

Abstract
Most epithelial tissues self-renew throughout adult life due to the presence of multipotent stem cells and/or unipotent progenitor cells. Epithelial stem cells are specified during development and are controlled by epithelial-mesenchymal interactions. Despite morphological and functional differences among epithelia, common signaling pathways appear to control epithelial stem cell maintenance, activation, lineage determination, and differentiation. Additionally, deregulation of these pathways can lead to human disorders including cancer. Understanding epithelial stem cell biology has major clinical implications for the diagnosis, prevention, and treatment of human diseases, as well as for regenerative medicine.
Epithelia are continuous sheets of tightly linked cells that constitute the surfaces (such as the epidermis and corneal epithelium) and linings (such as the digestive, respiratory, and uro-genital epithelia) of the body. At these locations, epithelia not only provide a protective envelope against the external environment but also regulate water and nutrient absorption as well as glandular secretions.
Although epithelia can be multilayered (stratified) or single-layered (simple) and may be derived from ectoderm, mesoderm, or endoderm, the epithelial tissues of the body share several molecular and cellular characteristics. In development, epithelia begin as a sheet of cells that adhere to a basement membrane, rich in extracellular matrix (ECM) and growth factors that are produced and deposited at the interface of the epithelium and the underlying mesenchyme. Epithelial cells express trans-membrane integrin heterodimers that bind to ECM ligands in the basement membrane and provide a link to the cellular cytoskeleton (Giancotti and Tarone, 2003). In addition, focal adhesions and hemidesmosomes regulate the dynamics of adhesion and detachment of cells and their underlying ECM, thus mediating cell migration, stratification, and differentiation (Fuchs and Raghavan, 2002; Watt, 2002). Epithelial cells also make intercellular connections through the formation of adherens junctions, tight junctions, and desmosomes that enable epithelial cells to communicate and function as a sheet. In conjunction with integrins, intercellular junctions and associated components distinguish the apical, basal, and lateral surfaces of the cell, which are essential for establishing epithelial cell polarity (Shin et al., 2006).
Most epithelia need to constantly replace damaged or dead cells throughout the life of the animal. The process of continual cell replacement is called tissue homeostasis and is critical for the maintenance of adult tissues. Typically, epithelial tissue homeostasis is maintained through the presence of stem cells. Stem cells are functionally defined by their ability to self-renew and to differentiate into the cell lineages of their tissue of origin (Moore and Lemischka, 2006). Once activated, epithelial stem cells can generate proliferating progeny, which are often referred to as transiently amplifying (TA) cells. In their normal environment, TA cells will divide actively for a restricted period of time, expanding the cellular pool that will then differentiate along a particular cell lineage to make the tissue.
The homeostatic replacement of cells varies substantially among different epithelia. The epithelium of the intestine completely self-renews within ∼5 days. By contrast, interfollicular epidermis takes ∼4 weeks to renew, whereas the lung epithelium can take as long as 6 months to be replaced. In addition, some epithelia present a cyclic mode of tissue homeostasis. Hair follicles, for example, cycle continuously through bouts of hair growth (anagen), degeneration (catagen), and rest (telogen) (Blanpain and Fuchs, 2006). Similarly, the mammary gland proceeds through cycles of growth and degeneration during and following pregnancy (Hennighausen and Robinson, 2005).
Unless the epithelial stem cells and associated mesenchyme are permanently damaged, most epithelia are also able to repair their tissues following injuries. Typically, tissue regeneration upon wounding involves recruitment of epithelial stem cells to replace the damaged cells. The adult liver offers an unusual example: although its epithelial cells do not turn over significantly under physiological conditions, they have an impressive capacity to regenerate tissue after injury. Oddly, following acute injuries, this tissue repair appears to occur without obvious participation of multipotent stem cells. Rather, liver regeneration after a partial hepatectomy occurs through proliferation of hepatocytes (Taub, 2004). A similar mode of regeneration called self-duplication has also been proposed to account for the renewal of pancreatic islet cells (Dor et al., 2004). There is still no consensus as to whether adult cells with the capacity to regenerate and/or contribute to only a single lineage should be referred to as unipotent stem cells, unipotent progenitors, or simply cells that have retained proliferative potential.
