

Abstract
The developmental neurotoxicity of organophosphate pesticides targets serotonin (5HT) systems, which are involved in emotional and appetitive behaviors. We exposed neonatal rats to daily doses of diazinon on postnatal days 1–4, using doses (0.5 or 2 mg/kg) spanning the threshold for barely-detectable cholinesterase inhibition. We then evaluated the effects on 5HT1A and 5HT2 receptors, and on the 5HT transporter in cerebral cortical regions and the brainstem in adolescence through adulthood. Diazinon evoked a lasting deficit in 5HT1A receptors in males only, whereas it caused a small but significant increase in 5HT transporters in females; neither effect showed a significant regional selectivity. This pattern differed substantially from that seen in earlier work with another organophosphate, chlorpyrifos, which at pharmacodynamically similar doses spanning the threshold for cholinesterase inhibition, evoked a much more substantial, global upregulation of 5HT receptor expression; with chlorpyrifos, effects on receptors were seen in females, albeit to a lesser extent than in males, and were also regionally distinct. The effects of diazinon were nonmonotonic, showing larger alterations at the lower dose, likely reflecting positive trophic effects of cholinergic stimulation once the threshold for cholinesterase inhibition is exceeded. Our results reinforce the idea that different organophosphates have fundamentally distinct effects on the developmental trajectories of specific neurotransmitter systems, unrelated to their shared action as cholinesterase inhibitors. The effects on 5HT circuits expands the scope of behavioral endpoints that need to be considered in evaluating the developmental neurotoxicity of organophosphates.
Keywords: Brain development, Diazinon, Organophosphate insecticides, Serotonin receptors, Serotonin transporter
INTRODUCTION
Organophosphate pesticides are undergoing increasing scrutiny because of their propensity to elicit developmental neurotoxicity at lower exposures than those which cause overt symptoms of intoxication, or even below the threshold for cholinesterase inhibition, the biomarker most commonly used for exposure and risk assessment [12,14,29,30,38,53–55,65–67,86]. Indeed, a wealth of information now shows that these agents disrupt neural cell replication and differentiation, interfere with axonogenesis and synaptogenesis, and impair the functional development of neurotransmitter and neurotrophin systems, culminating in aberrant behavioral performance [9–12,19,55–57,65–67,77,92]. Consequently, the organophosphates produce developmental damage extending far beyond acetylcholine systems, notably including serotonin (5HT), which appears to be particularly sensitive to disruption by fetal or neonatal organophosphate exposure [1–5,58,64,72–75]. In keeping with the known role of 5HT abnormalities in affective disorders [49,50], rats exposed to low doses of chlorpyrifos as neonates show depression-like behavioral patterns [1]; further, a clear connection appears to be emerging between human organophosphate exposure and depression and suicide [32,36].
If the developmental neurotoxicity of organophosphates resides in mechanisms other than their shared ability to inhibit cholinesterase, then it is likely that the various members of this class might evoke dissimilar effects reflecting other mechanisms. We recently compared the ability of three different organophosphates, chlorpyrifos, diazinon and parathion, to elicit immediate changes in 5HT systems after exposure of neonatal rats to doses spanning the threshold for barely-detectable, nonsymptomatic inhibition of cholinesterase [4,73,79]. Although both chlorpyrifos and diazinon evoked an immediate increase in the concentration of 5HT1A and 5HT2 receptors at these low doses, parathion evoked a decrease in the 5HT1A subtype, confirming disparate actions of the three agents. Furthermore, when we examined expression patterns for the gene families encoding the 5HT biosynthetic enzymes, transporters and receptors, we also found major divergence between chlorpyrifos and diazinon, especially for the 5HT1A and 5HT2 receptor subtypes [73], suggesting that functional differences might emerge later. Accordingly, in the present study, we examined the long-term effects of neonatal diazinon exposure on the developmental profile of these receptors and the 5HT transporter (5HTT) in adolescence through adulthood, the period when lasting changes emerged in our earlier studies with chlorpyrifos [1–3,5,72,74,75]. We administered diazinon during the immediate postnatal period (postnatal days PN1-4), a stage where we previously found high sensitivity of 5HT systems to disruption by chlorpyrifos [2,4,5,75]. We evaluated two nonsymptomatic diazinon regimens [69,73,79], 0.5 mg/kg/day, which produces no discernible cholinesterase inhibition, and 2 mg/kg/day, which elicits approximately 20% inhibition, equivalent to that obtained with 1 mg/kg/day of chlorpyrifos as used in our earlier work [81]. Because the effects of chlorpyrifos on 5HT systems are strongly sex-selective [1,3,5,72], we evaluated both males and females for comparable effects of diazinon. Measurements were conducted for 5HT1A and 5HT2 receptors, which converge on common endpoints in 5HT cell signaling [8,47,63] and are key players in 5HT-related mental disorders, particularly depression [7,17,93,94]. In addition, we assessed binding to the 5HTT site, which regulates the synaptic concentration of 5HT and is the major target for antidepressant drugs [37,49,50]. Evaluations were conducted in the forebrain, which contains a high concentration of 5HT projections, and in the brainstem, which contains the corresponding 5HT cell bodies.
METHODS
Animal treatments
All experiments were carried out humanely and with regard for alleviation of suffering, with protocols approved by the Institutional Animal Care and Use Committee and in accordance with all federal and state guidelines. Timed-pregnant Sprague–Dawley rats (Charles River, Raleigh, NC) were housed in breeding cages, with a 12 h light–dark cycle and free access to food and water. On the day after birth, all pups were randomized and redistributed to the dams with a litter size of 10 (5 males, 5 females) to maintain a standard nutritional status. Because of its poor water solubility, diazinon (Chem Service, West Chester, PA) was dissolved in dimethylsulfoxide to provide consistent absorption [69,73,79,89] and was injected subcutaneously in a volume of 1 ml/kg once daily on postnatal days (PN) 1–4; control animals received equivalent injections of the dimethylsulfoxide vehicle, which does not itself produce developmental neurotoxicity [89]. Doses of 0.5 and 2 mg/kg/day were chosen because they lie below the threshold for signs of systemic toxicity in developing rats as evidenced by impaired viability or reduced weight gain [69] and they straddle the threshold for barely-detectable cholinesterase inhibition [73,79]. These treatments thus resemble the nonsymptomatic exposures reported in pregnant women [16] and are pharmacodynamically comparable to expected fetal and childhood exposures after routine home application or in agricultural communities [20,51]. Randomization of pup litter assignments within treatment groups was repeated at intervals of several days up until weaning, and in addition, dams were rotated among litters to distribute any maternal caretaking differences randomly across litters and treatment groups. Offspring were weaned on PN21.
On PN30, 60 and 100, one male and one female were selected from each litter of origin and were decapitated. The cerebellum (including flocculi) was removed and the midbrain/brainstem was separated from the forebrain by a cut rostral to the thalamus. The striatum and hippocampus were then dissected from these larger divisions and the midbrain and brainstem were divided from each other. The cerebral cortex was divided down the midline and then further sectioned into anterior and posterior regions (frontal/parietal cortex and temporal/occipital cortex, respectively). The current studies were performed on the frontal/parietal cortex and temporal/occipital cortex, which contain the major cerebrocortical 5HT projections, and the brainstem, which contains 5HT cell bodies; the remaining regions were reserved for future work. Tissues were frozen with liquid nitrogen and stored at −45° C.
