

Abstract
Ca2+ liberation through inositol 1,4,5-trisphosphate receptors (IP3Rs) plays a universal role in cell regulation, and specificity of cell signalling is achieved through the spatiotemporal patterning of Ca2+ signals. IP3Rs display Ca2+-induced Ca2+ release (CICR), but are grouped in clusters so that regenerative Ca2+ signals may remain localized to individual clusters, or propagate globally between clusters by successive cycles of Ca2+ diffusion and CICR. We used confocal microscopy and photoreleased IP3 in Xenopus oocytes to study how these properties are modulated by mobile cytosolic Ca2+ buffers. EGTA (a buffer with slow ‘on-rate’) speeded Ca2+ signals and ‘balkanized’ Ca2+ waves by dissociating them into local signals. In contrast, BAPTA (a fast buffer with similar affinity) slowed Ca2+ responses and promoted ‘globalization’ of spatially uniform Ca2+ signals. These actions are likely to arise through differential effects on Ca2+ feedback within and between IP3R clusters, because Ca2+ signals evoked by influx through voltage-gated channels were little affected. We propose that cell-specific expression of Ca2+-binding proteins with distinct kinetics may shape the time course and spatial distribution of IP3-evoked Ca2+ signals for specific physiological roles.
Cytosolic Ca2+ ions regulate numerous aspects of cellular activity in virtually all cell types (Berridge et al. 2000). This versatility is made possible by the diverse mechanisms by which Ca2+ signals are generated and transmitted to act over very different time and distance scales (Berridge et al. 2000; Marchant & Parker, 2000). Endogenous Ca2+ binding proteins play a key role in determining the magnitude, kinetics and spatial distribution of cytosolic Ca2+ signals, and typically buffer most (> 90 %) of the total Ca2+ ions in the cytosol (Berridge et al. 2000). As well as simply reducing the availability of free cytosolic Ca2+ ions, buffers also affect their diffusional mobility. Immobile buffers reduce the effective diffusion coefficient for Ca2+, whereas mobile buffers can act as a ‘shuttle’ to speed Ca2+ diffusion in the presence of immobile buffers (Stern, 1992; Roberts, 1994). It is thus likely that cells utilize buffers to shape Ca2+ signals for their specific functions; a notion supported by observations that different cell types - particularly sub-populations of neurons - selectively express mobile Ca2+ binding proteins (e.g. parvalbumin and calretinin) with differing properties (Andressen et al. 1993). However, the functional consequences of selective buffer expression remain unclear (Neher, 2000; Schwaller et al. 2002).
Most experimental and theoretical investigations of cellular Ca2+ buffers have concentrated on signals arising from a fixed ‘pulse’ of Ca2+, e.g. Ca2+ entry through voltage- or ligand-gated membrane channels (e.g. Lee et al. 2000a,b). A more complex case involves Ca2+ liberation from intracellular stores through IP3Rs. The opening of these channels shows dual positive and negative regulation by cytosolic Ca2+ (Iino, 1990; Finch et al. 1991; Bezprozvanny et al. 1991; Mak et al. 1998), leading to a mechanism of CICR whereby small elevations of cytosolic [Ca2+] promote channel opening to liberate further Ca2+. Buffers are thus expected to exert a powerful effect on signals arising through intracellular Ca2+ liberation, as they will affect not only the fate of Ca2+ ions already deposited into the cytosol, but also influence the subsequent liberation of Ca2+ by modulating feedback onto IP3Rs.
IP3-evoked Ca2+ signals display complex spatial and temporal patterning. Stimulation by relatively high [IP3] typically gives rise to repetitive, propagating Ca2+ waves (Lechleiter & Clapham, 1992). Lower [IP3] evokes transient, localized Ca2+ elevations, christened ‘Ca2+ puffs’, which have been visualized in oocytes and other cell types (Yao et al. 1995; Bootman et al. 1997). Puffs are believed to arise because localized CICR causes the concerted opening of channels within localized clusters of a few tens of IP3Rs, which act as functionally discrete Ca2+ release units (Callamaras et al. 1998a,b; Swillens et al. 1999). Puffs may serve both as local Ca2+ signals in their own right, and as ‘building blocks’ from which Ca2+ waves are generated. When [IP3] is sufficiently high, Ca2+ diffusing from one release site triggers CICR at neighbouring sites a few microns away, giving rise to saltatory waves propagating from site to site (Yao et al. 1995; Bootman et al. 1997; Berridge, 1997; Callamaras et al. 1998a,b). Thus, Ca2+ diffusion and feedback onto the IP3R act over two very different distance scales: rapid (microsecond) interactions over nanometre scales between individual IP3Rs within a cluster to generate local Ca2+ puffs, and slower (tens of milliseconds) interactions over micrometre scales between release sites to generate global Ca2+ waves.
In this paper, we studied how Ca2+ buffers with differing kinetics modulate Ca2+ interactions over these widely divergent distance and time scales. Our objectives were to: (1) explore how cells might employ buffers to shape Ca2+ signals for their specific purposes, and (2) use buffers as a tool to derive mechanistic information about Ca2+ interactions at scales otherwise inaccessible to direct visualization.
We utilized Xenopus oocytes as a model cell system to image the perturbations of Ca2+ signalling resulting from intracellular injections of two mobile buffers, EGTA and BAPTA. The oocyte has advantages that intracellular Ca2+ liberation is mediated solely through type 1 IP3Rs (Parys et al. 1992); its large size greatly facilitates intracellular injections; and it is among the best characterized cells for Ca2+ signalling. EGTA and BAPTA were selected for study because these well-characterized exogenous buffers display similar affinities yet have very different Ca2+ binding kinetics (Tsien, 1980). EGTA has an on-rate (≈3-10 μm−1 s−1: Table 1) sufficiently fast to bind Ca2+ ions diffusing over the micrometre distances between Ca2+ release sites (Roberts et al. 1994; Horne & Meyer, 1997; Song et al. 1998; Kidd et al. 1999), but too slow to influence short range feedback of Ca2+ between individual IP3Rs at individual puff sites. On the other hand, BAPTA has a much faster on-rate (100–1000 μm−1 s−1: Table 1) and, whereas it is expected to disrupt local Ca2+ feedback within IP3R clusters, it may actually enhance coupling between release sites by acting as a ‘shuttle buffer’ (Stern, 1992; Roberts, 1994). We show here that injection of the slow buffer EGTA into the oocyte promotes a ‘balkanization’ of Ca2+ liberation as transient, autonomous local release events, whereas the fast buffer BAPTA results in a ‘globalization’ of spatially diffuse Ca2+ signals. Moreover, these spatial changes are accompanied by profound changes in kinetics of Ca2+ signals, suggesting that Ca2+ interactions between release sites play a large part in setting the duration of Ca2+ spikes.
Table 1.
Summary of kinetic parameters and diffusion distances for binding and unbinding of Ca2+ to EGTA and BAPTA
| EGTA | BAPTA | |
|---|---|---|
| Apparent Kd (nM) (at pH 7.2) | 150 | 160 |
| Kon (μM−1 s−1) | 3–10 | 100–1000 |
| Koff (s−1) | 0.5–1.5 | 16–160 |
| τdwell (ms) | 700–2000 | 6–60 |
| dshuttle (μm) | 28–50 | 3–9 |
| τcapture (mS) (for B = 270μm) | 4–10 | 0.04–0.4 |
| dcapture (μm) (for B = 270μm) | 0.7–1 | 0.07–0.2 |
Apparent affinities (Morris et al. 1999) and lower and upper estimates for on-rates (Kon) (Naraghi, 1997; Nagerl et al. 2000), were used to derive off-rates (Koff = KdKon) for EGTA and BAPTA. Dwell time (τdwell) reflects how long Ca2+ will remain bound to each buffer, where τdwell = 1/Koff. The corresponding mean distances over which the Ca2+-buffer complex will diffuse before releasing bound Ca2+ (dshuttle) were estimated as √ (6DCabufferτdwell), assuming a diffusion coefficient (DCabuffer) of 200 μm2 s−1. Mean capture times before a Ca2+ ion binds to EGTA or BAPTA were calculated as τcapture = 1/ (KonB) (Stern, 1992; Roberts, 1994), where B is the concentration of Ca2+-free binding sites on the buffer, assuming that Ca2+ ions in the cytosol are bound to immobile endogenous buffers for 90% of the time. The corresponding mean distances that a Ca2+ ion would diffuse before capture (dcapture) were calculated as dcapture = √ (DCaτcapture), assuming an apparent diffusion coefficient (DCa) of 20 μm2 s−1 for Ca2+ in the presence of immobile endogenous buffers.
