

Abstract
Telomeres, consisting of nucleotide repeats and a protein complex at chromosome ends, are essential in maintaining chromosomal integrity. Dyskeratosis congenita (DC) is the inherited bone marrow failure syndrome (IBMFS) that epitomizes the effects of abnormal telomere biology. Patients with DC have extremely short telomere lengths (<1st percentile) and many have mutations in telomere biology genes. Interpretation of telomere length in other IBMFSs is less straightforward. Abnormal telomere shortening has been reported in patients with apparently acquired hematologic disorders, including aplastic anemia, myeolodysplasia, paroxysmal nocturnal hemoglobinuria, and leukemia. In these disorders, the shortest lived cells have the shortest telomeres, suggestive of increased hematopoietic stress. Telomeres are also markers of replicative and/or oxidative stress in other complex disease pathways, such as inflammation, stress, and carcinogenesis.
The spectrum of related disorders caused by mutations in telomere biology genes extends beyond classical DC to include marrow failure that does not respond to immunosuppression, idiopathic pulmonary fibrosis, and possibly other syndromes. We suggest that such patients are categorized as having an inherited disorder of telomere biology. Longitudinal studies of patients with very short telomeres but without classical DC are necessary to further understand the long-term sequelae, such as malignancy, osteonecrosis/osteoporosis, and pulmonary and liver disease.
BONE MARROW FAILURE
Bone marrow failure (BMF), also referred to as aplastic anemia (AA), is the result of the inability of the bone marrow to produce an adequate number of circulating blood cells (white blood cells [WBCs], red blood cells and/or platelets). Varying degrees of peripheral blood leukopenia, anemia, and thrombocytopenia are often present. The severity of BMF ranges from incidental findings on routine blood counts to life-threatening cytopenias. BMF is typically classified as inherited or acquired. Inherited bone marrow failure syndromes (IBMFS), such as Fanconi anemia (FA) and dyskeratosis congenita (DC), usually come to clinical attention in childhood, while acquired AAs are more common in adulthood. However, it is important to recognize that inherited syndromes, especially those with variable phenotypes, can present at any age, as can acquired AA.
Telomere length and telomere dynamics have been shown to be abnormal in almost all patients with DC, and in some patients with an IBMFS other than DC, as well as in some cases of acquired AA or other bone marrow disorders. Making the distinction between an IBMFS and acquired AA is critical in order to provide proper counseling and treatment for individual patients. In addition, since IBMFS are also cancer predisposition syndromes proper surveillance for this increased risk of cancer is a necessary component of clinical care.
TELOMERES AND TELOMERE LENGTH
Telomeres consist of long TTAGGG nucleotide repeats and associated proteins at the ends of chromosomes that are essential for the maintenance of chromosomal integrity. In order to preserve the chromosome end, telomerase reverse transcriptase (gene name TERT), its RNA component (TERC) and an ordered protein complex, termed shelterin, protect the telomere from end-to-end fusion (Collins and Mitchell 2002; de Lange 2005; Nakamura and Cech 1998). Shelterin consists of six proteins (gene name, protein name abbreviations), telomeric repeat-binding factor 1 (TERF1, TRF1), terlomeric-repeat binding factor 2 (TERF2, TRF2), TRF1-interacting nuclear factor 2 (TINF2, TIN2), TERF2-interacting protein (TERF2IP, Rap1), TIN2-interacting protein 1 (ACD, protein names include TPP1, TINT1, PIP1, and PTOP) and protection of telomeres (POT1, POT1). Telomeric repeats are lost with each cell division, in part due to incomplete replication of the 3’ end of the chromosome. Telomeric attrition can result in critically short telomeres, prompting cellular senescence or cellular crisis, including apoptosis, genomic instability or a reduction in cellular proliferative potential (Maser and DePinho 2002; Shay et al. 2001).
Telomere shortening with age is very well described. It occurs most rapidly during infancy and subsequently slows but continues at approximately 30 to 40 base pairs per year (Frenck, Jr. et al. 1998; Rufer et al. 1999; Slagboom et al. 1994; Zeichner et al. 1999). Intra-individual telomere lengths appear to be consistent between tissues including cerebral cortex, myocardium, liver, renal cortex, and liver (Takubo et al. 2002) as well as between whole blood WBCs, umbilical artery and foreskin specimens (Okuda et al. 2002). There does appear to be more intra-individual variation in telomere length within bone marrow-derived cells: specifically intra-individual granulocyte telomere lengths in subjects with BMF are often shorter than those of other WBC subsets (Alter et al. 2007a; Ball et al. 1998; Brummendorf et al. 2001c; Lee et al. 2001; Polychronopoulou and Koutroumba 2004; Rufer et al. 1998; Weng et al. 1998).
Numerous methods are available to determine telomere length in blood and other cell types, and have been reviewed elsewhere (Baird 2005; Lin and Yan 2005). Telomere length in genomic DNA is often determined by terminal restriction fragment (TRF) measurement on Southern blots, or by quantitative polymerase chain reaction (QPCR). Fluorescence in situ hybridization (FISH) and immunostaining (telomere/immunostaining-FISH [TELI-FISH] and single telomere length analysis [STELA]) are used on tissue specimens and for individual chromosome measurements. FISH combined with flow-cytometry (flow-FISH) uses fresh blood samples and has the advantage of providing telomere length data on specific white blood cell subsets (Baerlocher and Lansdorp 2003). The method of telomere length measurement, cell type studied, as well as study design, must be considered in data interpretation and clinical application.
Until recently, most studies of BMF, inherited or acquired, used TRF to determine telomere lengths in whole blood WBCs or mononuclear cells and reported either mean telomere length in patients compared with age-matched controls, or the difference between the average telomere lengths of patients and controls (deltaTEL). The focus of those reports was the average telomere length in groups of patients compared with groups of controls, rather than in specific individuals whose telomeres were abnormally short. Refinements in telomere length measurements and advances in understanding of telomere biology and clinical phenotype are progressing rapidly. Large case-control studies of telomere length and disease association are being conducted to evaluate telomere length in groups of cases with telomere length in groups of age-matched control subjects. In order to apply this growing literature to the individual patient, interpretation of telomere length data must consider the presence of age-appropriate controls, the method of telomere length measurement, the cell type studied, as well as the definition of short telomeres used in the study.
TELOMERE LENGTH INHERITANCE AND GENETICS
Numerous studies suggest that telomere length is a heritable trait, despite the high degree of inter-individual variation. A study of monozygotic and dyzygotic twin pairs which measured telomere length in DNA isolated from whole blood WBCs showed high levels of concordance between twin pairs and telomere length, with an estimated heritability of 78% (Slagboom et al. 1994); this was subsequently confirmed (Jeanclos et al. 2000). Others have evaluated telomere length in female sibling pairs and parent-child trios and found varying degrees of telomere length heritability (Andrew et al. 2006; Bischoff et al. 2005; Graakjaer et al. 2006).
Identification of the genetic region and/or genes responsible for telomere length has proven difficult. X-linked inheritance of telomere length was suggested by analysis of multi-generational pedigrees and comparison of parent/child pairs (Nawrot et al. 2004). A quantitative-trait linkage analysis of 400 microsatellite markers in 258 sibling pairs showed significant linkage on chromosome 12, and identified a candidate gene in that region, DDX11, a DNA helicase (Vasa-Nicotera et al. 2005). Significant linkage at 14q23.2 and 3p26.1 was found in another quantitative-trait linkage analysis study of 1025 female dizygotic twin pairs (Andrew et al. 2006). Single nucleotide polymorphisms (SNPs), the most common germline genetic variants in the human genome, may also play a role in telomere length regulation. Individuals with one or two copies of the −1381 promoter T allele in the telomerase gene (TERT, T>C, rs2735940) had significantly longer telomeres and increased telomerase activity than in individuals with the CC genotype (Matsubara et al. 2006). These data suggest that polygenic inheritance of telomere length may occur.
DISORDERS WITH MUTATIONS IN TELOMERE BIOLOGY GENES
Dyskeratosis Congenita
DC is a clinically heterogeneous IBMFS that epitomizes the clinical consequences of very short telomeres and mutations in telomere biology (Walne et al. 2005; Yamaguchi 2007). The classical diagnosis of DC requires the triad of dysplastic nails, lacey reticular pigmentation of the upper chest and neck, and oral leukoplakia (Drachtman and Alter 1992). This diagnostic triad is still important in defining clinically significant disease, but as more patients with DC and their family members have been studied, an extremely broad clinical phenotype is now appreciated. Additional features include epiphora, blepharitis, premature gray hair, alopecia, developmental delay, short stature, cerebellar hypoplasia, microcephaly, esophageal stenosis, urethral stenosis, pulmonary fibrosis, liver disease, avascular necrosis of hips or shoulders, epithelial cancers, myelodysplastic syndrome (MDS), and leukemia (Alter 2005; Walne et al. 2005; Yamaguchi 2007). Hoyeraal-Hreidarsson (HH) Syndrome is a severe form of DC with BMF, immunodeficiency, microcephaly, cerebellar hypoplasia, intrauterine growth retardation, and developmental delay (Sznajer et al. 2003; Vulliamy et al. 2006). Revesz Syndrome, characterized by bilateral exudative retinopathy, bone marrow hypoplasia, nail dystrophy, fine hair, cerebellar hypoplasia, and growth retardation, also appears to be in the DC disease spectrum (Kajtar and Mehes 1994; Revesz et al. 1992).
Many cases of DC are not diagnosed until the development of cytopenias, but correct diagnosis has important clinical implications. Patients with DC and BMF do not respond to immunosuppressive therapy (Al Rahawan et al. 2006). They are at high risk of bone marrow transplantation-related complications due to underlying pulmonary and liver disease (Brazzola et al. 2005; de la Fuente and Dokal 2007; Dror et al. 2003; Ostronoff et al. 2007; Yabe et al. 1997). Selection of related stem cell donors may be complicated by the variable phenotype seen in DC, including silent carriers (Alter et al. 2007b). In addition, patients with DC must be carefully screened for cancers, such as squamous cell carcinomas of the head and neck or anogenital region, as well as hematologic malignancies.
Abnormalities in telomere biology are present in essentially all patients with DC (Marrone et al. 2005; Walne et al. 2005). These include abnormally short telomeres for age and reduced telomerase enzyme activity (Mitchell et al. 1999; Vulliamy et al. 2001). Mutations in dyskerin (DKC1) on the X chromosome were the first to be identified in patients with DC (Heiss et al. 1998). Subsequently mutations were found in TERC in families with autosomal dominant inheritance (Vulliamy et al. 2004). TERT mutations have been described in patients with DC and in patients initially classified as having acquired AA (Savage et al. 2006b; Vulliamy et al. 2005; Yamaguchi et al. 2005). Recessive mutations in NOP10 were recently discovered in one out of 16 consanguineous families with DC (Walne et al. 2007). In a 2006 review of the DC registry in the United Kingdom, it was estimated that a mutated gene is currently not identified in approximately 60% of patients with DC (Vulliamy et al. 2006).
The clinical heterogeneity and lack of molecular characterization in more than half of known patients with DC makes it a diagnostic challenge. We recently conducted a study of the utility of telomere length determination by flow-FISH as a diagnostic test of DC where very short telomeres were stringently defined as <1st percentile for age (Alter et al. 2007a). The normal range of telomere lengths was based on data from 400 healthy controls ranging in age from newborn (cord blood) to 100 years. The diagnostic sensitivity and specificity of very short telomeres was >90% in total lymphocytes, naïve T-cells, and B-cells for the diagnosis of DC in comparison with non-DC IBMFS patients, or with healthy relatives of patients with DC (Figure 1A). It is important to note that this study found that in many patients with non-DC IBMFS telomere lengths were also short in granulocytes; a similar result was reported in patients with acquired AA (Ball et al. 1998). This is most likely due to increased granulocyte turnover and short lifespan in comparison to other cell types and is discussed below.
Figure 1. Telomere lengths in inherited bone marrow failure syndromes.