Assays
Assays were conducted on each individual tissue, so that each determination represented a value from the corresponding brain region of one animal. Each tissue was thawed and homogenized (Polytron, Brinkmann Instruments, Westbury, NY) in ice-cold 50 mM Tris (pH 7.4), and aliquots of the homogenate were withdrawn for measurement of total protein [80]. The remaining homogenate was sedimented at 40,000 × g for 15 min and the resultant pellet was washed by resuspension (Polytron) in homogenization buffer followed by resedimentation, and was then dispersed with a homogenizer (smooth glass fitted with Teflon pestle) in 50 mM Tris buffer (pH 7.4). An aliquot was withdrawn for the determination of membrane protein [80]. Two radioligands were used to determine 5HT receptor binding [91]: 1 nM [3H]8-hydroxy-2-(di-n-propylamino)tetralin (PerkinElmer Life Sciences, Boston, MA; specific activity, 135 Ci/mmol) for 5HT1A receptors [52,82], and 0.4 nM [3H]ketanserin (PerkinElmer; specific activity, 63 Ci/mmol) for 5HT2 receptors [35,52]. For 5HT1A receptors, incubations lasted for 30 min at 25°C in a buffer consisting of 50 mM Tris (pH 8), 2 mM MgCl2 and 2 mM sodium ascorbate; 100 μM 5HT (Sigma) was used to displace specific binding. For 5HT2 receptors, incubations lasted 15 min at 37°C in 50 mM Tris (pH 7.4) and specific binding was displaced with 10 μM methylsergide (Sandoz Pharmaceuticals, E. Hanover, NJ). Incubations were stopped by the addition of a large excess of ice-cold buffer and the labeled membranes were trapped by rapid vacuum filtration onto glass fiber filters that were pre-soaked in 0.15% polyethyleneimine (Sigma). The filters were then washed repeatedly and radiolabel was determined. For binding to the presynaptic 5HTT [46,70,71,76,90], the membrane suspension was incubated with 85 pM [3H]paroxetine (PerkinElmer; specific activity 19.4 Ci/mmol) with or without addition of 100 μM 5HT to displace specific binding, and incubations lasted 120 min at 20° C. Binding was calculated relative to membrane protein.
Data analysis
Data were compiled as means and standard errors. Because we evaluated multiple neurochemical variables that were all related to 5HT synapses, the initial comparison was conducted by a global ANOVA (data log-transformed because of heterogeneous variance among ages, regions and measures) incorporating all the variables and measurements so as to avoid an increased probability of type 1 errors that might otherwise result from multiple tests of the same data set: treatment, age, sex, region and the three repeated measures (5HT1A receptors, 5HT2 receptors, 5HTT). Where we identified interactions of treatment with the other variables, data were then subdivided for lower-order ANOVAs to evaluate individual treatments that differed from the corresponding control. Significance for all tests was assumed at the level of p < 0.05. For convenience, some of the results are presented as the percent change from control values but statistical comparisons were conducted only on the original data. For reference, the corresponding control values are shown in Table 1.
TABLE 1.
Control Values for 5HT Markers
| Brain region | Age | 5HT1A Receptors | 5HT2 Receptors* | 5HT Transporter | |||
|---|---|---|---|---|---|---|---|
| male | female | male | female | male | female | ||
| Frontal/parietal cortex | PN30 | 104 ± 5 | 111 ± 2 | 125 ± 4 | 137 ± 3 | 277 ± 7 | 291 ± 10 |
| PN60 | 68 ± 7 | 68 ± 7 | 116 ± 3 | 126 ± 3 | 339 ± 14 | 343 ± 9 | |
| PN100 | 56 ± 2 | 49 ± 4 | 88 ± 2 | 93 ± 2 | 299 ± 9 | 287 ± 6 | |
| Temporal/occipital cortex | PN30 | 138 ± 4 | 124 ± 5 | 69 ± 2 | 75 ± 2 | 199 ± 9 | 187 ± 7 |
| PN60 | 99 ± 5 | 103 ± 6 | 51 ± 3 | 55 ± 2 | 207 ± 7 | 203 ± 10 | |
| PN100 | 92 ± 7 | 99 ± 7 | 52 ± 2 | 61 ± 3 | 220 ± 13 | 231 ± 10 | |
| Brainstem | PN30 | 38 ± 2 | 31 ± 2 | 19 ± 1 | 20 ± 1 | 276 ± 14 | 290 ± 7 |
| PN60 | 22 ± 1 | 24 ± 1 | 16 ± 1 | 15 ± 1 | 251 ± 14 | 244 ± 8 | |
| PN100 | 18 ± 1 | 19 ± 1 | 13 ± 1 | 14 ± 1 | 234 ± 6 | 257 ± 6 | |
Data are means and standard errors obtained from 6 animals of each sex at each age.
Females significantly higher than males; main effect of sex, p < 0.0001.
RESULTS
Neonatal diazinon exposure did not cause any significant deficits in body or brain region weights on PN30, 60 or 100 (data not shown). Nevertheless, there were significant overall effects on 5HT synaptic proteins, as revealed by a global ANOVA incorporating all variables and measures: p < 0.0002 for the interaction of treatment × sex, p < 0.04 for treatment × age, and p < 0.05 for treatment × sex × measure. Since the most robust interaction was with sex, data were subdivided for males and females, both of which showed significant effects of diazinon exposure (males, main treatment effect, p < 0.02; females, main treatment effect, p <0.02). For presentation, each measure was evaluated separately in males and females, in light of the interactions seen in the global test.
For 5HT1A receptors, males exposed to the lower dose of diazinon showed a significant overall decrement of about 10–20% whereas the higher dose produced no consistent alterations (Fig. 1A); the effect in the low dose group was statistically distinguishable from the lack of effect in the high dose group (p < 0.05). In contrast, no statistically significant effects were seen in females who, if anything, showed a tendency toward an increase rather than the decrease seen in males (p < 0.008 for the interaction of treatment × sex in the low dose group, Fig. 1B). For 5HT2 receptors, neither males (Fig. 2A) nor females (Fig. 2B) showed statistically significant alterations. Similarly, males did not display any consistent changes in 5HTT sites (Fig. 3A), but females exhibited a small, statistically significant increase in the group given 0.5 mg/kg diazinon (Fig. 3B). Again, the dose-effect relationship was nonmonotonic, since no increase over control values was seen with the higher dose of diazinon; the low dose group was significantly different (p < 0.05) from the high dose group.
Figure 1.