METHODS
Preparation of Xenopus oocytes
Xenopus laevis were anaesthetized by immersion in 0.17 % MS-222 for 15 min and killed by decapitation in adherence with protocols approved by the UC Irvine Institutional Animal Care and Use Committee. Oocytes (stage V-VI) were manually plucked and collagenase-treated (0.5 mg ml−1 for 30 min) before storage in Barth's solution (composition (mm): NaCl, 88; KCl, 1; NaHCO3, 2.4; MgSO4, 0.82; Ca(NO3)2, 0.33; CaCl2, 0.41; Hepes, 5; pH 7.4) containing 0.1 mg ml−1 gentamicin at 17 °C for 1–7 days before use.
Microinjection of oocytes
Intracellular microinjections were performed using a Drummond microinjector. Oocytes were first loaded with Oregon Green 488 BAPTA 1 (OG-1) together with caged IP3 (d-myo-inositol 1,4,5-trisphosphate, P4(5)-(1-(2-nitrophenyl)ethyl) ester) to respective final intracellular concentrations of 48 and 8 μm; assuming 1 μl cytosolic volume. After allowing 30 min for intracellular distribution, control Ca2+ responses were imaged. A specified volume of buffer solution was then injected through a fresh micropipette and the pipette was immediately removed to minimize leakage. Ca2+ responses were then imaged after allowing ≈20 min for intracellular equilibration of buffer concentration, and sequential injections and recordings were made in this way to examine the effects of stepwise increases in buffer concentration. Injection solutions contained 5 mm of either EGTA or BAPTA together with 2.5 mm CaCl2 (pH 7.25, with KOH), such that the buffers were half-saturated with Ca2+. To obtain high final intracellular concentrations (> 270 μm), stock buffer concentrations were increased to 10 mm (including 5 mm CaCl2) and buffer was administered by a series of repeated 10 nl injections. Control experiments, where high cytosolic buffer concentrations were attained following single injections of EGTA or BAPTA, gave similar results to those where the concentration was raised in a stepwise manner. Moreover, responses evoked in control oocytes (not injected with buffer) remained consistent over periods up to 3 h.
Confocal laser scanning microscopy
Confocal Ca2+ images were obtained using a custom-built line-scan confocal scanner interfaced to an Olympus IX70 inverted microscope (Parker et al. 1997). Recordings were made at room temperature, imaging in the animal hemisphere of oocytes bathed in normal Ringer solution (composition (mm): NaCl2, 120; KCl, 2; CaCl2, 1.8; Hepes, 5; pH 7.3). The laser spot of a 488 nm argon ion laser was focused with a ×40 oil immersion objective (NA 1.35) and scanned every 16 ms along a 50 μm line, of which representative trimmed sections are shown in the figures. Emitted fluorescence was detected at wavelengths > 510 nm through a confocal pinhole providing lateral and axial resolutions of about 0.3 and 0.5 μm, respectively. The scan line was focused at the level of the pigment granules and images were collected through a coverglass forming the base of the recording chamber. Fluorescence signals are expressed as ratios (F/F0 or ΔF/F0) of the fluorescence (F) at each pixel relative to the mean resting fluorescence (F0) at that pixel prior to stimulation. IP3 was photo-released from a caged precursor by delivering flashes of UV light, focused uniformly throughout a 200 μm spot surrounding the image scan line (Callamaras & Parker, 1998). The amount of photo-released IP3 was controlled (in a linear manner) by varying flash duration. Each flash consumes only a negligible fraction of the caged IP3 (Callamaras & Parker, 1998), thus it was possible to acquire numerous consistent responses using repeated flashes. Intervals of > 60 s were allowed between recordings to allow IP3Rs to recover from desensitization and for cytosolic [Ca2+] to recover to basal levels. Images were constructed and processed using custom routines written in the IDL programming environment (Research Systems, Boulder, CO, USA), and measurements were exported to Microcal Origin 6.0 (OriginLab, Northampton, MA, USA) for analysis and graphing.
Expression of N-type Ca2+ channels
Plasmids containing cDNA clones coding for the N-type Ca2+ channel α1B-d and β3 subunits (Lin et al. 1997) were linearized and transcribed in vitro. Equal quantities of each subunit cRNA were mixed to a final concentration of 0.1-1 μg μl−1 and injected (50 nl) into defolliculated oocytes 3–5 days prior to imaging and electrophysiological recording. Membrane potential was held at −60 mV using a two-electrode voltage clamp (Gene Clamp 500, Axon Instruments, Foster City, CA, USA), and stepped to +30 mV to induce opening of N-type Ca2+ channels expressed in the plasma membrane. Ca2+ imaging was as described above, except that [Ca2+] in the Ringer solution was 6 mm.
Reagents
OG-1, caged IP3 and BAPTA were from Molecular Probes Inc. (Eugene, OR, USA); all other reagents were from Sigma Chemical Co. (St Louis, MO, USA).
RESULTS
Actions of buffers on IP3-evoked Ca2+ signals
We compared the actions of fast (BAPTA) and slow (EGTA) Ca2+ buffers on IP3-evoked intracellular Ca2+ signals. Xenopus oocytes were loaded with the indicator OG-1 together with caged IP3, and confocal line-scan images of Ca2+-dependent fluorescence were acquired along a 50 μm scan line focused ≈5 μm inward from the cell surface where Ca2+ release sites are concentrated (Callamaras & Parker, 1999). IP3 was photo-released by UV light flashes, with durations set by an electronic shutter to provide a linear control of the relative amount of IP3 liberation. To control for variation between oocytes, flash strengths were normalized relative to that evoking a half-maximal response under control conditions (i.e. before adding buffer) in each cell.
Figure 1 illustrates Ca2+ images obtained at various buffer concentrations in response to a fixed flash strength, and Fig. 2 shows superimposed fluorescence traces in response to various flash strengths at each buffer concentration. Following acquisition of control records (upper panels in Fig. 1 and Fig. 2), a volume of buffer solution (EGTA, Fig. 1A and Fig. 2A; or BAPTA, Fig. 1B and Fig. 2B) was injected into each oocyte to give a final intracellular concentration of 23 μm (see Methods for details). Microinjection pipettes were removed immediately following injection to prevent leakage, and oocytes were left for ≈20 min to ensure uniform buffer distribution and re-equilibration of Ca2+ ions in the cytosol before imaging responses to the same series of UV flashes. This procedure was then repeated, sequentially injecting increasing volumes of buffer solutions to image Ca2+ signals at progressively higher buffer concentrations (46–540 μm).
Figure 1. Modulation of IP3-evoked Ca2+ signals by EGTA and BAPTA.
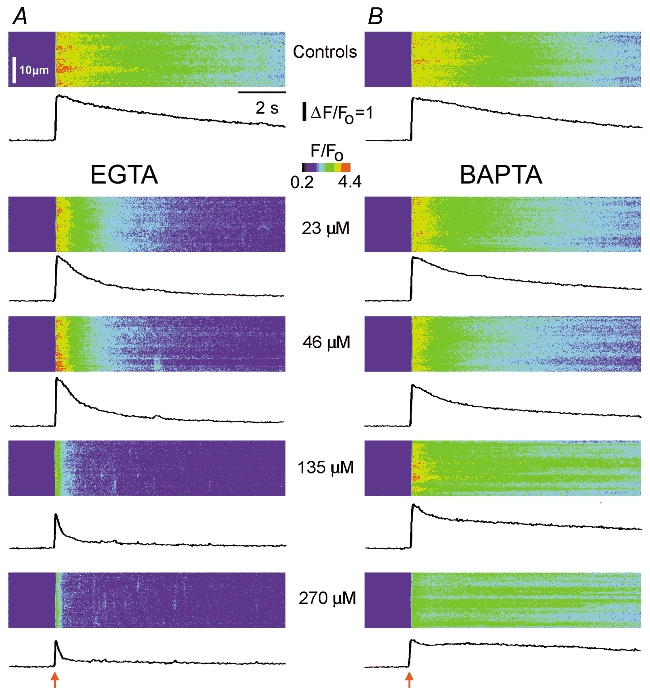
Confocal line-scan images illustrate Ca2+ signals evoked by photoreleased IP3 in the presence of increasing concentrations of buffer. In each image, distance is depicted vertically, time runs from left to right and increasing Oregon Green 488 BAPTA 1 (OG-1) fluorescence ratio (ΔF/F0: proportional to [Ca2+]free) is represented by ‘warmer’ colours as denoted by the colour bar. Identical photolysis flashes (normalized intensities of 1.5 in A and 1.9 in B) were delivered at the arrows. Traces below each image show fluorescence profiles averaged over 100 pixel (6.6 μm) regions. A, upper panel shows response before loading buffer, and subsequent panels illustrate responses after sequentially loading the same oocyte with EGTA to the final intracellular concentrations stated. B, similar records from a different oocyte showing the effects of increasing concentrations of BAPTA.
Figure 2. Buffer actions at varying [IP3].

