

Abstract
Young birds and mammals frequently solicit food by means of extravagant and apparently costly begging displays. Much attention has been devoted to the idea that these displays are honest signals of need, and that their apparent cost serves to maintain their honesty. Recent analyses, however, have shown that the cost needed to maintain a fully informative, honest signal may often be so great that both offspring (signaler) and parent (receiver) would do better to refrain from communication. This apparently calls into question the relevance of the costly signaling hypothesis. Here, I show that this argument overlooks the impact of sibling competition. When multiple signalers must compete for the attention of a receiver (as is commonly the case in parent–offspring interactions), I show that (all other things being equal) individual equilibrium signal costs will typically be lower. The greater the number of competitors, the smaller the mean cost, though the maximum level of signal intensity employed by very needy signalers may actually increase with the number of competitors. At the same time, costs become increasingly sensitive to relatedness among signalers as opposed to relatedness between signalers and receivers. As a result of these trends, signaling proves profitable for signalers under a much wider range of conditions when there is competition (though it is still likely to be unprofitable for receivers).
The young of birds and mammals often solicit food from their parents by means of extravagant and apparently costly begging displays (1, 2). Initial analyses of this phenomenon emphasized scramble competition among sibs as an explanation for the evolution of begging (3–6). More recently, much attention has been devoted to the idea that solicitation displays are reliable signals of offspring need (1, 2, 7). According to the signaling hypothesis, the costly nature of begging reflects the fact that offspring are selected to demand more food than parents are selected to give (2, 8–10), so that a degree of cost is required to prevent chicks from advertising a dishonestly high level of need (7, 11, 12).
Empirical studies have yielded some support for the predictions of signaling models of begging (1). However, recent theoretical analyses have revealed an apparent problem for the hypothesis. The cost required for the maintenance of honesty may often be so great that both offspring (signaler) and parent (receiver) suffer a net loss in fitness as a consequence of communication (13, 14). In other words, although honest and costly signaling is evolutionarily stable once established in a population, both offspring and parents might do better at a nonsignaling equilibrium. This raises questions about how begging can evolve in the first place.
Questions about the accessibility of costly signaling equilibria appear particularly pressing because it has also been shown that cost is not always necessary for the maintenance of honesty. Although a fully informative signal will prove stable only if it is costly, a limited degree of information transfer is possible (between relatives) even in the absence of signal cost (15, 16). Given that fully informative signals will typically be unprofitable, might selection not favor less informative, but cheaper displays?
Here, I investigate this issue further by assessing the impact of sibling competition on the cost of honesty. This is a topic that has not previously been addressed. Although, as early analyses of begging emphasized (2–6), offspring must typically compete with one another for parental attention, most models of the honest signaling of need feature only a single signaler (7, 11–18). Only one signaling model developed by Godfray (19) incorporates sibling competition. Moreover, even this analysis does not allow easy assessment of the impact of competition on overall signal cost, because it assumes that offspring can influence the division of food brought by the parent, but not the total amount delivered. Because models that feature only one signaler focus perforce on the latter possibility, a direct comparison of the two is difficult.
Below, I describe a simple extension of the continuous Philip Sidney game (11, 12), a model that has been widely used to analyze signaling of need. Whereas the basic game features only a single signaler, I allow for any number of competitors. I also assume that, regardless of the number present, signaling can affect both the likelihood of resources being delivered and the offspring to whom they are given. This assumption permits me to investigate the effect of different levels of competition (i.e., different numbers of rival signalers) on the cost required for the maintenance of an honest, fully informative signaling equilibrium.
Competition in the Continuous Philip Sidney Game.
The Philip Sidney game (11), in its original formulation, features two players: a donor and a beneficiary. The former controls some indivisible resource, which he may choose to keep for himself or to give to the beneficiary. Whichever player ends up with the resource gains some additive fitness benefit; the magnitude of this benefit (which may differ for the two players) will be denoted d in the case of the donor and b in the case of the beneficiary. The coefficient of relatedness between the two players will be denoted r.
Each player knows his own level of need (i.e., the amount he stands to gain from acquisition of the resource), but is ignorant as to the need of the other. The only information each possesses about the other’s need is that it is drawn from the probability distribution f(b) or g(d) [the corresponding cumulative probability distributions will be denoted F(b) and G(d)]. The beneficiary, however, may advertise its need to the donor by means of a costly display.
