

Abstract
Although humanity depends on the continued, aggregate functioning of natural ecosystems, few studies have explored the impact of community structure on the stability of aggregate community properties. Here we derive the stability of the aggregate property of community biomass as a function of species’ competition coefficients for a two-species model. The model predicts that the stability of community biomass is relatively independent of the magnitude of the interaction strengths. Instead, the degree of asymmetry of the interactions appears to be key to community stability.
With the increasing pressure from humans on natural ecosystems, there is growing interest in what makes ecological systems stable (1, 2). Researchers have focused on the fluctuations in aggregate community variables, such as total biomass (3-5), nutrient retention (6), and CO2 flux (7). Here we construct a two-species theoretical model to examine how species interaction strength affects the variance of community biomass over time.
We define community biomass (B) as the sum of the number of individuals (N) of all species and refer to the variance in B as the degree of “biomass stability.” Low variance of B indicates high biomass stability. Similarly, we refer to the variance of individual species’ abundances (Ni) (calculated in the presence of the other species) as the degree of “population stability.” For a two-species community, community biomass remains constant along “biomass isoclines,” or the contours where N1 + N2 = B (Fig. 1). Variation in total biomass occurs perpendicular to the isoclines (in the direction of the line N1 − N2 = 0). In contrast, population variation takes place parallel to the axes of each species.
Figure 1.

Biomass isoclines for a two-species community. N1 and N2 are the number of individuals of species 1 and 2, respectively. Total community biomass is constant along any line where N1 + N2 equals a constant, B. Total biomass varies in the direction shown by the arrows, perpendicular to the biomass isoclines.
The model is derived from traditional density-dependent, Lotka-Volterra competition equations. Fig. 2 illustrates a simulation of the model. A two-species community begins at an equilibrium point where both species have positive abundances. At every time step, the abundances of each species are perturbed independently. The dots in Fig. 2 represent the population sizes of the species at each time step. The spread of the points perpendicular to the biomass isoclines corresponds to community stability. The spread of the points parallel to the axes corresponds to the stability of the individual species’ populations. Thus, the community in Fig. 2a is more stable than the community in Fig. 2b, both in terms of biomass stability and population stability.
Figure 2.

Simulations of the model where the abundance of each species is perturbed at each time step (σz2 = 8). The curves show the trajectories of the nonlinear model without perturbations (Eq. 1). The dots represent the abundances of both species at 200 time steps. The community in (a) (r0 = 1) is more stable than (b) (r0 = 0.3) in terms of both biomass and population stability.
We use the model to consider how interspecific competition affects biomass and population stability. The variance of species’ populations and the variance of community biomass is derived in terms of interaction strengths, intrinsic growth rates, and the variance of the stochastic fluctuations.
Before proceeding, two points should be clarified. First, there are many meanings of interaction strength in the ecological literature (8). In this paper, interaction strength refers to the Lotka-Volterra competition coefficient, α. This coefficient represents the intensity of per-capita interspecific competition relative to per-capita intraspecific competition (9).
Second, just as there are many meanings of stability, there are a variety of ways to measure each type of stability. Here we are examining the resilience of a system, or how fast a variable (biomass) returns to its initial value after a perturbation (10). Empirical studies measuring resilience have measured variance (5, 7), coefficient of variation (CV) (3, 4), and the proportional return to an initial value after a given time (4). Although we use variance to measure population and biomass stability, we also show how our results change if the variation is standardized by the mean values (that is, if the CV is used).