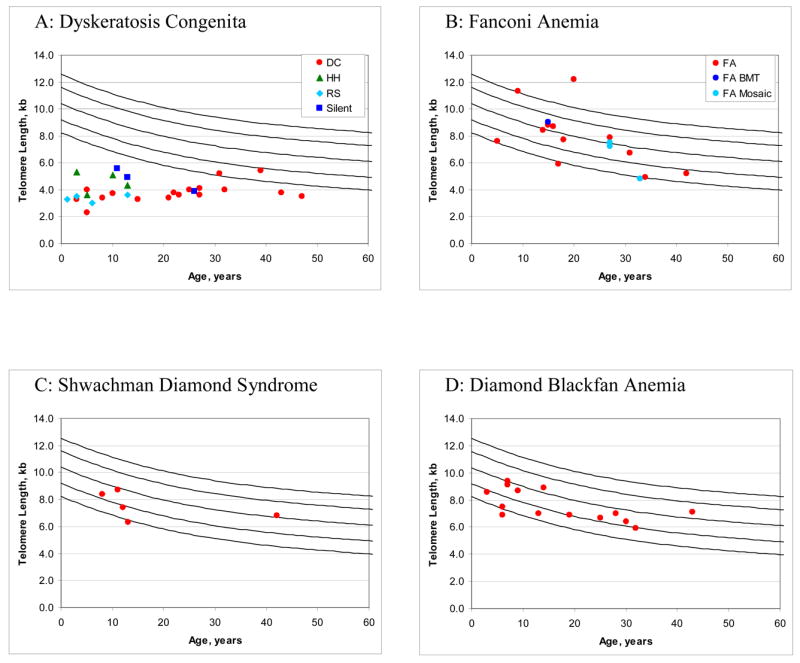
Telomere lengths in lymphocytes were determined by multi-color flow-FISH. Telomere lengths determined on 400 healthy subjects from newborn to 100 years of age were used to determine the normal range. The top line represents the 99th %ile and the bottom line represents the 1st %ile. This figure was derived from previously published data (Alter et al. 2007).
A: Telomere lengths in Dyskeratosis Congenita (DC). Red circles indicate patients with DC. Green triangles indicate the Hoyeraal-Hreidarsson Syndrome (HH) and light blue diamonds indicate the Revesz Syndrome (RS) variants of DC. Dark blue squares are silent carriers of gene mutations who remain clinically asymptomatic.
B: Telomere lengths in Fanconi Anemia (FA). Red circles indicate patients with FA. The dark blue circle is a patient with FA who underwent bone marrow transplantation. Mosaic FA patients are noted with the light blue circle.
C: Telomere lengths in Shwachman Diamond Syndrome.
D: Telomere lengths in Diamond Blackfan Anemia.
It is also important to note that in many previous studies of telomere length in DC deltaTELwas calculated by subtracting the patient’s value from that of an age-matched control. In normal individuals, as well as those with a non-DC inherited BMF, telomere length decreases with age in a nonlinear, S-shaped manner. However, telomere length does not appear to decrease with age in DC, and in some cell types even appears to increase slightly (Alter et al. 2007a). Thus deltaTEL will also decrease with age, and cannot be used to compare telomere lengths across ages. The lack of age-associated decline in telomere length in DC was not known at the time of the publications using deltaTEL.
We further confirmed the validity of very short telomere lengths as a sensitive and specific diagnostic test for DC in a genetic linkage scan of a large, non-consanguineous family with autosomal DC and no mutations in DKC1, TERC, or TERT (Savage et al. 2007). Using very short telomeres as the defining feature of DC instead of the heterogeneous clinical phenotypes, we identified novel mutations in TRF1-interacting nuclear factor 2 (TINF2, protein name TIN2) in all family members with telomere lengths <1st percentile but not in those with normal telomere lengths. TINF2 mutations were also present in four additional, unrelated patients with DC, but not in their unaffected relatives. The mutation prevalence in DC probands currently represented in our cohort are 31.2% TINF2, 18.8% DKC1, 18.8% TERC, 6.2% TERT, and 4 patients remain molecularly uncharacterized (25%). Larger studies are warranted to determine the true prevalence of TINF2 mutations in DC.
Acquired Aplastic Anemia
Acquired AA is a complex, heterogeneous disorder usually characterized by immune-mediated BMF. It can present at any age and is often associated with exposure to putative environmental factors, chemicals such as benzene, idiosyncratic reactions to drugs such as chloramphenicol, and certain infections (Young et al. 2006). Immune-mediated T-cell destruction of the bone marrow has been shown both in vitro and in vivo, but the reason for T-cell activation in these patients is not clear. A combination of genetic risk factors, including SNPs in tumor necrosis factor-alpha (Demeter et al. 2002; Peng et al. 2003) and gamma-interferon (Dufour et al. 2004) may be contributory.
Several studies have described telomere shortening in approximately one-third of individuals with acquired AA (Ball et al. 1998; Brummendorf et al. 2001a; Lee et al. 2001). These studies evaluated either TRF or flow-FISH telomere lengths in acquired AA patients in mononuclear cells and/or granulocytes. In general, TRF values were significantly lower in granulocytes than in mononuclear cells, a difference that may be a result of increased cell turnover in the former. Increased telomere loss (i.e. shorter telomere length) was also associated with longer duration of cytopenias. Telomerase activity was shown to be up-regulated in acquired AA, possibly to counteract telomere loss (Shen et al. 2002). Patients with acquired AA may have short telomeres because of the rapid turnover of hematopoietic progenitor cells in order to compensate for a failing bone marrow (Brummendorf et al. 2001a; Young 2006). Therefore, the specific cell type studied must be considered when interpreting these data. The presence of granulocytes in cell fractions may skew the data towards the appearance of abnormally short telomeres due to rapid cell turnover and short lifespan. Methods such as flow-FISH that determine telomere length in white blood cell subsets can separately evaluate longer lived cells such as lymphocytes; other methods to study telomere lengths of non-hematogenous cells may be better suited for such studies (Baerlocher et al. 2006).
Individuals with apparently acquired AA who did not respond to immunosuppression had the shortest telomeres in several studies (Ball et al. 1998; Brummendorf et al. 2001a; Lee et al. 2001). This is consistent with at least two different pathological mechanisms causing BMF, immune-mediated (acquired AA) and genetic (an IBMFS). The immune-mediated forms of BMF are likely to be caused by environmental exposures, such as infection and chemicals, and respond to immunosuppressive therapy. The forms of BMF associated with short telomeres and failure to respond to immunosuppressive therapy are most likely derived from different genetic and environmental risk factors. The role that these risk factors play in BMF associated with short telomeres may vary from highly penetrant, such as mutations in DKC1 or other genes causing DC, to single nucleotide polymorphisms that act as markers of disease risk (Tables 1 and 2). For example, single nucleotide polymorphisms in TERF1 (telomeric repeat binding factor 1) were associated with an increased risk of acquired AA (Savage et al. 2006a). In the middle of this spectrum are individuals with what was initially thought to be an acquired AA, because it occurred later in life, but who have mutations in TERC (Yamaguchi et al. 2003) or TERT (Yamaguchi et al. 2005). Semantics may now play a role: persons (and their relatives) with germline mutations in genes in the telomere maintenance pathway are at risk of BMF, MDS, leukemia, and specific solid tumors, and should be managed and counseled accordingly, independent of their clinical and laboratory phenotype.
Table 1.
Disorders with mutations in telomere biology genes.
| Specific Disorder | Major Clinical Findings* | Family History | Relative degree of telomere shortening | Mutations |
|---|---|---|---|---|
| Dyskeratosis Congenita | Diagnostic triad of dyskeratotic nails, lacey reticular pigmentation, and oral leukoplakia may not always be present.
BMF, pulmonary fibrosis, cancer predisposition, and a broad range of phenotypes may develop over time |
XLR, AD, AR, or de novo | Very Short, <1st percentile | DKC1, TERC, TERT, NOP10, TINF2 |
| Acquired Aplastic Anemia** | BMF of varying degrees, idiopathic or associated with an environmental exposure | +/− | Short to very short, <10th percentile | TERC, TERT TERF1 (SNP) |
| Idiopathic Pulmonary Fibrosis** | Pulmonary fibrosis progressing to interstitial pneumonia | +/− | Short to very short, <10th percentile | TERC, TERT |
Abbreviations: BMF, bone marrow failure, XLR, X-linked recessive, AD, autosomal dominant, AR, autosomal recessive, SNP, single nucleotide polymorphism.
Numerous, more specific clinical findings are described in the text. DC is the only disorder in which all patients have very short telomeres (< 1st percentile).
The authors of these studies used a more liberal cut-off for short telomeres, generally <10th percentile, than we have used to define DC.
Table 2.
Complex disorders associated with short telomeres.
| Disorder | Major Clinical Findings* | Inheritance / Family History | Usual Telomere Length | Notes |
|---|---|---|---|---|
| Shwachman- Diamond Syndrome | Malabsorption, short stature, metaphyseal dysostosis, neutropenia, MDS, leukemia, SBDS gene mutations | AR | Normal for age, >1st percentile | |
| Diamond-Blackfan Anemia | Abnormal thumbs, short stature, reticulocytopenia, cancer predisposition, RPS17, RPS19, RPS24 gene mutations | AD | Normal for age, >1st percentile | |
| Paroxysmal Nocturnal Hemoglobinuria | Intravascular hemolysis, venous thrombosis, variable degrees of BMF, PIGA mutations produce GPI-negative cell clones | Somatic muations | Shorter in GPI- negative cells than in GPI-positive cells | May indicate replicative stress or stem cell exhaustion |
| Myelodysplastic Syndrome | Dysplastic blood cells, cytogenetic abnormalities | Multifactorial | Shorter in granulocytes and monocytes | Suggests presence of chromosomal instability |
| Hematopoietic Malignancies | Variable, dependent on malignancy | Multifactorial | Shorter telomeres in leukemic cells | Suggests presence of chromosomal instability |
| Hematopoietic Stem Cell Transplantation | Dependent on the disease that requires transplant HSCT | NA | Shorter than donor telomeres | May indicate ageing replicative stress and ageing |
| Solid Tumors | Dependent on disease; includes bladder, lung, renal, head and neck, and breast cancers | Multifactorial | Not as short as DC on the individual patient level, but statistically significant differences between cases and controls | May be markers of disease predisposition, ageing, and/or oxidative stress |
| Inflammation | Ulcerative Colitis | Multifactorial | ||
| Arthritis | HLA associations | |||
| Ageing and Stress | Cardiovascular disease, smoking, diabetes, obesity, caregivers of the chronically ill | Multifactorial |
Numerous, more specific clinical findings are described in the text. DC is the only disorder in which all patients have very short telomeres (< 1st percentile).
Abbreviations: AD, autosomal dominant; AR, autosomal recessive; DC, dyskeratosis congenita; HSCT, hematopoietic stem cell transplantation; GPI, glycosylphosphatidylinositol; MDS, myelodysplastic syndrome; OR, odds ratio; CI, confidence interval; NA, not applicable
Idiopathic Pulmonary Fibrosis
Idiopathic pulmonary fibrosis (IPF) is a complex disorder in which genetic and environmental factors have been implicated. In a study of 73 probands from families with IPF, six were found to be heterozygous for mutations in TERT or TERC (Armanios et al. 2007). They also had short telomeres compared with related, wild-type family members as determined in lymphocytes by flow-FISH. One of these probands had other relatives with aplastic anemia. Additional TERT and/or TERC mutations were also identified in a separate study of familial and sporadic IPF (Tsakiri et al. 2007). Some family members in these IPF pedigrees had clinical features similar to DC, including anemia, cancer, and osteoporosis/osteopenia. Variable clinical phenotypes, including IPF, were also present in subjects with novel TERC mutations (Marrone et al. 2007). Pulmonary fibrosis and other pulmonary diseases have been noted in 20% of individuals with DC (Marrone et al. 2005) and patients with DC who undergo bone marrow transplantation are at increased risk of pulmonary complications as well (Yabe et al. 1997). Thus patients with IPF due to germline mutations in genes in TERC and TERT might also be considered to be within the DC spectrum, and should be managed and counseled in the same way as patients with classical DC.
