., 1996). Nevertheless, it is worth noting that disruption of the consensus binding site for p60, the PI3K adaptor, leads to phenotypes indistinguishable from those resulting from the complete elimination of chico function. Furthermore, the Grb2/Drk binding site is conserved in Drosophila, suggesting that Ras/MAPK activation via Chico in response to DILPs may have a minor or, perhaps, context-specific role undetectable in our experiments. In fact, Ras has been shown to activate PI3K in Drosophila and mammals (Prober and Edgar, 2002; Rodriguez-Viciana et al., 1996) and, although Ras-mediated regulation of PI3K is dispensable for viability, it is required for maximal PI3K signaling in specific biological contexts (Orme et al., 2006). It remains unclear, however, whether an optimal effect of Ras requires the presence of an intact Grb2/Drk binding site in Chico.
We also demonstrate that the PI3K pathway regulates GSC division via dFOXO downstream of insulin receptor signaling. These results are similar to the earlier finding that dFOXO function is required to mediate the reduction in cell proliferation under low insulin signaling levels during development (Junger et al., 2003). Also, analogously to the reduced wing size reported in dFOXO mutants (Junger et al., 2003), we also find that dFOXO mutants appear to have a reduced rate of GSC division on a rich diet (see Table 2). This is probably due to either non-cell autonomous effects or to strain background differences, given that the rates of proliferation of dFOXO mutant GSCs are comparable to those of wild type GSCs in mosaic germaria (see Fig. 2G). Interestingly, elimination of dFOXO function is not sufficient to compensate for the effects of a poor diet on G2, suggesting further redundancy in the poor diet-induced repression of GSC division. These results are reminiscent of the reported starvation-induced G2 arrest of C. elegans germline precursors, which also does not require FOXO function (Fukuyama et al., 2006).
GSCs spend a large proportion of their division cycle in the G2 phase
The relatively long G2 phase in female Drosophila GSCs could conceivably represent an adaptation to their function. One speculative possibility is that new fusome components are synthesized in GSCs during G2, taking advantage of their doubled DNA content, for incorporation into new fusome structures that are assembled, fused to old fusome material, and partitioned between the GSC and newly formed cystoblast beginning early in the G1 phase of the next division cycle (see Fig. 4A) (see de Cuevas and Spradling, 1998). Although the fusome represents a developmental modification of the endoplasmic reticulum (Snapp et al., 2004), it is a very large structure that establishes intercellular connections and has a highly specialized function (Lin et al., 1994); thus, synthesis of all the components necessary to assemble it may potentially be a time-demanding process. Indeed, other instances exist in which long G2 phases allow for completion of time-consuming processes. In the fission yeast Schizosaccharomyces pombe, which normally has a very short G1 phase perhaps to minimize DNA damage during a stage when only one copy of its haploid genome is present, a long G2 phase allows for the increase in cell mass prior to M (Nasmyth et al., 1991). In fertilized Xenopus eggs, a prolonged G2 phase in the first mitotic cell cycle is thought to allow karyogamy to be successfully completed (Walter et al., 2000).
Alternatively, the extended G2 phase of GSCs may maximize the accuracy of the genetic information to be passed on to the next generation by compensating for potential decatenation checkpoint shortcomings. It has been reported that the decatenation checkpoint, which normally delays entry into mitosis until chromosomes have been fully disentangled, is highly inefficient in several stem and progenitor cells (Damelin et al., 2005). This may also be the case in GSCs. Yet another possibility is that the relatively long G2 of GSCs, coupled to its regulation by DILPs, allows for more immediate, post DNA synthesis changes in the rate of production of new daughter cells, similar to what has been proposed for wound repair in the transitional epithelium of the rat urinary bladder (Kaneko et al., 1984).