Epithelial Stem Cells and Their Lineages
Epithelial stem cells can generate tissues that display a fascinating array of cellular architectures, each of which are specifically tailored for distinct functions (Figure 1). In this review, we will focus on four well-characterized epithelial stem cells whose tissues possess diverse architectural designs and physiology: intestine, epidermis, mammary gland, and cornea. We will briefly introduce the organization of these tissues into their functional units, delineate which cell lineages compose the unit, and discuss where the stem cells are located within each unit and their routes of activation and differentiation.
Figure 1. Stem Cell Location in Different Epithelial Tissues.

The intestinal crypt. The putative stem cells of the intestine (red) reside in a narrow band near the base of the crypt. These stem cells are thought to move downward to differentiate into Paneth cells (blue) or upward to generate proliferative transiently amplifying (TA) cells (orange). The intestinal TA cells differentiate upward into three lineages, enteroendocrine cells, goblet cells, and enterocytes, to form the villus. The corneal limbus. The cornea (blue) is a stratified epithelium that is flanked by the limbus (red) and the conjunctiva (purple). Corneal stem cells are thought to reside in the limbus region. The hair-follicle bulge. Label-retaining stem cells of the hair follicle reside below the sebaceous gland in a region known as the bulge, which is connected to the arector pili muscle. During periods of rest, bulge stem cells form the base of the follicle, which is adjacent to the specialized mesenchymal cells (dermal papillae). At the start of each hair cycle, stem cells at the bulge base become activated to form the highly proliferative new hair germ. As the germ grows, a proliferative compartment of TA cells (matrix cells) engulfs the dermal papilla at the base. These cells progress to differentiate to form seven concentric shells of discrete cell lineages, which are from outer to inner: the companion layer, the three layers of the inner root sheath, and the three layers of the hair shaft. These differentiated layers are surrounded by the outer root sheath, which extends below the bulge and is thought to contain stem cells that continue to migrate down to the follicle base during the growth phase of the hair cycle (Oshima et al., 2001). The interfollicular epidermis is a stratified epithelium with a basal layer that contains unipotent progenitor cells and TA cells. Basal cells differentiate upward to form the spinous, granular, and stratum corneum layers of the epidermis. The mammary gland terminal end bud. Stem cells of the mammary gland reside in the terminal end bud at the end of the mammary duct. The end of the terminal end bud is composed of body cells surrounded by cap cells. During pregnancy, the stem cells form a complex ductal structure containing ductal cells that end in the alveoli that produce milk components.
The small intestine is a simple epithelium composed of crypt-villus units (Radtke and Clevers, 2005), which function to absorb water and nutrients and to form a functional barrier to protect against ingested pathogens. In the adult, the stem cells and their TA progeny reside in a region near the base of the crypt region, and as they migrate out of their niche, they cease to proliferate and initiate differentiation into the different cell lineages of the mature villi. Intestinal stem cells can differentiate into four different cell lineages: the absorptive enterocytes, mucin-secreting-goblet cells, peptide hormone-secreting neuroendocrine cells, and microbicide-secreting Paneth cells. The majority of cells in the villi are enterocytes with a few goblet and neuroendocrine cells located at various intervals. These three lineages form and mature as they migrate up the crypt to the tip of the villus. During tissue homeostasis, these cells are replaced and subsequently exfoliated into the intestinal lumen. Paneth cells differentiate as they travel down to the base of the crypt (Figure 1).