TELOMERE BIOLOGY IN INHERITED BONE MARROW FAILURE SYNDROMES
Fanconi Anemia
FA is the most common IBMFS (Alter 2003). As in DC, there is also a broad spectrum of disease phenotype in FA. The inheritance of FA is usually autosomal recessive, but X-linked disease has also been identified in one subtype (FANCB). To date, mutations have been found in 13 different genes in FA. The products of these genes are part of a complex DNA damage repair pathway (Shimamura 2006a; Tamary and Alter 2007). Approximately 75% of patients with FA present with congenital anomalies which include abnormal radii and/or thumbs, abnormal kidneys, short stature, café-au-lait spots, skin hyperpigmentation, triangular facies, microcephaly and decreased fertility. Patients with FA are at very high risk of BMF, MDS, leukemia and epithelial cancers. FA is diagnosed in the laboratory by detection of an increased rate of chromosomal aberrations in mitogen-stimulated peripheral blood lymphocytes after culture in the presence of a DNA-crosslinker such as diepoxybutane or mitomycin C (Tamary and Alter 2007). If the results from lymphocytes are inconclusive, but clinical suspicion for FA is high, fibroblasts are often used as a second test, as hematopoietic somatic mosaicism has been reported (Lo-Ten-Foe et al. 1997).
The first study of telomeres in FA was reported on six patients from four families (Ball et al. 1998). Telomere lengths were determined by the TRF method on genomic DNA isolated from total leukocytes, and in granulocytes and mononuclear cells in a subset of patients, but specific data on which cell types were measured in which patients was not reported. Two patients with FA had normal telomere lengths for age. The other four FA patients, consisting of two sibling pairs, had decreased telomere lengths for age. As noted above, granulocyte telomeres may be biased towards more rapid shortening, which suggests that telomere measurement in total leukocytes, which will include granulocytes, may not be indicative of true germline telomere length.
Increased telomere shortening and telomerase activation were found in another study of 45 patients with FA (Leteurtre et al. 1999). Overall, TRF length was shorter in patients with FA compared with controls, but there was a wide scatter of data. This report also suggested that telomere shortening correlated with the progression of bone marrow aplasia. The authors proposed that telomere shortening may play a role in the evolution of BMF in patients with FA. They conducted a follow-up study with an additional 26 additional patients with FA (Li et al. 2003). Individual annual telomere-shortening rates (IATSR) were higher (195 base pairs/year) among patients with FA with severe BMF compared with those with non-severe BMF (115 base pairs/year). Sixty-six percent of patients with severe BMF had high telomere shortening rates, whereas 75 % of those with non-severe BMF had low telomere-shortening rates. In contrast, shorter TRF lengths were not noted in patients with FA with malignant or clonal bone marrow abnormalities compared with patients with FA without these abnormalities (Ball et al. 1998). However, telomerase activity was found to be 4.8-fold higher in FA cells (from 25 patients) than in controls (n=15).
Our study of the role of flow-FISH determination of telomere length in white blood cell subsets (granulocytes, lymphocytes, naïve T-cells, memory T-cells, B-cells, NK/NKT cells, and total leukocytes) in IBMFS patients showed that seven out of 17 patients with FA had very short telomeres (defined as <1st percentile for age) in at least one cell type (Figure 1B) (Alter et al. 2007a). These cell types more commonly included total leukocytes and granulocytes; i.e. cells or groups that include cells with a much higher turnover rate. Only three of 16 patients with FA who had not received a bone marrow transplant had very short telomeres in three or four leukocyte subsets. In contrast, the majority of patients with DC had very short telomeres in all cell lines, including long lived T-cells. Restriction of the analysis to the combination of the three most informative subsets (total lymphocytes, naïve T cells, and B cells) revealed that none of the patients with FA had very short telomeres in all three, compared with 26 of 28 patients with DC (including three silent carriers).
Proposed mechanisms for accelerated telomere shortening in FA include increased chromosomal breakage, excess free radical formation and/or abnormal regulation of cell proliferation related to deregulation of apoptosis (Adelfalk et al. 2001; Franco et al. 2004; Li et al. 2003). For example, fibroblasts from FA patients have higher rates of telomere loss and grow more slowly than control fibroblasts (Adelfalk et al. 2001). Lymphocytes from patients with FA had a higher frequency of extra-chromosomal TTAGGG signals by direct FISH as well as reduced signal at the telomere (Callen et al. 2002; Hanson et al. 2001). The FA genetic defects that induce alterations in cell proliferation through cell cycle delays and a high rate of apoptosis can result in stress hematopoiesis and BMF. Telomere shortening appears to be a marker of ageing and rapid cell turnover in FA. When the ability of hematopoietic stem/progenitor FA cells to proliferate becomes impaired due to TRF shortening, progressive pancytopenia may occur. Thus the very short telomeres found in patients with FA may be a consequence of hematopoietic failure or stress, and/or a result of treatment, in some patients, rather than constitutionally short telomeres.
Shwachman-Diamond Syndrome
Shwachman-Diamond Syndrome (SDS) is an autosomal recessive disease that typically presents in early childhood with malabsorption due to pancreatic exocrine insufficiency and neutropenia (Rothbaum et al. 2002; Shimamura 2006b; Tamary and Alter 2007). Other features include short stature, metaphyseal dysostosis, protuberant abdomen, ichthyotic skin rash, and other less commonly observed malformations. Patients with SDS are at high risk of pancytopenia, MDS and leukemia. Decreased serum trypsinogen and isoamylase are useful diagnostic tests but the latter is not reliable for children less than three years of age (Ip et al. 2002). At least 90% of patients with SDS have been found to have mutations in the SBDS gene (Woloszynek et al. 2004). The role of SBDS, a member of a highly conserved gene family that functions in RNA metabolism, is currently being elucidated.
There have been very few studies of telomere length in SDS to date. The first measured telomere lengths in 12 cases by TRF on DNA isolated from either neutrophils (n=3), whole marrow nucleated cells (n=7) or marrow mononuclear cells (n=2) and from neutrophils of 41 healthy controls (Thornley et al. 2002). The mean age-adjusted telomere length in SDS patients was 1.4 kilobase pairs shorter than in the age-adjusted controls. The degree of telomere shortening seen in SDS was similar to that reported in FA but not as profound as seen in DC.
Five patients with SDS were evaluated in our study of flow-FISH determination of telomere length in WBC subsets (Figure 1C) (Alter et al. 2007a). One of these patients had telomere lengths just below the 1st percentile in five of the seven white blood cell subsets studied, although in only two of the three sentinel subtypes (total lymphocytes and B cells, but not naïve T cells). These data suggest that differences in telomere length in SDS patients could be attributed to stress hematopoiesis, as the main cell lines with telomere shortening are granulocytes. Our data also indicate that short telomeres are not an intrinsic part of SDS.
In a study of 91 patients with apparently acquired AA, four were found to be heterozygous for the 258+2 T-C SBDS gene mutation (Calado et al. 2007). These patients did not have clinical evidence of pancreatic exocrine failure or other anomalies associated with SDS. Telomere lengths in granulocytes of cases with heterozygous mutations in SBDS were short for their age, and the authors speculated that this heterozygosity predisposes to BMF by accelerating telomere shortening.
Diamond-Blackfan Anemia
Patients with Diamond-Blackfan Anemia (DBA) typically present with pure red blood cell aplasia in the first year of life (Lipton 2006). This reticulocytopenic anemia is usually macrocytic, and the bone marrow of these patients has a paucity of erythroid precursors. Elevated fetal hemoglobin and increased red blood cell adenosine deaminase are common features in the patients, but may also be seen in some otherwise asymptomatic relatives. Other clinical features sometimes seen in DBA include short stature, abnormal thumbs, craniofacial abnormalities, as well as renal and cardiac defects. Patients with DBA are also at increased risk of AML, osteosarcoma and possibly other malignancies (Lipton et al. 2001).
Heterozygous mutations in RPS19, a ribosomal protein, have been identified in 20–25% of patients with DBA (Draptchinskaia et al. 1999). Subsequent studies suggest haploinsufficiency of RPS19 instead of a dominant negative effect (Gazda et al. 2004). A second gene, RPS24, was found to be mutated in a small subset (2%) of patients with DBA (Gazda et al. 2006) and a third gene, RPS17, in one patient (Cmejla et al. 2007).
Our study is the only one to report on telomere length in patients with DBA to date (Alter et al. 2007a). We studied WBC subsets by flow-FISH in 14 patients with DBA, four of whom had mutations in RPS19. Twelve patients had telomere lengths >1st percentile in all cells studied, one patient had very short telomeres (<1st percentile) only in granulocytes, and one other patient had very short telomeres in all six white blood cell types analyzed (Figure 1D). The latter patient was transfusion-dependent and had microcephaly and developmental delay, suggestive of a complex syndrome. He did not have mutations in RPS19, RPS24, DKC1, or TERC.
Paroxysmal Nocturnal Hemoglobinuria
Paroxysmal nocturnal hemoglobinuria (PNH) is clinically characterized by intravascular hemolysis, venous thrombosis, and variable degrees of BMF. It is a clonal disorder in which hematopoietic clones are derived from stem cells with somatic mutations in the PIGA gene (Bessler et al. 1994; Brodsky 2006; Miyata et al. 1994). These mutations result in variable degrees of clonal expansion of cells lacking expression of glycosylphosphatidylinositol anchored protein (GPI-negative cells). The disease manifestations of PNH are highly variable and correlated to the clonal expansion of GPI-negative cells. Several studies have evaluated telomere length in subjects with PNH in an effort to understand the role of telomere length in the replicative history of GPI-positive and GPI-negative cells and PNH disease progression.
The first studies of telomere length in PNH did not separately study GPI-negative and GPI-positive cells but did show that three patients with hemolytic PNH at presentation had shorter telomeres than control subjects (Ball et al. 1998). However, a second study of granulocyte telomere length in six patients with PNH did not show a significant difference between telomere lengths in these patients compared with controls (Brummendorf et al. 2001a). Subsequent studies have taken advantage of advances in the separation of GPI-positive from GPI-negative cells and in telomere length measurement to clarify the differences in telomere length between these cell types in PNH patients and in comparison with healthy subjects. These studies of telomere length in granulocytes have shown that GPI-negative cells have shorter telomeres than GPI-positive cells (Baerlocher et al. 2007; Beier et al. 2005; Karadimitris et al. 2003). The study by Baerlocher et al. also showed that within the same patient, telomere length in GPI-negative cells was significantly shorter than in GPI-positive cells and that the difference in telomere length between GPI-negative and GPI-positive cells was also highly correlated with the size of the PNH clone. In PNH, telomere length appears to correlate well with the replicative history of the cells; shorter telomeres are present in GPI-negative, clonal cells with higher cellular proliferation than GPI-positive cells.
Myelodysplastic Syndrome
MDSs are clonal disorders of hematopoietic stem cells that are characterized by dysplastic features in blood cells and chromosomal abnormalities (Bennett 2005; Hofmann and Koeffler 2005). Patients with IBMFS or acquired AA are at high risk of MDS, but it can also develop spontaneously in previously healthy individuals. MDS is classified as a pre-leukemic disorder because approximately 30% of patients with MDS will progress to acute myeloid leukemia (AML).
Studies of telomere biology in spontaneous MDS suggest that increased telomere shortening is present in granulocytes and/or monocytes, by both TRF and flow-FISH methods (Ball et al. 1998; Brummendorf et al. 2001c; Ohyashiki et al. 1994; Rigolin et al. 2004; Sieglova et al. 2004). Increased telomerase activity has been reported in approximately 40% of MDS patients, but correlations with outcome or telomere length have been inconsistent (Engelhardt et al. 2004; Gurkan et al. 2005; Ohyashiki et al. 1994; Ohyashiki et al. 1999). MDS appears to be in the pathway of progression from BMF (acquired AA or an IBMFS) to leukemia. The presence of abnormalities in telomere biology in these patients suggests that insufficient telomere length maintenance may predispose to abnormalities in cellular replication and the development of malignancy.