Maynard Smith (11) considered the case in which the beneficiary chooses whether or not to perform a display of fixed, additive cost. Subsequently, Johnstone and Grafen (12) extended his analysis to allow for variation in signal intensity; in their continuous game, the beneficiary chooses how much effort to invest in display, i.e., what level of cost, c, it will expend. The donor, in turn, decides whether to relinquish the resource on the basis of the signal. Johnstone and Grafen (12) focused on the case in which b and d are evenly distributed between 0 and 1; however, as shown by Bergstrom and Lachmann (14), the model can readily be generalized to allow for nonuniform distributions of need. Under a wide range of conditions, this continuous model yields a perfectly informative signaling equilibrium, at which signal cost is a smoothly increasing function of beneficiary need. The donor, at this equilibrium, is able to make the best possible decision in every case, which implies that he will transfer the resource if, and only if, d < rb. Imperfectly informative equilibria are also possible (14–16, 20) but I will not consider these solutions here.
Incorporating competition among signalers.
Let us now suppose that there are n potential beneficiaries to whom the donor can transfer the resource. The amount that each stands to gain from acquisition of the resource, denoted bi for the ith beneficiary, is independently drawn from the same continuous distribution f(b). Each knows its own level of need, but not that of any other beneficiary (or of the donor), and each must make an independent decision as to what level of signal it will employ, with no opportunity for exchange of information among signalers. The donor must then decide whether to transfer the resource, and if it does so, whom to give it to (the resource is assumed to be indivisible and thus cannot be shared among several beneficiaries). The coefficient of relatedness between the donor and each of the beneficiaries will, as before, be denoted r; the coefficient of relatedness between or among different beneficiaries will be denoted s.
Is a perfectly informative signaling equilibrium still possible when there are multiple potential beneficiaries? If so, what form must it take? At such an equilibrium, the donor is able to make the best possible decision in every case, implying that if it transfers the resource, it will do so to the beneficiary who signals most strongly, because signal strength is indicative of the greatest level of need. As in the two-player model, the transfer will take place only if d < rbmax, where bmax denotes the amount that the neediest beneficiary stands to gain from the resource. The expected inclusive fitness of a beneficiary of need b, who adopts the level of signal cost typical of an individual of need b̂ denoted w(b, b̂), is thus given by
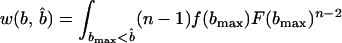 |
1 |
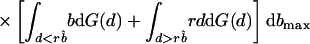 |
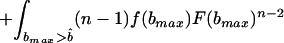 |
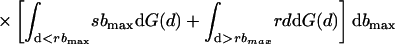 |
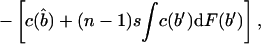 |
where c(b) denotes the level of signal cost typically employed by a beneficiary of need b.
In Eq. 1, bmax denotes the maximum level of need among the other n − 1 beneficiaries. The first two integrals give the expected payoff from possession of the resource. The first covers those cases in which bmax is less than the apparent level of need of the focal beneficiary, i.e., cases in which all of the others employ signals of lower cost. The second covers those cases in which one or more of the other beneficiaries are more needy than the focal individual appears to be, i.e., cases in which one or more adopt signals of greater cost. The integrand in both cases contains two further integrals; the first covers those cases in which the donor transfers the resource, and the second covers those cases in which it does not. Finally, the third integral in Eq. 1 gives the expected cost of begging to the focal beneficiary; the expected indirect cost of begging by its competitors is obtained by integrating over all possible levels of need b′ that each might exhibit.
Now, at equilibrium, we also know that the level of signal cost adopted by a beneficiary of need b must be that which maximizes its expected inclusive fitness. This implies that
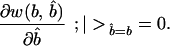 |
2 |
Substituting the expression for w(b, b̂) given in Eq. 1 into the above, we obtain, after some rearrangement, the following differential equation.
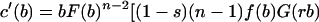 |
3 |
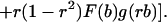 |
To complete our specification of the equilibrium signaling strategy c(b), we require only a starting value. Because beneficiaries of the lowest possible level of need bmin gain nothing from signaling, we may conclude that they will adopt a signal of zero cost, i.e., that
 |
4 |
Together, Eqs. 3 and 4 completely characterize the signaling equilibrium.
An illustrative solution.
To proceed further, one must specify the distributions of donor and beneficiary need, f(b) and g(d). As an illustrative example, I will focus on a case that has received much attention in previous analyses (12, 14, 16), in which b and d are evenly distributed between 0 and 1.
Given the assumption of uniform distributions of need, we can substitute the relevant expressions for f(b), F(b), g(d), G(d) and bmin, into Eqs. 3 and 4 to obtain (by solving the differential equation) the following equilibrium signaling function:
 |
5 |
Having derived this expression for c(b), we can readily calculate mean signal cost (for an individual beneficiary), c̄ which is given by
 |
6 |
It is also easy to derive the expected inclusive fitness of a beneficiary and of a donor at the signaling equilibrium. As the resulting expressions are large and complex, however, I will not give them here. Instead, the results of the model are illustrated graphically in the next section.