Encasing the body, the skin epidermis is composed of pilo-sebaceous units containing a hair follicle, sebaceous gland, and interfollicular epidermis (Blanpain and Fuchs, 2006). The interfollicular epidermis is a stratified squamous epithelium consisting of an innermost (basal) layer of proliferative cells that differentiate outward to form the distinctive suprabasal layers of the differentiated tissue: spinous, granular, and finally outermost stratum corneum layers to form an impermeable body surface. Resident basal stem cells strongly adhere to their underlying basement membrane and maintain homeostasis of the interfollicular epidermis by continually replenishing the suprabasal terminally differentiating cells as dead stratum corneum cells (squames) eventually reach and are sloughed from the skin surface.
The hair follicle is formed during embryogenesis as an appendage of the epidermis. A condensation of specialized mesenchymal cells (dermal papilla) in the dermis stimulates cells within the overlying epidermal basal layer. The epidermal cells respond by changing their shape to develop a bud, or “placode,” of hair progenitor cells that subsequently proliferate and grow downward. Postnatally, the base of the mature follicle contains rapidly proliferating (TA) matrix cells, which after several rounds of cell division differentiate upward to form the hair and its surrounding channel, the inner root sheath. Eventually, the supply of differentiating matrix cells ceases, and the follicle regresses and draws the dermal papillae upward. After the lower two-thirds of the follicle have degenerated, the dermal papillae come to rest at the base of the remaining permanent follicle segment, called the “bulge.” At the start of the next hair cycle, quiescent stem cells residing at the base of the bulge are stimulated to migrate and proliferate to supply the cells needed for hair-follicle regeneration and hair growth.
The surface of the eye is covered by the corneal epithelium, which acts as a protective outer barrier and helps to focus light into the retina. Like the epidermis, the cornea is a stratified epithelium. Adjacent to the corneal epithelium is the limbal region, a junctional zone between the cornea and the conjunctival epithelium. Corneal stem cells are thought to reside in this zone, giving rise to cells that migrate toward the center of the cornea, where they stratify and differentiate (Sun and Lavker, 2004).
Initially, mammary gland morphogenesis proceeds in a fashion that bears a marked resemblance to hair-follicle development. Ectodermal cells are instructed by their underlying mesenchyme to adopt a mammary epithelial fate and to grow downward as a germ. The germ then penetrates into the specialized breast mesenchyme, the fat pad, to form a rudimentary ductal tree that terminates into club-shaped structures, called terminal end buds (Hennighausen and Robinson, 2005). Mammary stem cells and their TA progeny reside in this structure along the basement membrane and have the capacity to form epithelial precursors that are committed to either a ductal or alveolar fate. During puberty, cells of terminal end buds proliferate rapidly to produce cells for the growth and branching of new ducts, which fill the entire fat pad. During pregnancy, alveoli are formed by the differentiation of alveoli precursors to form an external, loose network of myoepithelial cells. Differentiated alveoli epithelial cells then secrete milk into the lumen of the ducts during lactation. Contraction of the myoepithelial cells surrounding the ducts subsequently allows milk release. At the conclusion of lactation, alveoli structures undergo involution, and the mammary epithelium returns to its virgin appearance, ready for another cycle of alveolar growth and differentiation during the next pregnancy.
