

Abstract
Acetylcholinesterase (AChE) in the echinoid Lytechinus variegatus has been characterized. Kinetic parameters Vmax, Km, Kss, and b were 2594 ± 1048 nmoles ATCh hydrolyzed/min/mg tissue wet weight, 185 ± 11 μM, 308 ± 100 mM, and 0.2, respectively for the substrate ATCh and 17.8 ± 6.87 nmoles BTCh hydrolyzed/min/mg tissue wet weight, 654 ± 424 μM, 36 ± 31 mM, and 0.6, respectively for BTCh. Pharmacologic analyses were performed with four inhibitors of cholinesterases, physostigmine, BW284c51, ethopropazine, and iso-OMPA, and yielded IC50 values of 106 ± 4 nM, 718 ± 118 nM, 2.57 ± 0.6 mM, and > 0.0300 M, respectively. Both kinetic and pharmacologic results confirmed the existence of AChE in larval L. variegatus. Dimeric and tetrameric globular forms (determined by velocity sedimentation on sucrose gradients) were present in L. variegatus larvae. Activity of AChE increased significantly as larvae progressed in development from embryos to eight-arm larvae. Acetylcholinesterase activity of F1 larvae derived from sea urchins collected from wild populations and of F1 larvae derived from sea urchins cultured in the laboratory and fed two different diets suggest that the nutritional and/or environmental history of the adult sea urchin affect the developmental progression of AChE activity in the F1 offspring.
Keywords: Acetylcholinesterase, Nutrition, Sea Urchin, Lytechinus variegatus, Larvae, Acetylcholine
Introduction
Acetylcholinesterase (EC 3.1.1.7) is a cholinesterase found in vertebrates as well as invertebrates. It is a serine hydrolase that rapidly and efficiently hydrolyzes the neurotransmitter acetylcholine (ACh) into choline and acetic acid (Massoulié et al., 1993) within the cholinergic synapses of neuromuscular junctions of the nervous system and is found throughout the central and peripheral nervous systems. Cholinergic molecules are also found in non-neuromuscular tissues and various pre-nervous structures of developing vertebrates and invertebrates (Harrison et al., 2002).
There are two classes of the quaternary structure of AChE: the asymmetric and the globular molecular forms (Massoulié and Rieger, 1969). Asymmetric forms interact with the extracellular matrix by way of the triple-helical-collagen tail. Globular forms differ from the asymmetric forms in both quaternary structure and method of attachment. Globular forms can exist as monomers (G1), dimers (G2), or tetramers (G4). The globular forms do not contain collagen tails; however, globular forms can exist as either membrane-anchored or soluble forms (Gibney and Taylor, 1990; Duval et al., 1992). These various molecular forms are generally identified by their sedimentation coefficients, various structural elements, and solubility characteristics. Vertebrates possess both globular and asymmetric forms of the enzyme; however, asymmetric forms have not been found in invertebrates, suggesting that the collagen tail arose in early vertebrate phylogenetic divergence (Massoulié et al., 1993).
The presence of cholinergic molecules and receptors has been reported in several sea urchin species. ACh and ACh receptors are present in unfertilized sea urchin eggs and other pre-nervous developmental stages (Harrison et al., 2002; Qiao et al., 2003). Augustinsson and Gustafson, pioneers in the study of AChE in sea urchins, observed that cholinesterase (later determined to be AChE) activity is not observed in the unfertilized eggs of sea urchins (Augustinsson and Gustafson, 1949); however, with a more sensitive enzymatic assay (the Ellman assay, Ellman et al. (1961)), Ozaki (1974) has shown that AChE activity is present even in unfertilized Pseudocentrotus depressus eggs. The same assay method was used to obtain similar results revealing AChE activity associated with Strongylocentrotus purpuratus egg ghosts (Barber and Foy, 1973).
AChE activity has been traced throughout the development of several species of sea urchins. These species include P. (Ozaki, 1974), S. purpuratus (Ozaki, 1976), and Hemicentrotus pulcherrimus (Akasaka et al., 1986). Not only is AChE activity present in sea urchin embryos and larvae, a characteristic trend of increasing activity throughout development is observed in various species reported in the literature. ACh and AChE are present in early cleavages of the developing embryo, but sustained increases in the levels of ACh and AChE activity are observed during gastrulation (Falugi et al., 2002; Akasaka et al., 1986) and post-gastrulation (Augustinsson and Gustafson, 1949; Ozaki, 1974; 1976). This rapid increase in AChE activity is thought to be the possible beginnings of neuronal differentiation (Akasaka et al., 1986). Through the use of an AChE staining method, the Cu-thiocholine method of Karnovsky and Roots (1964), Ozaki (1974; 1976) determined that AChE is localized in the mesenchyme cells of sea urchin larvae. The mesenchyme cells are associated with the larval skeleton, oral lobe, and arms (Ozaki, 1974; 1976).
The sea urchin has been proposed as a model organism for neurotoxicity (Buznikov et al., 2001; Qiao et al., 2003; Cunha et al., 2005). Qiao et al. (2003) used the embryos of the sea urchins S. purpuratus and Strongylocentrotus droebachiensis as invertebrate models for developmental neurotoxicity in mammals, focusing on the high-affinity choline transporter, and suggested that the sea urchin has cholinergic structures and activity similar to that found in a mammalian brain. It has also been proposed that the sea urchin be used as a model to test the effects of various pesticides and organic compounds on early development (Buznikov et al., 2001), and the AChE of Paracentrotus lividus has recently been considered as a biomarker of environmental contamination (Cunha et al., 2005).
The primary goals of this research were to characterize the enzyme AChE in the sea urchin Lytechinus variegatus and to evaluate its activity in developing larvae. The goals were accomplished by a kinetic analysis of the enzyme’s substrate specificity and pharmacological inhibition, as well as a determination of the various molecular forms present. Following the characterization, developmental progressions of AChE activity were evaluated in F1 embryos and larvae derived from adult sea urchins either collected from wild populations or cultured in the laboratory on nutritionally-different diets. There have been no studies that investigate the effect of parental nutrition on AChE expression of developing sea urchins F1 embryos and larvae. Although developmental progressions in at least three other cold water sea urchin species have been constructed, L. variegatus is a warm water species that is more readily adapted as an experimental animal model. Thus, a final goal was to evaluate the suitability of AChE in L. variegatus as a biomarker for determining the well-being of developing organisms.