Representative fluorescence profiles (from different oocytes to those in Fig. 1) show superimposed Ca2+ transients evoked by increasing photorelease of IP3 in the absence of exogenous buffer (upper panels) and after loading increasing concentrations of EGTA (A) or BAPTA (B). Traces correspond to different photolysis flash strengths, indicated in normalized units.
The data in Figs. 1 and 2 are representative of observations in 45 oocytes (EGTA, n = 21; BAPTA, n = 24). The results are analysed in the following sections, but the most striking finding was that EGTA (a Ca2+ buffer with slow Ca2+ binding kinetics) greatly abbreviated IP3-evoked Ca2+ signals, whereas BAPTA (a faster Ca2+ buffer) slowed their decay.
Changes in Ca2+ signals result from increased buffering, not changes in resting [Ca2+] or store content
The effects described in Fig. 1 and Fig. 2 might, in principle, arise either through buffering of Ca2+ liberated in response to IP3, or because the added buffers chelated oocyte Ca2+ to cause long-term changes in basal cytosolic free [Ca2+] or in the loading of Ca2+ stores. Observations that equal amounts of two buffers of similar affinity produced dissimilar effects already suggested that the latter possibilities were not the case. Moreover, experiments were carried out by injecting each buffer already half-saturated with Ca2+, so that the free [Ca2+] in the injection solutions would have been ≈150 nm, a little higher than basal free [Ca2+] in the oocyte cytosol (Yao et al. 1995). As an additional control, we repeated the experiments using solutions of EGTA and BAPTA that contained either no added Ca2+ or were saturated (1:1 ratio) with Ca2+. The results were essentially identical to those in Fig. 1 and Fig. 2 (data not shown), indicating that the effects of EGTA and BAPTA on Ca2+ signals did not result from addition or removal of available Ca2+ ions within the oocyte.
Positive and negative modulation of IP3-evoked Ca2+ signals by buffers
Figure 3A and B shows families of curves plotting the mean peak amplitude of fluorescence signals evoked by increasing photorelease of IP3 at various concentrations of EGTA and BAPTA, respectively. High cytosolic concentrations (> 135 μm) of both buffers appreciably reduced the amplitude of Ca2+ signals evoked by strong photorelease of IP3, with BAPTA being more potent than EGTA. This reduction was expected - in part because the buffers will compete with indicator dye for free Ca2+ ions. In contrast, Ca2+ signals evoked by weak photolysis flashes were potentiated by EGTA; as shown in Fig. 3C, plotting Ca2+ signals evoked by weak (0.6-0.8) and strong (1.9-2.6) flashes at various buffer concentrations relative to control responses without added buffer. Potentiation was strongest at low (< 300 μm) concentrations of EGTA, but responses remained greater than control values at concentrations as high as 2 mm EGTA. The slow binding kinetics of EGTA appear to be important for this effect, because little potentiation was apparent with corresponding concentrations of BAPTA (Fig. 3D).
Figure 3. Buffers both facilitate and depress IP3-evoked Ca2+ signals.
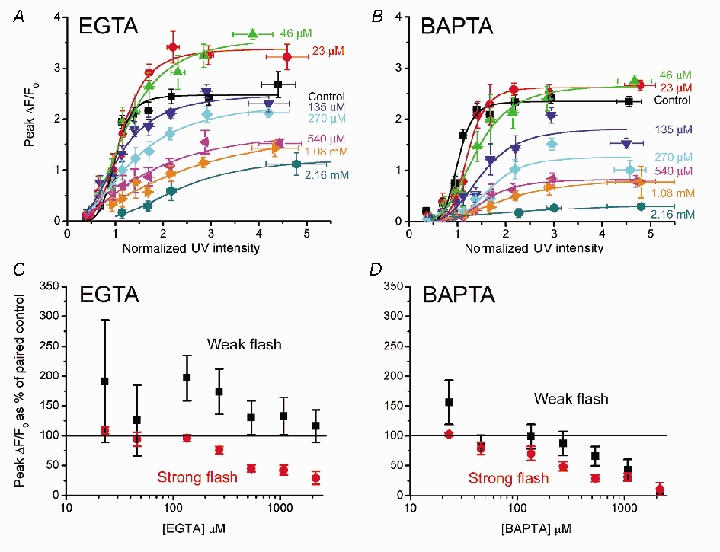
A and B, mean peak amplitude (ΔF/F0) of Ca2+ signals as a function of photolysis flash strength, plotted for various intracellular concentrations of EGTA (A: n = 21 oocytes) and BAPTA (B: n = 24). Curves were fitted using the Hill equation. C and D, changes in peak fluorescence signal as a function of increasing [buffer] plotted as a percentage of that in the same oocyte before loading buffer. Data are pooled for weak (black symbols) and strong (red) photolysis flashes (respective normalized flash strengths 0.6–0.8 and 1.9–2.6) in the presence of EGTA (C, n = 14) and BAPTA (D, n = 17).
Buffers reduce the apparent cooperativity of IP3-evoked Ca2+ liberation
The concentration-response relationships in Figs 3A and B are fitted by Hill curves (y = Vmax(xnH/(knH+xnH): where y = peak fluorescence ratio (ΔF/F0) at any given [IP3] (x); Vmax = peak ΔF/F0 at saturating [IP3]; and nH is the Hill coefficient, which gives a measure of the apparent cooperativity of IP3 action in evoking Ca2+ liberation. Data from these curves are plotted in Fig. 4, showing the dependence of nH (Fig. 4A) and Vmax (Fig. 4B) on concentration of EGTA or BAPTA.
Figure 4. Ca2+ buffers reduce the apparent cooperativity of IP3 action.
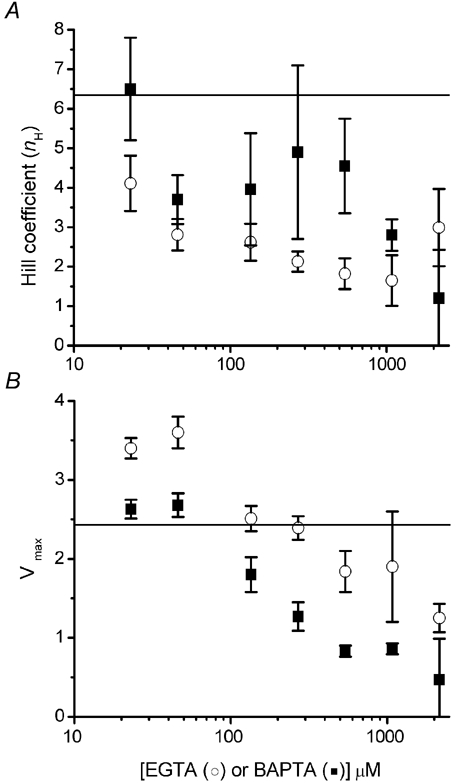
A, Hill coefficients, derived from the curves fitted to data in Fig. 3A and B, are plotted as functions of [EGTA] (open circles) and [BAPTA] (filled squares). B, plot shows Vmax (maximal fluorescence signal at infinite [IP3] derived from Hill-fits) as functions of [EGTA] (open circles) and [BAPTA] (filled squares). Horizontal lines on each graph represent control values (i.e. no exogenous buffer).
The steep dependence of Ca2+ liberation on [IP3] is likely to arise both from the requirement for binding to more than one subunit of the tetrameric receptor for channel opening and because of positive cooperativity resulting from CICR (Meyer et al. 1990; Hirota et al. 1995; Marchant & Taylor, 1997; Callamaras et al. 1998a). In agreement with this dual mechanism, nH in control conditions (before adding buffer) was ≈6.3, a value greater than expected even if channel opening requires binding to all four subunits. Moreover, progressive increases in buffer concentrations reduced nH to a minimal value of about 2 (Fig. 4A). A probable interpretation is that Ca2+ buffers disrupt the diffusion of Ca2+ ions between IP3Rs that normally leads to CICR, and that this communication contributes appreciably to the apparent cooperativity of IP3 action. Indeed, the minimal nH at high buffer concentrations suggests that binding of IP3 to two (or possibly even one) subunits may be sufficient for channel opening. Finally, moderate concentrations of EGTA caused a greater reduction in nH than equivalent concentrations of BAPTA (Fig. 4A). Because EGTA impairs Ca2+ diffusion between clusters of IP3Rs rather than between individual IP3Rs within a cluster (see Discussion), it seems that cluster–cluster interactions predominate in establishing the normally high cooperativity of IP3 action.
Vmax values derived from Hill fits provide a measure of maximal fluorescence at saturating [IP3], and are plotted as a function of [buffer] in Fig. 4B. Vmax declined progressively with increasing concentrations of BAPTA, whereas a biphasic relationship was apparent with EGTA, with an initial potentiation apparent at 23 and 46 μm EGTA.