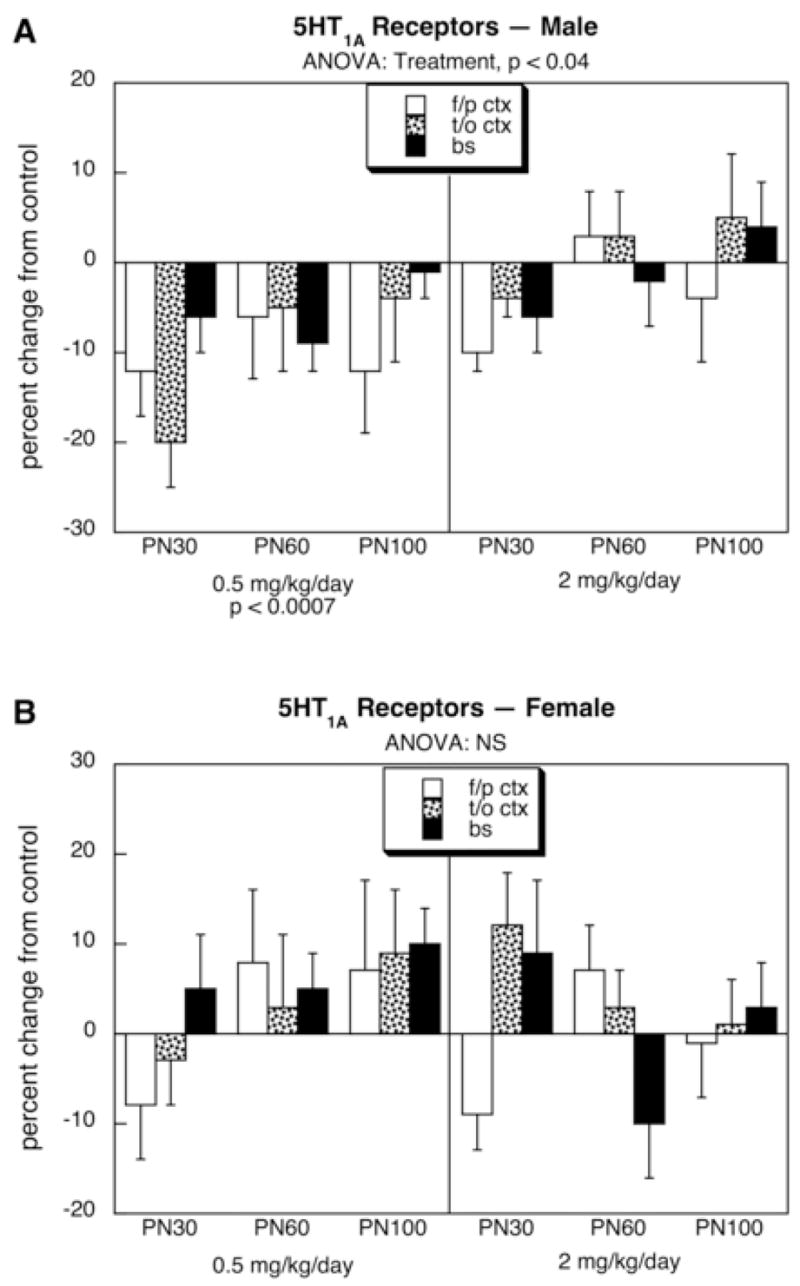
Effects of neonatal diazinon exposure on 5HT1A receptors in males (A) and females (B). Data represent means and standard errors obtained from six animals of each sex in each treatment group at each age, presented as the percentage change from control values shown in Table 1. ANOVA across all variables (all treatments, both sexes, all ages, all regions) indicates an interaction of treatment × sex (p < 0.02); accordingly, lower-order ANOVAs for each sex appear at the top of the panels and significance for individual treatments are at the bottom. Abbreviation: NS, not significant.
Figure 2.
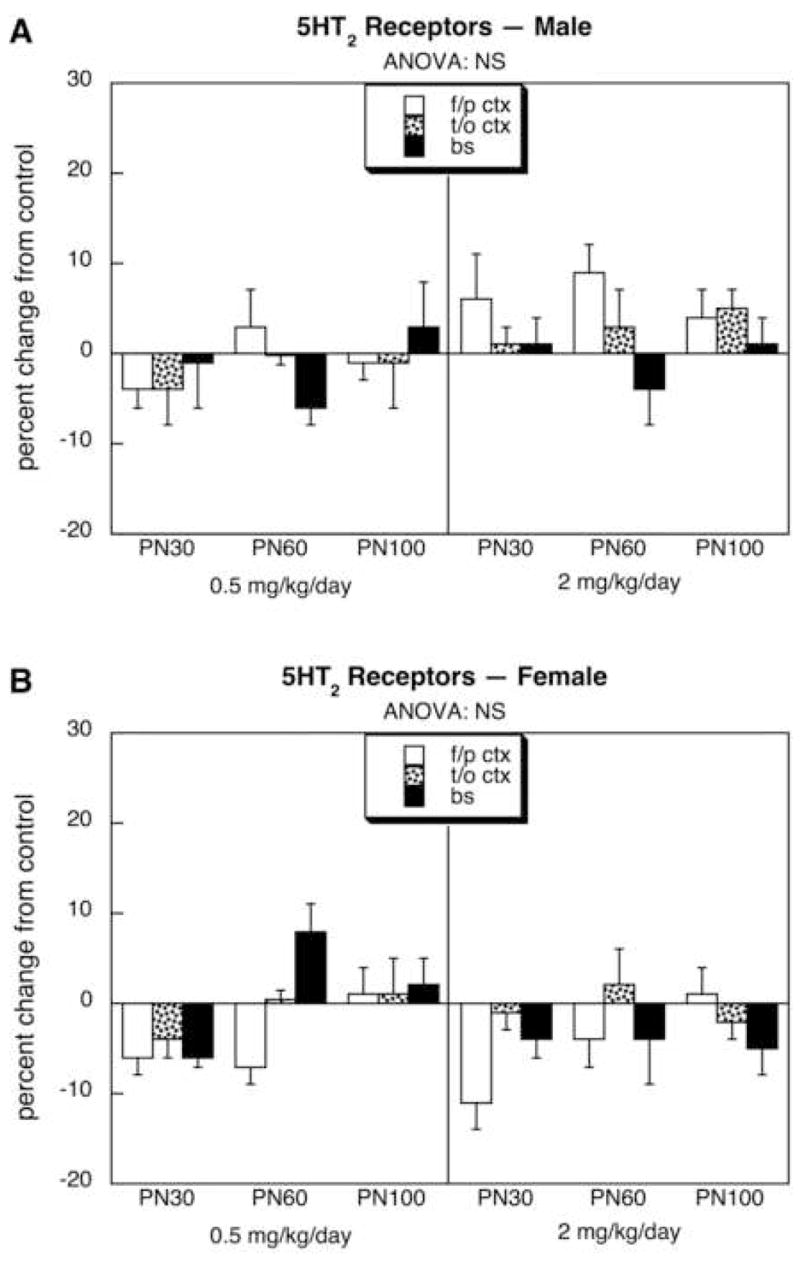
Effects of neonatal diazinon exposure on 5HT2 receptors in males (A) and females (B). Data represent means and standard errors obtained from six animals of each sex in each treatment group at each age, presented as the percentage change from control values shown in Table 1. ANOVA across all variables (all treatments, both sexes, all ages, all regions) indicates an interaction of treatment × sex (p < 0.008); accordingly, lower-order ANOVAs for each sex appear at the top of the panels. Abbreviation: NS, not significant.
Figure 3.