The Model
For two species, the density-dependent, Lotka-Volterra competition equations in discrete time are
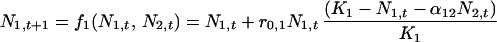 |
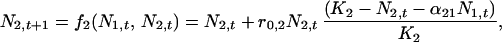 |
1 |
where for species i, Ni,t is the population abundance at time t, r0,i is the intrinsic rate of increase, Ki is the carrying capacity, and αij is the effect of species j on species i. By definition, an equilibrium, (N̂1, N̂2), satisfies the two equations simultaneously
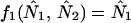 |
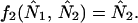 |
2 |
We introduce the variables n1,t and n2,t which describe the amount that N1 and N2 deviate from the equilibrium values at time t (11). Now we can write
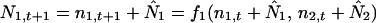 |
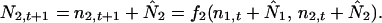 |
3 |
Performing a Taylor expansion of f1 and f2 about (N̂1, N̂2) and dropping the higher-order terms yields the linear model in discrete time
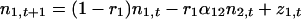 |
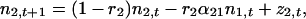 |
4 |
where ni,t is the deviation at time t of the population size of species i from its long-term average value. Stochastic perturbations (zi) are added to the population abundance of species i and are independently identically distributed random variables with mean zero and variance σz2. (For simplification, we assume that the perturbations of both species have the same distributions.) Here ri are the effective rates of increase of the species and equal r0N̂i/Ki (11), or
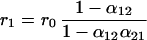 |
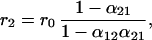 |
5 |
where r0 is the intrinsic growth rate, which, for simplicity, is the same for both species.
In the following steps, we derive the variance of n1, n2, and n1 + n2 (equivalent to the variance of N1, N2, and N1 + N2). In matrix form Eq. 4 is
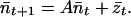 |
6 |
Let A represent the matrix
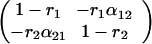 |
, and n̄t+1, n̄t and z̄t denote the vectors
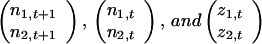 |
, respectively.
The eigenvectors of A describe the direction of the species’ response to perturbations from the equilibrium, and the eigenvalues describe the strength of the response in these directions. Below we arrange Eq. 6 so that we can derive the variance of the populations and total biomass in terms of the community’s eigenvectors and eigenvalues and the variance of the stochastic perturbations.
Let the matrix T be a matrix of the unit length eigenvectors (ē1 and ē2) of A.
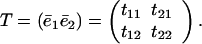 |
7 |
Eq. 6 can be rewritten as
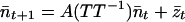 |
8 |
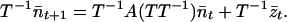 |
9 |
Because A has distinct eigenvalues and its eigenvectors are linearly dependent, A is equal to TDT−1, where D is a matrix with the eigenvalues of A as the diagonal entries and zeros elsewhere. Hence, D = T−1AT and Eq. 9 becomes
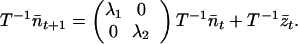 |
10 |
When |λi| < 1, then the equilibrium is stable.
The multiplication of each term in the above equation by T−1 transforms the vectors to a new basis along the inverse eigenvectors. We represent this change of basis by an asterisk:
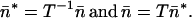 |
11 |
Eq. 10 now can be written
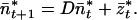 |
12 |
Considering the elements of the column vectors separately,
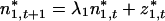 |
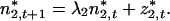 |
13 |
These equations are equivalent to Eq. 4, but written in terms of the eigenvectors and eigenvalues of A. Eq. 13 is a “first-order autoregressive process” and can be solved by successive substitution:
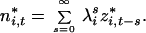 |
14 |
Then, the variance of the series is
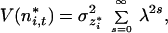 |
15 |
where σz*i2 is the variance of the perturbations of species i in the new coordinates (12). When |λi| < 1, Eq. 15 simplifies to
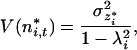 |
16 |
which does not depend on t. This equation gives the variance of a species’ population along the inverse eigenvectors when adding a perturbation z at every time step, where the perturbations have a normal distribution with mean 0 and variance σz2. The variance of σz*i2 can be written in terms of σz2. As defined in Eq. 11, z̄* = T−1z̄, or
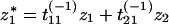 |
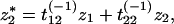 |
17 |
where tij(−1) represents the terms of the matrix T−1. Then,
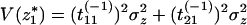 |
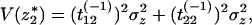 |
18 |
because the perturbations z1 and z2 are independent, that is, Cov(z1, z2) = 0.