Differential effects of buffers on kinetics of IP3-evoked Ca2+ signals
As noted earlier, EGTA and BAPTA exerted very different actions on the kinetics of IP3-evoked Ca2+ transients; respectively causing an acceleration or prolongation of their decay. This is illustrated in Fig. 5A, showing superimposed responses to strong photolysis flashes in the presence of increasing [EGTA] or [BAPTA] after normalizing amplitudes to facilitate comparison. Fluorescence decays were fit by mono- or bi-exponential curves (Fig. 5B). In the absence of exogenous buffer, the decay followed a single exponential, with a time constant (τslow) of a few seconds that slowed progressively with increasing photorelease of IP3. In the presence of EGTA the decay phase became markedly biphasic (Fig. 5B), with a prominent fast component (τfast) and smaller slower component (τslow). In the presence of BAPTA a slow decay was prominent, and some oocytes (13/20) also showed a small fast component (Fig. 5B).
Figure 5. Ca2+ transients are speeded by EGTA but prolonged by BAPTA.
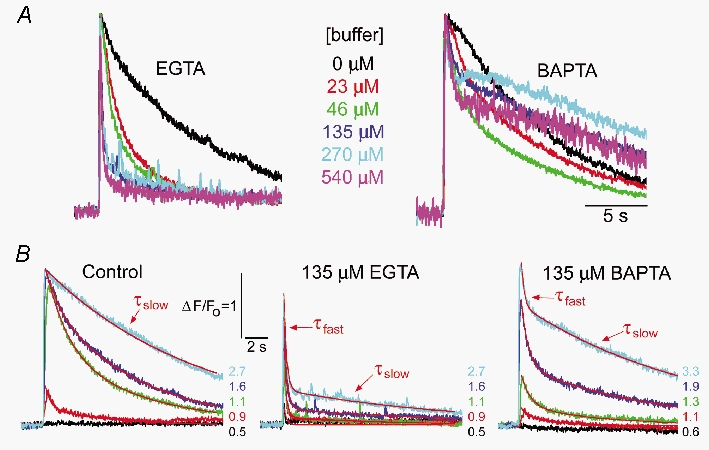
A, families of curves illustrate Ca2+ transients evoked by a fixed photolysis flash (normalized strength 2.5) in the presence of the indicated concentrations of EGTA (left) and BAPTA (right). Responses are scaled to same peak height to facilitate comparison. B, Ca2+ transients evoked by various photolysis flashes of various strengths (normalized strengths indicated) before loading buffer (left); in the same oocyte after loading 135 μm EGTA (middle); and in a different oocyte loaded with 135 μm BAPTA (right). Red curves illustrate fitting of single or double exponentials to derive fast (τfast) and slow (τslow) decay time constants.
Figure 6 shows measurements of time constants of the fast and slow decay components, and their relative weights in contributing toward the overall decline in [Ca2+], plotted as a function of flash strength for various concentrations of EGTA or BAPTA. Several features are evident. (1) In the absence of added buffer, Ca2+ transients showed a mono-exponential decay with a time constant slowing from ≈1 s with just-suprathreshold photorelease of IP3 to nearly 10 s with stronger photo-release (black symbols, Fig. 6A–D). (2) EGTA caused the decay to become markedly biphasic, with an initial fast decline (τfast) that contributed > 80 % of the total decay (Fig. 6E) and shortened with increasing [EGTA] (Fig. 6A). The value of τfast slowed with increasing [IP3] at low (< 50 μm) concentrations of EGTA, but was relatively independent of [IP3] at high (> 135 μm) concentrations of EGTA (Fig. 6A). Following the large, fast decay, the remaining small component declined with a time constant (τslow) that lengthened slightly with increasing photorelease of IP3, and was slower than in the absence of added buffer (Fig. 6C). (3) Addition of BAPTA greatly slowed (τslow 10–30 s) the major part of the Ca2+ signal (Fig. 6D). In those oocytes that displayed biphasic kinetics, the initial fast component was much smaller than with EGTA (Fig. 6E), and its time constant (τfast) was faster than in controls, but slower than with EGTA (Fig. 6B).
Figure 6. EGTA and BAPTA differentially affect the fast and slow decay components of IP3-evoked Ca2+ transients.
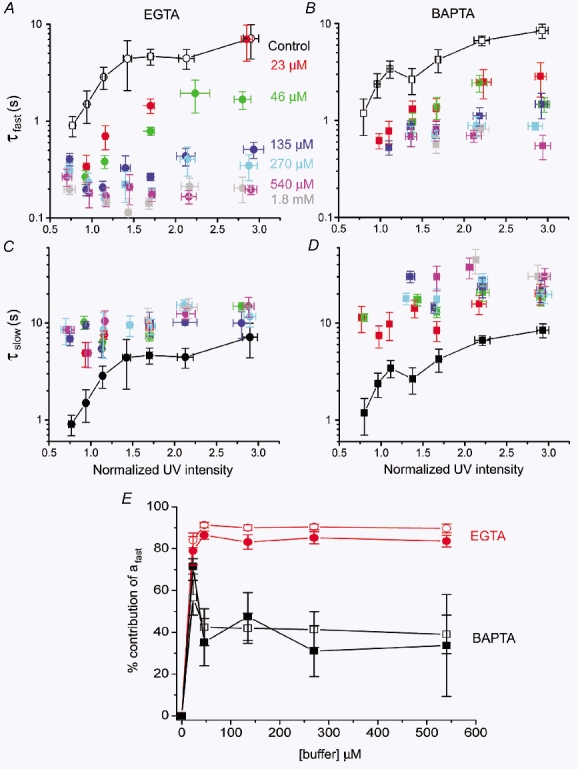
A and C, graphs show, respectively, the time constants for the fast (τfast) and slow (τslow) decay components as a function of photolysis flash strength, in the presence of various concentrations of EGTA. B and D, similar data, with corresponding concentrations of BAPTA. Because the decay of Ca2+ transients without added buffer was mono-exponential, control time constants are repeated in the upper and lower panels (black symbols joined by lines). E, plot shows the relative magnitude of the fast decay component (τfast: measured as shown in Fig. 5B) at various concentrations of EGTA (red symbols) and BAPTA (black symbols). The amplitudes of the fast and slow components (afast and aslow, respectively) were derived at the time of the peak signal from bi-exponential fits, and the graph shows the percentage contribution of afast to the total signal (i.e. afast/(afast+aslow) × 100). Open and filled symbols represent signals evoked, respectively, by weak and strong photolysis flashes.
Decay of Ca2+ transients is dominated by release kinetics, not sequestration
IP3-evoked Ca2+ signals reflect a balance between Ca2+ liberation through IP3Rs into the cytosol and its subsequent removal by processes including buffering, extrusion, re-sequestration via sarco(endo)plasmic reticulum Ca2+-ATPase (SERCA) pumps and diffusion into the enormous interior volume of the oocyte (Parker et al. 1996). The differing effects of EGTA and BAPTA on the kinetics of Ca2+ signals (Fig. 6) might thus arise either through modulation of Ca2+ liberation or through actions on subsequent cytosolic Ca2+ clearance. To discriminate between these possibilities, we examined the effects of buffers on signals arising from Ca2+ flux into the cytosol through voltage-gated N-type Ca2+ channels expressed in the plasma membrane. Opening of these channels by a depolarizing pulse is expected to induce a fixed ‘pulse’ of Ca2+ influx - without complications of CICR as with IP3Rs - and the decay of the resulting Ca2+ transient should thus provide a good measure of cytosolic Ca2+ clearance. Moreover, the spatial distribution of Ca2+ influx across the plasma membrane is expected to closely mimic Ca2+ liberation from IP3-sensitive sites, as these are distributed as a thin shell close under the membrane (Callamaras & Parker, 1999). Ca2+ arising from both sources will thus approximate a 2-dimensional sheet, with subsequent diffusion along a radial axis into the interior of the oocyte.
Oocytes expressing N-type channels were voltage clamped at −60 mV and stepped to +30 mV for 300 ms to induce Ca2+ influx with a duration matching the rise time of IP3-evoked Ca2+ signals (Fig. 7). After the fluorescence signal had returned to baseline (10 s), a UV flash was applied to photorelease IP3 (red arrow). Before loading Ca2+ buffers, signals induced by activation of N-type Ca2+ channels decayed much more rapidly than those evoked by IP3, even though they were of similar amplitude (Fig. 7A). This marked disparity indicates that the slow decay of the IP3-evoked signals did not arise through dye saturation - a conclusion further supported by findings of similar decay rates with the low affinity (Kd≈4 μm) indicator Fluo-4 dextran (not shown).
Figure 7. The decay of IP3-evoked Ca2+ transients is largely determined by the kinetics of Ca2+ release, not sequestration.