Hematopoietic Malignancies
Studies of hematopoietic malignancies have shown that progressive telomere shortening is present in chronic and acute myeloid and lymphoid leukemias. The contribution of telomere shortening to the cytogenetic abnormalities commonly seen in leukemia is an active area of investigation; many of these studies have been reviewed in detail by others (Deville et al. 2006; Drummond et al. 2007). The majority of these studies were performed on pre-therapeutic specimens, but it should be noted that cytotoxic chemotherapy can also contribute to telomere shortening (Fern et al. 2004; Franco et al. 2003).
It is possible that there is overlap in telomere dysfunction in IBMFS, acquired AA, MDS, and hematopoietic malignancies. Many acute leukemias have reduced telomere length at diagnosis when compared with normal cells and increased telomerase activity (Engelhardt et al. 1998; Hartmann et al. 2005). The effect of telomerase activity and telomere length on acute leukemia outcomes is not consistent. A study of adult T-cell leukemia suggested that patients with elevated telomerase activity and shorter telomeres were more likely to have a poorer prognosis (Kubuki et al. 2005). However, a study of pediatric T-cell leukemia did not find a relationship between telomerase activity or telomere length and outcome (Kleideiter et al. 2005).
The role of telomere biology in chronic myeloid leukemia (CML) has been relatively well-studied. Its well-characterized chronic and blast phases, BCR-ABL translocation t(11;22)(q25;q13) and fusion protein, and proven response to therapy targeting the fusion protein with the tyrosine kinase inhibitor imatinib mesylate have facilitated these studies. Malignant clones with BCR-ABL translocations were shown to have decreased telomere length (measured by flow-FISH) compared with normal cells without BCR-ABL translocations (Brummendorf et al. 2000). In addition, successful treatment with imatinib mesylate resulted in an increase in the mean telomere length (Brummendorf et al. 2003).
Recent studies of telomere length and telomerase activity in hematopoietic malignancies have not always been consistent with regard to the effect of outcome, but they do point to the role that telomere loss plays in the development of malignancy. Abnormal marrow stem cell clones could result from chromosomal instability generated by abnormal telomere lengths. The malignant cells may be able to survive, despite critically short telomeres, due to up-regulation of telomerase activity or activation of alternative pathways (Artandi and Attardi 2005; Henson et al. 2002). However, in all of these studies, further evaluation of telomere length and telomere dynamics in specific cell subtypes will be important to advance our understanding of their contributions to normal hematopoiesis, BMF, and hematopoietic malignancy.
Hematopoietic Stem Cell Transplantation
Accelerated telomere shortening after allogeneic hematopoietic stem cell transplantation (HSCT) was suggested in several studies (Brummendorf et al. 2001b; Mathioudakis et al. 2000; Notaro et al. 1997; Robertson et al. 2001; Rufer et al. 2001; Wynn et al. 1999; Wynn et al. 1998). Increased telomere rates were observed after both myeloablative and nonmyeloablative condition for allogeneic HSCT (Lahav et al. 2005; Rocci et al. 2007; Roelofs et al. 2003) and after the use of umbilical cord blood for HSCT (Pipes et al. 2006). However, long-term follow-up data suggest that there may be some variability in telomere length after hematopoietic stem cell reconstitution. Transient telomere shortening followed by a period of telomere lengthening resulting in telomere length homeostasis was reported after both allogeneic and autologous HSCT (Bhatia et al. 2005; Roelofs et al. 2003). Others found that telomere lengths remain shorter than expected after autologous HSCT (Rocci et al. 2007). These findings have led to speculation that the telomeres of elderly hematopoietic stem cells donors might become too short for acceptable lifetime risks in a young recipient (Akiyama et al. 2000). Hematopoietic stem cell replicative stress might contribute to the increased rates of telomere shortening observed after HSCT. The combination of chemotherapy, which has been shown to reduce telomere lengths (Engelhardt et al. 1998; Ricca et al. 2005; Rocci et al. 2007), in addition to the underlying bone marrow disorder that made HSCT necessary could be contributory mechanisms. The high degree of variability which has been reported between HSCT patients, and the various methods and times studied, make it difficult to determine whether there really is a significant degree of telomere shortening after HSCT. Despite the differences noted in telomere length after HSCT, there does not appear to be evidence of stem cell exhaustion in long-term follow-up in these and other studies (de Pauw et al. 2002). Additional, larger longitudinal studies are required to further understand these differences.
OTHER COMPLEX DISORDERS ASSOCIATED WITH SHORT TELOMERES
Advances in methods to measure telomere lengths in substantial sample sizes have led to large association studies of the role of germline telomere length as a risk factor for complex diseases. The differences in telomere length between disease cases and healthy controls, while statistically significant in these studies, are not as extreme as the differences between individuals with DC and healthy subjects. However, they do illustrate the complex and dynamic role that telomeres play in chromosomal stability and cell division (Table 2).
The connection between telomere abnormalities in cancer tissues (Meeker et al. 2002; Meeker et al. 2004; Meeker and Argani 2004; O'Sullivan et al. 2002; Plentz et al. 2004) and in cancer predisposition syndromes such as DC, have led to studies of the role of telomere length as a cancer risk factor. Three independent studies of bladder cancer found that individuals with bladder cancer had shorter telomeres in either blood or buccal cells than healthy control subjects (Broberg et al. 2005; McGrath et al. 2007; Wu et al. 2003). Similar findings were noted in patients with head, neck, lung, and breast cancers (Shen et al. 2007; Wu et al. 2003). In these studies, telomere length differences between cases and controls were statistically significant, but the profound telomere shortening seen in DC was not present in germline DNA from these cases. In most reports, these findings were in excess of telomere shortening associated with smoking.
Chronic inflammation also contributes to telomere shortening, possibly due to oxidative stress. Short telomeres in T-cells and granulocytes have been reported in rheumatoid arthritis (Koetz et al. 2000). In addition, a study that evaluated telomere shortening by quantitative FISH from biopsy specimens from patients with ulcerative colitis found that higher rates of telomere shortening were present in the dysplastic biopsies than in non-dysplastic samples from colitis patients or control subjects (O'Sullivan et al. 2002).
Numerous studies have also sought to use germline telomere length as a marker of biological ageing and chronic disease. Several studies found that telomere shortening is associated with cardiovascular disease (CVD) and/or environmental factors associated with CVD, such as smoking or stress (Bekaert et al. 2007; Brouilette et al. 2007; Epel et al. 2006; Fitzpatrick et al. 2007). The presence of psychological stress has also been associated with oxidative stress and increased telomere shortening in caregivers of chronically ill family members (Damjanovic et al. 2007; Epel et al. 2004). Additional factors that appear to contribute to accelerated telomere shortening include smoking, socioeconomic status, obesity, metabolic syndrome, and diabetes (Aviv et al. 2006; Broberg et al. 2005; Cherkas et al. 2006; Epel et al. 2006; McGrath et al. 2007; Valdes et al. 2005; Wu et al. 2003). Germline telomere length may serve as a marker of the degree of cellular ageing in this context and could be a biomarker of risk in these complex disorders
CLINICAL IMPLICATIONS OF SHORT TELOMERE LENGTHS
Rapid advances in methods of telomere length measurement, the role of telomere biology in a growing list of complex clinical disorders, and the identification of mutations in genes important in telomere biology associated with many of these disorders makes the interpretation of the literature challenging, and the application to clinical care even more complex. Several studies have suggested that these measurements may be good markers of outcome in hematopoietic malignancies (Brummendorf et al. 2000; Gowda and Byrd 2006; Iwama et al. 1997). A recent study of CVD suggested that mean leukocyte telomere length could help predict CVD in high-risk men and also identify individuals who would benefit from statin treatment (Brouilette et al. 2007). As our understanding of the role of telomere biology in complex disorders grow, we will be better able to develop direct clinical applications.
Thus far, DC appears to be the best studied example of the utility of measurement of telomere length. While the clinical diagnosis of classical DC may remain contingent upon the triad of dysplastic nails, lacey reticular skin pigmentation, and oral leukoplakia, it is important to recognize that individuals may have very short telomeres and mutations in telomere biology genes in the absence of the DC diagnostic triad, or even of any clinical phenotype beyond BMF. Children and young adults with BMF are commonly screened for FA, even in the absence of the common dysmorphic findings. In BMF patients who have normal chromosome breakage in cells cultured with clastogenic agents, we suggest screening for DC by determination of telomere length in white blood cell subsets by flow-FISH. Diagnosis of DC in the context of BMF has important clinical implications; patients with DC do not respond to immunosuppressive therapy for BMF and are at increased risk of complications of bone marrow transplantation (Al Rahawan et al. 2006; de la Fuente and Dokal 2007). Patients with DC may benefit from reduced intensity conditioning (non-myeloablative) prior to HSCT as well as, when possible, avoidance of agents with known pulmonary and hepatic toxicities. Clinical trials are currently underway to evaluate this. It may also be beneficial for the majority of DC HSCTs to be conducted at a small number of large HSCT centers in order to increase the experience of the clinical team with DC-specific complications.