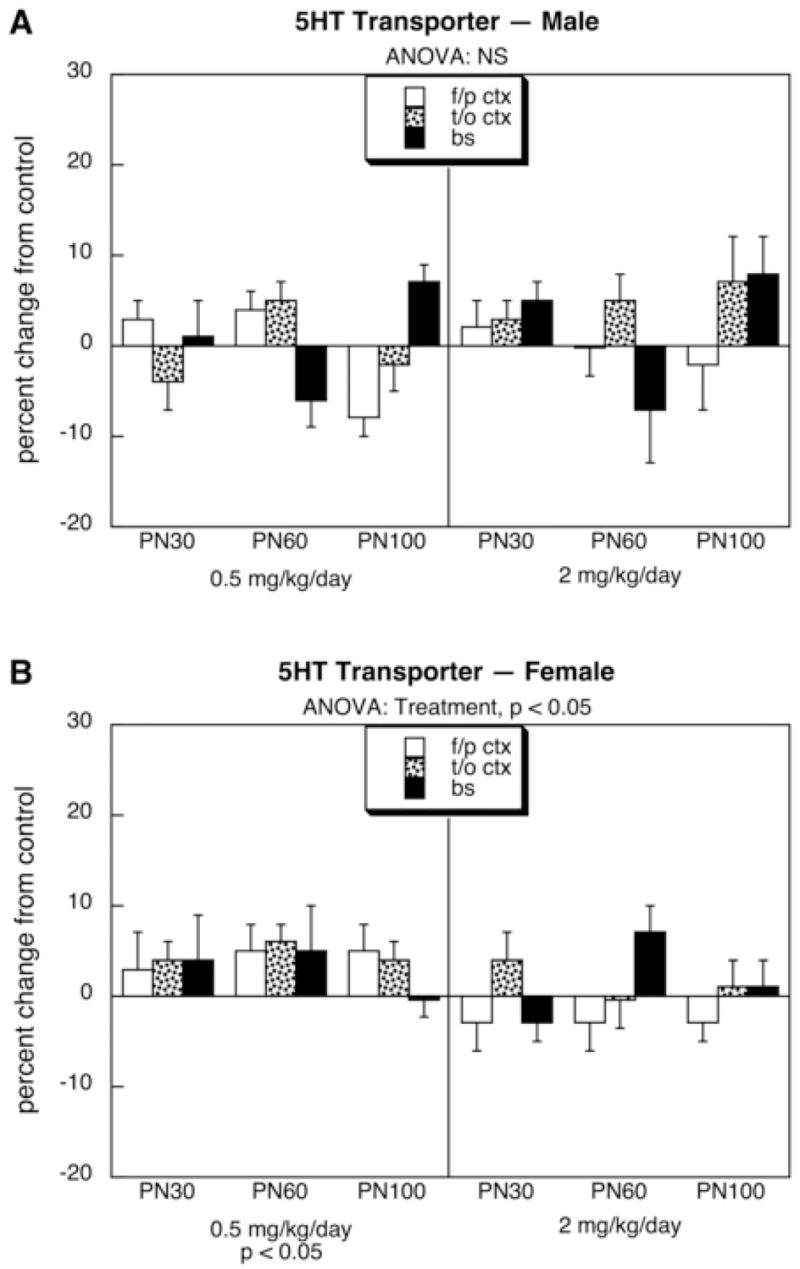
Effects of neonatal diazinon exposure on the 5HT transporter in males (A) and females (B). Data represent means and standard errors obtained from six animals of each sex in each treatment group at each age, presented as the percentage change from control values shown in Table 1. ANOVA across all variables (all treatments, both sexes, all ages, all regions) indicates an interaction of treatment × sex (p < 0.02); accordingly, lower-order ANOVAs for each sex appear at the top of the panels and significance for individual treatments are at the bottom. Abbreviation: NS, not significant.
DISCUSSION
In our earlier work with neonatal chlorpyrifos exposure, we found permanent deficits in 5HT neurotransmission, likely reflecting an underlying “miswiring” of 5HT circuits [1,2,5,72]. Consequently, 5HT receptors were globally upregulated and 5HT presynaptic activity was elevated but 5HT-related behaviors remained deficient nevertheless. Here, we performed a parallel study of the effects of diazinon, using doses that, as with our earlier work, lie below or just above the threshold for cholinesterase inhibition [79,81] and well below the 70% inhibition required for symptomatic exposure [13]. Although we found persistent changes in 5HT receptors and the 5HTT site, the pattern of effects was entirely different from that seen with chlorpyrifos. Whereas chlorpyrifos evoked parallel upregulation of both receptor subtypes, diazinon downregulated 5HT1A receptors. Chlorpyrifos showed strong targeting of brain regions containing 5HT cell bodies, probably reflecting reactive sprouting after damage to nerve terminal areas [5,72]; diazinon showed no such selectivity. Chlorpyrifos affected both males and females but with a notably greater effect in males; diazinon did indeed also show a preferential effect for 5HT1A receptors in males but the change was in the opposite direction from that obtained with chlorpyrifos; diazinon also showed a preferential upregulation of 5HTT in females, a specificity not seen with chlorpyrifos, which instead increased values in the brainstem for both sexes and reduced cerebrocortical values in females. But perhaps most notably, the effects of chlorpyrifos reported earlier were far more robust than those seen here for diazinon; indeed, compared to chlorpyrifos, the persistent effects of diazinon on 5HT systems were small, albeit statistically significant.
These results provide several important conclusions. First, there are major differences in the outcomes from the two organophosphates, even when they are administered in doses that occupy the same region of the dose-response curve in terms both of the amount given and of pharmacodynamic effect as assessed by cholinesterase inhibition [79,81]. It is thus inescapable that the effects of these low doses on 5HT systems are unrelated to the shared, anticholinesterase mechanism of chlorpyrifos and diazinon, and in turn, measurements of different exposure paradigms on that particular enzyme do not provide meaningful information about the threshold for developmental neurotoxicity in these circuits. Second, the disparate outcomes from chlorpyrifos and diazinon validate predictions made from the immediate impact of neonatal exposure on patterns of gene expression related to 5HT receptors and indices of neuronal damage [25,73], indicating that at least some of the differences reflect direct effects of the organophosphates on neuronal cell differentiation. Indeed, we recently identified a number of neurotrophic interactions and processes involved in cell damage/repair, indicative of distinctly different toxicant actions of chlorpyrifos and diazinon [25,73,77]. Third, based on the neurochemical findings, the functional outcomes can be expected to diverge in several key features centered around 5HT-dependent behaviors, such as emotional and appetitive functions. We recently tested this prediction with several measures of anxiety and hedonia [1,62]. Whereas chlorpyrifos had an anxiolytic effect on males in the plus maze, diazinon was anxiogenic, just as would be predicted from their opposite effects on 5HT1A receptors. Chlorpyrifos produced appetitive anhedonia in both males and females in the chocolate milk preference test, whereas diazinon exerted the effect in males only. The two agents also differed in their impact on the role of 5HT in cognitive function. With neonatal chlorpyrifos exposure, cognitive performance that ordinarily depends on acetylcholine circuits was instead taken over by 5HT, so that administration of a 5HT2 antagonist produced cognitive impairment, an effect not seen in controls [1]. In keeping with the lessened impact of diazinon on 5HT systems, we did not observe any such change after exposure to this organophosphate [84].
Although the specific outcomes of developmental exposures may differ among the various organophosphates, the disparities between effects on males and females seen here for diazinon continues a pattern of sex-selectivity reported for other agents in this class [1,15,18,33,34,48,60,67,68,75,78]. Although there may be effects on gonadal hormones [6,26,43,59,85], these effects generally require much higher exposures than those used in the present study and in our earlier work with chlorpyrifos. There are two other factors that are more likely to contribute to the sex differences. First, the exposure period studied here corresponds to the major phase of sexual differentiation of the brain [39,44], and for chlorpyrifos, we found that this is the peak period of sensitivity, both for effects on serotonergic systems and related behaviors [1,2,4,5]. In fact, the net effect of chlorpyrifos is to obtund the normally-occurring sex differences, likely reflecting interference with sexual differentiation of the brain. It is therefore reasonable to expect that diazinon, although differing in other ways, may similarly alter sex-related developmental events when exposure occurs during this critical period. The second likely factor is the inherent difference in brain plasticity between males and females [40,44,83]. The outcome from an early neurodevelopmental insult represents the net effect of the initial injury as well as the subsequent plastic responses that may produce adaptive or maladaptive changes due to rewiring of the affected circuits. In fact, we already have significant evidence demonstrating the subsequent maturational changes that occur after neonatal chlorpyrifos exposure [5,72]: alterations present in young adulthood in females tend to disappear by five months of age, whereas those in males do not. It is therefore likely that similar factors operate for the emergence of sex-selective neurodevelopmental differences in the effects of diazinon.
We found that the dose-effect relationship for the actions of diazinon on 5HT systems was nonmonotonic, displaying significant alterations at the low dose that were no longer apparent when the dose was raised to just above the threshold for cholinesterase inhibition. This same pattern was reported earlier for chlorpyrifos, both in terms of neurochemistry [56] and behavior [24,33]. For diazinon, the disparity in dose-effect curves can also be detected in the immediate impact of exposure on patterns of gene expression delineating cell damage/repair and alterations in differentiation related to specific neurotransmitter phenotypes [73]; as seen here, these extend to the ultimate effects on 5HT function in adolescence and adulthood, and again correspond to nonmonotonic behavioral outcomes [62,84]. As discussed previously, acetylcholine subserves important trophic functions in brain development centered around this specific period of neonatal exposure [22,23,31], so that a small degree of cholinergic activation just as the dose exceeds the threshold for cholinesterase inhibition may serve to offset some of the adverse, noncholinergic effects on brain development, provided that these are in just the right relative balance, akin to the positive effect of choline supplementation [41,42,45]. Alternatively, the increasing damage evoked by higher doses that recruit other mechanisms such as cholinergic hyperstimulation, may result in more widespread effects on other neurotransmitter systems that suppress some of the long-term changes in 5HT circuits evoked by the direct actions of the organophosphates. In either case, a reduced effect on specific biomarkers or small subset of behaviors should not be interpreted as a lack of a global, adverse impact on brain development.