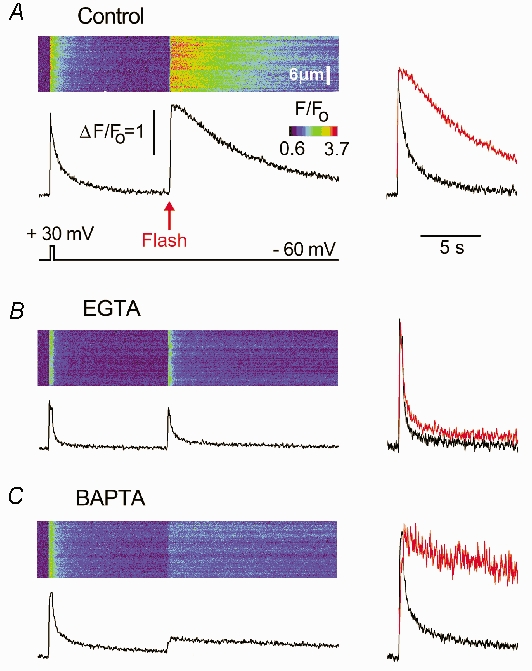
A, line-scan image shows a Ca2+ transient evoked by a 300 ms duration depolarization to +30 mV to activate expressed N-type channels, followed 10 s later by a UV flash to photorelease IP3. The lower trace shows the fluorescence profile averaged across a 6.6 μm region of the scan line, and superimposed traces on the right compare the voltage-gated signal (black) with the IP3-evoked signal (red) after normalizing to the same peak height. B, corresponding image and traces recorded under identical conditions in the same oocyte after loading 270 μm EGTA. C, similar records in a different oocyte loaded with 270 μm BAPTA. Control records in this oocyte before loading BAPTA were similar to one in A.
Thus, the decay of IP3-evoked signals does not simply reflect Ca2+ clearance, but instead is determined primarily by a persistent liberation of Ca2+ through IP3Rs lasting for several seconds. Furthermore, the effects of EGTA and BAPTA are most readily explained by their actions on this prolonged Ca2+ liberation, rather than on Ca2+ clearance from the cytosol. Both EGTA (Fig. 7B) and BAPTA (Fig. 7C) produced only a modest speeding of decay of signals resulting from Ca2+ influx through N-type channels, in marked contrast to their respective actions to accelerate or slow IP3-evoked Ca2+ signals.
Biphasic Ca2+ liberation through IP3Rs
Data from experiments similar to those in Fig. 7 were then used to derive the kinetics of Ca2+ flux into the cytosol through IP3R, assuming that subsequent Ca2+ clearance follows a first order process with a rate constant (k) corresponding to the decay of Ca2+ signals following influx through N-type channels (Fig. 8). Ca2+ flux from intracellular stores is thus proportional to d[Ca2+]/dt+k[Ca2+], where [Ca2+] is the free cytosolic Ca2+ concentration as signalled by changes in fluorescence (Parker et al. 1996). Consistent with previous observations (Parker et al. 1996), control oocytes showed an initial fast spike of IP3-evoked Ca2+ liberation, followed by a lingering ‘tail’ of Ca2+ efflux persisting for several seconds (upper traces, Fig. 8B and D). Although the tail component is small (Fig. 8A and C), over time it contributes one-half or more of the total Ca2+ liberated, and is responsible for the prolonged Ca2+ transients evoked by IP3 under normal conditions. EGTA almost abolished the tail component of Ca2+ liberation (Fig. 8B), thus accelerating the decay of the fluorescent Ca2+ profile (Fig. 8A). Different to this, the tail component was not significantly altered by BAPTA (Fig. 8D), suggesting that prolonged release of Ca2+ through the IP3R may involve cluster–cluster interactions that are selectively disrupted by slow buffers.
Figure 8. EGTA abolishes the ‘tail’ of IP3-evoked Ca2+ liberation that normally prolongs Ca2+ transients.
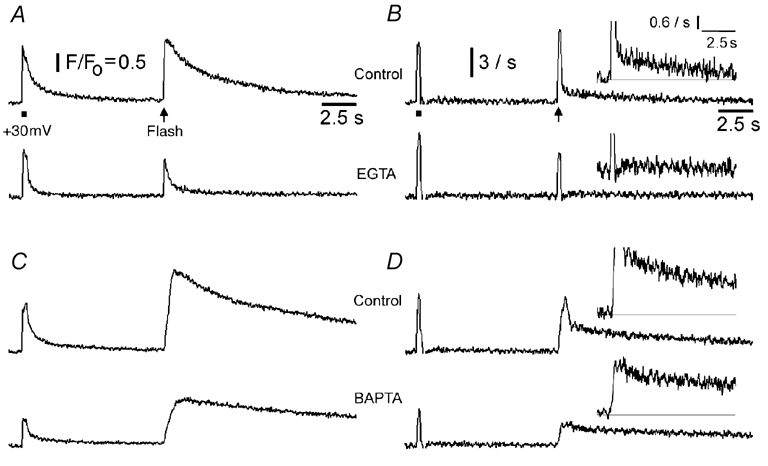
A, fluorescence signals evoked by Ca2+ entry through voltage-gated channels (bar) and by photoreleased IP3 (arrow), before (upper) and after (lower) loading 270 μm EGTA, derived from images similar to Fig. 7A and B. B, rates of Ca2+ flux into the cytosol derived from the records in A, as described in the text. Traces were smoothed using 15 point adjacent averaging. Calibration bar corresponds to a rate of increase in fluorescence (d(ΔF/F0)/dt) of 3 s−1. Inset graphs show the same records with expanded vertical axes to better illustrate the abolition of the persistent ‘tail’ of Ca2+ liberation by EGTA. C and D, similar data from a different oocyte, illustrating the lack of action of BAPTA (270 μm) on the tail of Ca2+ liberation.
Changes in spatial distribution of Ca2+ signals
In addition to their actions on the amplitude and kinetics of IP3-evoked Ca2+ signals, EGTA and BAPTA produced characteristic, and very different, changes in the spatial distribution of Ca2+ signals (Fig. 9). Control oocytes (without added buffer) often display transient localized Ca2+ signals (puffs) in response to photolysis flashes within a narrow ‘window’ of flash strengths (Yao et al. 1995), but these were not evident in the oocytes illustrated, and only spatially diffuse Ca2+ waves were observed (Fig. 9A). However, following injection of EGTA (135 μm), the same oocytes showed discretely localized puffs (Fig. 9B: representative of 14/15 oocytes). With weak stimuli puffs arose at discrete sites following differing latencies. Stronger photorelease of IP3 caused sites to respond almost simultaneously, giving rise to the abrupt, rapidly decaying Ca2+ transient described earlier. Puffs continued after this initial transient and became more frequent, but smaller, with progressively stronger stimuli. In marked contrast, the same concentration (135 μm) of BAPTA abolished Ca2+ responses to weak photolysis flashes (10/18 oocytes with normalized flash strengths <1.1), and at higher flash strengths oocytes showed only spatially uniform and slowly decaying Ca2+ signals. We never (0/24 oocytes) observed localized Ca2+ puffs.
Figure 9. 2+ signals into discrete, autonomous units, whereas BAPTA promotes spatially uniform global signals.

A, line-scan images and fluorescence profiles (averaged over 4 μm regions) showing responses to photolysis flashes (red arrows) of increasing strength (indicated in normalized units) before injecting buffer. B, corresponding records in the same oocyte after loading 135 μm EGTA. Two representative fluorescence profiles are illustrated from each image, recorded at different puff sites (arrowed). C, corresponding records in a different oocyte loaded with 135 μm BAPTA. Records in this oocyte before loading BAPTA were similar to those in A.
DISCUSSION
We show that two mobile Ca2+ buffers with similar affinities but very different binding kinetics affect the spatiotemporal characteristics of IP3-evoked Ca2+ liberation in contrasting ways. EGTA - a buffer with a slow binding rate - causes Ca2+ signals to become more transient and ‘balkanizes’ Ca2+ liberation such that individual release sites act autonomously to generate discrete puffs rather than functioning in concert to generate Ca2+ waves. On the other hand, BAPTA - a fast buffer - prolongs Ca2+ transients and promotes a ‘globalization’ of Ca2+ signalling as a spatially uniform elevation without evidence of local release events. These strikingly different actions cannot be explained merely by chelation of Ca2+ subsequent to its liberation into the cytosol, because Ca2+ signals generated by influx through voltage-gated channels were affected to a much lesser degree by EGTA and BAPTA (Fig. 7). Moreover, they do not arise through changes in resting free [Ca2+] or alterations in Ca2+ store filling, as similar results were obtained whether buffers were injected either without Ca2+ or saturated with Ca2+. Instead, EGTA and BAPTA appear to act over different time and distance scales to modulate the processes of Ca2+ diffusion and CICR that shape the regenerative nature of IP3-evoked Ca2+ liberation.