Results and Discussion
Fig. 1 shows equilibrium signal cost as a function of beneficiary need, when r = s = 0.5. The different curves give results for different numbers of competing signalers (1, 2, 4, and 8, as indicated on the graph). The figure reveals that increasing levels of competition lead, surprisingly, to a reduction in signal intensity, except among very needy signalers. To understand why, one must focus on the benefits to be gained from a small increase in signal intensity. A slight intensification can have two effects: first, it may cause the donor to transfer rather than keep the resource; second, it may cause a donor with multiple beneficiaries to give the resource to the focal beneficiary rather than to a competitor. As the number of signalers increases, the former possibility becomes less important, and the latter more important; i.e., conflict between donor and beneficiary is gradually overtaken by conflict among beneficiaries.
Figure 1.

Equilibrium signal cost (or intensity) as a function of beneficiary need for different numbers (as indicated on the respective curves) of competing beneficiaries. Relatedness among all individuals is 0.5; i.e., r = s = 0.5.
When there are few beneficiaries, increasing conflict among signalers tends to favor more intense display; signal cost nevertheless decreases as the number of competing signalers rises, because the increase in signaler/signaler competition is outweighed by the decline in signaler/receiver conflict. When there are many beneficiaries, increased conflict among signalers actually leads to a reduction in mean signal intensity; the majority simply “give up” (when there are many competitors, the maximum level of need among them is likely to be high; thus for an individual of moderate need, any slight increase in display is unlikely to capture the resource). The only exception to this trend occurs among signalers of very high need, who signal more strongly when there are many signalers (for such individuals, a slight increase in signal intensity is of little value unless there are many competitors, because the maximum level of need among them is otherwise likely to prove too low to pose a challenge).
Fig. 2 shows the overall mean signal cost incurred by a beneficiary, relative to the probability of resource transfer, as a function of r and of s, for different numbers of competing signalers. The graph clearly reveals that increasing levels of competition not only lead to lower signaling costs, but also to a shift in the relative influence of these two parameters (r and s) on cost. With one beneficiary, signal cost (per benefit transferred) tends to decrease as r (donor–beneficiary relatedness) increases, but is unaffected by s. When multiple beneficiaries compete, signal cost also tends to increase as s (relatedness among the beneficiaries) decreases. Moreover, the greater the number of competitors, the more significant the influence of s becomes relative to that of r. With two competitors, for instance, r and s have approximately equal influence on cost; with eight competitors, by contrast, cost is almost entirely dependent on s, and decreases only slightly with increasing r, reflecting the increasing importance of conflict among beneficiaries relative to conflict between beneficiary and donor.
Figure 2.

Mean signal cost incurred by individual beneficiaries per benefit transferred, and as a function of r (signaler/receiver relatedness) and s (signaler/signaler relatedness) for different numbers of competing beneficiaries. Darker shading indicates lower cost, paler shading indicates higher cost.
Figs. 3 and 4 illustrate the net fitness payoff from signaling to donors and to beneficiaries, respectively. More precisely, each figure shows the range of values of r and s over which either donors or beneficiaries would enjoy higher expected inclusive fitness at the signaling equilibrium than they would in the absence of signaling (if no signal is given, the best decision a donor can make is to transfer the resource to a random beneficiary if, and only if, d < r/2). From Fig. 3, it is clear that donors benefit from signaling only when relatedness is very high (14). Increasing the number of beneficiaries does not alter this conclusion, but merely leads to a shift in the importance of relatedness among beneficiaries compared with relatedness between beneficiary and donor. That is, with few beneficiaries, high values of r make signaling profitable, largely irrespective of s. With many beneficiaries, by contrast, high values of s make signaling profitable, largely irrespective of r. Again, these circumstances reflect the increasing importance of conflict among signalers (as opposed to conflict between signaler and receiver) as the number of beneficiaries increases.
Figure 3.

Profitability of signaling for donors as a function of r (signaler/receiver relatedness) and s (signaler/signaler relatedness) for different numbers of competing beneficiaries. Unshaded regions are those in which donors enjoy higher inclusive fitness at the signaling equilibrium than they would in the absence of signaling. Shaded regions are those in which donor fitness is lower when signals are given.
Figure 4.

Profitability of signaling for beneficiaries as a function of r (signaler/receiver relatedness) and s (signaler/signaler relatedness) for different numbers of competing beneficiaries. Unshaded regions are those in which beneficiaries enjoy higher inclusive fitness at the signaling equilibrium than they would in the absence of signaling. Shaded regions are those in which beneficiary fitness is lower when signals are given.
More surprisingly, Fig. 4 reveals that the incorporation of competition into the model greatly increases the range of conditions under which signaling is profitable for beneficiaries. Signaling is not profitable for a single signaler except at very high values of r (14); signaling is, however, profitable for two signalers when s ≥ 0.5, regardless of the value of r. For larger numbers of competitors, signaling is actually more likely to prove profitable for low values of r, though s continues to exert a greater influence. In general, for plausible levels of relatedness, costly signaling will be profitable for signalers if, and only if, there is competition.