The location of stem cells within each of these epithelia was identified by taking advantage of the relative quiescence of stem cells within epithelial tissues. Despite their reduced mitotic index, epithelial stem cells can be labeled by continuous administration of nucleotide analogs (pulse) such as BrdU or 3H-TdR for a prolonged period. During the subsequent chase period, cells dilute their label through cell divisions. Those cells that divide less frequently during the chase period retain the label and hence have been referred to as label-retaining cells (Bickenbach, 1981). Label-retaining cells have been found in discrete locations within these epithelia. These sites of putative stem cells include the limbal region of the cornea (Cotsarelis et al., 1989), the bulge of the hair follicle (Cotsarelis et al., 1990), the terminal end bud of the mammary gland (Zeps et al., 1998), and a narrow band near the bottom of the intestinal crypts (Potten et al., 2002). To circumvent the use of BrdU or 3H-TdR in order to mark, isolate, and characterize living label-retaining cells, Tumbar and colleagues engineered transgenic mice expressing histone 2B-green fluorescent protein (H2B-GFP) under the control of a tetracycline-regulated enhancer element (Tumbar et al., 2004). To isolate label-retaining cells from stratified and glandular epithelia, they mated them to mice expressing the tetracycline-activatable repressor under control of a keratin 5 promoter (K5-tetoff) (Tumbar et al., 2004). In the absence of tetracycline, H2B-GFP is synthesized and incorporated into the nucleus of proliferating cells, but upon tetracycline addition, H2B-GFP expression is repressed. Assuming that chromosomally packaged H2B-GFP is very stable, GFP fluorescence intensity is likely to correlate with the number of cell divisions during the chase period. In mouse skin epithelium, a chase period of a month specifically marks bulge cells and has been used to isolate and characterize living bulge label-retaining cells. This method may be applicable to other epithelia, allowing the characterization and isolation of epithelial stem cells for which few or no cell-surface markers are currently known.
Epithelial-Mesenchymal Interactions Regulate Epithelial Fate
One defining characteristic of an epithelium is the close contact these cells have with the underlying mesenchyme. A number of classical tissue recombination experiments have demonstrated the inductive ability of mesenchymal tissues in the control of epithelial cell-fate determination. In the embryo, incubation of mesenchyme from different tissues can induce epithelia to adopt the cell fate of the mesenchymal tissue of origin. For instance, murine back-skin dermis can induce nonhairy epidermis to form hair follicles (Hardy, 1992). Similarly, epithelial lung buds are instructed to form gastric glands when placed in contact with stomach mesenchyme, villi epithelium when recombined with intestinal mesenchyme, and hepatic cords when incubated with liver mesenchyme (Birchmeier and Birchmeier, 1993). Prospective mammary mesenchyme can induce dorsal epidermis to differentiate into mammary epithelium (Cunha et al., 1995). Central corneal cells can be reprogrammed to form pilosebaceous units when transplanted with embryonic back-skin dermis (Ferraris et al., 2000). These experiments support the notion that epithelial appendage and tissue formation is instructed by mesenchymal signals. Whether the inductive potential is enhanced and maintained in adult stem cells is an intriguing question, as yet unexplored.
Additional epithelial-mesenchymal crosstalk is required to complete morphogenesis and differentiation. When combined with mouse, chicken, or lizard epidermis, murine dermis can induce hair follicle, feather, or scale placodes, respectively, but these placodes do not grow or differentiate further, due to the absence of dermal papillae. These results suggest that a second signal emanating from the epidermis is required for the differentiation of mesenchymal cells into a proper organizing center (Hardy, 1992). When taken together, these tissue recombination studies demonstrate that the reciprocal inductive interactions between the mesenchyme and the overlying epithelium are essential for the fate induction of various epithelia. Although the precise mediators of these epithelial-mesenchymal interactions are not well defined, secreted Wnt ligands may be one of the first signals involved in this communication as discussed later.
Multipotency of Epithelial Stem Cells
Epithelial homeostasis is typically maintained by unipo-tent progenitor cells, which have the ability to differentiate into one particular cell lineage. The direct demonstration of the existence of these progenitors and their lineage potential relies on clonal transplantation of isolated progenitors and genetic lineage tracing experiments.