Spatiotemporal scales of Ca2+ signalling
IP3Rs are organized in Xenopus oocytes as tight (ca 50 nm diameter) clusters containing several tens of channels, with a mean cluster–cluster spacing of ≈3 μm (Callamaras et al. 1998a,b; Swillens et al. 1999; Shuai, 2002,Jung, 2003). CICR can thus act over two very different spatiotemporal scales - via fast diffusion over nanometre distances between IP3Rs within a cluster to give local Ca2+ puffs, and much more slowly across micrometre distances between clusters to generate propagating saltatory waves (Fig. 10A: Yao et al. 1995; Bootman et al. 1997; Berridge, 1997; Callamaras et al. 1998a,b; Marchant & Parker, 2000). Depending on their kinetics, buffers may selectively act on one or other of these processes.
Figure 10. Cartoon illustrating the distribution of Ca2+ ions around IP3R clusters, and how communication between IP3Rs may be affected by mobile buffers with differing binding kinetics.
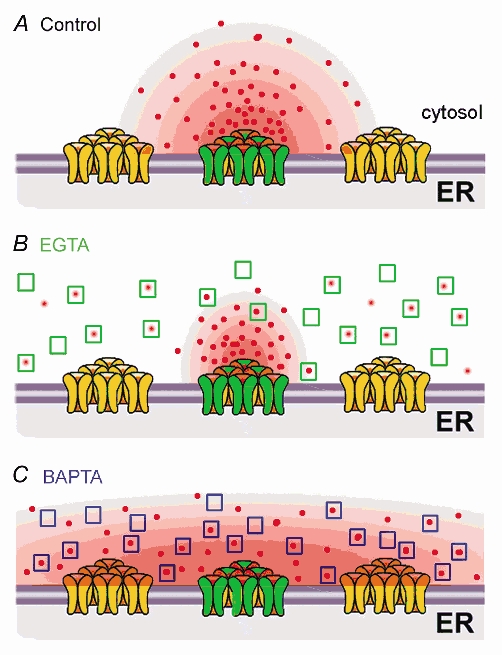
A, without added buffer, Ca2+ ions liberated through open IP3 at a cluster diffuse a few micrometres and, depending on [IP3], may or may not trigger CICR at adjacent clusters. B, EGTA acts too slowly to disrupt Ca2+ diffusion and CICR within clusters, but binds Ca2+ ions as they diffuse between clusters and rapidly ‘shuttles’ them away, so as to disrupt cluster–cluster interactions. C, BAPTA binds Ca2+ ions sufficiently quickly to disrupt Ca2+ communication between IP3Rs within individual clusters, whilst promoting Ca2+ communication between clusters by acting as a rapid Ca2+ shuttle.
Key factors determining the interactions of buffers with Ca2+ ions are summarized in Table 1, and include: (1) the mean time (τcapture) and distance (dcapture) that a Ca2+ ion will diffuse before it becomes bound to a buffer molecule; (2) the mean time (dwell time; τdwell) a Ca2+ ion will remain bound to a buffer before dissociating, and the corresponding mean distance (‘shuttle’ distance; dshutttle) by which the Ca2+-buffer complex will diffuse before dissociation. Further, diffusion of Ca2+ ions in the cytosol is slowed by binding to endogenous, immobile buffers. Less than 10 % of the total Ca2+ ions in the cytosol are free at any given time, and the apparent diffusion coefficient for Ca2+ in the oocyte is thereby slowed about 10-fold as compared to free aqueous diffusion (respectively, 20 vs. 200 μm2 s−1: Allbritton et al. 1992; Yao et al. 1995). Buffers, such as EGTA and BAPTA can thus act as a ‘shuttle’ to facilitate diffusion (Stern, 1992; Roberts, 1994) because, when bound to a mobile buffer molecule, a Ca2+ ion is ‘protected’ from binding to immobile sites and is thus carried appreciable distances at a rate only a little slower than that for free aqueous diffusion.
Buffer actions on inter- and intra-cluster signalling
We present here a qualitative model to account for the actions of EGTA and BAPTA on IP3-evoked Ca2+ signalling, which is summarized in Fig. 10. This is necessarily simplified, and will require mathematical modelling for rigorous verification, but nevertheless offers useful insights.
Dealing first with EGTA, we consider how cytosolic diffusion of Ca2+ would be altered in the presence of 270 μm of this buffer. Its slow on-rate implies that a Ca2+ ion will diffuse an average distance of about 1 μm before binding to an EGTA molecule (Table 1). EGTA is thus expected to have little effect on short-range Ca2+ feedback over nanometre distances between individual IP3Rs within a cluster, but it will efficiently capture Ca2+ ions diffusing over micrometre distances between neighbouring clusters (Roberts, 1994; Horne & Meyer, 1997; Song et al. 1998; Callamaras et al. 1998a,b; Kidd et al. 1999). Once bound Ca2+ ions will be unavailable to trigger CICR for prolonged periods (τdwell = 0.7-2 s: Falcke, 2003), and Ca2+ bound to EGTA will be shuttled long distances (dshuttle = 28–50 μm) before being ‘dumped’ in the interior of the oocyte where release sites are absent (Callamaras & Parker, 1999). The overall effect will thus be to more sharply restrict Ca2+ signals around individual clusters, and reduce free [Ca2+] between clusters so as to functionally uncouple them (Fig. 10B). In agreement, EGTA has little effect on puffs (Callamaras & Parker, 2000), yet strongly inhibits Ca2+ waves and dissociates global IP3-evoked Ca2+ signals into discrete, localized Ca2+ release events (Fig. 9: Horne & Meyer, 1997; Kidd et al. 1999; Callamaras & Parker, 2000).
In contrast, BAPTA has an on-rate about 100 times faster than EGTA, and binds Ca2+ ions within a distance (tens of nanometres; Table 1) comparable to the dimensions of an IP3R cluster, thereby reducing CICR between individual receptors within the cluster. Subsequently, Ca2+ bound to BAPTA will rapidly (ca 10 ms) be ‘shuttled’ a few micrometres before dissociating. This is comparable to the inter-cluster spacing, so whereas BAPTA will inhibit intra-cluster feedback by Ca2+ it may actually facilitate inter-cluster communication. Moreover, discrete puffs arise only when the Ca2+ release time at a site (ca 50 ms: Sun et al. 1998) is short in relation to the inter-cluster diffusion time (Dawson et al. 1999; Shuai & Jung, 2003). This condition is fulfilled in the absence of exogenous buffer, but the ability of BAPTA to rapidly shuttle Ca2+ and thereby speed its diffusion effectively ‘smears’ the distribution of Ca2+, so that the endoplasmic reticulum (e.r.) approximates a deterministic, spatially continuous Ca2+ source, rather than an array of loosely coupled, discrete stochastic sources (Dawson et al. 1999; Shuai & Jung, 2003).
Cooperativity of IP3-evoked Ca2+ release
In the absence of added buffer, Ca2+ liberation in the oocyte increases steeply (nH > 6) with [IP3]. This is likely to reflect both the requirement for binding of multiple IP3 molecules to open the homo-tetrameric IP3R, and positive cooperativity introduced by CICR. Our results suggest that the latter is the more prominent factor. High (millimolar) concentrations of both EGTA and BAPTA reduced nH to ≈2, suggesting that binding of two (or possibly one) molecules of IP3 is sufficient for channel opening. Interestingly, moderate concentrations of EGTA (approximately 25 μm) caused a much greater reduction in nH than the same concentrations of BAPTA, further implying that inter-, rather than intra-cluster interactions are the main factor contributing to the high cooperativity under physiological conditions.
Potentiation of Ca2+ signalling by low [EGTA]
Ca2+ signals evoked by strong photorelease of IP3 became progressively smaller with increasing concentrations of EGTA and BAPTA, as might be expected because the buffers will compete with the indicator dye for available Ca2+ as well as reducing Ca2+ liberation by impairing CICR. On the other hand, EGTA (but not BAPTA) actually potentiated responses evoked by weak photorelease of IP3 by as much as 100 % (Fig. 3C). This surprising finding is consistent with a recent demonstration that slow, mobile Ca2+ buffers (parvalbumin and EGTA) promote puff activity in Xenopus oocytes, probably by sensitizing IP3R to resting levels of IP3 (John et al. 2001). Potentiation of Ca2+ excitability by expression of specific endogenous buffers may, therefore, play an important role in regulating cellular signalling. The underlying mechanism is unclear (John et al. 2001), but effects on cluster–cluster interactions are implicated because potentiation is most prominent with slow buffers, appears not to result from changes in resting free [Ca2+], and does not correlate with binding of Ca2+-free forms of different buffers to IP3Rs (Richardson & Taylor, 1993; John et al. 2001).