The targeting of 5HT function by organophosphates is important for a number of reasons. Early in brain development, 5HT is a morphogen, so that disruption of this system leads to abnormal architectural assembly of the brain [21,87,88]. Next, early stages of synaptic communication imprint the future reactivity of 5HT circuits, particularly involving the expression of 5HT1A receptors [1,2,5,28], the subtype found to be most highly targeted by chlorpyrifos [4,5,72] or, as seen here, by diazinon. Third, alterations in 5HT function elicit changes in emotion, cognition, appetite and sleep patterns [1,49,50,61], thus expanding the scope of behavioral endpoints that need to be considered after early organophosphate exposure, issues that are just now being pursued in a number of laboratories [1,27,62]. Fourth, the current results demonstrate that a longstanding, basic assumption about the developmental neurotoxicity of organophosphates is incorrect: these agents do not produce a parallel set of neurobehavioral outcomes, and consequently, the various agents will need to be evaluated separately and in a comparative framework that incorporates endpoints other than cholinesterase inhibition. Finally, the heterogeneity of the developmental neurotoxicity of different organophosphates provides an opportunity to design new and safer pesticides within this class of compounds.
Acknowledgments
Acknowledgments/disclaimers: Research was supported by NIH ES10356. The authors state that they have no conflicts of interest. Theodore Slotkin and Frederic Seidler have provided expert witness testimony on behalf of government agencies, corporations and/or individuals.
Abbreviations
- 5HT
5-hydroxytrypamine, serotonin
- ANOVA
analysis of variance
- PN
postnatal day
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Aldridge JE, Levin ED, Seidler FJ, Slotkin TA. Developmental exposure of rats to chlorpyrifos leads to behavioral alterations in adulthood, involving serotonergic mechanisms and resembling animal models of depression. Environ Health Perspect. 2005;113:527–531. doi: 10.1289/ehp.7867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Aldridge JE, Meyer A, Seidler FJ, Slotkin TA. Alterations in central nervous system serotonergic and dopaminergic synaptic activity in adulthood after prenatal or neonatal chlorpyrifos exposure. Environ Health Perspect. 2005;113:1027–1031. doi: 10.1289/ehp.7968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Aldridge JE, Meyer A, Seidler FJ, Slotkin TA. Developmental exposure to terbutaline and chlorpyrifos: pharmacotherapy of preterm labor and an environmental neurotoxicant converge on serotonergic systems in neonatal rat brain regions. Toxicol Appl Pharmacol. 2005;203:134–144. doi: 10.1016/j.taap.2004.08.002. [DOI] [PubMed] [Google Scholar]
- 4.Aldridge JE, Seidler FJ, Meyer A, Thillai I, Slotkin TA. Serotonergic systems targeted by developmental exposure to chlorpyrifos: effects during different critical periods. Environ Health Perspect. 2003;111:1736–1743. doi: 10.1289/ehp.6489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Aldridge JE, Seidler FJ, Slotkin TA. Developmental exposure to chlorpyrifos elicits sex-selective alterations of serotonergic synaptic function in adulthood: critical periods and regional selectivity for effects on the serotonin transporter, receptor subtypes, and cell signaling. Environ Health Perspect. 2004;112:148–155. doi: 10.1289/ehp.6713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Andersen HR, Vinggaard AM, Rasmussen TH, Gjermandsen IM, Bonefeld-Jorgensen EC. Effects of currently used pesticides in assays for estrogenicity, androgenicity, and aromatase activity in vitro. Toxicol Appl Pharmacol. 2002;179:1–12. doi: 10.1006/taap.2001.9347. [DOI] [PubMed] [Google Scholar]
- 7.Arango V, Underwood MD, Boldrini M, Tamir H, Kassir SA, Hsiung S, Chen JJ, Mann JJ. Serotonin-1A receptors, serotonin transporter binding and serotonin transporter mRNA expression in the brainstem of depressed suicide victims. Neuropsychopharmacology. 2001;25:892–903. doi: 10.1016/S0893-133X(01)00310-4. [DOI] [PubMed] [Google Scholar]
- 8.Barnes NM, Sharp T. A review of central 5-HT receptors and their function. Neuropharmacology. 1999;38:1083–1152. doi: 10.1016/s0028-3908(99)00010-6. [DOI] [PubMed] [Google Scholar]
- 9.Barone S, Das KP, Lassiter TL, White LD. Vulnerable processes of nervous system development: a review of markers and methods. Neurotoxicology. 2000;21:15–36. [PubMed] [Google Scholar]
- 10.Betancourt AM, Burgess SC, Carr RL. Effect of developmental exposure to chlorpyrifos on the expression of neurotrophin growth factors and cell-specific markers in neonatal rat brain. Toxicol Sci. 2006;92:500–506. doi: 10.1093/toxsci/kfl004. [DOI] [PubMed] [Google Scholar]
- 11.Betancourt AM, Carr RL. The effect of chlorpyrifos and chlorpyrifos-oxon on brain cholinesterase, muscarinic receptor binding, and neurotrophin levels in rats following early postnatal exposure. Toxicol Sci. 2004;77:63–71. doi: 10.1093/toxsci/kfh003. [DOI] [PubMed] [Google Scholar]
- 12.Casida JE, Quistad GB. Organophosphate toxicology: safety aspects of nonacetylcholinesterase secondary targets. Chem Res Toxicol. 2004;17:983–998. doi: 10.1021/tx0499259. [DOI] [PubMed] [Google Scholar]
- 13.Clegg DJ, van Gemert M. Determination of the reference dose for chlorpyrifos: proceedings of an expert panel. J Toxicol Environ Health. 1999;2:211–255. doi: 10.1080/109374099281179. [DOI] [PubMed] [Google Scholar]
- 14.Costa LG. Current issues in organophosphate toxicology. Clin Chim Acta. 2006;366:1–13. doi: 10.1016/j.cca.2005.10.008. [DOI] [PubMed] [Google Scholar]
- 15.Dam K, Seidler FJ, Slotkin TA. Chlorpyrifos exposure during a critical neonatal period elicits gender-selective deficits in the development of coordination skills and locomotor activity. Dev Brain Res. 2000;121:179–187. doi: 10.1016/s0165-3806(00)00044-4. [DOI] [PubMed] [Google Scholar]
- 16.De Peyster A, Willis WO, Molgaard CA, MacKendrick TM, Walker C. Cholinesterase and self-reported pesticide exposure among pregnant women. Arch Environ Health. 1993;48:348–352. doi: 10.1080/00039896.1993.9936724. [DOI] [PubMed] [Google Scholar]
- 17.Fujita M, Charney DS, Innis RB. Imaging serotonergic neurotransmission in depression: hippocampal pathophysiology may mirror global brain alterations. Biol Psychiat. 2000;48:801–812. doi: 10.1016/s0006-3223(00)00960-4. [DOI] [PubMed] [Google Scholar]