The unusual pattern of cyclin E expression in GSCs may contribute to their “stemness”
In most proliferating cells, CycE expression starts in late G1 phase, reaches its maximum at S-phase entrance, and then undergoes ubiquitin-mediated proteolysis (Moroy and Geisen, 2004). In Drosophila GSCs, however, we show that CycE is also expressed at high levels during most of G2 and M, suggesting potential differences in the cell cycle regulation of GSCs. It is possible, for example, that the short G1 phase of GSCs is a consequence of the overall high levels of CycE throughout most of their cell cycle, including in the preceding M phase. In mammalian cultured cells, overexpression of CycE shortens G1 phase, suggesting that cyclin levels are rate-limiting for the transition from G1 to S (Ohtsubo and Roberts, 1993). Interestingly, mammalian embryonic stem cells also have a very short G1 phase (Becker et al., 2006; Fluckiger et al., 2006; Savatier et al., 1994; Stead et al., 2002) and constitutive expression of CycE throughout the cell cycle (Fluckiger et al., 2006; Stead et al., 2002). Given that the length of the G1 phase corresponds to a window of increased sensitivity to differentiation signals (Dehay and Kennedy, 2007), it is conceivable that shortening the G1 phase might contribute to the maintenance of the undifferentiated state of GSCs, as previously proposed for mammalian embryonic stem cells (Burdon et al., 2002).
Stem cells and tumors share the ability to respond to diet
GSC tumors respond to diet as normal GSCs do and have a similar pattern of CycE expression. Many human cancer cell lines also exhibit abnormal regulation of CycE expression (Darzynkiewicz et al., 1996; Juan and Cordon-Cardo, 2001; Keyomarsi et al., 1995). Our finding that tumorous and normal GSCs respond to diet similarly is interesting in light of the well documented influence of dietary factors on the incidence and prognosis of cancer (Uauy and Solomons, 2005), the connection between cancer risk and elevated levels of circulating insulin and IGF-1 (Voskuil et al., 2005), and the growing evidence suggesting that cancer stem cells are essential for tumor formation and maintenance (Clarke and Fuller, 2006).
ACKNOWLEDGEMENTS
We thank E. Hafen, B. Ohlstein, C. Ferguson, and the Bloomington Stock Center for Drosophila stocks. We also thank D. Branton, H. Richardson, T.Orr-Weaver, and the Developmental Studies Hybridoma Bank for antibodies. We are grateful to B. Appel, D. Greenstein, and E. Ables for valuable comments on this manuscript. Supported by Development Funds from the Vanderbilt University School of Medicine, by National Institutes of Health grant no. GM 069875, and by American Cancer Society grant no. RSG-07-182-01-DDC. Support for L.L. was provided by National Institutes of Health research training grant 2T32HD007502.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
REFERENCES
- Adesanya OO, et al. Insulin-like growth factor 1 is required for G2 progression in the estradiol-induced mitotic cycle. Proc. Natl. Acad. Sci. U S A. 1999;96:3287–91. doi: 10.1073/pnas.96.6.3287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barthel A, et al. FoxO proteins in insulin action and metabolism. Trends Endocrinol. Metab. 2005;16:183–9. doi: 10.1016/j.tem.2005.03.010. [DOI] [PubMed] [Google Scholar]
- Bassermann F, et al. Mitotic entry: a matter of oscillating destruction. Cell Cycle. 2005;4:1515–7. doi: 10.4161/cc.4.11.2192. [DOI] [PubMed] [Google Scholar]