The spectrum of disorders caused by germline mutations in telomere biology genes is growing beyond what has traditionally been thought of as classical DC (i.e. patients with the DC diagnostic triad). We suggest that the definition of DC be expanded to include patients with germline mutations in telomere biology genes and/or telomere lengths <1st percentile for age in lymphocytes. This group will include patents with telomere biology abnormalities as well as BMF that fails to respond to immunosuppression, IPF, and possibly other other yet-to-be-identified disorders (Table 1). We recommend assigning such patients to the category of an inherited disorder because this will impact medical management, bone marrow transplantation, cancer surveillance, and family counseling. Longitudinal studies of patients with very short telomeres but without classical DC are necessary to further understand the long-term sequelae, such as malignancy, osteonecrosis/osteoporosis, pulmonary and liver disease. It is highly plausible that since DC is a cancer predisposition syndrome, individuals with abnormally short telomeres and/or germline mutations in telomere biology genes will also be at very high risk of cancer. Careful long-term follow-up of patient cohorts and cancer screening of these patients will be necessary for optimal patient care.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Reference List
- Adelfalk C, Lorenz M, Serra V, von Zglinicki T, Hirsch-Kauffmann M, Schweiger M. Accelerated telomere shortening in Fanconi anemia fibroblasts--a longitudinal study. FEBS Lett. 2001;506:22–26. doi: 10.1016/s0014-5793(01)02869-1. [DOI] [PubMed] [Google Scholar]
- Akiyama M, Asai O, Kuraishi Y, Urashima M, Hoshi Y, Sakamaki H, Yabe H, Furukawa T, Yamada O, Mizoguchi H, Yamada H. Shortening of telomeres in recipients of both autologous and allogeneic hematopoietic stem cell transplantation. Bone Marrow Transplant. 2000;25:441–447. doi: 10.1038/sj.bmt.1702144. [DOI] [PubMed] [Google Scholar]
- Al Rahawan MM, Giri N, Alter BP. Intensive immunosuppression therapy for aplastic anemia associated with dyskeratosis congenita. Int J Hematol. 2006;83:275–276. doi: 10.1532/IJH97.06030. [DOI] [PubMed] [Google Scholar]
- Alter BP. Inherited bone marrow failure syndromes. 2003;6:280–365. [Google Scholar]
- Alter BP. Bone marrow failure: a child is not just a small adult (but an adult can have a childhood disease) Hematology Am Soc Hematol Educ Program. 2005:96–103. doi: 10.1182/asheducation-2005.1.96. [DOI] [PubMed] [Google Scholar]
- Alter BP, Baerlocher GM, Savage SA, Chanock SJ, Weksler BB, Willner JP, Peters JA, Giri N, Lansdorp PM. Very short telomere length by flow fluorescence in situ hybridization identifies patients with dyskeratosis congenita. Blood. 2007a;110:1439–1447. doi: 10.1182/blood-2007-02-075598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alter BP, Denny CC, Peters JA, Giri N, Wilfond BS. Disclosure of "Unwanted" Genetic Information To Benefit a Family Member. Pediatric Academic Societies 2007 Annual Meeting Online E-PAS2007:615863.4.2007b. [Google Scholar]
- Andrew T, Aviv A, Falchi M, Surdulescu GL, Gardner JP, Lu X, Kimura M, Kato BS, Valdes AM, Spector TD. Mapping genetic Loci that determine leukocyte telomere length in a large sample of unselected female sibling pairs. Am J Hum Genet. 2006;78:480–486. doi: 10.1086/500052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armanios MY, Chen JJ, Cogan JD, Alder JK, Ingersoll RG, Markin C, Lawson WE, Xie M, Vulto I, Phillips JA, III, Lansdorp PM, Greider CW, Loyd JE. Telomerase mutations in families with idiopathic pulmonary fibrosis. N Engl J Med. 2007;356:1317–1326. doi: 10.1056/NEJMoa066157. [DOI] [PubMed] [Google Scholar]
- Artandi SE, Attardi LD. Pathways connecting telomeres and p53 in senescence, apoptosis, and cancer. Biochem Biophys Res Commun. 2005;331:881–890. doi: 10.1016/j.bbrc.2005.03.211. [DOI] [PubMed] [Google Scholar]
- Aviv A, Valdes A, Gardner JP, Swaminathan R, Kimura M, Spector TD. Menopause modifies the association of leukocyte telomere length with insulin resistance and inflammation. J Clin Endocrinol Metab. 2006;91:635–640. doi: 10.1210/jc.2005-1814. [DOI] [PubMed] [Google Scholar]
- Baerlocher GM, Lansdorp PM. Telomere length measurements in leukocyte subsets by automated multicolor flow-FISH. Cytometry A. 2003;55:1–6. doi: 10.1002/cyto.a.10064. [DOI] [PubMed] [Google Scholar]
- Baerlocher GM, Sloand EM, Young NS, Lansdorp PM. Telomere length in paroxysmal nocturnal hemoglobinuria correlates with clone size. Exp Hematol. 2007 Aug 11; doi: 10.1016/j.exphem.2007.06.010. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- Baerlocher GM, Vulto I, de Jong G, Lansdorp PM. Flow cytometry and FISH to measure the average length of telomeres (flow FISH) Nat Protoc. 2006;1:2365–2376. doi: 10.1038/nprot.2006.263. [DOI] [PubMed] [Google Scholar]
- Baird DM. New developments in telomere length analysis. Exp Gerontol. 2005;40:363–368. doi: 10.1016/j.exger.2005.02.008. [DOI] [PubMed] [Google Scholar]
- Ball SE, Gibson FM, Rizzo S, Tooze JA, Marsh JC, Gordon-Smith EC. Progressive telomere shortening in aplastic anemia. Blood. 1998;91:3582–3592. [PubMed] [Google Scholar]
- Beier F, Balabanov S, Buckley T, Dietz K, Hartmann U, Rojewski M, Kanz L, Schrezenmeier H, Brummendorf TH. Accelerated telomere shortening in glycosylphosphatidylinositol (GPI)-negative compared with GPI-positive granulocytes from patients with paroxysmal nocturnal hemoglobinuria (PNH) detected by proaerolysin flow-FISH. Blood. 2005;106:531–533. doi: 10.1182/blood-2004-10-3996. [DOI] [PubMed] [Google Scholar]
- Bekaert S, De Meyer T, Rietzschel ER, De Buyzere ML, De Bacquer D, Langlois M, Segers P, Cooman L, Van Damme P, Cassiman P, Van Criekinge W, Verdonck P, De Backer GG, Gillebert TC, Van Oostveldt P. Telomere length and cardiovascular risk factors in a middle-aged population free of overt cardiovascular disease. Aging Cell. 2007;6:639–647. doi: 10.1111/j.1474-9726.2007.00321.x. [DOI] [PubMed] [Google Scholar]
- Bennett JM. A comparative review of classification systems in myelodysplastic syndromes (MDS) Semin Oncol. 2005;32:S3–10. doi: 10.1053/j.seminoncol.2005.06.021. [DOI] [PubMed] [Google Scholar]
- Bessler M, Mason PJ, Hillmen P, Miyata T, Yamada N, Takeda J, Luzzatto L, Kinoshita T. Paroxysmal nocturnal haemoglobinuria (PNH) is caused by somatic mutations in the PIG-A gene. EMBO J. 1994;13:110–117. doi: 10.1002/j.1460-2075.1994.tb06240.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhatia R, Van Heijzen K, Palmer A, Komiya A, Slovak ML, Chang KL, Fung H, Krishnan A, Molina A, Nademanee A, O'Donnell M, Popplewell L, Rodriguez R, Forman SJ, Bhatia S. Longitudinal assessment of hematopoietic abnormalities after autologous hematopoietic cell transplantation for lymphoma. J Clin Oncol. 2005;23:6699–6711. doi: 10.1200/JCO.2005.10.330. [DOI] [PubMed] [Google Scholar]
- Bischoff C, Graakjaer J, Petersen HC, Hjelmborg JB, Vaupel JW, Bohr V, Koelvraa S, Christensen K. The heritability of telomere length among the elderly and oldest-old. Twin Res Hum Genet. 2005;8:433–439. doi: 10.1375/183242705774310141. [DOI] [PubMed] [Google Scholar]
- Brazzola P, Duval M, Fournet JC, Gauvin F, Dalle JH, Champagne J, Champagne MA. Fatal diffuse capillaritis after hematopoietic stem-cell transplantation for dyskeratosis congenita despite low-intensity conditioning regimen. Bone Marrow Transplant. 2005;36:1103–1105. doi: 10.1038/sj.bmt.1705171. [DOI] [PubMed] [Google Scholar]
- Broberg K, Bjork J, Paulsson K, Hoglund M, Albin M. Constitutional short telomeres are strong genetic susceptibility markers for bladder cancer. Carcinogenesis. 2005;26:1263–1271. doi: 10.1093/carcin/bgi063. [DOI] [PubMed] [Google Scholar]
- Brodsky RA. New insights into paroxysmal nocturnal hemoglobinuria. Hematology Am Soc Hematol Educ Program. 2006:24–8. 516. doi: 10.1182/asheducation-2006.1.24. [DOI] [PubMed] [Google Scholar]
- Brouilette SW, Moore JS, McMahon AD, Thompson JR, Ford I, Shepherd J, Packard CJ, Samani NJ. Telomere length, risk of coronary heart disease, and statin treatment in the West of Scotland Primary Prevention Study: a nested case-control study. Lancet. 2007;369:107–114. doi: 10.1016/S0140-6736(07)60071-3. [DOI] [PubMed] [Google Scholar]
- Brummendorf TH, Ersoz I, Hartmann U, Bartolovic K, Balabanov S, Wahl A, Paschka P, Kreil S, Lahaye T, Berger U, Gschaidmeier H, Bokemeyer C, Hehlmann R, Dietz K, Lansdorp PM, Kanz L, Hochhaus A. Telomere length in peripheral blood granulocytes reflects response to treatment with imatinib in patients with chronic myeloid leukemia. Blood. 2003;101:375–376. doi: 10.1182/blood-2002-08-2557. [DOI] [PubMed] [Google Scholar]
- Brummendorf TH, Holyoake TL, Rufer N, Barnett MJ, Schulzer M, Eaves CJ, Eaves AC, Lansdorp PM. Prognostic implications of differences in telomere length between normal and malignant cells from patients with chronic myeloid leukemia measured by flow cytometry. Blood. 2000;95:1883–1890. [PubMed] [Google Scholar]
- Brummendorf TH, Maciejewski JP, Mak J, Young NS, Lansdorp PM. Telomere length in leukocyte subpopulations of patients with aplastic anemia. Blood. 2001a;97:895–900. doi: 10.1182/blood.v97.4.895. [DOI] [PubMed] [Google Scholar]
- Brummendorf TH, Rufer N, Baerlocher GM, Roosnek E, Lansdorp PM. Limited telomere shortening in hematopoietic stem cells after transplantation. Ann NY Acad Sci. 2001b;938:1–7. doi: 10.1111/j.1749-6632.2001.tb03568.x. [DOI] [PubMed] [Google Scholar]
- Brummendorf TH, Rufer N, Holyoake TL, Maciejewski J, Barnett MJ, Eaves CJ, Eaves AC, Young N, Lansdorp PM. Telomere length dynamics in normal individuals and in patients with hematopoietic stem cell-associated disorders. Ann NY Acad Sci. 2001c;938:293–303. doi: 10.1111/j.1749-6632.2001.tb03598.x. [DOI] [PubMed] [Google Scholar]
- Calado RT, Graf SA, Wilkerson KL, Kajigaya S, Ancliff PJ, Dror Y, Chanock SJ, Lansdorp PM, Young NS. Mutations in the SBDS gene in acquired aplastic anemia. Blood. 2007 doi: 10.1182/blood-2007-03-080044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callen E, Samper E, Ramirez MJ, Creus A, Marcos R, Ortega JJ, Olive T, Badell I, Blasco MA, Surralles J. Breaks at telomeres and TRF2-independent end fusions in Fanconi anemia. Hum Mol Genet. 2002;11:439–444. doi: 10.1093/hmg/11.4.439. [DOI] [PubMed] [Google Scholar]