- 18.Garcia SJ, Seidler FJ, Qiao D, Slotkin TA. Chlorpyrifos targets developing glia: effects on glial fibrillary acidic protein. Dev Brain Res. 2002;133:151–161. doi: 10.1016/s0165-3806(02)00283-3. [DOI] [PubMed] [Google Scholar]
- 19.Gupta RC. Brain regional heterogeneity and toxicological mechanisms of organophosphates and carbamates. Toxicol Mech Meth. 2004;14:103–143. doi: 10.1080/15376520490429175. [DOI] [PubMed] [Google Scholar]
- 20.Gurunathan S, Robson M, Freeman N, Buckley B, Roy A, Meyer R, Bukowski J, Lioy PJ. Accumulation of chlorpyrifos on residential surfaces and toys accessible to children. Environ Health Perspect. 1998;106:9–16. doi: 10.1289/ehp.981069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hamon M, Bourgoin S, Chanez C, De Vitry F. Do serotonin and other neurotransmitters exert a trophic influence on the immature brain? Dev Neurobiol. 1989;12:171–183. [Google Scholar]
- 22.Hohmann CF. A morphogenetic role for acetylcholine in mouse cerebral neocortex. Neurosci Biobehav Rev. 2003;27:351–63. doi: 10.1016/s0149-7634(03)00066-6. [DOI] [PubMed] [Google Scholar]
- 23.Hohmann CF, Berger-Sweeney J. Cholinergic regulation of cortical development and plasticity: new twists to an old story. Perspect Dev Neurobiol. 1998;5:401–425. [PubMed] [Google Scholar]
- 24.Icenogle LM, Christopher C, Blackwelder WP, Caldwell DP, Qiao D, Seidler FJ, Slotkin TA, Levin ED. Behavioral alterations in adolescent and adult rats caused by a brief subtoxic exposure to chlorpyrifos during neurulation. Neurotoxicol Teratol. 2004;26:95–101. doi: 10.1016/j.ntt.2003.09.001. [DOI] [PubMed] [Google Scholar]
- 25.Jameson RR, Seidler FJ, Slotkin TA. Nonenzymatic functions of acetylcholinesterase splice variants in the developmental neurotoxicity of organophosphates: chlorpyrifos, chlorpyrifos oxon and diazinon. Environ Health Perspect. 2007;115:65–70. doi: 10.1289/ehp.9487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kang HG, Jeong SH, Cho JH, Kim DG, Park JM, Cho MH. Chlorpyrifos-methyl shows anti-androgenic activity without estrogenic activity in rats. Toxicology. 2004;199:219–230. doi: 10.1016/j.tox.2004.02.025. [DOI] [PubMed] [Google Scholar]
- 27.Kofman O, Ben-Bashat G. Diisopropylfluorophosphate administration in the pre-weanling period induces long-term changes in anxiety behavior and passive avoidance in adult mice. Psychopharmacology. 2006;183:452–461. doi: 10.1007/s00213-005-0208-z. [DOI] [PubMed] [Google Scholar]
- 28.Kusserow H, Davies B, Hörtnagl H, Voigt I, Stroh T, Bert B, Deng DR, Fink H, Veh RW, Theuring F. Reduced anxiety-related behaviour in transgenic mice overexpressing serotonin-1A receptors. Mol Brain Res. 2004;129:104–116. doi: 10.1016/j.molbrainres.2004.06.028. [DOI] [PubMed] [Google Scholar]
- 29.Landrigan PJ. Pesticides and polychlorinated biphenyls (PCBs): an analysis of the evidence that they impair children's neurobehavioral development. Mol Genet Metab. 2001;73:11–17. doi: 10.1006/mgme.2001.3177. [DOI] [PubMed] [Google Scholar]
- 30.Landrigan PJ, Claudio L, Markowitz SB, Berkowitz GS, Brenner BL, Romero H, Wetmur JG, Matte TD, Gore AC, Godbold JH, Wolff MS. Pesticides and inner-city children: exposures, risks, and prevention. Environ Health Perspect. 1999;107(suppl 3):431–437. doi: 10.1289/ehp.99107s3431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lauder JM, Schambra UB. Morphogenetic roles of acetylcholine. Environ Health Perspect. 1999;107(Suppl 1):65–69. doi: 10.1289/ehp.99107s165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lee WJ, Alavanja MCR, Hoppin JA, Rusiecki JA, Kamel F, Blair A, Sandler DP. Mortality among pesticide applicators exposed to chlorpyrifos in the agricultural health study. Environ Health Perspect. 2007;115:528–534. doi: 10.1289/ehp.9662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Levin ED, Addy N, Baruah A, Elias A, Christopher NC, Seidler FJ, Slotkin TA. Prenatal chlorpyrifos exposure in rats causes persistent behavioral alterations. Neurotoxicol Teratol. 2002;24:733–741. doi: 10.1016/s0892-0362(02)00272-6. [DOI] [PubMed] [Google Scholar]
- 34.Levin ED, Addy N, Christopher NC, Seidler FJ, Slotkin TA. Persistent behavioral consequences of neonatal chlorpyrifos exposure in rats. Dev Brain Res. 2001;130:83–89. doi: 10.1016/s0165-3806(01)00215-2. [DOI] [PubMed] [Google Scholar]
- 35.Leysen JE, Niemegeers CJ, Van Nueten JM, Laduron PM. [3H]Ketanserin ( R41468), a selective 3H-ligand for serotonin2 receptor binding sites: binding properties, brain distribution, and functional role. Mol Pharmacol. 1982;21:301–314. [PubMed] [Google Scholar]
- 36.London L, Flisher AJ, Wesseling C, Mergler D, Kromhout H. Suicide and exposure to organophosphate insecticides: cause or effect? Am J Ind Med. 2005;47:308–321. doi: 10.1002/ajim.20147. [DOI] [PubMed] [Google Scholar]
- 37.Maes M, Meltzer H. The serotonin hypothesis of major depression. In: Bloom FE, Kupfer DJ, Bunney BS, Ciaranello RD, Davis KL, Koob GF, Meltzer HY, Schuster CR, Shader RI, Watson SJ, editors. Psychopharmacology: The Fourth Generation of Progress. Raven Press; New York: 1995. pp. 933–944. [Google Scholar]
- 38.May M. Disturbing behavior: neurotoxic effects in children. Environ Health Perspect. 2000;108:A262–A267. doi: 10.1289/ehp.108-a262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.McCarthy MM. Molecular aspects of sexual differentiation of the rodent brain. Psychoneuroendocrinology. 1994;19:415–427. doi: 10.1016/0306-4530(94)90029-9. [DOI] [PubMed] [Google Scholar]
- 40.McEwen BS. Sex, stress and the hippocampus: allostasis, allostatic load and the aging process. Neurobiol Aging. 2002;23:921–939. doi: 10.1016/s0197-4580(02)00027-1. [DOI] [PubMed] [Google Scholar]
- 41.Meck WH, Williams CL. Characterization of the facilitative effects of perinatal choline supplementation on timing and temporal memory. Neuroreport. 1997;8:2831–2835. doi: 10.1097/00001756-199709080-00005. [DOI] [PubMed] [Google Scholar]
- 42.Meck WH, Williams CL. Choline supplementation during prenatal development reduces proactive interference in spacial memory. Dev Brain Res. 1999;118:51–59. doi: 10.1016/s0165-3806(99)00105-4. [DOI] [PubMed] [Google Scholar]
- 43.Meeker JD, Ryan L, Barr DB, Hauser R. Exposure to nonpersistent insecticides and male reproductive hormones. Epidemiology. 2006;17:61–68. doi: 10.1097/01.ede.0000190602.14691.70. [DOI] [PubMed] [Google Scholar]
- 44.Mong JA, McCarthy MM. Steroid-induced developmental plasticity in hypothalamic astrocytes: implications for synaptic patterning. J Neurobiol. 1999;40:602–619. doi: 10.1002/(sici)1097-4695(19990915)40:4<602::aid-neu14>3.0.co;2-o. [DOI] [PubMed] [Google Scholar]