The variance of the species’ deviations in the original coordinates is similarly related to the deviations in the new basis given in Eq. 16:
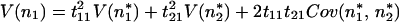 |
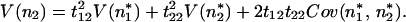 |
19 |
In the same manner,
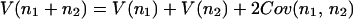 |
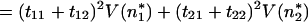 |
20 |
 |
The covariance of n*1 and n*2 can also be expressed in terms of the eigenvectors and eigenvalues of A (see Appendix):
 |
21 |
The CV of population abundance is simply the SD divided by the equilibrium value (the mean abundance), or
 |
 |
22 |
Similarly, the CV of community biomass is the SD of the sum of the species divided by the sum of species’ equilibrium values, or
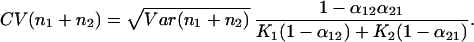 |
23 |
Results
We are concerned with the behavior of a community when it is perturbed from a stable equilibrium where all species have positive values (an interior equilibrium). An equilibrium is stable when the absolute value of the eigenvalues of the community matrix (A) is less than one and assures that when a species is moved from the equilibrium it eventually will return (i.e., the perturbation goes to zero). Here we explore a range of parameter values where the interaction coefficients and the intrinsic growth rate satisfy the conditions for a stable interior equilibrium (0 < α12 < 1, 0 < α21 < 1, 0 < r0 < 2).
The Symmetric Case.
For a two-species community with symmetric competition (where α12 = α21 = α), the eigenvalues of the matrix A (Eq. 6) are
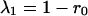 |
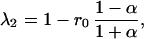 |
24 |
and the matrix of unit eigenvectors is
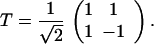 |
25 |
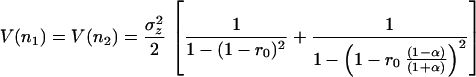 |
26 |
and
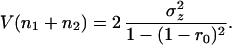 |
27 |
Fig. 3a plots Eqs. 26 and 27 as a function of the species’ interaction strength (α) for particular values of r0 and σz2. Population stability decreases (the variance increases) as interactions become stronger, with a sharp decline in stability as α approaches 1. Biomass stability does not depend on interaction strength, however. When α is zero—that is, the species are independent—the variance of the sum of the species simply equals the sum of the variances of both species individually. If interactions increase, biomass variance remains equivalent to two independent species, and eventually the population variance of the competing species surpasses the biomass variance. Simulations reveal that the linear model is a good approximation of the original Lotka-Volterra equations (Fig. 3a).
Figure 3.

The symmetric case σz2 = 5; r0 = 1). (a) The variance of n1 + n2 (solid line) and n1 (dashed line), as a function of the α coefficient. Calculations of the variance from a simulation of the original Lotka-Volterra model agree with the results of the linear approximation [each point is variance after 1,000 time steps; variance of N1 + N2 (o), variance of N1 (x)]. (b) The CV of n1 + n2 (solid line) and n1 (dashed line), as a function of the α coefficient (K1 = K2 = 100).
The independence of biomass stability and species interaction strengths can be explained by the symmetric competition. The species affect one another’s populations in the same manner, so that the species exactly compensate for one another regardless of interaction strength. In contrast, an increase in interaction strength increases the variance of each population, because a species responds not only to its own perturbations, but increasingly to the perturbations of the other species.
Because the equilibrium values of species’ abundances are dependent on α, the CV of the individual populations and the CV of the total biomass are functions of the species’ interaction strength (Fig. 3b). Fig. 3b illustrates another qualitative change: the stability (measured by the CV) of total biomass is greater than that of individual populations for all interaction strengths between 0 and 1. Using CV as a measure of stability, however, does not alter the result that biomass stability is much less influenced by interaction strength compared to population stability.
For any pair of symmetric competition coefficients between 0 and 1, the variance of n1 + n2 is lowest when the species’ intrinsic growth rates (r0) are 1, and increases to infinity as r0 approaches 0 or 2 (Fig. 4). When r0 = 1, the species returns to the equilibrium biomass in the next time step. In this case, the biomass variance is simply the variance of the environmental perturbations (σz2). If the growth rate falls below 1, the species are slow to respond to perturbations, and the biomass variance increases. Similarly, if the growth rate is above 1, the species overcompensate for perturbations and again, the biomass variance increases.
Figure 4.

The variance of total biomass, n1 + n2, as a function of the intrinsic rate of increase, r0 (α = 0.5; σz2 = 1).
The variance of the environmental perturbations (σz2) simply scales both the variance of the biomass and the individual populations. For instance, a doubling of the variance of the environmental shocks decreases both biomass stability and population stability by one-half.