Materials and Methods
Collection and Culture of Sea Urchins for the Characterization of AChE
Adult L. variegatus sea urchins were collected from St. Joseph Bay, Florida in May of 2006 and transported to the University of Alabama at Birmingham (UAB). Individuals were held in recirculating seawater systems containing synthetic seawater (Instant Ocean, 32ppt; 22–24°C) and fed a formulated feed (Hammer, 2006) until analysis. Adult sea urchins were spawned by injection of approximately 1 mL of 0.1 M ACh. Gametes were collected by inverting females over a beaker while sperm was collected dry by removing expressed sperm by pipette. Fertilization tests were performed by obtaining a sample of eggs and fertilizing with a sample of diluted sperm on a microscope slide. Eggs were fertilized with diluted sperm to reduce the possibility for polyspermy. After fertilization, zygotes were placed in a shallow glass fingerbowl in synthetic seawater (32 ± 1 ppt). After the first cell divisions were complete, embryos were placed into a larger volume of aerated synthetic seawater and were fed twice daily combined mixtures of the algae Dunaliella tertiolecta, Isochrysis galbana, and Rhodomonas salina (obtained from the University of Texas, Port Aransas, TX) to apparent satiation (stomachs were observed to be full).
At eight days post-fertilization, a subsample of eight-arm larvae was collected by siphoning excess culture seawater through a Nitex screen (mesh size 75 micron), while retaining the larvae and concentrating the organisms. Once concentrated, the larvae were transferred to several 50 mL centrifuge tubes to further concentrate the sample. Samples were centrifuged at ≤470 g (Beckman TJ-6R Tabletop Centrifuge, TH-4 rotor), depending upon the developmental stage, at 4°C for 10 min. After centrifugation, the supernatant was removed by aspiration and remaining pellets were combined. Synthetic seawater was added to resuspend the combined pellets to a final volume of 25 mL. Organisms in three 50 μL aliquots were counted on a microscope slide to obtain the number of organisms in each sample. Also, a 1 mL aliquot from each sample was preserved in 10% buffered formalin to confirm developmental stage. Samples were re-centrifuged, the supernatant removed by aspiration, and the pellet homogenized with 5 mL of high-ionic strength (HIS) buffer (10 mM NaHPO4, 1 M NaCl, 1 mM EDTA, 1% Triton X-100, pH 7). Samples were then frozen at −30°C until analysis.
Characterization of AchE
Determination of AChE Activity
AChE specific activity was determined by a colorimetric assay as described by Ellman et al. (1961). In this assay, two reactions are coupled to determine AChE activity. AChE hydrolyzes the substrate acetylthiocholine (ATCh) into thiocholine and acetate. Thiocholine then reacts with 5-(3-carboxy-4-nitrophenyl) disulfanyl-2-nitrobenzoic acid (DTNB) to produce a yellow color. The rate of the reaction was determined colorimetrically at 405 nm. This assay is sensitive to low concentrations of AChE and allows for kinetic analysis of the enzyme (Ellman et al., 1961). Samples were homogenized by a Polytron homogenizer and centrifuged at 17,000 g at 4°C for 20 min. In the assay, 50 μL of each supernatant were transferred into individual wells of a 96-well microtitre plate. To each well, 250 μL of Ellman’s solution (pH = 7; final concentrations of components: 100 mM NaHPO4, 2.5 mM DTNB, and 0.833 mM ATCh, 167 mM NaCl, and 0.17% Triton X-100) were added. The colorimetric determination was performed using an ELx808 Ultra Microplate Reader (Biotek Instruments, Inc. Winooski, Vermont).
Similar concentrations of the algae D. tertiolecta, I. galbana, and R. salina were analyzed for the presence of AChE activity. No significant enzyme activity was detected in these samples.
AChE Kinetic Analysis: Substrate Specificity
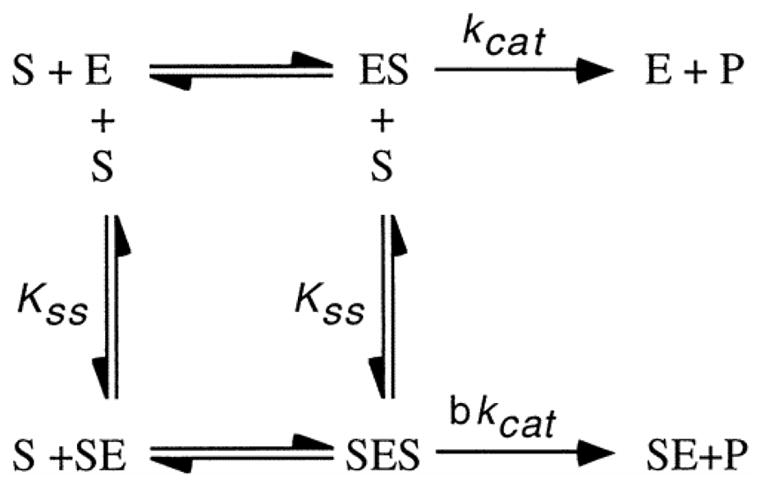
AChE is generally identified by the substrate it will hydrolyze (Augustinsson, 1948). ATCh is hydrolyzed by AChE; however, butyrylthiocholine (BTCh) is not hydrolyzed by AChE. To determine the kinetics of AChE, Ellman’s colorimetric assay was performed separately with various concentrations of the substrates ATCh iodide (minimum 98% TLC, Sigma-Aldrich) and BTCh iodide (minimum 98%, Sigma-Aldrich). The kinetic parameters Vmax, Km, Kss, and b were calculated. AChE activity versus substrate concentration curves were constructed with Sigma Plot software using the scheme and the equation described by Radiæ et al. (1993) to determine kinetic parameters.
In this scheme, substrate (S) can bind to enzyme (E) at two different sites. Kss is the dissociation constant for S in both the SE and SES complexes. The parameter, b, indicates the relative catalytic efficiency of the SES complex compared to SE. If b < 1, the enzyme shows substrate inhibition; if b > 1, the enzyme shows substrate activation; and if b = 1, Michaelis-Menten kinetics is observed.
AChE Kinetic Analysis: Pharmacological Inhibition
Inhibitors of cholinesterases including physostigmine (eserine), an inhibitor of all cholinesterases, BW284c51, a preferential inhibitor of AChE, and iso-OMPA and ethopropazine, preferential inhibitors of BuChE, were tested (Austin and Berry, 1953; Silver, 1974). Dilutions of inhibitors were incubated with the enzyme samples for 20 min at room temperature. The Ellman’s colorimetric enzyme assay was then performed as described previously, except that inhibitor was present in the sample and the assay solution, and plots of the fractional AChE activity versus inhibitor concentration for each inhibitor were constructed with Sigma Plot software. IC50 values were also calculated with Sigma Plot. This method is similar to the original IC50 experiments conducted by Austin and Berry (1953) in their paper describing iso-OMPA and BW284c51, has been used numerous times, and is appropriate for diagnostic purposes, if not detailed kinetic studies (Meedel and Whittaker, 1979; Rotundo, 1984; Toutant et al., 1985; Sanders et al., 1996).