- 45.Montoya DAC, White AM, Williams CL, Blusztajn JK, Meck WH, Swartzwelder HS. Prenatal choline exposure alters hippocampal responsiveness to cholinergic stimulation in adulthood. Dev Brain Res. 2000;123:25–32. doi: 10.1016/s0165-3806(00)00075-4. [DOI] [PubMed] [Google Scholar]
- 46.Moret C, Briley M. Platelet 3H-paroxetine binding to the serotonin transporter is insensitive to changes in central serotonergic innervation in the rat. Psychiat Res. 1991;38:97–104. doi: 10.1016/0165-1781(91)90035-n. [DOI] [PubMed] [Google Scholar]
- 47.Morin D, Sapena R, Zini R, Tillement JP. Serotonin enhances the β-adrenergic response in rat brain cortical slices. Eur J Pharmacol. 1992;225:273–274. doi: 10.1016/0922-4106(92)90029-u. [DOI] [PubMed] [Google Scholar]
- 48.Moser VC, Padilla S. Age- and gender-related differences in the time course of behavioral and biochemical effects produced by oral chlorpyrifos in rats. Toxicol Appl Pharmacol. 1998;149:107–119. doi: 10.1006/taap.1997.8354. [DOI] [PubMed] [Google Scholar]
- 49.Nemeroff CB. The neurobiology of depression. Sci Am. 1998;278(6):42–49. doi: 10.1038/scientificamerican0698-42. [DOI] [PubMed] [Google Scholar]
- 50.Nutt DJ. The neuropharmacology of serotonin and noradrenaline in depression. Int Clin Psychopharmacol. 2002;17:S1–S12. doi: 10.1097/00004850-200206001-00002. [DOI] [PubMed] [Google Scholar]
- 51.Ostrea EM, Morales V, Ngoumgna E, Prescilla R, Tan E, Hernandez E, Ramirez GB, Cifra HL, Manlapaz ML. Prevalence of fetal exposure to environmental toxins as determined by meconium analysis. Neurotoxicology. 2002;23:329–339. doi: 10.1016/s0161-813x(02)00077-3. [DOI] [PubMed] [Google Scholar]
- 52.Park S, Harrold JA, Widdowson PS, Williams G. Increased binding at 5-HT1A, 5-HT1B, and 5-HT2A receptors and 5-HT transporters in diet-induced obese rats. Brain Res. 1999;847:90–97. doi: 10.1016/s0006-8993(99)02055-7. [DOI] [PubMed] [Google Scholar]
- 53.Perera FP, Rauh V, Whyatt RM, Tang D, Tsai WY, Bernert JT, Tu YH, Andrews H, Barr DB, Camann DE, Diaz D, Dietrich J, Reyes A, Kinney PL. A summary of recent findings on birth outcomes and developmental effects of prenatal ETS, PAH, and pesticide exposures. Neurotoxicology. 2005;26:573–587. doi: 10.1016/j.neuro.2004.07.007. [DOI] [PubMed] [Google Scholar]
- 54.Physicians for Social Responsibility, Pesticides and Children, Physicians for Social Responsibility, Washington DC, 1995.
- 55.Pope CN. Organophosphorus pesticides: do they all have the same mechanism of toxicity? J Toxicol Environ Health. 1999;2:161–181. doi: 10.1080/109374099281205. [DOI] [PubMed] [Google Scholar]
- 56.Qiao D, Seidler FJ, Padilla S, Slotkin TA. Developmental neurotoxicity of chlorpyrifos: What is the vulnerable period? Environ Health Perspect. 2002;110:1097–1103. doi: 10.1289/ehp.021101097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Qiao D, Seidler FJ, Tate CA, Cousins MM, Slotkin TA. Fetal chlorpyrifos exposure: adverse effects on brain cell development and cholinergic biomarkers emerge postnatally and continue into adolescence and adulthood. Environ Health Perspect. 2003;111:536–544. doi: 10.1289/ehp.5828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Raines KW, Seidler FJ, Slotkin TA. Alterations in serotonin transporter expression in brain regions of rats exposed neonatally to chlorpyrifos. Dev Brain Res. 2001;130:65–72. doi: 10.1016/s0165-3806(01)00211-5. [DOI] [PubMed] [Google Scholar]
- 59.Recio R, Ocampo-Gomez G, Moran-Martinez J, Borja-Aburto V, Lopez-Cervante M, Uribe M, Torres-Sanchez L, Cebrian ME. Pesticide exposure alters follicle-stimulating hormone levels in Mexican agricultural workers. Environ Health Perspect. 2005;113:1160–1163. doi: 10.1289/ehp.7374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ricceri L, Venerosi A, Capone F, Cometa MF, Lorenzini P, Fortuna S, Calamendrei G. Developmental neurotoxicity of organophosphorous pesticides: fetal and neonatal exposure to chlorpyrifos alters sex-specific behaviors at adulthood in mice. Toxicol Sci. 2006;93:105–113. doi: 10.1093/toxsci/kfl032. [DOI] [PubMed] [Google Scholar]
- 61.Risch CS, Nemeroff CB. Neurochemical alterations of serotonergic neuronal systems in depression. J Clin Psychiat. 1991;53(suppl 10):3–6. [PubMed] [Google Scholar]
- 62.Roegge CS, Timofeeva OA, Seidler FJ, Slotkin TA, Levin ED. Developmental diazinon neurotoxicity in rats: later effects on emotional response. Brain Res Bull. 2007 doi: 10.1016/j.brainresbull.2007.08.008. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Rovescalli AC, Brunello N, Perez J, Vitali S, Steardo L, Racagni G. Heterologous sensitization of adenylate cyclase activity by serotonin in the rat cerebral cortex. Eur Neuropsychopharmacol. 1993;3:463–475. doi: 10.1016/0924-977x(93)90271-m. [DOI] [PubMed] [Google Scholar]
- 64.Sachana M, Flaskos J, Nikolaidis E, Hargreaves A, Alexaki-Tzivanidou E. Inhibition of rat platelet 5-hydroxytryptamine uptake by chlorpyrifos and carbaryl. Pharmacol Toxicol. 2001;89:195–200. doi: 10.1111/j.0901-9928.2001.890409.x. [DOI] [PubMed] [Google Scholar]
- 65.Slotkin TA. Developmental cholinotoxicants: nicotine and chlorpyrifos. Environ Health Perspect. 1999;107(suppl 1):71–80. doi: 10.1289/ehp.99107s171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Slotkin TA. Cholinergic systems in brain development and disruption by neurotoxicants: nicotine, environmental tobacco smoke, organophosphates. Toxicol Appl Pharmacol. 2004;198:132–151. doi: 10.1016/j.taap.2003.06.001. [DOI] [PubMed] [Google Scholar]
- 67.Slotkin TA. Developmental neurotoxicity of organophosphates: a case study of chlorpyrifos. In: Gupta RC, editor. Toxicity of Organophosphate and Carbamate Pesticides. Elsevier Academic Press; San Diego: 2005. pp. 293–314. [Google Scholar]
- 68.Slotkin TA, Cousins MM, Tate CA, Seidler FJ. Persistent cholinergic presynaptic deficits after neonatal chlorpyrifos exposure. Brain Res. 2001;902:229–243. doi: 10.1016/s0006-8993(01)02387-3. [DOI] [PubMed] [Google Scholar]
- 69.Slotkin TA, Levin ED, Seidler FJ. Comparative developmental neurotoxicity of organophosphate insecticides: effects on brain development are separable from systemic toxicity. Environ Health Perspect. 2006;114:746–751. doi: 10.1289/ehp.8828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Slotkin TA, McCook EC, Ritchie JC, Carroll BJ, Seidler FJ. Serotonin transporter expression in rat brain regions and blood platelets: aging and glucocorticoid effects. Biol Psychiat. 1997;41:172–183. doi: 10.1016/S0006-3223(96)00215-6. [DOI] [PubMed] [Google Scholar]