An explanation of the eigenvectors and eigenvalues of the community matrix, A, may help to clarify further Eqs. 26 and 27. The eigenvectors of A describe the directions of the species’ response to perturbations, and the eigenvalues describe the strength of the species’ response along these directions. In the symmetric case, the first eigenvector is perpendicular to and the second is parallel to the biomass isoclines. Because biomass stability is measured as variation perpendicular to the biomass isoclines, only changes along the first eigenvector matter. The strength of the response along this eigenvector (the first eigenvalue) does not depend on α thus, biomass stability is independent of species’ interaction strength.
Conversely, components of both eigenvectors affect population stability, because the eigenvectors are not parallel to either species’ axis. Therefore, both eigenvalues (including the second eigenvalue that depends on α) appear in Eq. 26.
The Asymmetric Case.
The general two-species case, where species interactions may be asymmetrical (α12 ≠ α21), is given in the Appendix. In this section, we describe the solution with figures and a special case. The dominant trend is that biomass stability is highest when the interaction coefficients of the two species are equal and decreases as the difference between α12 and α21 increases (Fig. 5a). In other words, as the competitive abilities of the species become more disparate, the response to the perturbations by each species becomes more unequal and biomass stability decreases.
Figure 5.

The asymmetric case. The variance of (a) n1 + n2 and (b) n1 as a function of α12 and α21. The CV of (c) n1 + n2 and (d) n1 as a function of α12 and α21 (K1 = K2 = 100). In all cases, σz2 = 1 and r0 = 1.
This pattern holds when r0 is not high (<∼1.5), and is strongest for species with low intrinsic growth rates. When r0 is high (>∼1.5), there are asymmetric pairs of alphas that yield a lower biomass variance than symmetric pairs.
Just as in the symmetric case, the larger the interaction coefficients, the lower the population stability. The results are complicated by the intrinsic growth rates of the species, but the trends are robust for most values. In general, the variance of species 1 increases as the effect of species 2 on 1 (α12) increases (Fig. 5b). Similarly, increasing the strength of the effect of species 1 on 2 (α21) increases the variance of species 1, particularly when the intrinsic growth rates are high.
The main results do not change when measuring stability as the CV rather than variance. The CV of total biomass generally increases as the interaction strengths become more different (Fig. 5c), and the CV of species 1 increases as α12 increases (Fig. 5d). One trend does change: the CV of species 1 decreases as α21 increases (Fig. 5d). The increased abundance of species 1 at high values of α21 outweighs the increase in variance because of stronger interactions. Finally, as in the symmetric case, standardizing the stability measure by the expected mean changes the relative degree of biomass stability versus population stability for some interaction strengths. For most combinations of alphas, the CV of total biomass is lower than the CV of individual populations (note the different scales in Fig. 5 c and d).
It is useful to examine the special case where α12 + α21 = 1 to interpret the complex relationship between the biomass variance and the three parameters (α12, α21, and r0). In this case, biomass variance reduces to
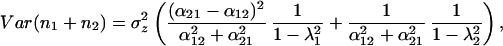 |
28 |
where
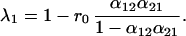 |
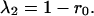 |
29 |
Substituting the constraint α12 + α21 = 1 into Eq. 5, the first eigenvalue (λ1) is equal to 1 − r1α12 or 1 − r2α21. The terms r1α12 and r2α21 also appear in Eq. 4, as the response of one species to the abundance of the other species. Hence, λ1 is inversely related to the strength of the response of one species to the other. Just as in the symmetric case, the second eigenvalue depends only on r0, the growth rate of the species by itself.
Eq. 28 now can be divided into two components, which we will refer to as the response term (including λ1, which depends on r1α12), and the self term (including λ2, which depends only on r0). In other words, the biomass variance is the sum of the response term and the self term multiplied by σz2. The first part of each term is a coefficient determined by the alphas. These coefficients reflect the direction of the eigenvectors and scale accordingly the contributions of eigenvalues to the biomass variance. As long as the interaction strengths are not equal, neither eigenvector is parallel to the biomass isoclines, and both eigenvalues contribute to the biomass variance.