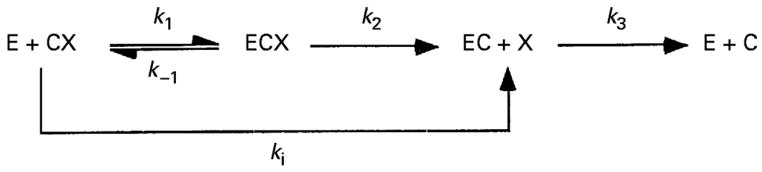
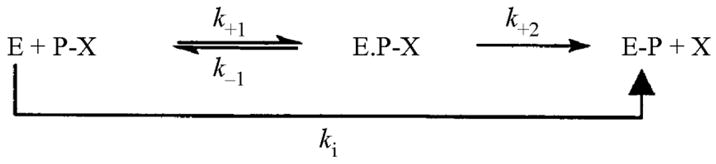
The time course for inhibition by covalent inhibitors of cholinesterases including the carbamates physostigmine and neostigmine, and the organophosphates dichlorvos and paraoxon were also determined. The reaction schemes for carbamylation and phosphonylation of the enzyme are below. In these schemes, CX and PX represent carbamate and organophosphate, respectively. X is the leaving group. Inhibition by carbamates is slowly reversible, while for organophosphates it is essentially irreversible.
Inhibitors (5 μL per well) were incubated with the enzyme sample (45 μL per well) for 0–10 min at room temperature. After incubation, 250 μL of Ellman’s solution were added to each well of a microtitre plate and AChE activity determined. To determine kobs (min−1), the first-order inactivation rate constant, Sigma Plot was used to fit the data to the equation for a single exponential decay (Equation 1):
The apparent bimolecular carbamylation and phosphonylation rate constants (kobs; min−1) were divided by the molar concentration of the inhibitors to obtain ki (M−1 min−1) (Ariel et al., 1998; Ordentlich et al., 1999; Patel et al., 2006).
Determination of Larval Molecular Forms of AChE
The determination of the molecular forms of AChE was completed by velocity sedimentation of the enzyme sample on sucrose gradients as described by Massoulié and Toutant (1988). Homogenized samples and a catalase marker (11.3 S, as described by Rosenberry and Scoggin (1984)) were loaded onto sucrose gradients (5–25% sucrose, 10 mM NaHPO4, pH = 7, 1 mM EDTA, 1mg/mL bovine serum albumin) in the presence and absence of 1% Triton X-100. Samples were centrifuged for 20 h at 35,000 rpm and 4°C (Beckman L7-55 Ultracentrifuge, SW 41 rotor). After centrifugation, each gradient was fractionated and assayed for catalase and AChE activity. Data were analyzed with Sigma Plot to determine the sedimentation coefficients of the various molecular forms. One-way t-tests using SPSS statistical software version 12.0 for Windows (2006; Chicago, Illinois) were performed to determine the significance of the sedimentation coefficients calculated in the presence and absence of Triton X-100.
A sequential extraction of AChE in eight-arm larval L. variegatus followed by velocity sedimentation on sucrose gradients (5–25% sucrose) was performed. Larval L. variegatus were collected as described previously and stored in synthetic seawater at −30°C until analysis. The larval sample was centrifuged at 17,000 g at 4°C for 20 min (Sorvall RC 5C). The supernatant was discarded. An aliquot of 2.5 mL of low ionic strength (LIS) buffer (10 mM NaHPO4, 1 mM EDTA, 1% Triton X-100, pH = 7) with protease inhibitors was added to the pellet. The sample was homogenized with a Polytron homogenizer and centrifuged at 17,000 g at 4°C for 20 min to extract the globular molecular forms of AChE. The supernatant was saved as supernatant one (S1). Another 2.5 mL of LIS buffer were added to the pellet to serve as a wash for any remaining globular forms. The sample was homogenized and centrifuged as before, and the supernatant was saved as supernatant two (S2). Another 2.5 mL of HIS buffer with protease inhibitors were added to the pellet to extract any asymmetric molecular forms. The sample was homogenized and centrifuged as before, and the supernatant was saved as supernatant three (S3). S1, S2, and S3 were assayed for AChE activity. The S2 fraction is normally considered as a wash and not analyzed on gradients (Bon et al., 1979; Rosenberry and Scoggin, 1984).
S1 and S3 were loaded onto sucrose gradients (5–25% sucrose) in the presence of the detergent, Triton X-100 and centrifuged at 35,000 rpm at 4°C for 19 h. Following centrifugation, sucrose gradients were fractionated and assayed for catalase and AChE activity. Sedimentation coefficients were determined by Sigma Plot.
Developmental Progression of AChE Activity in Larval L. variegatus
Adult L. variegatus (ca. 40 mm in diameter) were collected from St. Joseph Bay, Florida in December of 2006. Sea urchins were transported to UAB where they were held as described previously and starved for approximately three months. Sea urchins (n = 32 per treatment) were randomly assigned a diet treatment of either organic romaine lettuce and organic carrots (EPA reference diet (USEPA, 1995; USGS, 2000)) or a nutritionally-complete formulated feed (Hammer, 2006). Sea urchins were photographed and weighed at the beginning and end of the experiment. Sea urchins were placed into mesh baskets partitioned into four individual cages (ca. 11.5 × 11.5 × 15.5 cm). Eight baskets (32 individual cages), elevated by a piece of 3″ PVC pipe split length-wise, were dispersed among four recirculating raceway systems (Jennings, 2007). Sea urchins were maintained in synthetic sea water (Instant Ocean Sea Salt, 32 ± 2 ppt; 23 ± 2°C) with a 12 h light:12 h dark photoperiod. Sea urchins were fed daily ad libitum for ca. 12 weeks (n = 32 individuals per diet treatment; additional diets were tested during this experiment but the results will not be presented here). Water chemistry parameters, including ammonia, nitrate, nitrite, pH, and alkalinity, were monitored biweekly and maintained at appropriate levels.