Cluster–cluster interactions prolong Ca2+ liberation
Ca2+ signals during local Ca2+ puffs persist for only a few hundred milliseconds (Yao et al. 1995; Sun et al. 1998), whereas Ca2+ elevations during global waves decay over several seconds (e.g. Fig. 2). We originally thought that this difference might reflect differences in Ca2+ clearance, and that the kinetics of Ca2+ liberation were similar during these local and global events. Specifically, it seemed that Ca2+ liberated at a ‘point’ source (as during a puff) would rapidly dissipate by 3-dimensional diffuse into the vast volume of the oocyte, whereas the spatially extensive Ca2+ release during a wave would be cleared primarily by slower processes such as sequestration back into the e.r. (Yao et al. 1995). However, the experiment of Fig. 7 indicates that this is not a full explanation, because IP3-evoked Ca2+ signals decayed much more slowly than equivalent signals evoked by extracellular Ca2+ influx through voltage-gated channels. Instead, we conclude that IP3-evoked waves are prolonged largely because Ca2+ efflux from the e.r. is biphasic. That is to say, a large, but rapidly terminating flux during the rising phase of the wave is followed by a ‘tail’ of Ca2+ efflux which, though small, is sufficiently prolonged that it greatly slows the decline of cytosolic [Ca2+] (Fig. 8A). The actions of EGTA and BAPTA on global Ca2+ signals to respectively speed and slow their decay can then be understood in terms of their relative effects on the fast and slow components of Ca2+ efflux. EGTA almost abolishes the slow component while having little effect on the fast component, whereas BAPTA selectively attenuates the fast component and prolongs the slow component (Fig. 8B and C).
Termination of Ca2+ flux through IP3Rs thus appears to involve two distinct processes. Flux at individual clusters acting in isolation (at low [IP3], or after uncoupling with EGTA) ceases abruptly and stochastically within ≈100 ms, and shows an ‘adaptive’ property such that it can be reactivated by further elevation of [IP3] (Callamaras & Parker, 2000). On the other hand, clusters recruited during a wave show an initial large Ca2+ efflux with a time course similar to that of puffs, and then abruptly transition to a state in which they continue to ‘leak’ Ca2+ at a low rate for several seconds. The mechanisms underlying this transition remain unknown, but the striking action of EGTA to attenuate the slow release suggests that it involves diffusional spread of Ca2+ between clusters, such that each continues to maintain others in an activated state. Moreover, the importance of the slow release component in shaping Ca2+ transients further emphasizes the role of buffers in cell signalling. Although the slow Ca2+ flux is small, its persistence means that the total amount of Ca2+ liberated is greater than that during the fast component; and attenuation of the slow component by EGTA shortens the duration of Ca2+ spikes as much as 10-fold.
Physiological roles of buffers in cellular Ca2+ signalling
Our results point to the importance of buffering kinetics in shaping the spatial and temporal patterning of Ca2+ signals generated by intracellular Ca2+ liberation through Ca2+-sensitive release channels - effects that are more complex than for signals arising from a fixed ‘pulse’ of Ca2+, as with Ca2+ entry through voltage-gated channels (e.g. Lee et al. 2000). For convenience we utilized the synthetic buffers EGTA and BAPTA, which are not normally present within cells. However, preliminary experiments with endogenous mobile Ca2+-binding proteins (Dargan et al. 2003) point out the physiological relevance of our results. In particular, parvalbumin - a mobile Ca2+ buffer with slow Ca2+ binding kinetics (Lee et al. 2000b; Schmidt et al. 2003) - balkanized Ca2+ signals and accelerated their decay similar to EGTA; whereas the fast buffer calretinin (Edmonds et al. 2000) produced BAPTA-like effects. Although there are obvious differences between small-molecule and protein buffers (such as differing mobility and possibility of protein-protein interactions), the emergent theme is that the kinetics of a buffer are the key parameters defining its actions in shaping IP3-mediated Ca2+ signals. Moreover, this theme is likely to extend to mammalian systems because the clustered distribution of IP3Rs in the Xenopus oocyte is mirrored in diverse mammalian cells (Bootman et al. 1997; Simpson et al. 1997). We finally note that ryanodine receptors (RyRs), the other major class of intracellular Ca2+ release channel, also communicate via CICR (Berridge, 1997), so that their signalling may similarly be susceptible to shaping by mobile Ca2+ buffers.
Acknowledgments
Plasmids containing cDNA clones coding for the N-type Ca2+ channel α1B-d and β3 subunits were a kind gift from Diane Lipscombe. We thank Dr Angelo Demuro for in vitro transcription, expressing N-type Ca2+ channels in oocytes, and helping with experiments of Fig. 7; and Dr Brian Edmonds for helpful discussion. Funded by NIH grants GM48071 and GM58329.
REFERENCES
- Allbritton NL, Meyer T, Stryer L. Range of messenger action of calcium ion and inositol 1,4,5-trisphosphate. Science. 1992;258:1812–1815. doi: 10.1126/science.1465619. [DOI] [PubMed] [Google Scholar]
- Andressen C, Blumcke I, Celio MR. Calcium-binding proteins: selective markers of nerve cells. Cell Tissue Res. 1993;271:181–208. doi: 10.1007/BF00318606. [DOI] [PubMed] [Google Scholar]
- Berridge MJ. Elementary and global aspects of calcium signalling. J Physiol. 1997;499:291–306. doi: 10.1113/jphysiol.1997.sp021927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berridge MJ, Lipp P, Bootman MD. The versatility and universality of calcium signalling. Nat Rev Mol Cell Biol. 2000;1:11–21. doi: 10.1038/35036035. [DOI] [PubMed] [Google Scholar]
- Bezprozvanny I, Watras J, Ehrlich BE. Bell-shaped calcium-response curves of Ins(1,4,5)P3- and calcium-gated channels from endoplasmic reticulum of cerebellum. Nature. 1991;351:751–754. doi: 10.1038/351751a0. [DOI] [PubMed] [Google Scholar]
- Bootman M, Niggli E, Berridge M, Lipp P. Imaging the hierarchical Ca2+ signalling system in HeLa cells. J Physiol. 1997;499:307–314. doi: 10.1113/jphysiol.1997.sp021928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callamaras N, Marchant JS, Sun XP, Parker I. Activation and co-ordination of InsP3-mediated elementary Ca2+ events during global Ca2+ signals in Xenopus oocytes. J Physiol. 1998a;509:81–91. doi: 10.1111/j.1469-7793.1998.081bo.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callamaras N, Parker I. Caged inositol 1,4,5-trisphosphate for studying release of Ca2+ from intracellular stores. Methods Enzymol. 1998b;291:380–403. doi: 10.1016/s0076-6879(98)91024-2. [DOI] [PubMed] [Google Scholar]
- Callamaras N, Parker I. Radial localization of inositol 1,4,5-trisphosphate-sensitive Ca2+ release sites in Xenopus oocytes resolved by axial confocal linescan imaging. J Gen Physiol. 1999;113:199–213. doi: 10.1085/jgp.113.2.199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callamaras N, Parker I. Phasic characteristic of elementary Ca2+ release sites underlies quantal responses to IP3. EMBO J. 2000;19:3608–3617. doi: 10.1093/emboj/19.14.3608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callamaras N, Sun XP, Ivorra I, Parker I. Hemispheric asymmetry of macroscopic and elementary calcium signals mediated by InsP3 in Xenopus oocytes. J Physiol. 1998b;511:395–405. doi: 10.1111/j.1469-7793.1998.395bh.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dargan SL, Demuro A, Marchant J, Callamaras N, Schwaller B, Parker I. Shaping of IP3-mediated Ca2+ signals by Ca2+ buffers and Ca2+ binding proteins. Biophys J. 2003;84(suppl.):384a. [Google Scholar]
- Dawson SP, Keizer J, Pearson JE. Fire-diffuse-fire model of dynamics of intracellular calcium waves. Proc Natl Acad Sci U S A. 1999;96:6060–6063. doi: 10.1073/pnas.96.11.6060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edmonds B, Reyes R, Schwaller B, Roberts WM. Calretinin modifies presynaptic calcium signaling in frog saccular hair cells. Nat Neurosci. 2000;3:786–790. doi: 10.1038/77687. [DOI] [PubMed] [Google Scholar]