Fig. 6 plots the response and self terms as functions of r0 and the difference between α12 and α21. For a given pair of alphas, how does r0 affect the two component’s contributions to biomass variance? An increase in r0 increases the response of a species to the other (r1α12 = r2α21), which corresponds to a decrease in the response term’s contribution to biomass variance. For the self term, r0 is related to biomass variance in the same manner as the symmetric case: the minimum biomass variance occurs when r0 = 1 and increases as r0 goes to 0 and 2.
Figure 6.

The contribution of the response and self terms (see text) to the variance of community biomass [Var(n1 + n2)] as functions of the intrinsic growth rate (r0) and the interaction coefficient (α12) where α12 + α21 = 1.
Conversely, for a given growth rate, how does the disparity of the alphas affect the component’s contributions to biomass variance? The response term changes in two ways as the interaction strengths become more different: (i) the response of one species to the other (r1α12) decreases, and (ii) the first eigenvector becomes more perpendicular to the biomass isoclines. Both factors lead to an increase in overall biomass variance. In contrast, the self term decreases as the interaction strengths become more unequal, because the direction of the second eigenvector becomes more parallel to the biomass isoclines.
The relationships described above and depicted in Fig. 6 explain why, at high intrinsic growth rates, the relationship between the interaction coefficients and biomass stability changes from the simple form in Fig. 5a. When r0 is very high, the increased ability of the species to respond to one another makes up for some of the variation in biomass caused by unequal interaction strengths.
Discussion
The model suggests that the predictability of an aggregate community property may be relatively independent of the strength of the species’ interactions. Evenness of the species competitive abilities appears to be key to community stability. This result conflicts with the idea that more tightly linked communities may be more stable (13, 14). Using a similar analysis, Ives (15, 16) studied the resilience of population abundances in stochastic systems. He also concluded that the greater the similarity (in terms of competitive abilities) of two species, the greater the buffering of densities against stochastic fluctuations.
The model also highlights that the stability of individual species’ populations may not necessarily be related to overall community stability, as others have pointed out (4). Although the magnitude of species competition is largely independent of biomass stability, strong interactions sharply decreased population stability.
The different relationships between population and community stability and interaction strength predict that as the intensity of competition increases, population variation will become increasingly greater than biomass variation (both in terms of variance and the CV). In a grassland ecosystem, Tilman (4) found that plant species’ abundances fluctuated more than total plant biomass over an 11-year period and, in fact, these plant species appear to be competing strongly. Reexamining other published studies may help to test this hypothesis.
The “point of entry” of random fluctuations can influence the results of stochasticity on population growth models (e.g., refs. 17-19). Here random fluctuations are incorporated as direct perturbations to abundances. Random temporal variation also can be added to parameters such as r, K, and α. The consequences of this type of stochasticity should be explored for this model.
We used a linear model hoping to capture the same biological spirit of the Lotka-Volterra equations. The variance of numerical simulations of the original Lotka-Volterra equations closely agree with the prediction of the linear model. This result coincides with previous analyses where stochastic fluctuations are added to the carrying capacity (20). Although the fit of the linear model does decrease as the variance of the perturbations (σz2) increases, the chance of species extinction increases at the same time. Thus, before the approximation becomes problematic, the probability of species extinction becomes consequential.
The technique used in this paper to model the predictability of an aggregate community variable is generalizable for other indexes. For example, total nutrient retention (R) of a plant community might be modeled as R = aN1 + bN2 + … + xNn, where Ni is the abundance of each of the n species. The contours of the index then could be compared to the direction of the eigenvectors. The model also could be used to address the influence of species diversity on the predictability of community variables. As human beings continue to alter the planet’s ecosystems, understanding the relationship between community structure and the maintenance of ecosystem functioning will become increasingly valuable in managing and preserving essential ecosystem services (21).
Acknowledgments
We thank Sean Connolly, Mark Tanaka, and two anonymous reviewers for valuable comments. Support for J.B.H. was provided by the Koret Foundation and the generosity of Helen and Peter Bing. J.R. received support from the Stanford University Research Incentive Fund.
ABBREVIATION
- CV
coefficient of variation
Appendix
Derivation of Eq. 21.