- Becker KA, et al. Self-renewal of human embryonic stem cells is supported by a shortened G1 cell cycle phase. J. Cell. Physiol. 2006;209:883–93. doi: 10.1002/jcp.20776. [DOI] [PubMed] [Google Scholar]
- Burdon T, et al. Signalling, cell cycle and pluripotency in embryonic stem cells. Trends Cell Biol. 2002;12:432–8. doi: 10.1016/s0962-8924(02)02352-8. [DOI] [PubMed] [Google Scholar]
- Cameron IL, Cleffmann G. Initiation of Mitosis in Relation to the Cell Cycle Following Feeding of Starved Chickens. J. Cell Biol. 1964;21:169–74. doi: 10.1083/jcb.21.2.169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casanueva MO, Ferguson EL. Germline stem cell number in the Drosophila ovary is regulated by redundant mechanisms that control Dpp signaling. Development. 2004;131:1881–90. doi: 10.1242/dev.01076. [DOI] [PubMed] [Google Scholar]
- Chan S. Targeting the mammalian target of rapamycin (mTOR): a new approach to treating cancer. Br. J. Cancer. 2004;91:1420–4. doi: 10.1038/sj.bjc.6602162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke MF, Fuller M. Stem cells and cancer: two faces of eve. Cell. 2006;124:1111–5. doi: 10.1016/j.cell.2006.03.011. [DOI] [PubMed] [Google Scholar]
- Damelin M, et al. Decatenation checkpoint deficiency in stem and progenitor cells. Cancer Cell. 2005;8:479–84. doi: 10.1016/j.ccr.2005.11.004. [DOI] [PubMed] [Google Scholar]
- Darzynkiewicz Z, et al. Cytometry of cyclin proteins. Cytometry. 1996;25:1–13. doi: 10.1002/(SICI)1097-0320(19960901)25:1<1::AID-CYTO1>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- de Cuevas M, et al. alpha-spectrin is required for germline cell division and differentiation in the Drosophila ovary. Development. 1996;122:3959–68. doi: 10.1242/dev.122.12.3959. [DOI] [PubMed] [Google Scholar]
- de Cuevas M, Spradling AC. Morphogenesis of the Drosophila fusome and its implications for oocyte specification. Development. 1998;125:2781–9. doi: 10.1242/dev.125.15.2781. [DOI] [PubMed] [Google Scholar]
- de Nooij JC, et al. Expression of the cyclin-dependent kinase inhibitor Dacapo is regulated by cyclin E. Mech. Dev. 2000;97:73–83. doi: 10.1016/s0925-4773(00)00435-4. [DOI] [PubMed] [Google Scholar]
- Dehay C, Kennedy H. Cell-cycle control and cortical development. Nat. Rev. Neurosci. 2007;8:438–50. doi: 10.1038/nrn2097. [DOI] [PubMed] [Google Scholar]
- Drummond-Barbosa D. Regulation of stem cell populations. In: Meyers RA, editor. Encyclopedia of Molecular Cell Biology and Molecular Medicine. Vol. 12. WILEY-VCH; Weinheim: 2005. pp. 67–98. [Google Scholar]
- Drummond-Barbosa D, Spradling AC. Stem cells and their progeny respond to nutritional changes during Drosophila oogenesis. Dev. Biol. 2001;231:265–78. doi: 10.1006/dbio.2000.0135. [DOI] [PubMed] [Google Scholar]
- Ekholm SV, et al. Accumulation of cyclin E is not a prerequisite for passage through the restriction point. Mol. Cell. Biol. 2001;21:3256–65. doi: 10.1128/MCB.21.9.3256-3265.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fluckiger AC, et al. Cell cycle features of primate embryonic stem cells. Stem Cells. 2006;24:547–56. doi: 10.1634/stemcells.2005-0194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frederick TJ, Wood TL. IGF-I and FGF-2 coordinately enhance cyclin D1 and cyclin E-cdk2 association and activity to promote G1 progression in oligodendrocyte progenitor cells. Mol. Cell. Neurosci. 2004;25:480–92. doi: 10.1016/j.mcn.2003.11.015. [DOI] [PubMed] [Google Scholar]
- Fukuyama M, et al. C. elegans DAF-18/PTEN mediates nutrient-dependent arrest of cell cycle and growth in the germline. Curr. Biol. 2006;16:773–9. doi: 10.1016/j.cub.2006.02.073. [DOI] [PubMed] [Google Scholar]