At week 12, lab-cultured sea urchins fed the lettuce and carrot diet or the formulated feed were spawned as described previously. Fertilization tests for each diet treatment, as described previously, were performed to ensure gametes were viable. There were no noticeable differences in the fertilization efficiencies between diet treatment groups. For each diet treatment, gametes of 3+ females and 3+ males were pooled and fertilized. Various stages of embryological and larval development (1.5, 3, 24, 48, 96, and 192 h post-fertilization) were collected as described previously. Embryos from each diet treatment were not appreciably different in size; however, larvae collected at later developmental stages exhibited slight but noticeable differences in arm length.
Adult L. variegatus were collected from wild populations at approximately the same time as cultured urchins were analyzed. The adult urchins were spawned, a fertilization test was performed, and gametes from one male and one female from the wild were fertilized. Again, there was no noticeable difference in the fertilization efficiency of this feed treatment compared to the two treatments described previously. Various stages of embryological and larval development were collected as described previously. Homogenates of embryos and larvae from both cultured and wild sea urchins were stored at −30°C.
At the time of analysis, previously-collected samples were thawed and evaluated to ensure cell lysis. Advanced larval stages were further homogenized with the Polytron homogenizer. Five 50 μL aliquots of each homogenate were assayed for AChE activity in the presence of 250 μL of Ellman’s solution. AChE activity values (mAb/min) for each developmental stage were normalized to nmoles of ATCh hydrolyzed per min per 104 organisms. Mean values of AChE activity and standard errors for each developmental stage were determined. AChE activity from wild and lab-cultured larvae were compared statistically with SPSS statistical software version 12.0 for Windows (2006; Chicago, Illinois). An analysis of variance (ANOVA) was performed to evaluate the effect of feed treatment on AChE activity at specific post-fertilization times. Significant differences were determined at P < 0.05 and were compared by the Tukey HSD post-hoc test.
Results
AChE Kinetics: Substrate Specificity
ATCh was the preferred substrate for larval L. variegatus cholinesterase; BTCh was not substantially hydrolyzed (Fig. 1). The low VmaxBTCh/VmaxATCh ratio (Table 1) confirmed that only ATCh is hydrolyzed significantly, indicating the homogenate contained AChE. AChE also exhibits substrate inhibition at high substrate concentrations, indicated by a b value of less than one (Fig. 1, Table 1). Estimates of the catalytic efficiency of the enzyme, Vmax/Km, for ATCh and BTCh also show that the enzyme has a marked preference for ATCh.
Fig. 1.

Substrate specificity of AChE. AChE was extracted from L. variegatus larvae with HIS buffer and assayed with ATCh (●) or BTCh (○).
Table 1.
Kinetic analysis of AChE. Kinetic analysis was performed on samples of L. variegatus larval homogenates. Values are means ± SE (n = 4 determinations).
| Parameter (units) | Value |
|---|---|
| VmaxATCh (nmol min−1 mg−1) | 2594 ± 1048 |
| KmATCh (μM) | 185 ± 11 |
| KssATCh (mM) | 308 ± 100 |
| bATCh | 0.20 ± 0.12 |
| VmaxBTCh (nmol min−1 mg−1) | 17.8 ± 6.87 |
| KmBTCh (μM) | 654 ± 424 |
| KssBTCh (mM) | 36 ± 31 |
| bBTCh | 0.60 ± 0.60 |
| VmaxBTCh/VmaxATCh | 0.007 ± 0.007 |
| VmaxATCh/KmATCh | 14.02 |
| VmaxBTCh/KmBTCh | 0.030 |
AChE Kinetics: Pharmacology
Physostigmine (an inhibitor of all cholinesterases) and BW284c51 (a specific inhibitor of AChE) inhibited L. variegatus AChE activity at lower inhibitor concentrations than ethopropazine and iso-OMPA (specific inhibitors of BuChE) (Fig. 2). The enzyme sample assayed independently with physostigmine and BW284c51 exhibited monophasic dose response curves. IC50 values were 106 ± 4 nM (mean ± SE; n = 3 determinations), 718 ± 118 nM, 2.57 ± 0.6 mM, and > 0.03 M for physostigmine, BW284c51, ethopropazine, and iso-OMPA respectively.
Fig. 2.

Pharmacological inhibition of AChE. Four inhibitors of cholinesterases were individually incubated with samples of L. variegatus larval homogenates for 20 min, and Ellman’s assay was performed (□ = physostigmine, ■ = BW284c51, ○ = ethopropazine, ● = iso-OMPA).
The time course for inhibition and the bimolecular rate constants (ki) were determined for two carbamates, physostigmine and neostigmine, and two organophosphates, paraoxon and dichlorvos. The curves were fit to the equation for a first-order exponential decay; higher-order equations did not improve the fits (Fig. 3).
Figure 3.

Inhibition of AChE by carbamates and organophosphates. Two carbamates and 2 organophosphates were individually incubated with samples of 8-arm L. variegatus larval homogenates for 0–10 min, and the Ellman’s assay was performed as described in Materials and Methods [● = physostigmine (1 × 10−6 M, final concentration; ki = 1.77 × 105 ± 1.51 × 104 M−1 min−1; mean ± SE), ○ = neostigmine (3 × 10−6 M, ki = 4.13 × 104 ± 7.27 × 103 M−1 min−1), ■ = dichlorvos (3 × 10−6 M, ki = 2.36 × 104 ± 1.10 × 103 M−1 min−1), □ = paraoxon (1 × 10−3 M, ki = 4.45 × 101 ± 6.05 M−1 min−1), n = 3 for each inhibitor].
Characterization of the Molecular Forms of AChE in L. variegatus
Two peaks of approximately 7.94 S and 10.38 S suggested that G2 and G4 molecular forms are present in L. variegatus larvae (Fig. 4). Average sedimentation coefficients of 7.62 S (SE = 0.17 S, n = 3) and 10.42 (SE = 0.02 S, n = 3) in the presence of Triton X-100 and 8.80 (SE = 0.31 S, n = 3) and 11.46 (SE= 0.40 S, n = 3) in the absence of Triton X-100 for all gradients also suggested that putative G2 and G4 molecular forms are present. There were significant differences between sedimentation coefficients in the presence and absence of Triton X-100. A sequential extraction of the larval L. variegatus resulted in two peaks corresponding to the G2 and G4 forms in the LIS fraction. When HIS extracts were analyzed, there was insufficient AChE activity in each fraction to detect discrete peaks suggesting the absence of asymmetric forms. The percentages of total AChE activity recovered in each sequential extraction were as follows: S1, 68.54%; S2, 15.32%; S3, 16.14%.
Fig. 4.