The interfollicular epidermis and sebaceous glands contain unipotent progenitors that can maintain homeostasis of their respective tissue. Histological analyses of mouse skin reveals that the epidermis is organized in stacks of cells with a hexagonal surface area covering about ten basal cells (Potten, 1974). This structure has been proposed to function as an epidermal proliferative unit, harboring one putative stem cell per unit. More recently, this notion has been demonstrated experimentally by lineage tracing analysis, either by grafting virally tagged keratinocytes onto the backs of Nude mice (Ghazizadeh and Taichman, 2001; Kolodka et al., 1998; Mackenzie, 1997) or by inducing permanent expression of GFP clonally in the interfollicular epidermis and monitoring the fate of marked cells over time (Ro and Rannala, 2004). Stem cells from cultured human interfollicular epidermis have been isolated based on elevated expression of integrins (Jones and Watt, 1993). Recent fate-mapping analysis has also revealed the existence of a small group of unipo-tent sebaceous progenitor cells residing at the base of the gland that express the transcriptional repressor Blimp1 (Horsley et al., 2006).
In addition to these unipotent progenitors, multipotent stem cells reside in the bulge (Cotsarelis et al., 1990; Oshima et al., 2001; Taylor et al., 2000). Multipotent stem cells can differentiate into all epithelial cell lineages residing in the tissue. In skin, they not only serve as a reservoir of cells for regeneration during the normal cyclic periods of hair growth but also in conditions of hyperproliferation of sebaceous glands (Horsley et al., 2006) and in the repair of the interfollicular epidermis following wounding (Ito et al., 2005; Levy et al., 2005; Taylor et al., 2000; Tumbar et al., 2004) (Figure 2). Whether progenitors resident in the interfollicular epidermis or sebaceous glands can similarly become multipotent when the situation mandates is an intriguing question for future studies.
Figure 2. Multipotency of Hair-Follicle and Mammary Gland Stem Cells.
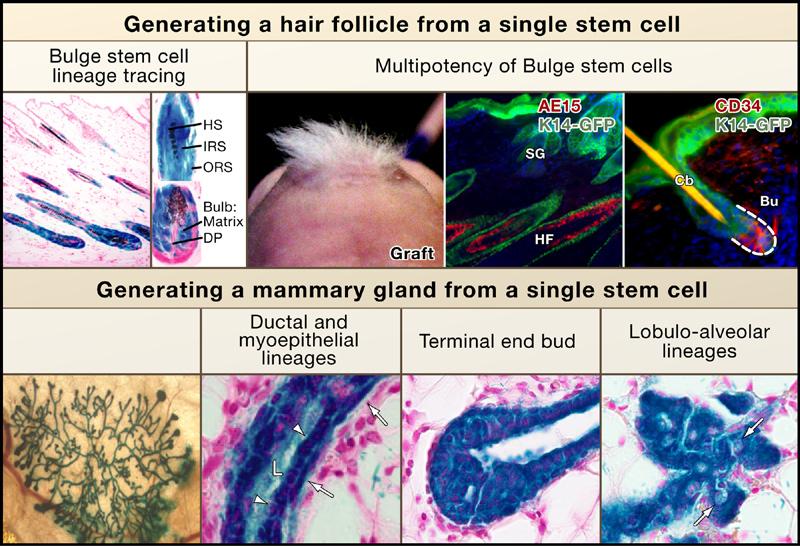
Bulge stem cell lineage tracing in vivo. Skin sections from mice expressing a regulatable Cre-recombinase under the control of a bulge-specific (K15) enhancer element and a lacZ transgene active in all Cre-expressing cells and their progeny (reprinted by permission from MacMillan Publishers Ltd.; Morris et al., 2004). Following Cre activation, all lineages of the new hair follicle expressed LacZ, demonstrating that they arose from bulge cells. Multipotency of bulge cells. CD34+, α6 integrin+ bulge cells were purified by flow cytometry from skin of mice expressing GFP-actin under the control of the K14 promoter (Blanpain et al., 2004). Bulge cells were cultured, and clonally derived keratinocytes from a single cell were used for grafting with unlabeled mesenchymal cells. Nude mice developed tufts of hair at grafted sites (left), and sectioning revealed that the GFP+ cells contributed to the epidermis, sebaceous glands (SG), and hair follicles (reprinted from Cell with permission from Elsevier; Blanpain et al., 2004). Multipotency of mammary gland stem cells. Lin−CD29hiCD24+ myoepithelial cells were purified by flow cytometry from lacZ+ mice, and individual cells were transplanted into cleared fat pads (Shackleton et al., 2006; Stingl et al., 2006) Epithelial grafts from virgin (left and middle) or pregnant recipients (right) demonstrate the ability of single lacZ+ stem cells (blue) to form a complete mammary gland structure. Sections of mammary grafts demonstrate the contribution of LacZ+ (blue) progeny from the single mammary stem cell to the ductal (arrowheads) and myoepithelial (arrows) cell lineages, the terminal end bud in a virgin recipient, and the lobulo-alveolar epithelium in a mid-term pregnant recipient (right, arrows indicate lipid droplets) (reprinted with permission from MacMillan Publishers Ltd: Shackleton et al., 2006).