- Gao X, et al. Drosophila PTEN regulates cell growth and proliferation through PI3K-dependent and -independent pathways. Dev. Biol. 2000;221:404–18. doi: 10.1006/dbio.2000.9680. [DOI] [PubMed] [Google Scholar]
- Gems D, et al. Two pleiotropic classes of daf-2 mutation affect larval arrest, adult behavior, reproduction and longevity in Caenorhabditis elegans. Genetics. 1998;150:129–55. doi: 10.1093/genetics/150.1.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goberdhan DC, Wilson C. The functions of insulin signaling: size isn’t everything, even in Drosophila. Differentiation. 2003;71:375–97. doi: 10.1046/j.1432-0436.2003.7107001.x. [DOI] [PubMed] [Google Scholar]
- Hafen E. Cancer, type 2 diabetes, and ageing: news from flies and worms. Swiss Med. Wkly. 2004;134:711–9. doi: 10.4414/smw.2004.09885. [DOI] [PubMed] [Google Scholar]
- Hatfield SD, et al. Stem cell division is regulated by the microRNA pathway. Nature. 2005;435:974–8. doi: 10.1038/nature03816. [DOI] [PubMed] [Google Scholar]
- Juan G, Cordon-Cardo C. Intranuclear compartmentalization of cyclin E during the cell cycle: disruption of the nucleoplasm-nucleolar shuttling of cyclin E in bladder cancer. Cancer Res. 2001;61:1220–6. [PubMed] [Google Scholar]
- Junger MA, et al. The Drosophila forkhead transcription factor FOXO mediates the reduction in cell number associated with reduced insulin signaling. J. Biol. 2003;2:20. doi: 10.1186/1475-4924-2-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaneko H, et al. The presence of G1 and G2 populations in normal epithelium of rat urinary bladder. Basic Appl. Histochem. 1984;28:41–57. [PubMed] [Google Scholar]
- Keyomarsi K, et al. Deregulation of cyclin E in breast cancer. Oncogene. 1995;11:941–50. [PubMed] [Google Scholar]
- Kirilly D, Xie T. The Drosophila ovary: an active stem cell community. Cell Res. 2007;17:15–25. doi: 10.1038/sj.cr.7310123. [DOI] [PubMed] [Google Scholar]
- Koga A, Kimura S. Influence of restricted diet on the cell cycle in the crypt of mouse small intestine. J. Nutr. Sci. Vitaminol. 1980;26:33–8. doi: 10.3177/jnsv.26.33. [DOI] [PubMed] [Google Scholar]
- Kramer JM, et al. Expression of Drosophila FOXO regulates growth and can phenocopy starvation. BMC Dev. Biol. 2003;3:5. doi: 10.1186/1471-213X-3-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaFever L, Drummond-Barbosa D. Direct control of germline stem cell division and cyst growth by neural insulin in Drosophila. Science. 2005;309:1071–3. doi: 10.1126/science.1111410. [DOI] [PubMed] [Google Scholar]
- Li L, Xie T. Stem cell niche: structure and function. Annu. Rev. Cell Dev. Biol. 2005;21:605–31. doi: 10.1146/annurev.cellbio.21.012704.131525. [DOI] [PubMed] [Google Scholar]
- Lilly MA, Spradling AC. The Drosophila endocycle is controlled by Cyclin E and lacks a checkpoint ensuring S-phase completion. Genes Dev. 1996;10:2514–26. doi: 10.1101/gad.10.19.2514. [DOI] [PubMed] [Google Scholar]
- Lin H, et al. The Drosophila fusome, a germline-specific organelle, contains membrane skeletal proteins and functions in cyst formation. Development. 1994;120:947–56. doi: 10.1242/dev.120.4.947. [DOI] [PubMed] [Google Scholar]
- Lu K, Campisi J. Ras proteins are essential and selective for the action of insulin-like growth factor 1 late in the G1 phase of the cell cycle in BALB/c murine fibroblasts. Proc. Natl. Acad. Sci. U S A. 1992;89:3889–93. doi: 10.1073/pnas.89.9.3889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKearin D, Ohlstein B. A role for the Drosophila bag-of-marbles protein in the differentiation of cystoblasts from germline stem cells. Development. 1995;121:2937–47. doi: 10.1242/dev.121.9.2937. [DOI] [PubMed] [Google Scholar]