- 71.Slotkin TA, Miller DB, Fumagalli F, McCook EC, Zhang J, Bissette G, Seidler FJ. Modeling geriatric depression in animals: biochemical and behavioral effects of olfactory bulbectomy in young versus aged rats. J Pharmacol Exp Ther. 1999;289:334–345. [PubMed] [Google Scholar]
- 72.Slotkin TA, Seidler FJ. The alterations in CNS serotonergic mechanisms caused by neonatal chlorpyrifos exposure are permanent. Dev Brain Res. 2005;158:115–119. doi: 10.1016/j.devbrainres.2005.06.008. [DOI] [PubMed] [Google Scholar]
- 73.Slotkin TA, Seidler FJ. Comparative developmental neurotoxicity of organophosphates in vivo: transcriptional responses of pathways for brain cell development, cell signaling, cytotoxicity and neurotransmitter systems. Brain Res Bull. 2007;72:232–274. doi: 10.1016/j.brainresbull.2007.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Slotkin TA, Seidler FJ. Developmental exposure to terbutaline and chlorpyrifos, separately or sequentially, elicits presynaptic serotonergic hyperactivity in juvenile and adolescent rats. Brain Res Bull. 2007;73:301–309. doi: 10.1016/j.brainresbull.2007.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Slotkin TA, Seidler FJ. Prenatal chlorpyrifos exposure elicits presynaptic serotonergic and dopaminergic hyperactivity at adolescence: critical periods for regional and sex-selective effects. Reprod Toxicol. 2007;23:421–427. doi: 10.1016/j.reprotox.2006.07.010. [DOI] [PubMed] [Google Scholar]
- 76.Slotkin TA, Seidler FJ, Ali SF. Cellular determinants of reduced adaptability of the aging brain: neurotransmitter utilization and cell signaling responses after MDMA lesions. Brain Res. 2000;879:163–173. doi: 10.1016/s0006-8993(00)02767-0. [DOI] [PubMed] [Google Scholar]
- 77.Slotkin TA, Seidler FJ, Fumagalli F. Exposure to organophosphates reduces the expression of neurotrophic factors in neonatal rat brain regions: similarities and differences in the effects of chlorpyrifos and diazinon on the fibroblast growth factor superfamily. Environ Health Perspect. 2007;115:909–916. doi: 10.1289/ehp.9901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Slotkin TA, Tate CA, Cousins MM, Seidler FJ. Functional alterations in CNS catecholamine systems in adolescence and adulthood after neonatal chlorpyrifos exposure. Dev Brain Res. 2002;133:163–173. doi: 10.1016/s0165-3806(02)00284-5. [DOI] [PubMed] [Google Scholar]
- 79.Slotkin TA, Tate CA, Ryde IT, Levin ED, Seidler FJ. Organophosphate insecticides target the serotonergic system in developing rat brain regions: disparate effects of diazinon and parathion at doses spanning the threshold for cholinesterase inhibition. Environ Health Perspect. 2006;114:1542–1546. doi: 10.1289/ehp.9337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, Fujimoto EK, Goeke NM, Olson BJ, Klenk DC. Measurement of protein using bicinchoninic acid. Anal Biochem. 1985;150:76–85. doi: 10.1016/0003-2697(85)90442-7. [DOI] [PubMed] [Google Scholar]
- 81.Song X, Seidler FJ, Saleh JL, Zhang J, Padilla S, Slotkin TA. Cellular mechanisms for developmental toxicity of chlorpyrifos: targeting the adenylyl cyclase signaling cascade. Toxicol Appl Pharmacol. 1997;145:158–174. doi: 10.1006/taap.1997.8171. [DOI] [PubMed] [Google Scholar]
- 82.Stockmeier CA, Shapiro LA, Dilley GE, Kolli TN, Friedman L, Rajkowska G. Increase in serotonin-1A autoreceptors in the midbrain of suicide victims with major depression: postmortem evidence for decreased serotonin activity. J Neurosci. 1998;18:7394–7401. doi: 10.1523/JNEUROSCI.18-18-07394.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Suzuki S, Brown CM, Wise PM. Mechanisms of neuroprotection by estrogen. Endocrine. 2006;29:209–215. doi: 10.1385/ENDO:29:2:209. [DOI] [PubMed] [Google Scholar]
- 84.Timofeeva OA, Roegge CS, Seidler FJ, Slotkin TA, Levin ED. Persistent cognitive alterations in rats after early postnatal exposure to low doses of the organophosphate pesticide, diazinon. Neurotoxicol Teratol. 2007 doi: 10.1016/j.ntt.2007.10.002. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Vinggaard AM, Hnida C, Breinholt V, Larsen JC. Screening of selected pesticides for inhibition of CYP19 aromatase activity in vitro. Toxicol in Vitro. 2000;14:227–234. doi: 10.1016/s0887-2333(00)00018-7. [DOI] [PubMed] [Google Scholar]
- 86.Weiss B, Amler S, Amler RW. Pesticides. Pediatrics. 2004;113:1030–1036. [PubMed] [Google Scholar]
- 87.Whitaker-Azmitia PM. Role of serotonin and other neurotransmitter receptors in brain development: basis for developmental pharmacology. Pharmacol Rev. 1991;43:553–561. [PubMed] [Google Scholar]
- 88.Whitaker-Azmitia PM. Serotonin and brain development: role in human developmental diseases. Brain Res Bull. 2001;56:479–485. doi: 10.1016/s0361-9230(01)00615-3. [DOI] [PubMed] [Google Scholar]
- 89.Whitney KD, Seidler FJ, Slotkin TA. Developmental neurotoxicity of chlorpyrifos: cellular mechanisms. Toxicol Appl Pharmacol. 1995;134:53–62. doi: 10.1006/taap.1995.1168. [DOI] [PubMed] [Google Scholar]
- 90.Xu Z, Seidler FJ, Ali SF, Slikker W, Slotkin TA. Fetal and adolescent nicotine administration: effects on CNS serotonergic systems. Brain Res. 2001;914:166–178. doi: 10.1016/s0006-8993(01)02797-4. [DOI] [PubMed] [Google Scholar]
- 91.Xu Z, Seidler FJ, Cousins MM, Slikker W, Slotkin TA. Adolescent nicotine administration alters serotonin receptors and cell signaling mediated through adenylyl cyclase. Brain Res. 2002;951:280–292. doi: 10.1016/s0006-8993(02)03174-8. [DOI] [PubMed] [Google Scholar]
- 92.Yanai J, Vatury O, Slotkin TA. Cell signaling as a target and underlying mechanism for neurobehavioral teratogenesis. Ann NY Acad Sci. 2002;965:473–478. doi: 10.1111/j.1749-6632.2002.tb04188.x. [DOI] [PubMed] [Google Scholar]
- 93.Yatham LN, Liddle PF, Dennie J, Shiah IS, Adam MJ, Lane CJ, Lam RW, Ruth TJ. Decrease in brain serotonin-2 receptor binding in patients with major depression following desipramine treatment: a positron emission tomography study with fluorine-18-labeled setoperone. Arch Gen Psychiat. 1999;56:705–711. doi: 10.1001/archpsyc.56.8.705. [DOI] [PubMed] [Google Scholar]
- 94.Yatham LN, Liddle PF, Shiah IS, Scarrow G, Lam RW, Adam MJ, Zis AP, Ruth TJ. Brain serotonin-2 receptors in major depression: a positron emission tomography study. Arch Gen Psychiat. 2000;57:850–858. doi: 10.1001/archpsyc.57.9.850. [DOI] [PubMed] [Google Scholar]