Determination of larval molecular forms by velocity sedimentation in the presence of Triton X-100. L. variegatus larvae (eight-arm stage) were collected and homogenized in HIS buffer. The larval homogenate and a catalase marker (11.3 S) were loaded onto sucrose gradients (5–25% sucrose) in the presence of Triton X-100. Sucrose gradients were centrifuged at 35,000 rpm for 20 h at 4°C. The gradients were fractionated and assayed for catalase and AChE activity.
Developmental Progression of AChE Activity
Very little AChE activity was observed in pre-gastrulation stages of development. However, there was a substantial increase in AChE activity post-gastrulation, and AChE activity continued to increase exponentially throughout larval development in all groups (Fig. 5). Significant differences between groups were observed throughout all developmental stages.
Fig. 5.

Developmental progression of AChE activity in embryos and larvae obtained from adults collected from wild populations or cultured in the laboratory and fed lettuce/carrots or a formulated feed. Various stages of the same cohort of L. variegatus embryological and larvae were collected and assayed for AChE activity. AChE activity was standardized per 104 organisms for each developmental stage. Values presented are means (n = 4+ determinations). Same letters represent no significant difference in AChE activity among treatments at the various post-fertilization times (ANOVA, P < 0.05; Tukey HSD post-hoc test).
Discussion
Characterization of AChE
Results from the kinetic analysis, including substrate specificity and pharmacology, indicated that L. variegatus express AChE. Analysis of the substrate specificity of larval L. variegatus enzyme homogenates resulted in the preferential hydrolysis of ATCh and virtually no significant hydrolysis of BTCh. Ozaki (1976) also confirmed that enzyme homogenates of S. purpuratus, Strongylocentrotus franciscanus, and Dentraster excentricus were specific for substrate ATCh and not BTCh. Because AChE is specific for its substrate ACh while BuChE hydrolyzes butyrylcholine more efficiently than ACh (Alles and Hawes, 1940; Augustinsson, 1948), the results indicated that AChE is present in both larval and adult (Jennings, 2007) tissues. X-ray analysis of the Pacific electric ray, Torpedo californica, AChE revealed that the enzyme has a very narrow catalytic gorge that is lined with 14 aromatic residues, is approximately 20 Å deep, and has a narrow acyl pocket that restricts the binding of large substrates such as butyrylcholine (Sussman et al., 1991). Fromson and Whittaker (1970) confirmed that cholinesterase from larval homogenates of the ascidian, Ciona intestinalis, preferentially hydrolyzed substrate acetylthiocholine iodide (100% hydrolytic rate) while only hydrolyzing the substrate BTCh at 4.5% of that rate suggesting a low VmaxBTCh/VmaxATCh ratio that is indicative of AChE. Kinetic analysis further indicated that inhibition of L. variegatus AChE occurred at high ATCh concentrations, indicated by a fractional activity (b) value of less than one, which is also characteristic of AChE. Substrate inhibition of AChE activity at high substrate concentrations was also shown in the sea urchin P. lividus (Augustinsson and Gustafson, 1949) and in C. intestinalis (Meedel and Whittaker, 1979). Because of a secondary substrate binding site, the peripheral site, located near the entrance of the catalytic gorge, AChE has the potential for substrate inhibition (Sussman et al., 1991; Massoulié et al., 1993). Furthermore, the kinetic parameters, Km and Kss calculated are similar to those reported by Meedel and Whittaker (1979). These results are indicative that the cholinesterase in the L. variegatus larval homogenate was AChE.
Physostigmine, an inhibitor of all cholinesterases, and BW284c51, a preferential inhibitor of AChE, inhibited the enzyme sample at low inhibitor concentrations, while ethopropazine as well as iso-OMPA, both preferential inhibitors of BuChE, only inhibited enzyme activity at much higher inhibitor concentrations. Comparative studies indicated that the observed inhibition was characteristic of AChE (Fromson and Whittaker, 1970; Silver, 1974). Bisquaternary inhibitors, such as BW284c51, are able to penetrate the deep and narrow catalytic gorge of AChE and in doing so can bind to both the active and peripheral sites, therefore resulting in significant inhibition of AChE (Sussman et al., 1991; Radiæ et al., 1993). Inhibitors of BuChE are too bulky to penetrate the narrow catalytic gorge of AChE, and therefore result in insignificant inhibition. Monophasic dose response curves also suggest that the larval sample contained only one cholinesterase. The first-order decay curves for the carbamates and organophosphates also suggest that a single enzyme is present; however, multiple enzymes with identical activities can not be ruled out as a possibility (Frasco et al., 2006), albeit an unlikely one in our opinion. Multiple candidate genes for AChE have been annotated for the S. purpuratus genome by automated computational analysis (Sea Urchin Genome Sequencing Consortium, 2006) and deposited in GenBank. However, although a detailed sequence analysis is beyond the scope of this paper, most of the sequences are incomplete, and all but one of the complete sequences raise questions about their authenticity as AChEs. Moreover, to our knowledge, none of these genes has been cloned and shown to possess biological activity. Additionally, Cunha et al. (2005) concluded that the sea urchin P. lividus possesses a single cholinesterase activity that hydrolyzes ATCh and proprionylthiocholine (PTCh), but not BTCh; demonstrates substrate inhibition with ATCh and PTCh; and is inhibited by low concentrations of physostigmine and BW284c51, but only high concentrations of iso-OMPA. These are kinetic and pharmacologic characteristics similar to what we have seen for L. variegatus. Thus, the current evidence suggests a single enzyme activity.
Once L. variegatus cholinesterase was confirmed as AChE, the molecular forms of AChE in larvae were determined. Velocity sedimentation of larval L. variegatus AChE samples on sucrose gradients suggested that dimeric (G2) and tetrameric (G4) globular molecular forms are present. Average sedimentation coefficients (S) of L. variegatus AChE were similar to G2 and G4 sedimentation coefficients of Electrophorus (7.7 and 11.8 S, respectively; Bon et al., 1976), Torpedo (7.8 and 11.1 S, respectively; Bon and Massoulié, 1980), bovine (7.2 and 10.9 S, respectively; Bon et al., 1979), and chicken (7.9 and 11.8 S, respectively; Allemand et al., 1981). The change in sedimentation coefficients of both globular forms in the presence and absence of Triton X-100 suggested that the G2 and G4 forms were amphiphilic and interacted with the detergent, which lowered their densities and sedimentation coefficients.
Molecular forms of AChE have been identified in other sea urchin species. Ozaki (1976) reported the presence of a monomer (7.6 S) and dimer (10.6 S) in S. purpuratus, which we now know to be a dimer and a tetramer, respectively. Ozaki (1974) reported the presence of two forms in P. depressus having molecular weights of 180,000 and 280,000. However, only one form was detected in embryos of H. pulcherrimus having a molecular weight of approximately 300,000 (Akasaka et al., 1986). Thus, the number of molecular forms of AChE may vary depending upon species. Akasaka et al. (1986) suggested that the production of the various molecular forms of AChE in sea urchin species is controlled by different mechanisms.