By the definition of covariance,
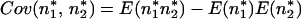 |
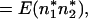 |
because E(n*i) = 0. As in Eq. 14, n*i can be solved by successive substitution, so
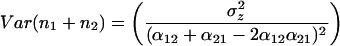 |
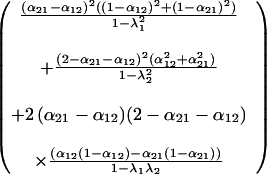 |
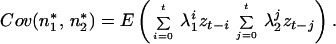 |
Where i ≠ j, the terms are zero, and the equation above becomes
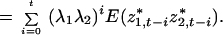 |
As t → ∞, this becomes
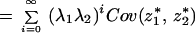 |
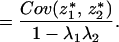 |
Because E(z*1) = E(z*2) = 0, the above equals
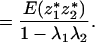 |
Substituting Eq. 17 for z*1 and z*2,
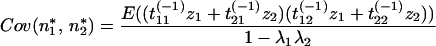 |
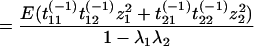 |
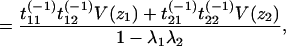 |
because E(z1z2) = 0 and E(z1) = E(z2) = 0.
The Asymmetric Case.
The variances of the species’ populations are
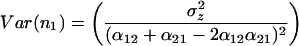 |
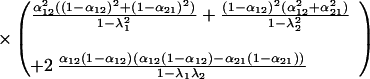 |
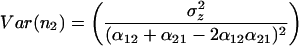 |
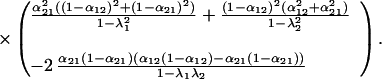 |
The variance of community biomass is
where
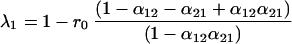 |
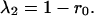 |
References
- 1.Pimm S L. The Balance of Nature? Chicago: Univ. of Chicago; 1991. [Google Scholar]
- 2.Schulze E-D, Mooney H A. Biodiversity and Ecosystem Function. Berlin: Springer; 1994. [Google Scholar]
- 3.McNaughton S J. Am Naturalist. 1977;111:515–525. [Google Scholar]
- 4.Tilman D. Ecology. 1996;77:350–363. [Google Scholar]
- 5.Naeem S, Shibin L. Nature (London) 1997;390:507–509. [Google Scholar]
- 6.Ewel J J, Mazzarino M J, Berish C W. Ecol Appl. 1991;1:289–302. doi: 10.2307/1941758. [DOI] [PubMed] [Google Scholar]
- 7.McGrady-Steed J, Harris P M, Morin P J. Nature (London) 1997;390:162–165. [Google Scholar]
- 8.Laska M S, Wootton J T. Ecology. 1998;79:461–476. [Google Scholar]
- 9.MacArthur R. In: Growth by Intussusception. Deevey E S, editor. New Haven, CT: Connecticut Academy of Arts and Sciences; 1972. pp. 177–188. [Google Scholar]
- 10.Pimm S L. Nature (London) 1984;307:321–326. [Google Scholar]
- 11.Roughgarden J. Theory of Population Genetics and Evolutionary Ecology. Upper Saddle River, NJ: Simon & Schuster; 1979. [Google Scholar]
- 12.Cox D R, Miller H D. The Theory of Stochastic Processes. New York: Wiley; 1968. [Google Scholar]
- 13.Elton C S. The Ecology of Invasions by Animals and Plants. London: Chapman & Hall; 1958. [Google Scholar]
- 14.Odum H. Fundamentals of Ecology. Philadelphia: Saunders; 1971. [Google Scholar]
- 15.Ives A R. Ecol Monogr. 1995;65:217–233. [Google Scholar]
- 16.Ives A R. Ecology. 1995;76:926–941. [Google Scholar]
- 17.May R M. Stability and Complexity in Model Ecosystems. Princeton: Princeton Univ.; 1973. [PubMed] [Google Scholar]
- 18.Levins R. Proc Natl Acad Sci USA. 1969;62:1061–1065. doi: 10.1073/pnas.62.4.1061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Roughgarden J. Theor Popul Biol. 1975;7:1–12. doi: 10.1016/0040-5809(75)90002-7. [DOI] [PubMed] [Google Scholar]
- 20.Roughgarden J. Am Naturalist. 1975;109:713–736. [Google Scholar]
- 21.Daily G C. Nature’s Services: Societal Dependence on Natural Ecosystems. Washington, DC: Island; 1997. [Google Scholar]