- Moroy T, Geisen C. Cyclin E. Int. J. Biochem. Cell Biol. 2004;36:1424–39. doi: 10.1016/j.biocel.2003.12.005. [DOI] [PubMed] [Google Scholar]
- Narbonne P, Roy R. Inhibition of germline proliferation during C. elegans dauer development requires PTEN, LKB1 and AMPK signalling. Development. 2006a;133:611–9. doi: 10.1242/dev.02232. [DOI] [PubMed] [Google Scholar]
- Narbonne P, Roy R. Regulation of germline stem cell proliferation downstream of nutrient sensing. Cell Div. 2006b;1:29. doi: 10.1186/1747-1028-1-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nasmyth K, et al. Some facts and thoughts on cell cycle control in yeast. Cold Spring Harb. Symp. Quant. Biol. 1991;56:9–20. doi: 10.1101/sqb.1991.056.01.004. [DOI] [PubMed] [Google Scholar]
- Ohtsubo M, Roberts JM. Cyclin-dependent regulation of G1 in mammalian fibroblasts. Science. 1993;259:1908–12. doi: 10.1126/science.8384376. [DOI] [PubMed] [Google Scholar]
- Oldham S, Hafen E. Insulin/IGF and target of rapamycin signaling: a TOR de force in growth control. Trends Cell Biol. 2003;13:79–85. doi: 10.1016/s0962-8924(02)00042-9. [DOI] [PubMed] [Google Scholar]
- Oldham S, et al. The Drosophila insulin/IGF receptor controls growth and size by modulating PtdInsP(3) levels. Development. 2002;129:4103–9. doi: 10.1242/dev.129.17.4103. [DOI] [PubMed] [Google Scholar]
- Orme MH, et al. Input from Ras is required for maximal PI(3)K signalling in Drosophila. Nat. Cell Biol. 2006;8:1298–302. doi: 10.1038/ncb1493. [DOI] [PubMed] [Google Scholar]
- Potten CS, Loeffler M. Stem cells: attributes, cycles, spirals, pitfalls and uncertainties. Lessons for and from the crypt. Development. 1990;110:1001–20. doi: 10.1242/dev.110.4.1001. [DOI] [PubMed] [Google Scholar]
- Prober DA, Edgar BA. Interactions between Ras1, dMyc, and dPI3K signaling in the developing Drosophila wing. Genes Dev. 2002;16:2286–99. doi: 10.1101/gad.991102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puig O, et al. Control of cell number by Drosophila FOXO: downstream and feedback regulation of the insulin receptor pathway. Genes Dev. 2003;17:2006–20. doi: 10.1101/gad.1098703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puig O, Tjian R. Nutrient availability and growth: regulation of insulin signaling by dFOXO/FOXO1. Cell Cycle. 2006;5:503–5. doi: 10.4161/cc.5.5.2501. [DOI] [PubMed] [Google Scholar]
- Richardson H, et al. Ectopic cyclin E expression induces premature entry into S phase and disrupts pattern formation in the Drosophila eye imaginal disc. Development. 1995;121:3371–9. doi: 10.1242/dev.121.10.3371. [DOI] [PubMed] [Google Scholar]
- Rodriguez-Viciana P, et al. Phosphatidylinositol 3′ kinase: one of the effectors of Ras. Philos Trans. R. Soc. Lond. B Biol. Sci. 1996;351:225–32. doi: 10.1098/rstb.1996.0020. [DOI] [PubMed] [Google Scholar]
- Savatier P, et al. Contrasting patterns of retinoblastoma protein expression in mouse embryonic stem cells and embryonic fibroblasts. Oncogene. 1994;9:809–18. [PubMed] [Google Scholar]
- Snapp EL, et al. The fusome mediates intercellular endoplasmic reticulum connectivity in Drosophila ovarian cysts. Mol. Biol. Cell. 2004;15:4512–21. doi: 10.1091/mbc.E04-06-0475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spradling A, et al. Stem cells find their niche. Nature. 2001;414:98–104. doi: 10.1038/35102160. [DOI] [PubMed] [Google Scholar]