Based on previous reports, only globular molecular forms of AChE were predicted in L. variegatus larval homogenates. Larval and adult (Jennings, 2007) sequential extraction analysis confirmed the apparent lack of asymmetric forms in L. variegatus.
Effect of Parental Nutrition on AChE Activity throughout Sea Urchin Development
AChE activity of F1 embryos and larvae, from adults (F0) obtained from wild populations or from cultured adults fed either a diet of organic lettuce and carrots, or a nutritionally-complete formulated feed, exhibited similar developmental progressions of AChE activity as reported previously in four other sea urchin species (Augustinsson and Gustafson, 1949; Ozaki, 1974; Ozaki, 1976; Akasaka et al., 1986). Unfertilized L. variegatus eggs and pre-gastrulation stages expressed minimal AChE activity. Augustinsson and Gustafson (1949) have shown that AChE activity in unfertilized eggs and other pre-gastrula stages of the sea urchin, P. lividus, is insignificant when compared to advanced plutei. Ozaki (1974), Ozaki (1976), and Akasaka et al. (1986) have all reported very little AChE activity in pre-gastrula stages in P. depressus, S. purpuratus, and H. pulcherrimus, respectively. A significant increase in AChE activity occurred post-gastrulation in all experimental groups of L. variegatus as well as the three previously reported sea urchin species (Ozaki, 1974; Ozaki, 1976; Akasaka et al., 1986). We suggest that the increase in AChE activity at this developmental stage is related to increased motility. Development of the larval arms occurs post-gastrulation and is the stage at which the larvae begin to feed by swimming up through the water column. The exponential increase in AChE activity post-gastrulation would suggest that increased AChE activity is required for the locomotion of the ciliated band, which allows the larvae to swim through the water column and feed (McEdward and Miner, 2007). Nerves have been reported to be associated with the ciliated band and the larval esophagus (Burke, 1978). Histochemical stains of sea urchin larvae have indicated the presence of AChE in regions specialized for locomotion and feeding such as the mesenchyme cells associated with the larval skeleton, the oral lobe, and the anal arms (Ozaki, 1974; Akasaka et al., 1986). Augustinsson and Gustafson (1949) indicated that at 48 h of development, larval P. lividus had a moving intestine and exhibited high AChE activity. The larval intestine has complex ciliary activity and muscular movement which is highly coordinated. They suggested that increased AChE activity may be needed for digestion.
There was a slight but noticeable difference in the arm length of the larvae in the various feed treatments. McEdward and Miner (2007) suggested that arm length is dependent upon the amount of food present. If larvae have been fed poorly, arm length would increase in order to swim more efficiently through the water column to obtain more food. However, larvae from each diet treatment were fed to apparent satiation. There was no indication that this increased arm length in some individuals was a result of poor feeding.
The level of AChE activity throughout larval development in L. variegatus is influenced by the nutritional history of the parents. Larvae of individuals collected from the wild may have different nutrient reserves than offspring of cultured sea urchins, possibly requiring metabolic adjustments. As such, the significantly higher AChE activity levels seen in the wild organisms might be a result of differential locomotion and feeding patterns necessary for survival.
The increased variability of the AChE activity levels seen in the 192 hr developmental stage, in particular the lettuce/carrot diet treatment, could be a result of a difference in the amount of food ingested by the adult sea urchins and the amount of nutrients accumulated.
Environmental factors may also affect differential expression of AChE activity. Parental sea urchins were obtained from the wild only five days before spawning. These individuals had been exposed to changing temperatures, salinities, and/or other seasonal or cyclical factors, resulting in a variable environmental history. Cultured sea urchins were held in the laboratory for six months before spawning in defined and constant conditions of temperature, salinity, and photoperiod. The effects of the environmental history on enzyme expression patterns have not been described and will require further study.
Regardless of the mechanism(s) inducing differential developmental expression of AChE activity, these data suggest a significant plasticity in activity expression patterns. The consequence of plasticity may be adaptive, allowing larvae of different environmental or nutritional histories to adapt, thereby increasing survival and fitness. Expression plasticity may also represent a physiological difference in these offspring, with consequence to the developing organisms. To determine whether this plasticity confers an adaptive advantage or physiological consequence would require further culture and evaluation of the individuals. Besides the levels of AChE reflecting the physiological and environmental conditions, the assembly and distribution of the various molecular forms could also be modulated, as seen in other organisms (Witzemann and Boustead, 1982; Atkins and Pezzementi, 1993; Anselmet et al., 1994; Bertrand et al., 2001; Talesa et al., 2001; Falugi et al., 2002; Romani et al., 2005), which could be another focus of future studies with L. variegatus. Such additional research will help resolve whether the AChE of L. variegatus could be used as an invertebrate biomarker of environmental, nutritional, and toxicological conditions.
Scheme 1.
Enzyme kinetic mechanism for cholinesterases.
Scheme 2.
Reaction scheme for carbamate inhibition.
Scheme 3.
Reaction scheme for organophosphate inhibition.