- Cherkas LF, Aviv A, Valdes AM, Hunkin JL, Gardner JP, Surdulescu GL, Kimura M, Spector TD. The effects of social status on biological aging as measured by white-blood-cell telomere length. Aging Cell. 2006;5:361–365. doi: 10.1111/j.1474-9726.2006.00222.x. [DOI] [PubMed] [Google Scholar]
- Cmejla R, Cmejlova J, Handrkova H, Petrak J, Pospisilova D. Ribosomal protein S17 gene (RPS17) is mutated in Diamond-Blackfan anemia. Hum Mutat. 2007 doi: 10.1002/humu.20608. [DOI] [PubMed] [Google Scholar]
- Collins K, Mitchell JR. Telomerase in the human organism. Oncogene. 2002;21:564–579. doi: 10.1038/sj.onc.1205083. [DOI] [PubMed] [Google Scholar]
- Damjanovic AK, Yang Y, Glaser R, Kiecolt-Glaser JK, Nguyen H, Laskowski B, Zou Y, Beversdorf DQ, Weng NP. Accelerated telomere erosion is associated with a declining immune function of caregivers of Alzheimer's disease patients. J Immunol. 2007;179:4249–4254. doi: 10.4049/jimmunol.179.6.4249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de la Fuente J, Dokal I. Dyskeratosis congenita: advances in the understanding of the telomerase defect and the role of stem cell transplantation. Pediatr Transplant. 2007;11:584–594. doi: 10.1111/j.1399-3046.2007.00721.x. [DOI] [PubMed] [Google Scholar]
- de Lange T. Shelterin: the protein complex that shapes and safeguards human telomeres. Genes Dev. 2005;19:2100–2110. doi: 10.1101/gad.1346005. [DOI] [PubMed] [Google Scholar]
- de Pauw ES, Otto SA, Wijnen JT, Vossen JM, van Weel MH, Tanke HJ, Miedema F, Willemze R, Roelofs H, Fibbe WE. Long-term follow-up of recipients of allogeneic bone marrow grafts reveals no progressive telomere shortening and provides no evidence for haematopoietic stem cell exhaustion. Br J Haematol. 2002;116:491–496. doi: 10.1046/j.0007-1048.2001.03283.x. [DOI] [PubMed] [Google Scholar]
- Demeter J, Messer G, Schrezenmeier H. Clinical relevance of the TNF-alpha promoter/enhancer polymorphism in patients with aplastic anemia. Ann Hematol. 2002;81:566–569. doi: 10.1007/s00277-002-0544-6. [DOI] [PubMed] [Google Scholar]
- Deville L, Hillion J, Lanotte M, Rousselot P, Segal-Bendirdjian E. Diagnostics, prognostic and therapeutic exploitation of telomeres and telomerase in leukemias. Curr Pharm Biotechnol. 2006;7:171–183. doi: 10.2174/138920106777549768. [DOI] [PubMed] [Google Scholar]
- Drachtman RA, Alter BP. Dyskeratosis congenita: clinical and genetic heterogeneity. Report of a new case and review of the literature. Am J Pediatr Hematol Oncol. 1992;14:297–304. [PubMed] [Google Scholar]
- Draptchinskaia N, Gustavsson P, Andersson B, Pettersson M, Willig TN, Dianzani I, Ball S, Tchernia G, Klar J, Matsson H, Tentler D, Mohandas N, Carlsson B, Dahl N. The gene encoding ribosomal protein S19 is mutated in Diamond-Blackfan anaemia. Nat Genet. 1999;21:169–175. doi: 10.1038/5951. [DOI] [PubMed] [Google Scholar]
- Dror Y, Freedman MH, Leaker M, Verbeek J, Armstrong CA, Saunders FE, Doyle JJ. Low-intensity hematopoietic stem-cell transplantation across human leucocyte antigen barriers in dyskeratosis congenita. Bone Marrow Transplant. 2003;31:847–850. doi: 10.1038/sj.bmt.1703931. [DOI] [PubMed] [Google Scholar]
- Drummond MW, Balabanov S, Holyoake TL, Brummendorf TH. Concise review: Telomere biology in normal and leukemic hematopoietic stem cells. Stem Cells. 2007;25:1853–1861. doi: 10.1634/stemcells.2007-0057. [DOI] [PubMed] [Google Scholar]
- Dufour C, Capasso M, Svahn J, Marrone A, Haupt R, Bacigalupo A, Giordani L, Longoni D, Pillon M, Pistorio A, Di Michele P, Iori AP, Pongiglione C, Lanciotti M, Iolascon A. Homozygosis for (12) CA repeats in the first intron of the human IFN-gamma gene is significantly associated with the risk of aplastic anaemia in Caucasian population. Br J Haematol. 2004;126:682–685. doi: 10.1111/j.1365-2141.2004.05102.x. [DOI] [PubMed] [Google Scholar]
- Engelhardt M, Ozkaynak MF, Drullinsky P, Sandoval C, Tugal O, Jayabose S, Moore MA. Telomerase activity and telomere length in pediatric patients with malignancies undergoing chemotherapy. Leukemia. 1998;12:13–24. doi: 10.1038/sj.leu.2400889. [DOI] [PubMed] [Google Scholar]
- Engelhardt M, Wasch R, Guo Y. Telomeres and telomerase in normal and leukemic hematopoietic cells. Leuk Res. 2004;28:1001–1004. doi: 10.1016/j.leukres.2004.01.015. [DOI] [PubMed] [Google Scholar]
- Epel ES, Blackburn EH, Lin J, Dhabhar FS, Adler NE, Morrow JD, Cawthon RM. Accelerated telomere shortening in response to life stress. Proc Natl Acad Sci USA. 2004;101:17312–17315. doi: 10.1073/pnas.0407162101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epel ES, Lin J, Wilhelm FH, Wolkowitz OM, Cawthon R, Adler NE, Dolbier C, Mendes WB, Blackburn EH. Cell aging in relation to stress arousal and cardiovascular disease risk factors. Psychoneuroendocrinology. 2006;31:277–287. doi: 10.1016/j.psyneuen.2005.08.011. [DOI] [PubMed] [Google Scholar]
- Fern L, Pallis M, Ian Carter G, Seedhouse C, Russell N, Byrne J. Clonal haemopoiesis may occur after conventional chemotherapy and is associated with accelerated telomere shortening and defects in the NQO1 pathway; possible mechanisms leading to an increased risk of t-AML/MDS. Br J Haematol. 2004;126:63–71. doi: 10.1111/j.1365-2141.2004.05006.x. [DOI] [PubMed] [Google Scholar]
- Fitzpatrick AL, Kronmal RA, Gardner JP, Psaty BM, Jenny NS, Tracy RP, Walston J, Kimura M, Aviv A. Leukocyte telomere length and cardiovascular disease in the cardiovascular health study. Am J Epidemiol. 2007;165:14–21. doi: 10.1093/aje/kwj346. [DOI] [PubMed] [Google Scholar]
- Franco S, Ozkaynak MF, Sandoval C, Tugal O, Jayabose S, Engelhardt M, Moore MA. Telomere dynamics in childhood leukemia and solid tumors: a follow-up study. Leukemia. 2003;17:401–410. doi: 10.1038/sj.leu.2402815. [DOI] [PubMed] [Google Scholar]
- Franco S, van de Vrugt HJ, Fernandez P, Aracil M, Arwert F, Blasco MA. Telomere dynamics in Fancg-deficient mouse and human cells. Blood. 2004;104:3927–3935. doi: 10.1182/blood-2003-10-3626. [DOI] [PubMed] [Google Scholar]
- Frenck RW, Jr, Blackburn EH, Shannon KM. The rate of telomere sequence loss in human leukocytes varies with age. Proc Natl Acad Sci USA. 1998;95:5607–5610. doi: 10.1073/pnas.95.10.5607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gazda HT, Grabowska A, Merida-Long LB, Latawiec E, Schneider HE, Lipton JM, Vlachos A, Atsidaftos E, Ball SE, Orfali KA, Niewiadomska E, Da Costa L, Tchernia G, Niemeyer C, Meerpohl JJ, Stahl J, Schratt G, Glader B, Backer K, Wong C, Nathan DG, Beggs AH, Sieff CA. Ribosomal protein S24 gene is mutated in Diamond-Blackfan anemia. Am J Hum Genet. 2006;79:1110–1118. doi: 10.1086/510020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gazda HT, Zhong R, Long L, Niewiadomska E, Lipton JM, Ploszynska A, Zaucha JM, Vlachos A, Atsidaftos E, Viskochil DH, Niemeyer CM, Meerpohl JJ, Rokicka-Milewska R, Pospisilova D, Wiktor-Jedrzejczak W, Nathan DG, Beggs AH, Sieff CA. RNA and protein evidence for haplo-insufficiency in Diamond-Blackfan anaemia patients with RPS19 mutations. Br J Haematol. 2004;127:105–113. doi: 10.1111/j.1365-2141.2004.05152.x. [DOI] [PubMed] [Google Scholar]
- Gowda A, Byrd JC. Use of prognostic factors in risk stratification at diagnosis and time of treatment of patients with chronic lymphocytic leukemia. Curr Opin Hematol. 2006;13:266–272. doi: 10.1097/01.moh.0000231425.46148.b0. [DOI] [PubMed] [Google Scholar]
- Graakjaer J, Der-Sarkissian H, Schmitz A, Bayer J, Thomas G, Kolvraa S, Londono-Vallejo JA. Allele-specific relative telomere lengths are inherited. Hum Genet. 2006;119:344–350. doi: 10.1007/s00439-006-0137-x. [DOI] [PubMed] [Google Scholar]
- Gurkan E, Tanriverdi K, Baslamisli F. Telomerase activity in myelodysplastic syndromes. Leuk Res. 2005;29:1131–1139. doi: 10.1016/j.leukres.2005.03.006. [DOI] [PubMed] [Google Scholar]
- Hanson H, Mathew CG, Docherty Z, Mackie Ogilvie C. Telomere shortening in Fanconi anaemia demonstrated by a direct FISH approach. Cytogenet Cell Genet. 2001;93:203–206. doi: 10.1159/000056985. [DOI] [PubMed] [Google Scholar]
- Hartmann U, Brummendorf TH, Balabanov S, Thiede C, Illme T, Schaich M. Telomere length and hTERT expression in patients with acute myeloid leukemia correlates with chromosomal abnormalities. Haematologica. 2005;90:307–316. [PubMed] [Google Scholar]
- Heiss NS, Knight SW, Vulliamy TJ, Klauck SM, Wiemann S, Mason PJ, Poustka A, Dokal I. X-linked dyskeratosis congenita is caused by mutations in a highly conserved gene with putative nucleolar functions. Nat Genet. 1998;19:32–38. doi: 10.1038/ng0598-32. [DOI] [PubMed] [Google Scholar]
- Henson JD, Neumann AA, Yeager TR, Reddel RR. Alternative lengthening of telomeres in mammalian cells. Oncogene. 2002;21:598–610. doi: 10.1038/sj.onc.1205058. [DOI] [PubMed] [Google Scholar]
- Hofmann WK, Koeffler HP. Myelodysplastic syndrome. Annu Rev Med. 2005;56:1–16. doi: 10.1146/annurev.med.56.082103.104704. [DOI] [PubMed] [Google Scholar]
- Ip WF, Dupuis A, Ellis L, Beharry S, Morrison J, Stormon MO, Corey M, Rommens JM, Durie PR. Serum pancreatic enzymes define the pancreatic phenotype in patients with Shwachman-Diamond syndrome. J Pediatr. 2002;141:259–265. doi: 10.1067/mpd.2002.125849. [DOI] [PubMed] [Google Scholar]
- Iwama H, Ohyashiki K, Ohyashiki JH, Hayashi S, Kawakubo K, Shay JW, Toyama K. The relationship between telomere length and therapy-associated cytogenetic responses in patients with chronic myeloid leukemia. Cancer. 1997;79:1552–1560. doi: 10.1002/(sici)1097-0142(19970415)79:8<1552::aid-cncr17>3.0.co;2-x. [DOI] [PubMed] [Google Scholar]
- Jeanclos E, Schork NJ, Kyvik KO, Kimura M, Skurnick JH, Aviv A. Telomere length inversely correlates with pulse pressure and is highly familial. Hypertension. 2000;36:195–200. doi: 10.1161/01.hyp.36.2.195. [DOI] [PubMed] [Google Scholar]
- Kajtar P, Mehes K. Bilateral coats retinopathy associated with aplastic anaemia and mild dyskeratotic signs. Am J Med Genet. 1994;49:374–377. doi: 10.1002/ajmg.1320490404. [DOI] [PubMed] [Google Scholar]
- Karadimitris A, Araten DJ, Luzzatto L, Notaro R. Severe telomere shortening in patients with paroxysmal nocturnal hemoglobinuria affects both GPI- and GPI+ hematopoiesis. Blood. 2003;102:514–516. doi: 10.1182/blood-2003-01-0128. [DOI] [PubMed] [Google Scholar]
- Kleideiter E, Bangerter U, Schwab M, Boukamp P, Koscielniak E, Klotz U, Greil J. Telomeres and telomerase in paediatric patients with T-cell acute lymphoblastic leukaemia (T-ALL) Leukemia. 2005;19:296–298. doi: 10.1038/sj.leu.2403596. [DOI] [PubMed] [Google Scholar]