The multipotency of stem cells within the bulge was first suggested from transplantation experiments in which a dissected bulge region was grafted onto immunodeficient mice (Oshima et al., 2001). Transplanted bulge cells were able to differentiate into the complete repertoire of skin epithelial cells: the interfollicular epidermis, sebaceous glands, and the eight cell lineages constituting mature hair follicles. Bulge cells purified by fluorescence-activated cell sorting (FACS) on the basis of their preferred expression of a GFP transgene also differentiated into these lineages upon transplantation (Morris et al., 2004). Clonal analyses were essential to demonstrate that bulge cells are multipotent stem cells and not simply a mixture of unipotent progenitors. This was accomplished by first culturing progeny derived from single FACS-purified bulge cells (Blanpain et al., 2004) or from microdissected tissue (Claudinot et al., 2005) and then transplantation.
The existence of multipotent stem cells in the mammary gland was posited from experiments in which fragments of mammary tissue containing cells harboring randomly inserted MMTV retroviruses were first transplanted into the epithelium-free mammary fat pads of an uninfected recipient mouse and then serially transplanting clonally derived outgrowths to create second-generation glands (Kordon and Smith, 1998). With each round of outgrowth, the viral integration site was mapped and shown to be identical, suggesting that an entire functional gland may arise from the progeny of a single mammary epithelial stem cell, which displays long-term self-renewal properties.
Two different groups recently demonstrated this rigorously, first by identifying and purifying a population of mammary cells (with the marker profile Lin−, α6 or β1High, and CD24+), which are enriched for stem cells, and then by transplantation studies (Shackleton et al., 2006; Stingl et al., 2006). By tagging these cells with a lacZ transgene, the researchers were able to directly demonstrate that individual mammary stem cells are able to generate a functional mammary gland upon transplantation (Figure 2). Similar to bulge stem cells, mammary stem cells display elevated surface integrins as well as keratins 5 and 14, which are characteristics of stratified and glandular epithelial cells that reside along a basement membrane and that have proliferative potential. They also appear to be slow-cycling cells, able to retain nucleotide label over an extended pulse-chase, and they express a number of genes that are also upregulated in bulge stem cells (Blanpain et al., 2004; Morris et al., 2004; Stingl et al., 2006; Tumbar et al., 2004).
Although the intestine is thought to have multipotent stem cells with the capacity to differentiate into enterocytes, goblet, Paneth, and neuroendocrine cells, the current biological evidence for the existence of a multipotent intestinal stem cell is not as well-established as it is for other epithelia. This is primarily because of the current paucity of cell-surface markers or promoters specific for intestinal stem cells that would permit their isolation from their putative location in the crypt . Recent molecular profiling of putative crypt stem cells captured by laser microdissection has revealed a number of new markers that could rapidly advance this field in the future (Giannakis et al., 2006).