Acknowledgments
I thank Dr. Mickie Powell for all her guidance and help in culturing sea urchins and algae. I also thank the Texas A&M Mariculture Lab, Port Aransas, Texas for providing the formulated feeds used in this study. Many thanks go to the members of the Watts Lab: Blake Alexander, Adele Cunningham, Heath Garris, Katie Gibbs, Warren Jones, Alicia Kindred, Dorothy Moseley, Anita Patel, Dr. Anthony Siccardi, Kimberly Trawick, and Laura Wright. This project was supported by the Mississippi-Alabama Sea Grant Consortium to SAW and the Academic Research Enhancement Award No. 1 R15 GM072510-01 from the National Institutes of Health to LP.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Akasaka K, Sasaki H, Shimada H, Shiroya T. Purification of acetylcholinesterase from sea urchin (Hemicentrotus pulcherrimus) embryos by affinity chromatography. Dev Grow Differ. 1986;28:85–94. doi: 10.1111/j.1440-169X.1986.00085.x. [DOI] [PubMed] [Google Scholar]
- Allemand P, Bon S, Massoulié J, Vigny M. The quaternary structure of chicken acetylcholinesterase and butyrylcholinesterase; effect of collagenase and trypsin. J Neurochem. 1981;36:860–867. doi: 10.1111/j.1471-4159.1981.tb01673.x. [DOI] [PubMed] [Google Scholar]
- Alles GA, Hawes RC. Cholinesterases in the blood of man. J Biol Chem. 1940;133:375–390. [Google Scholar]
- Anselmet A, Fauquet M, Chatel JM, Maulet Y, Massoulié J, Vallette FM. Evolution of acetylcholinesterase transcripts and molecular forms during the development in the central nervous system of the quail. J Neurochem. 1994;62:2158–2165. doi: 10.1046/j.1471-4159.1994.62062158.x. [DOI] [PubMed] [Google Scholar]
- Ariel N, Ordentlich A, Barak D, Bino T, Velan B, Shafferman A. The aromatic patch of three poximal residues in the human acetylcholinesterase active centre allows for versatile interaction modes with inhibitors. Biochem J. 1998;335:95–102. doi: 10.1042/bj3350095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atkins C, Pezzementi L. Developmental changes in the molecular forms of acetylcholinesterase during the life cycle of the lamprey Petromyzon marinus. Comp Biochem Physiol. 1993;106B:369–372. doi: 10.1016/0305-0491(93)90314-u. [DOI] [PubMed] [Google Scholar]
- Augustinsson KB. Cholinesterases. A study in comparative enzymology. Acta Physiol Scand Suppl. 1948;15:52. [Google Scholar]
- Augustinsson KB, Gustafson T. Cholinesterase in developing sea-urchin eggs. J Cell Comp Physiol. 1949;34:311–321. doi: 10.1002/jcp.1030340208. [DOI] [PubMed] [Google Scholar]
- Austin L, Berry WK. Two selective inhibitors of cholinesterases. Biochem J. 1953;54:695–700. [PMC free article] [PubMed] [Google Scholar]
- Barber ML, Foy JE. An enzymatic comparison of sea urchin egg ghost prepared before and after fertilization. J Exp Zool. 1973;184:157–166. [Google Scholar]
- Bertrand C, Chatonnet A, Takke C, Yan Y, Postlethwait J, Toutant JP, Cousin X. Zebrafish acetylcholinesterase is encoded by a single gene localized on linkage group 7. J Biol Chem. 2001;276:464–474. doi: 10.1074/jbc.M006308200. [DOI] [PubMed] [Google Scholar]
- Bon S, Huet M, Lemonnier M, Rieger F, Massoulié J. Molecular forms of Electrophorus acetylcholinesterase; Molecular weight and composition. Eur J Biochem. 1976;68:523–530. doi: 10.1111/j.1432-1033.1976.tb10840.x. [DOI] [PubMed] [Google Scholar]
- Bon S, Massoulié J. Collagen-tailed and hydrophobic components of acetylcholinesterase in Torpedo marmorata electric organ. Proc Natl Acad Sci USA. 1980;77:4464–4468. doi: 10.1073/pnas.77.8.4464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bon S, Vigny M, Massoulié J. Asymmetric and globular forms of acetylcholinesterase in mammals and birds. Proc Natl Acad Sci (USA) 1979;76:2546–2550. doi: 10.1073/pnas.76.6.2546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burke RD. Structure of the nervous system of pluteus larval of Strongylocentrotus purpuratus. Cell Tis Re. 1978;191:233–247. doi: 10.1007/BF00222422. [DOI] [PubMed] [Google Scholar]
- Buznikov GA, Nikitina LA, Bezuglov VV, Lauder JM, Padilla S, Slotkin TA. An invertebrate model of the developmental neurotoxicity of insecticides: Effects of chlorpyrifos and dieldrin in sea urchin embryos and larvae. Env Health Persp. 2001;109:651–661. doi: 10.1289/ehp.01109651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunha I, Garcia LM, Guilhermino L. Sea-urchin (Paracentrotus lividus) glutathione S-transferases and cholinesterase activities as biomarkers of environmental contamination. J Environ Monit. 2005;7:288–294. doi: 10.1039/b414773a. [DOI] [PubMed] [Google Scholar]
- Duval N, Massoulié J, Bon S. H and T subunits of acetylcholinesterase from Torpedo, expressed in COS cells, generate all types of globular forms. J Cell Biol. 1992;118:641–653. doi: 10.1083/jcb.118.3.641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellman GL, Courtney KD, Andres V, Jr, Featherstone RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmac. 1961;7:88–95. doi: 10.1016/0006-2952(61)90145-9. [DOI] [PubMed] [Google Scholar]
- Falugi C, Diaspro A, Angelini C, Pedrotti ML, Raimondo M, Robello M. Three-dimensional mapping of cholinergic molecules by confocal laser scanning microscopy in sea urchin larvae. Micron. 2002;33(3):233–239. doi: 10.1016/s0968-4328(01)00018-x. [DOI] [PubMed] [Google Scholar]
- Frasco MF, Fournier D, Carvalho F, Guilhermino L. Cholinesterase from the common prawn (Palaemon serratus) eyes: catalytic properties and sensitivity to organophosphate and carbamate compounds. Aquat Toxicol. 2006;77:412–421. doi: 10.1016/j.aquatox.2006.01.011. [DOI] [PubMed] [Google Scholar]
- Fromson D, Whittaker JR. Acetylcholinesterase activity in eserine-treated ascidian embryos. Biol Bull. 1970;139:239–247. doi: 10.2307/1540080. [DOI] [PubMed] [Google Scholar]
- Gibney G, Taylor P. Biosynthesis of Torpedo acetylcholinesterase in mammalian cells. Functional expression and mutagenesis of the glycophospholipid-anchored form. J Biol Chem. 1990;265:12576–12583. [PubMed] [Google Scholar]
- Hammer HS. Biology, vol. Doctor of Philosophy. The University of Alabama; Birmingham, Birmingham: 2006. Determination of dietary protein, carbohydrate, and lipid requirements for the sea urchin Lytechinus variegatus fed semi-purified feeds; p. 172. [Google Scholar]