To sum up, the present analysis reveals that the incorporation of competition into a widely used model of signaling of need typically leads to a reduction in the individual signal costs required for the maintenance of an honest, fully informative signal; it can also result in an increase in the information to be gained. Consequently, such a signal is far more likely to prove profitable (for signalers, at least). This surprising result suggests that when signalers compete, it may prove much easier for a costly signaling system to become established and sustained [see Rodriguez-Girones et al. (9) for a related suggestion].
This result does not imply that the possibility of cheaper, partially informative signals can be ignored. Given that many different signaling equilibria (that vary widely in cost and in the degree of information conveyed) are possible, still to be determined are the more likely endpoints for an evolving population (20, 21). Simulation of the dynamics of signal evolution may offer some insights into the problem (21), although the results of such analyses are likely to depend heavily on the details of the simulation process (21, 22). As with game-theoretical studies of signaling equilibria, however, sibling competition has not yet been incorporated into simulations of signal dynamics (but see refs. 13, 21, and 22 for discussions of the possibility). The main conclusion to be drawn from this paper is that future analyses of signaling of need are more likely to be successful in explaining the origin and maintenance of costly solicitation displays when they take into account the competitive nature of begging.
Acknowledgments
I thank Rebecca Kilner, Hanna Kokko, Geoff Parker, and Miguel Rodríguez-Gironés for helpful comments and discussion.
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
References
- 1.Kilner R, Johnstone R A. Trends Ecol Evol. 1997;12:11–15. doi: 10.1016/s0169-5347(96)10061-6. [DOI] [PubMed] [Google Scholar]
- 2.Mock D W, Parker G A. The Evolution of Sibling Rivalry. Oxford: Oxford Univ. Press; 1997. [Google Scholar]
- 3.Stamps J, Metcalfe R A, Krishnan V V. Behav Ecol Sociobiol. 1978;3:369–372. [Google Scholar]
- 4.Macnair M, Parker G A. Anim Behav. 1979;27:1202–1209. [Google Scholar]
- 5.Parker G A. Anim Behav. 1985;33:519–533. [Google Scholar]
- 6.Harper A B. Am Nat. 1986;128:99–114. [Google Scholar]
- 7.Godfray H C J. Nature (London) 1991;352:328–330. [Google Scholar]
- 8.Trivers R L. Am Zool. 1974;14:249–264. [Google Scholar]
- 9.Mock D W, Forbes L S. Trends Ecol Evol. 1992;7:409–413. doi: 10.1016/0169-5347(92)90022-4. [DOI] [PubMed] [Google Scholar]
- 10.Godfray H C J. Nature (London) 1995b;376:1133–1138. [Google Scholar]
- 11.Maynard Smith J. Anim Behav. 1991;42:1034–1035. [Google Scholar]
- 12.Johnstone R A, Grafen A. J Theor Biol. 1992;156:215–234. doi: 10.1016/s0022-5193(05)80674-5. [DOI] [PubMed] [Google Scholar]
- 13.Rodríguez-Gironés M A, Cotton P A, Kacelnik A. Proc Natl Acad Sci USA. 1996;93:14637–14641. doi: 10.1073/pnas.93.25.14637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bergstrom C T, Lachmann M. Phil Trans R Soc London Ser B. 1997;352:609–617. [Google Scholar]
- 15.Maynard Smith J. Anim Behav. 1994;47:1115–11120. [Google Scholar]
- 16.Bergstrom C T, Lachmann M. Proc Natl Acad Sci USA. 1998;95:5100–5105. doi: 10.1073/pnas.95.9.5100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Johnstone R A. Proc R Soc London Ser B. 1996;263:1677–1681. [Google Scholar]
- 18.Johnstone R A. Am Nat. 1998;152:45–58. doi: 10.1086/286148. [DOI] [PubMed] [Google Scholar]
- 19.Godfray H C J. Am Nat. 1995a;146:1–24. [Google Scholar]
- 20.Lachmann M, Bergstrom C T. Theor Popul Biol. 1998;54:146–160. doi: 10.1006/tpbi.1997.1372. [DOI] [PubMed] [Google Scholar]
- 21.Rodríguez-Gironés M A, Enquist M, Cotton P A. Proc Natl Acad Sci USA. 1998;95:4453–4457. doi: 10.1073/pnas.95.8.4453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Payne R J H, Rodríguez-Gironés M A. J Theor Biol. 1998;195:273–279. doi: 10.1006/jtbi.1998.0798. [DOI] [PubMed] [Google Scholar]