- Spradling AC. Developmental Genetics of Oogenesis. In: Bate M, Arias AM, editors. The Development of Drosophila melanogaster. Vol. 1. Cold Spring Harbor Laboratory Press; Plainview, NY: 1993. pp. 1–70. [Google Scholar]
- Stead E, et al. Pluripotent cell division cycles are driven by ectopic Cdk2, cyclin A/E and E2F activities. Oncogene. 2002;21:8320–33. doi: 10.1038/sj.onc.1206015. [DOI] [PubMed] [Google Scholar]
- Tanaka S, et al. Neoplastic transformation induced by insulin receptor substrate-1 overexpression requires an interaction with both Grb2 and Syp signaling molecules. J. Biol. Chem. 1996;271:14610–6. doi: 10.1074/jbc.271.24.14610. [DOI] [PubMed] [Google Scholar]
- Uauy R, Solomons N. Diet, nutrition, and the life-course approach to cancer prevention. J. Nutr. 2005;135:2934S–2945S. doi: 10.1093/jn/135.12.2934S. [DOI] [PubMed] [Google Scholar]
- Voskuil DW, et al. The insulin-like growth factor system in cancer prevention: potential of dietary intervention strategies. Cancer Epidemiol. Biomarkers Prev. 2005;14:195–203. [PubMed] [Google Scholar]
- Walter SA, et al. Activation of Wee1 by p42 MAPK in vitro and in cycling xenopus egg extracts. Mol Biol Cell. 2000;11:887–96. doi: 10.1091/mbc.11.3.887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Z, Lin H. The division of Drosophila germline stem cells and their precursors requires a specific cyclin. Curr. Biol. 2005;15:328–33. doi: 10.1016/j.cub.2005.02.016. [DOI] [PubMed] [Google Scholar]
- Weinkove D, et al. Regulation of imaginal disc cell size, cell number and organ size by Drosophila class I(A) phosphoinositide 3-kinase and its adaptor. Curr. Biol. 1999;9:1019–29. doi: 10.1016/s0960-9822(99)80450-3. [DOI] [PubMed] [Google Scholar]
- Weissman IL. Stem cells: units of development, units of regeneration, and units in evolution. Cell. 2000;100:157–68. doi: 10.1016/s0092-8674(00)81692-x. [DOI] [PubMed] [Google Scholar]
- Wong MD, et al. Molecular mechanisms of germline stem cell regulation. Annu. Rev. Genet. 2005;39:173–95. doi: 10.1146/annurev.genet.39.073003.105855. [DOI] [PubMed] [Google Scholar]
- Wu MY, et al. Insulin delays the progression of Drosophila cells through G2/M by activating the dTOR/dRaptor complex. Embo J. 2007;26:371–9. doi: 10.1038/sj.emboj.7601487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie T, Spradling AC. decapentaplegic is essential for the maintenance and division of germline stem cells in the Drosophila ovary. Cell. 1998;94:251–60. doi: 10.1016/s0092-8674(00)81424-5. [DOI] [PubMed] [Google Scholar]
- Yan YP, et al. Insulin-like growth factor-1 is an endogenous mediator of focal ischemia-induced neural progenitor proliferation. Eur. J. Neurosci. 2006;24:45–54. doi: 10.1111/j.1460-9568.2006.04872.x. [DOI] [PubMed] [Google Scholar]
- Yenush L, et al. The Drosophila insulin receptor activates multiple signaling pathways but requires insulin receptor substrate proteins for DNA synthesis. Mol. Cell. Biol. 1996;16:2509–17. doi: 10.1128/mcb.16.5.2509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zagon IS, et al. Use of topical insulin to normalize corneal epithelial healing in diabetes mellitus. Arch. Ophthalmol. 2007;125:1082–8. doi: 10.1001/archopht.125.8.1082. [DOI] [PubMed] [Google Scholar]
- Zhang CC, Lodish HF. Insulin-like growth factor 2 expressed in a novel fetal liver cell population is a growth factor for hematopoietic stem cells. Blood. 2004;103:2513–21. doi: 10.1182/blood-2003-08-2955. [DOI] [PubMed] [Google Scholar]
- Zhang Y, et al. Drosophila target of rapamycin kinase functions as a multimer. Genetics. 2006;172:355–62. doi: 10.1534/genetics.105.051979. [DOI] [PMC free article] [PubMed] [Google Scholar]