- Harrison PK, Falugi C, Angelini C, Whitaker MJ. Muscarinic signalling affects intracellular calcium concentration during the first cell cycle of sea urchin embryos. Cell Calcium. 2002;31:289–297. doi: 10.1016/s0143-4160(02)00057-x. [DOI] [PubMed] [Google Scholar]
- Jennings NA. Biology, vol. Master of Science. The University of Alabama; Birmingham, Birmingham: 2007. Acetylcholinesterase in the sea urchin Lytechinus variegatus: characterization and developmental expression in larvae; p. 55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karnovsky MJ, Roots L. A “direct-coloring” thiocholine method for cholinesterases. J Hist Cytol. 1964;12:219–221. doi: 10.1177/12.3.219. [DOI] [PubMed] [Google Scholar]
- Massoulié J, Rieger F. L’acetylcholiesterase des organes electriques de poissons (Torpille et Gymnote) Eur J Biochem. 1969;11:441–455. doi: 10.1111/j.1432-1033.1969.tb00794.x. [DOI] [PubMed] [Google Scholar]
- Massoulié J, Toutant JP. Vertebrate cholinesterases: structure and types of interactions. Handb Exp Pharmacol. 1988;86:167–224. [Google Scholar]
- Massoulié J, Pezzementi L, Bon S, Krejci E, Vallette FM. Molecular and cellular biology of cholinesterases. Prog Neurobiol. 1993;41:31–91. doi: 10.1016/0301-0082(93)90040-y. [DOI] [PubMed] [Google Scholar]
- McEdward LR, Miner BG. Echinoid larval ecology. In: Lawrence JM, editor. Developments in aquaculture and fisheries science, vol. 37. Edible sea urchins 2. Elsevier; Amsterdam: 2007. pp. 71–93. [Google Scholar]
- Meedel TH, Whittaker JR. Development of acetylcholinesterase during embryogenesis of the ascidian Ciona intestinalis. J Exp Zool. 1979;210:1–10. doi: 10.1002/jez.1402100102. [DOI] [PubMed] [Google Scholar]
- Ordentlich A, Barak D, Kronman C, Benschop HP, De Jong LPA, Ariel N, Barak R, Segal Y, Velan B, Shafferman A. Exploring the active center of hyman acetylcholinesterase with stereomers of an organophosphorus inhibitor with two chiral centers. Biochemistry. 1999;38:3055–3066. doi: 10.1021/bi982261f. [DOI] [PubMed] [Google Scholar]
- Ozaki H. Localization and multiple forms of acetylcholinesterase in sea urchin embryos. Dev Grow Differ. 1974;16:267–279. doi: 10.1111/j.1440-169X.1974.00267.x. [DOI] [PubMed] [Google Scholar]
- Ozaki H. Molecular properties and differentiation of acetylcholinesterase in sea urchin embryos. Dev Grow Differ. 1976;18:245–257. doi: 10.1111/j.1440-169X.1976.00245.x. [DOI] [PubMed] [Google Scholar]
- Patel R, Sanders R, Brown L, Baker S, Tsigelny I, Pezzementi L. A tryptophan in the bottleneck of the catalytic gorge of an invertebrate acetylcholinesterase confers relative resistance to carbamate and organophosphate inhibitors. Cell Biochem Biophys. 2006;46:253–264. doi: 10.1385/CBB:46:3:253. [DOI] [PubMed] [Google Scholar]
- Qiao D, Nikitina LA, Buznikov GA, Lauder JM, Seilder FJ, Slotkin TA. The sea urchin embryo as a model for mammalian developmental neurotoxicity: ontogenesis of the high-affinity choline transporter and its role in cholinergic trophic activity. Env Health Persp. 2003;111:1730–1735. doi: 10.1289/ehp.6429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radiæ Z, Pickering N, Vellom DC, Camp S, Taylor P. Three distinct domains in the cholinesterase molecule confer selectivity for acetyl- and butyrylcholinesterase inhibitors. Biochem. 1993;32:12074–12084. doi: 10.1021/bi00096a018. [DOI] [PubMed] [Google Scholar]
- Romani R, Isani G, De Santis A, Giovannini E, Rosi E. Effects of chlorpyrifos on the catalytic efficiency and expression level of acetylcholinesterases in the bivalve mollusk Scapharca inaequivalvis. Environ Toxicol Chem. 2005;24:2879–2886. doi: 10.1897/04-555r3.1. [DOI] [PubMed] [Google Scholar]
- Rosenberry TL, Scoggin DM. Structure of human erythrocyte acetylcholinesterase: characterization of intersubunit disulfide bonding and detergent interaction. J Biol Chem. 1984;259:5643–5652. [PubMed] [Google Scholar]
- Rotundo RL. Purification and properties of the membrane-bound form of acetylcholinesterase from chicken brain. J Biol Chem. 1984;259:13186–13194. [PubMed] [Google Scholar]
- Sanders M, Mathews B, Sutherland D, Soong W, Giles H, Pezzementi L. Biochemical and molecular characterization of acetylcholinesterase from the hagfish Myxine glutinosa. Comp Biochem Physiol. 1996;115B:97–109. doi: 10.1016/0305-0491(96)00088-0. [DOI] [PubMed] [Google Scholar]
- Sea Urchin Genome Sequencing Consortium. The genome of the sea urchin Strongylocentrotus purpuratus. Science. 2006;314:941–952. doi: 10.1126/science.1133609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silver A. The biology of cholinesterases. Elsevier; Amsterdam: 1974. [Google Scholar]
- Sussman JL, Harel M, Frolow F, Oefner C, Goldman A, Toker L, Silman I. Atomic structure of acetylcholinesterase from Torpedo californica: a prototypic acetylcholine-binding protein. Science. 1991;253:872–879. doi: 10.1126/science.1678899. [DOI] [PubMed] [Google Scholar]
- Talesa V, Romani R, Antognelli C, Giovannini E, Rosi G. Soluble and membrane-bound acetylcholinesterases in Mytilus galloprovincialis. Chemico-Biologic Interact. 2001;134:151–166. doi: 10.1016/s0009-2797(01)00152-1. [DOI] [PubMed] [Google Scholar]
- Toutant JP, Massoulié J, Bon S. Polymorphism of pseudocholinesterase in Torpedo marmorata tissues: comparative study of the catalytic and molecular properties of this enzyme with acetylcholinesterase. J Neurochem. 1985;44:580–592. doi: 10.1111/j.1471-4159.1985.tb05452.x. [DOI] [PubMed] [Google Scholar]
- United States Environmental Protection Agency [USEPA] EPA/600/R-95/136. Short-term methods for estimating the chronic toxicity of effluents and receiving waters to west coast marine and estuarine organisms. [accessed on 11/12/06];1995 URL: http://www.epa.gov/EERD/full_doc.pdf.
- United States Geological Survey [USGS] Corpus Christi SOP: F10.7. Sea urchin embryological development toxicity test 2000 [Google Scholar]
- Witzemann V, Boustead C. Changes in acetylcholinesterase molecular forms during the embryonic development of Torpedo marmorata. J Neurochem. 1982;39:747–755. doi: 10.1111/j.1471-4159.1982.tb07956.x. [DOI] [PubMed] [Google Scholar]