- Koetz K, Bryl E, Spickschen K, O'Fallon WM, Goronzy JJ, Weyand CM. T cell homeostasis in patients with rheumatoid arthritis. Proc Natl Acad Sci USA. 2000;97:9203–9208. doi: 10.1073/pnas.97.16.9203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubuki Y, Suzuki M, Sasaki H, Toyama T, Yamashita K, Maeda K, Ido A, Matsuoka H, Okayama A, Nakanishi T, Tsubouchi H. Telomerase activity and telomere length as prognostic factors of adult T-cell leukemia. Leuk Lymphoma. 2005;46:393–399. doi: 10.1080/10428190400018349. [DOI] [PubMed] [Google Scholar]
- Lahav M, Uziel O, Kestenbaum M, Fraser A, Shapiro H, Radnay J, Szyper-Kravitz M, Avihai S, Hardan I, Shem-Tov N, Nagler A. Nonmyeloablative conditioning does not prevent telomere shortening after allogeneic stem cell transplantation. Transplantation. 2005;80:969–976. doi: 10.1097/01.tp.0000173649.99261.df. [DOI] [PubMed] [Google Scholar]
- Lee JJ, Kook H, Chung IJ, Na JA, Park MR, Hwang TJ, Kwak JY, Sohn SK, Kim HJ. Telomere length changes in patients with aplastic anaemia. Br J Haematol. 2001;112:1025–1030. doi: 10.1046/j.1365-2141.2001.02669.x. [DOI] [PubMed] [Google Scholar]
- Leteurtre F, Li X, Guardiola P, Le Roux G, Sergere JC, Richard P, Carosella ED, Gluckman E. Accelerated telomere shortening and telomerase activation in Fanconi's anaemia. Br J Haematol. 1999;105:883–893. doi: 10.1046/j.1365-2141.1999.01445.x. [DOI] [PubMed] [Google Scholar]
- Li X, Leteurtre F, Rocha V, Guardiola P, Berger R, Daniel MT, Noguera MH, Maarek O, Roux GL, de la Salmoniere P, Richard P, Gluckman E. Abnormal telomere metabolism in Fanconi's anaemia correlates with genomic instability and the probability of developing severe aplastic anaemia. Br J Haematol. 2003;120:836–845. doi: 10.1046/j.1365-2141.2003.04225.x. [DOI] [PubMed] [Google Scholar]
- Lin KW, Yan J. The telomere length dynamic and methods of its assessment. J Cell Mol Med. 2005;9:977–989. doi: 10.1111/j.1582-4934.2005.tb00395.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipton JM. Diamond blackfan anemia: New paradigms for a "not so pure" inherited red cell aplasia. Semin Hematol. 2006;43:167–177. doi: 10.1053/j.seminhematol.2006.04.002. [DOI] [PubMed] [Google Scholar]
- Lipton JM, Federman N, Khabbaze Y, Schwartz CL, Hilliard LM, Clark JI, Vlachos A. Osteogenic sarcoma associated with Diamond-Blackfan anemia: a report from the Diamond-Blackfan Anemia Registry. J Pediatr Hematol Oncol. 2001;23:39–44. doi: 10.1097/00043426-200101000-00009. [DOI] [PubMed] [Google Scholar]
- Lo-Ten-Foe JR, Kwee ML, Rooimans MA, Oostra AB, Veerman AJ, van Weel M, Pauli RM, Shahidi NT, Dokal I, Roberts I, Altay C, Gluckman E, Gibson RA, Mathew CG, Arwert F, Joenje H. Somatic mosaicism in Fanconi anemia: molecular basis and clinical significance. Eur J Hum Genet. 1997;5:137–148. [PubMed] [Google Scholar]
- Marrone A, Sokhal P, Walne A, Beswick R, Kirwan M, Killick S, Williams M, Marsh J, Vulliamy T, Dokal I. Functional characterization of novel telomerase RNA (TERC) mutations in patients with diverse clinical and pathological presentations. Haematologica. 2007;92:1013–1020. doi: 10.3324/haematol.11407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marrone A, Walne A, Dokal I. Dyskeratosis congenita: telomerase, telomeres and anticipation. Curr Opin Genet Dev. 2005;15:249–257. doi: 10.1016/j.gde.2005.04.004. [DOI] [PubMed] [Google Scholar]
- Maser RS, DePinho RA. Connecting chromosomes, crisis, and cancer. Science. 2002;297:565–569. doi: 10.1126/science.297.5581.565. [DOI] [PubMed] [Google Scholar]
- Mathioudakis G, Storb R, McSweeney PA, Torok-Storb B, Lansdorp PM, Brummendorf TH, Gass MJ, Bryant EM, Storek J, Flowers ME, Gooley T, Nash RA. Polyclonal hematopoiesis with variable telomere shortening in human long-term allogeneic marrow graft recipients. Blood. 2000;96:3991–3994. [PubMed] [Google Scholar]
- Matsubara Y, Murata M, Yoshida T, Watanabe K, Saito I, Miyaki K, Omae K, Ikeda Y. Telomere length of normal leukocytes is affected by a functional polymorphism of hTERT. Biochem Biophys Res Commun. 2006;341:128–131. doi: 10.1016/j.bbrc.2005.12.163. [DOI] [PubMed] [Google Scholar]
- McGrath M, Wong JY, Michaud D, Hunter DJ, De Vivo., I Telomere length, cigarette smoking, and bladder cancer risk in men and women. Cancer Epidemiol Biomarkers Prev. 2007;16:815–819. doi: 10.1158/1055-9965.EPI-06-0961. [DOI] [PubMed] [Google Scholar]
- Meeker AK, Argani P. Telomere shortening occurs early during breast tumorigenesis: a cause of chromosome destabilization underlying malignant transformation? J Mammary Gland Biol Neoplasia. 2004;9:285–296. doi: 10.1023/B:JOMG.0000048775.04140.92. [DOI] [PubMed] [Google Scholar]
- Meeker AK, Hicks JL, Iacobuzio-Donahue CA, Montgomery EA, Westra WH, Chan TY, Ronnett BM, De Marzo AM. Telomere length abnormalities occur early in the initiation of epithelial carcinogenesis. Clin Cancer Res. 2004;10:3317–3326. doi: 10.1158/1078-0432.CCR-0984-03. [DOI] [PubMed] [Google Scholar]
- Meeker AK, Hicks JL, Platz EA, March GE, Bennett CJ, Delannoy MJ, De Marzo AM. Telomere shortening is an early somatic DNA alteration in human prostate tumorigenesis. Cancer Res. 2002;62:6405–6409. [PubMed] [Google Scholar]
- Mitchell JR, Wood E, Collins K. A telomerase component is defective in the human disease dyskeratosis congenita. Nature. 1999;402:551–555. doi: 10.1038/990141. [DOI] [PubMed] [Google Scholar]
- Miyata T, Yamada N, Iida Y, Nishimura J, Takeda J, Kitani T, Kinoshita T. Abnormalities of PIG-A transcripts in granulocytes from patients with paroxysmal nocturnal hemoglobinuria. N Engl J Med. 1994;330:249–255. doi: 10.1056/NEJM199401273300404. [DOI] [PubMed] [Google Scholar]
- Nakamura TM, Cech TR. Reversing time: origin of telomerase. Cell. 1998;92:587–590. doi: 10.1016/s0092-8674(00)81123-x. [DOI] [PubMed] [Google Scholar]
- Nawrot TS, Staessen JA, Gardner JP, Aviv A. Telomere length and possible link to X chromosome. Lancet. 2004;363:507–510. doi: 10.1016/S0140-6736(04)15535-9. [DOI] [PubMed] [Google Scholar]
- Notaro R, Cimmino A, Tabarini D, Rotoli B, Luzzatto L. In vivo telomere dynamics of human hematopoietic stem cells. Proc Natl Acad Sci USA. 1997;94:13782–13785. doi: 10.1073/pnas.94.25.13782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Sullivan JN, Bronner MP, Brentnall TA, Finley JC, Shen WT, Emerson S, Emond MJ, Gollahon KA, Moskovitz AH, Crispin DA, Potter JD, Rabinovitch PS. Chromosomal instability in ulcerative colitis is related to telomere shortening. Nat Genet. 2002;32:280–284. doi: 10.1038/ng989. [DOI] [PubMed] [Google Scholar]
- Ohyashiki JH, Iwama H, Yahata N, Ando K, Hayashi S, Shay JW, Ohyashiki K. Telomere stability is frequently impaired in high-risk groups of patients with myelodysplastic syndromes. Clin Cancer Res. 1999;5:1155–1160. [PubMed] [Google Scholar]
- Ohyashiki JH, Ohyashiki K, Fujimura T, Kawakubo K, Shimamoto T, Iwabuchi A, Toyama K. Telomere shortening associated with disease evolution patterns in myelodysplastic syndromes. Cancer Res. 1994;54:3557–3560. [PubMed] [Google Scholar]
- Okuda K, Bardeguez A, Gardner JP, Rodriguez P, Ganesh V, Kimura M, Skurnick J, Awad G, Aviv A. Telomere length in the newborn. Pediatr Res. 2002;52:377–381. doi: 10.1203/00006450-200209000-00012. [DOI] [PubMed] [Google Scholar]
- Ostronoff F, Ostronoff M, Calixto R, Florencio R, Domingues MC, Souto Maior AP, Sucupira A, Tagliari C. Fludarabine, cyclophosphamide, and antithymocyte globulin for a patient with dyskeratosis congenita and severe bone marrow failure. Biol Blood Marrow Transplant. 2007;13:366–368. doi: 10.1016/j.bbmt.2006.11.015. [DOI] [PubMed] [Google Scholar]
- Peng J, Liu C, Zhu K, Zhu Y, Yu Y, Li J, Hou M, Chen X, Xu C, Zhang M. The TNF2 allele is a risk factor to severe aplastic anemia independent of HLA-DR. Hum Immunol. 2003;64:896–901. doi: 10.1016/s0198-8859(03)00141-1. [DOI] [PubMed] [Google Scholar]
- Pipes BL, Tsang T, Peng SX, Fiederlein R, Graham M, Harris DT. Telomere length changes after umbilical cord blood transplant. Transfusion. 2006;46:1038–1043. doi: 10.1111/j.1537-2995.2006.00839.x. [DOI] [PubMed] [Google Scholar]
- Plentz RR, Caselitz M, Bleck JS, Gebel M, Flemming P, Kubicka S, Manns MP, Rudolph KL. Hepatocellular telomere shortening correlates with chromosomal instability and the development of human hepatoma. Hepatology. 2004;40:80–86. doi: 10.1002/hep.20271. [DOI] [PubMed] [Google Scholar]
- Polychronopoulou S, Koutroumba P. Telomere length variation and telomerase activity expression in patients with congenital and acquired aplastic anemia. Acta Haematol. 2004;111:125–131. doi: 10.1159/000076519. [DOI] [PubMed] [Google Scholar]
- Revesz T, Fletcher S, al Gazali LI, DeBuse P. Bilateral retinopathy, aplastic anaemia, and central nervous system abnormalities: a new syndrome? J Med Genet. 1992;29:673–675. doi: 10.1136/jmg.29.9.673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ricca I, Compagno M, Ladetto M, Rocci A, Dell'Aquila M, Omede P, De Marco F, D'Antico S, Caracciolo D, Ferrero D, Carlo-Stella C, Tarella C. Marked telomere shortening in mobilized peripheral blood progenitor cells (PBPC) following two tightly spaced high-dose chemotherapy courses with G-CSF. Leukemia. 2005;19:644–651. doi: 10.1038/sj.leu.2403652. [DOI] [PubMed] [Google Scholar]
- Rigolin GM, Porta MD, Bugli AM, Castagnari B, Mauro E, Bragotti LZ, Ciccone M, Cuneo A, Castoldi G. Flow cytometric detection of accelerated telomere shortening in myelodysplastic syndromes: correlations with aetiological and clinical-biological findings. Eur J Haematol. 2004;73:351–358. doi: 10.1111/j.1600-0609.2004.00305.x. [DOI] [PubMed] [Google Scholar]
- Robertson JD, Testa NG, Russell NH, Jackson G, Parker AN, Milligan DW, Stainer C, Chakrabarti S, Dougal M, Chopra R. Accelerated telomere shortening following allogeneic transplantation is independent of the cell source and occurs within the first year post transplant. Bone Marrow Transplant. 2001;27:1283–1286. doi: 10.1038/sj.bmt.1703069. [DOI] [PubMed] [Google Scholar]
- Rocci A, Ricca I, Dellacasa C, Longoni P, Compagno M, Francese R, Lobetti Bodoni C, Manzini P, Caracciolo D, Boccadoro M, Ferrero D, Ladetto M, Carlo-Stella C, Tarella C. Long-term lymphoma survivors following high-dose chemotherapy and autograft: evidence of permanent telomere shortening in myeloid cells, associated with marked reduction of bone marrow hematopoietic stem cell reservoir. Exp Hematol. 2007;35:673–681. doi: 10.1016/j.exphem.2006.12.006. [DOI] [PubMed] [Google Scholar]