- Falcke M. Buffers and oscillations in intracellular Ca2+ dynamics. Biophys J. 2003;84:28–41. doi: 10.1016/S0006-3495(03)74830-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finch EA, Turner TJ, Goldin SM. Calcium as a coagonist of inositol 1,4,5-trisphosphate-induced calcium release. Science. 1991;252:443–446. doi: 10.1126/science.2017683. [DOI] [PubMed] [Google Scholar]
- Hirota J, Michikawa T, Miyawaki A, Furuichi T, Okura I, Mikoshiba K. Kinetics of calcium release by immunoaffinity-purified inositol 1,4,5-trisphosphate receptor in reconstituted lipid vesicles. J Biol Chem. 1995;270:19046–19051. doi: 10.1074/jbc.270.32.19046. [DOI] [PubMed] [Google Scholar]
- Horne JH, Meyer T. Elementary calcium-release units induced by inositol trisphosphate. Science. 1997;276:1690–1693. doi: 10.1126/science.276.5319.1690. [DOI] [PubMed] [Google Scholar]
- Iino M. Biphasic Ca2+ dependence of inositol 1,4,5-trisphosphate-induced Ca2+ release in smooth muscle cells of the guinea pig taenia caeci. J Gen Physiol. 1990;95:1103–1122. doi: 10.1085/jgp.95.6.1103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- John LM, Mosquera-Caro M, Camacho P, Lechleiter JD. Control of IP3-mediated Ca2+ puffs in Xenopus laevis oocytes by the Ca2+-binding protein parvalbumin. J Physiol. 2001;535:3–16. doi: 10.1111/j.1469-7793.2001.t01-2-00003.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kidd JF, Fogarty KE, Tuft RA, Thorn P. The role of Ca2+ feedback in shaping InsP3-evoked Ca2+ signals in mouse pancreatic acinar cells. J Physiol. 1999;520:187–201. doi: 10.1111/j.1469-7793.1999.00187.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lechleiter JD, Clapham DE. Molecular mechanisms of intracellular calcium excitability in X. laevis oocytes. Cell. 1992;69:283–294. doi: 10.1016/0092-8674(92)90409-6. [DOI] [PubMed] [Google Scholar]
- Lee SH, Rosenmund C, Schwaller B, Neher E. Differences in Ca2+ buffering properties between excitatory and inhibitory hippocampal neurons from the rat. J Physiol. 2000a;525:405–418. doi: 10.1111/j.1469-7793.2000.t01-3-00405.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee SH, Schwaller B, Neher E. Kinetics of Ca2+ binding to parvalbumin in bovine chromaffin cells: implications for [Ca2+] transients of neuronal dendrites. J Physiol. 2000b;525:419–432. doi: 10.1111/j.1469-7793.2000.t01-2-00419.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Z, Haus S, Edgerton J, Lipscombe D. Identification of functionally distinct isoforms of the N-type Ca2+ channel in rat sympathetic ganglia and brain. Neuron. 1997;18:153–166. doi: 10.1016/s0896-6273(01)80054-4. [DOI] [PubMed] [Google Scholar]
- Mak DO, McBride S, Foskett JK. Inositol 1,4,5-trisphosphate activation of inositol trisphosphate receptor Ca2+ channel by ligand tuning of Ca2+ inhibition. Proc Natl Acad Sci U S A. 1998;95:15821–15825. doi: 10.1073/pnas.95.26.15821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marchant JS, Parker I. Functional interactions in Ca2+ signaling over different time and distance scales. J Gen Physiol. 2000;116:691–696. doi: 10.1085/jgp.116.5.691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marchant JS, Taylor CW. Cooperative activation of IP3 receptors by sequential binding of IP3 and Ca2+ safeguards against spontaneous activity. Curr Biol. 1997;7:510–518. doi: 10.1016/s0960-9822(06)00222-3. [DOI] [PubMed] [Google Scholar]
- Meyer T, Wensel T, Stryer L. Kinetics of calcium channel opening by inositol 1,4,5-trisphosphate. Biochemistry. 1990;29:32–37. doi: 10.1021/bi00453a004. [DOI] [PubMed] [Google Scholar]
- Morris SA, Correa V, Cardy TJ, O'Beirne G, Taylor CW. Interactions between inositol trisphosphate receptors and fluorescent Ca2+ indicators. Cell Calcium. 1999;25:137–142. doi: 10.1054/ceca.1998.0016. [DOI] [PubMed] [Google Scholar]
- Nagerl UV, Novo D, Mody I, Vergara JL. Binding kinetics of calbindin-D(28k) determined by flash photolysis of caged Ca2+ Biophys J. 2000;79:3009–3018. doi: 10.1016/S0006-3495(00)76537-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naraghi M. T-jump study of calcium binding kinetics of calcium chelators. Cell Calcium. 1997;22:255–268. doi: 10.1016/s0143-4160(97)90064-6. [DOI] [PubMed] [Google Scholar]
- Neher E. Calcium buffers in flash-light. Biophys J. 2000;79:2783–2784. doi: 10.1016/S0006-3495(00)76517-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker I, Callamaras N, Wier WG. A high-resolution, confocal laser-scanning microscope and flash photolysis system for physiological studies. Cell Calcium. 1997;21:441–452. doi: 10.1016/s0143-4160(97)90055-5. [DOI] [PubMed] [Google Scholar]
- Parker I, Yao Y, Ilyin V. Fast kinetics of calcium liberation induced in Xenopus oocytes by photoreleased inositol trisphosphate. Biophys J. 1996;70:222–237. doi: 10.1016/S0006-3495(96)79565-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parys JB, Sernett SW, Delisle S, Snyder PM, Welsh MJ, Campbell KP. Isolation, characterization, and localization of the inositol 1,4,5-trisphosphate receptor protein in Xenopus laevis oocytes. J Biol Chem. 1992;267:18776–18782. [PubMed] [Google Scholar]
- Richardson A, Taylor CW. Effects of Ca2+ chelators on purified inositol 1,4,5-trisphosphate (InsP3) receptors and InsP3-stimulated Ca2+ mobilization. J Biol Chem. 1993;268:11528–11533. [PubMed] [Google Scholar]
- Roberts WM. Localization of calcium signals by a mobile calcium buffer in frog saccular hair cells. J Neurosci. 1994;14:3246–3262. doi: 10.1523/JNEUROSCI.14-05-03246.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt H, Brown EB, Schwaller B, Eilers J. Diffusional mobility of parvalbumin in spiny dendrites of cerebellar purkinje neurons quantified by fluorescence recovery after photobleaching. Biophys J. 2003;84:2599–2608. doi: 10.1016/S0006-3495(03)75065-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwaller B, Meyer M, Schiffmann S. ‘New’ functions for ‘old’ proteins: The role of the Ca2+ binding proteins calbindin D-28K, calretinin and parvalbumin in cerebellar physiology. Studies with knockout mice. Cerebellum. 2002;1:241–258. doi: 10.1080/147342202320883551. [DOI] [PubMed] [Google Scholar]
- Shuai JW, Jung P. Stochastic properties of Ca2+ release of inositol 1,4,5-trisphosphate receptor clusters. Biophys J. 2002;83:87–97. doi: 10.1016/S0006-3495(02)75151-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shuai JW, Jung P. Optimal ion channel clustering for intracellular calcium signaling. Proc Natl Acad Sci U S A. 2003;100:506–510. doi: 10.1073/pnas.0236032100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson PB, Mehotra S, Lange GD, Russell JT. High density distribution of endoplasmic reticulum proteins and mitochondria at specialized Ca2+ release sites in oligodendrocyte processes. J Biol Chem. 1997;272:22654–22661. doi: 10.1074/jbc.272.36.22654. [DOI] [PubMed] [Google Scholar]
- Song LS, Sham JS, Stern MD, Lakatta EG, Cheng H. Direct measurement of SR release flux by tracking ‘Ca2+ spikes’ in rat cardiac myocytes. J Physiol. 1998;512:677–691. doi: 10.1111/j.1469-7793.1998.677bd.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stern MD. Buffering of calcium in the vicinity of a channel pore. Cell Calcium. 1992;13:183–192. doi: 10.1016/0143-4160(92)90046-u. [DOI] [PubMed] [Google Scholar]
- Sun XP, Callamaras N, Marchant JS, Parker I. A continuum of InsP3-mediated elementary Ca2+ signalling events in Xenopus oocytes. J Physiol. 1998;509:67–80. doi: 10.1111/j.1469-7793.1998.067bo.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swillens S, Dupont G, Combettes L, Champeil P. From calcium blips to calcium puffs: theoretical analysis of the requirements for interchannel communication. Proc Natl Acad Sci U S A. 1999;96:13750–13755. doi: 10.1073/pnas.96.24.13750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsien RY. New calcium indicators and buffers with high selectivity against magnesium and protons: design, synthesis, and properties of prototype structures. Biochemistry. 1980;19:2396–2404. doi: 10.1021/bi00552a018. [DOI] [PubMed] [Google Scholar]
- Yao Y, Choi J, Parker I. Quantal puffs of intracellular Ca2+ evoked by inositol trisphosphate in Xenopus oocytes. J Physiol. 1995;482:533–553. doi: 10.1113/jphysiol.1995.sp020538. [DOI] [PMC free article] [PubMed] [Google Scholar]