- Roelofs H, de Pauw ES, Zwinderman AH, Opdam SM, Willemze R, Tanke HJ, Fibbe WE. Homeostasis of telomere length rather than telomere shortening after allogeneic peripheral blood stem cell transplantation. Blood. 2003;101:358–362. doi: 10.1182/blood-2002-06-1832. [DOI] [PubMed] [Google Scholar]
- Rothbaum R, Perrault J, Vlachos A, Cipolli M, Alter BP, Burroughs S, Durie P, Elghetany MT, Grand R, Hubbard V, Rommens J, Rossi T. Shwachman-Diamond syndrome: report from an international conference. J Pediatr. 2002;141:266–270. doi: 10.1067/mpd.2002.125850. [DOI] [PubMed] [Google Scholar]
- Rufer N, Brummendorf TH, Chapuis B, Helg C, Lansdorp PM, Roosnek E. Accelerated telomere shortening in hematological lineages is limited to the first year following stem cell transplantation. Blood. 2001;97:575–577. doi: 10.1182/blood.v97.2.575. [DOI] [PubMed] [Google Scholar]
- Rufer N, Brummendorf TH, Kolvraa S, Bischoff C, Christensen K, Wadsworth L, Schulzer M, Lansdorp PM. Telomere fluorescence measurements in granulocytes and T lymphocyte subsets point to a high turnover of hematopoietic stem cells and memory T cells in early childhood. J Exp Med. 1999;190:157–167. doi: 10.1084/jem.190.2.157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rufer N, Dragowska W, Thornbury G, Roosnek E, Lansdorp PM. Telomere length dynamics in human lymphocyte subpopulations measured by flow cytometry. Nat Biotechnol. 1998;16:743–747. doi: 10.1038/nbt0898-743. [DOI] [PubMed] [Google Scholar]
- Savage SA, Calado RT, Xin ZT, Ly H, Young NS, Chanock SJ. Genetic variation in telomeric repeat binding factors 1 and 2 in aplastic anemia. Exp Hematol. 2006a;34:664–671. doi: 10.1016/j.exphem.2006.02.008. [DOI] [PubMed] [Google Scholar]
- Savage SA, Giri N, Baerlocher GM, Orr N, Lansdorp PM, Alter BP. TINF2, a Component of the Shelterin Telomere Protection Complex, is Mutated in Dyskeratosis Congenita. Am J Hum Genet. 2007 doi: 10.1016/j.ajhg.2007.10.004. In Press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savage SA, Stewart BJ, Weksler BB, Baerlocher GM, Lansdorp PM, Chanock SJ, Alter BP. Mutations in the reverse transcriptase component of telomerase (TERT) in patients with bone marrow failure. Blood Cells Mol Dis. 2006b;37:134–136. doi: 10.1016/j.bcmd.2006.07.001. [DOI] [PubMed] [Google Scholar]
- Shay JW, Zou Y, Hiyama E, Wright WE. Telomerase and cancer. Hum Mol Genet. 2001;10:677–685. doi: 10.1093/hmg/10.7.677. [DOI] [PubMed] [Google Scholar]
- Shen J, Terry MB, Gurvich I, Liao Y, Senie RT, Santella RM. Short telomere length and breast cancer risk: a study in sister sets. Cancer Res. 2007;67:5538–5544. doi: 10.1158/0008-5472.CAN-06-3490. [DOI] [PubMed] [Google Scholar]
- Shen JB, Tang JY, Zhao JC, Pan C, Chen J, Zhou X, Wang YP. Telomerase activity and its correlation with the proliferative potential of bone marrow in aplastic anemia in children. Acta Haematol. 2002;107:208–212. doi: 10.1159/000058316. [DOI] [PubMed] [Google Scholar]
- Shimamura A. Inherited bone marrow failure syndromes: molecular features. Hematology Am Soc Hematol Educ Program. 2006a:63–71. doi: 10.1182/asheducation-2006.1.63. [DOI] [PubMed] [Google Scholar]
- Shimamura A. Shwachman-Diamond syndrome. Semin Hematol. 2006b;43:178–188. doi: 10.1053/j.seminhematol.2006.04.006. [DOI] [PubMed] [Google Scholar]
- Sieglova Z, Zilovcova S, Cermak J, Rihova H, Brezinova D, Dvorakova R, Markova M, Maaloufova J, Sajdova J, Brezinova J, Zemanova Z, Michalova K. Dynamics of telomere erosion and its association with genome instability in myelodysplastic syndromes (MDS) and acute myelogenous leukemia arising from MDS: a marker of disease prognosis? Leuk Res. 2004;28:1013–1021. doi: 10.1016/j.leukres.2003.11.020. [DOI] [PubMed] [Google Scholar]
- Slagboom PE, Droog S, Boomsma DI. Genetic determination of telomere size in humans: a twin study of three age groups. Am J Hum Genet. 1994;55:876–882. [PMC free article] [PubMed] [Google Scholar]
- Sznajer Y, Baumann C, David A, Journel H, Lacombe D, Perel Y, Blouin P, Segura JF, Cezard JP, Peuchmaur M, Vulliamy T, Dokal I, Verloes A. Further delineation of the congenital form of X-linked dyskeratosis congenita (Hoyeraal-Hreidarsson syndrome) Eur J Pediatr. 2003;162:863–867. doi: 10.1007/s00431-003-1317-5. [DOI] [PubMed] [Google Scholar]
- Takubo K, Izumiyama-Shimomura N, Honma N, Sawabe M, Arai T, Kato M, Oshimura M, Nakamura K. Telomere lengths are characteristic in each human individual. Exp Gerontol. 2002;37:523–531. doi: 10.1016/s0531-5565(01)00218-2. [DOI] [PubMed] [Google Scholar]
- Tamary H, Alter BP. Current diagnosis of inherited bone marrow failure syndromes. Pediatr Hematol Oncol. 2007;24:87–99. doi: 10.1080/08880010601123240. [DOI] [PubMed] [Google Scholar]
- Thornley I, Dror Y, Sung L, Wynn RF, Freedman MH. Abnormal telomere shortening in leucocytes of children with Shwachman-Diamond syndrome. Br J Haematol. 2002;117:189–192. doi: 10.1046/j.1365-2141.2002.03371.x. [DOI] [PubMed] [Google Scholar]
- Tsakiri KD, Cronkhite JT, Kuan PJ, Xing C, Raghu G, Weissler JC, Rosenblatt RL, Shay JW, Garcia CK. Adult-onset pulmonary fibrosis caused by mutations in telomerase. Proc Natl Acad Sci USA. 2007;104:7552–7557. doi: 10.1073/pnas.0701009104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valdes AM, Andrew T, Gardner JP, Kimura M, Oelsner E, Cherkas LF, Aviv A, Spector TD. Obesity, cigarette smoking, and telomere length in women. Lancet. 2005;366:662–664. doi: 10.1016/S0140-6736(05)66630-5. [DOI] [PubMed] [Google Scholar]
- Vasa-Nicotera M, Brouilette S, Mangino M, Thompson JR, Braund P, Clemitson JR, Mason A, Bodycote CL, Raleigh SM, Louis E, Samani NJ. Mapping of a major locus that determines telomere length in humans. Am J Hum Genet. 2005;76:147–151. doi: 10.1086/426734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vulliamy T, Marrone A, Szydlo R, Walne A, Mason PJ, Dokal I. Disease anticipation is associated with progressive telomere shortening in families with dyskeratosis congenita due to mutations in TERC. Nat Genet. 2004;36:447–449. doi: 10.1038/ng1346. [DOI] [PubMed] [Google Scholar]
- Vulliamy TJ, Knight SW, Mason PJ, Dokal I. Very short telomeres in the peripheral blood of patients with X-linked and autosomal dyskeratosis congenita. Blood Cells Mol Dis. 2001;27:353–357. doi: 10.1006/bcmd.2001.0389. [DOI] [PubMed] [Google Scholar]
- Vulliamy TJ, Marrone A, Knight SW, Walne A, Mason PJ, Dokal I. Mutations in dyskeratosis congenita: their impact on telomere length and the diversity of clinical presentation. Blood. 2006;107:2680–2685. doi: 10.1182/blood-2005-07-2622. [DOI] [PubMed] [Google Scholar]
- Vulliamy TJ, Walne A, Baskaradas A, Mason PJ, Marrone A, Dokal I. Mutations in the reverse transcriptase component of telomerase (TERT) in patients with bone marrow failure. Blood Cells Mol Dis. 2005;34:257–263. doi: 10.1016/j.bcmd.2004.12.008. [DOI] [PubMed] [Google Scholar]
- Walne AJ, Marrone A, Dokal I. Dyskeratosis congenita: a disorder of defective telomere maintenance? Int J Hematol. 2005;82:184–189. doi: 10.1532/IJH97.05067. [DOI] [PubMed] [Google Scholar]
- Walne AJ, Vulliamy T, Marrone A, Beswick R, Kirwan M, Masunari Y, Al Qurashi FH, Aljurf M, Dokal I. Genetic heterogeneity in autosomal recessive dyskeratosis congenita with one subtype due to mutations in the telomerase-associated protein NOP10. Hum Mol Genet. 2007 doi: 10.1093/hmg/ddm111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weng NP, Hathcock KS, Hodes RJ. Regulation of telomere length and telomerase in T and B cells: a mechanism for maintaining replicative potential. Immunity. 1998;9:151–157. doi: 10.1016/s1074-7613(00)80597-x. [DOI] [PubMed] [Google Scholar]
- Woloszynek JR, Rothbaum RJ, Rawls AS, Minx PJ, Wilson RK, Mason PJ, Bessler M, Link DC. Mutations of the SBDS gene are present in most patients with Shwachman-Diamond syndrome. Blood. 2004;104:3588–3590. doi: 10.1182/blood-2004-04-1516. [DOI] [PubMed] [Google Scholar]
- Wu X, Amos CI, Zhu Y, Zhao H, Grossman BH, Shay JW, Luo S, Hong WK, Spitz MR. Telomere dysfunction: a potential cancer predisposition factor. J Natl Cancer Inst. 2003;95:1211–1218. doi: 10.1093/jnci/djg011. [DOI] [PubMed] [Google Scholar]
- Wynn R, Thornley I, Freedman M, Saunders EF. Telomere shortening in leucocyte subsets of long-term survivors of allogeneic bone marrow transplantation. Br J Haematol. 1999;105:997–1001. doi: 10.1046/j.1365-2141.1999.01450.x. [DOI] [PubMed] [Google Scholar]
- Wynn RF, Cross MA, Hatton C, Will AM, Lashford LS, Dexter TM, Testa NG. Accelerated telomere shortening in young recipients of allogeneic bone-marrow transplants. Lancet. 1998;351:178–181. doi: 10.1016/S0140-6736(97)08256-1. [DOI] [PubMed] [Google Scholar]
- Yabe M, Yabe H, Hattori K, Morimoto T, Hinohara T, Takakura I, Shimizu T, Shimamura K, Tang X, Kato S. Fatal interstitial pulmonary disease in a patient with dyskeratosis congenita after allogeneic bone marrow transplantation. Bone Marrow Transplant. 1997;19:389–392. doi: 10.1038/sj.bmt.1700674. [DOI] [PubMed] [Google Scholar]
- Yamaguchi H. Mutations of telomerase complex genes linked to bone marrow failures. J Nippon Med Sch. 2007;74:202–209. doi: 10.1272/jnms.74.202. [DOI] [PubMed] [Google Scholar]
- Yamaguchi H, Baerlocher GM, Lansdorp PM, Chanock SJ, Nunez O, Sloand E, Young NS. Mutations of the human telomerase RNA gene (TERC) in aplastic anemia and myelodysplastic syndrome. Blood. 2003;102:916–918. doi: 10.1182/blood-2003-01-0335. [DOI] [PubMed] [Google Scholar]
- Yamaguchi H, Calado RT, Ly H, Kajigaya S, Baerlocher GM, Chanock SJ, Lansdorp PM, Young NS. Mutations in TERT, the gene for telomerase reverse transcriptase, in aplastic anemia. N Engl J Med. 2005;352:1413–1424. doi: 10.1056/NEJMoa042980. [DOI] [PubMed] [Google Scholar]
- Young NS. Pathophysiologic mechanisms in acquired aplastic anemia. Hematology Am Soc Hematol Educ Program. 2006:72–77. doi: 10.1182/asheducation-2006.1.72. [DOI] [PubMed] [Google Scholar]
- Young NS, Calado RT, Scheinberg P. Current concepts in the pathophysiology and treatment of aplastic anemia. Blood. 2006;108:2509–2519. doi: 10.1182/blood-2006-03-010777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeichner SL, Palumbo P, Feng Y, Xiao X, Gee D, Sleasman J, Goodenow M, Biggar R, Dimitrov D. Rapid telomere shortening in children. Blood. 1999;93:2824–2830. [PubMed] [Google Scholar]