In the absence of pure crypt cells to test their potential stem cell qualities, studies have been limited largely to lineage tracing analyses. In chimeric animals, a mosaic polyclonal composition of the intestinal crypts has been observed at birth, but within two weeks, crypts become mostly monoclonal, and thereafter the epithelium of individual crypts of adult mice is composed of cells of a single parental type (Ponder et al., 1985; Schmidt et al., 1988). The switch between a polyclonal crypt at birth and a monoclonal crypt after crypt fission in young animals is suggestive that several stem/progenitor cells reside within the initial crypts. In the adult intestine, however, only one stem/progenitor cell population appears to remain per crypt.
Additional genetic fate-mapping studies support the notion that unipotent and multipotent progenitors coexist in the intestinal epithelium (Bjerknes and Cheng, 1999). Although it seems likely that there are multipotent stem cells within intestinal epithelium, documentation of their existence awaits experiments that transplant isolated stem cells and/or genetic lineage tracing experiments that use promoters specific for intestinal stem cells.
Wnt Signaling and Epithelial Stem Cells
To ensure the lifelong maintenance of tissue homeostasis, stem cells must have an inexhaustible capacity to self-renew and differentiate. To do so, at least one daughter cell must maintain the properties of the stem cell following each cell division. Wnt/β-catenin signaling pathways have been shown to control the specification, maintenance, and activation of stem cells, and deregulation of this pathway often results in the development of familial and/or sporadic epithelial cancers (Reya and Clevers, 2005). Conserved from worms to human, the Wnt/β-catenin signaling pathway is evolutionarily ancient and orchestrates development and morphogenesis in many different tissues (Logan and Nusse, 2004).
Wnt proteins are secreted cysteine-rich proteins, of which there are about 20 members in mammals. The majority of β-catenin in the cell resides at sites of intercellular (adherens) junctions where it binds to E-cadherin at one end and α-catenin at the other. In contrast, the cytoplasmic pool of β-catenin is normally phosphorylated and targeted for ubiquitin-mediated degradation by a multi-protein Axin scaffold consisting of adenomatous polyposis coli (APC) protein, β-catenin, and its GSK-3β kinase. When Wnt ligand binds their cognate Frizzled receptor, the scaffold is unable to function, causing β-catenin to accumulate in the cytoplasm. When a member of the Tcf/Lef family of DNA-binding proteins is present, stabilized cytoplasmic β-catenin can bind to them and act as a transcriptional cofactor to stimulate the transcription of their target genes. Despite casual reference to β-catenin/ Tcf/Lef target gene activity as “Wnt reporter activity,” it is important to be mindful of the fact that β-catenin can be stabilized by non-Wnt mechanisms (He, 2006).
β-catenin signaling pathways often have multiple roles in stem cell lineage determination within most if not all epithelial tissues (Figure 3). In the intestine and mammary gland, Tcf4 is prominently expressed in the stem compartment (Barker et al., 1999); in the follicular stem cell niche, Tcf4 is expressed in conjunction with Tcf3 (Blanpain et al., 2004; Nguyen et al., 2006). The importance of Tcf/Lef1/β-catenin signaling in the specification and/or maintenance of epithelial stem cells has been highlighted by gain- and loss-of-function studies in mice. Mice deficient for Lef1 do not develop hair follicles, mammary glands, or teeth (van Genderen et al., 1994), whereas those lacking Tcf4 display a neonatal intestinal epithelium composed entirely of differentiated, nondividing villus cells (Korinek et al., 1998). Ablation of β-catenin also results in an absence of follicle morphogenesis in the neonate (Huelsken et al., 2001) and a postnatal loss of the follicle stem cell niche (Lowry et al., 2005). Similarly, overexpression of Dkk1, a soluble inhibitor of Wnt ligands, results in the loss of intestinal crypts (Pinto et al., 2003) and a failure to develop hair-follicle and mammary glands (Andl et al., 2002; Chu et al., 2004). Together, these studies underscore a specific role for Wnt signaling in the development and/or maintenance of epithelial stem cells.
Figure 3. Wnt/β-Catenin Signaling Regulates Epithelial Stem Cells.
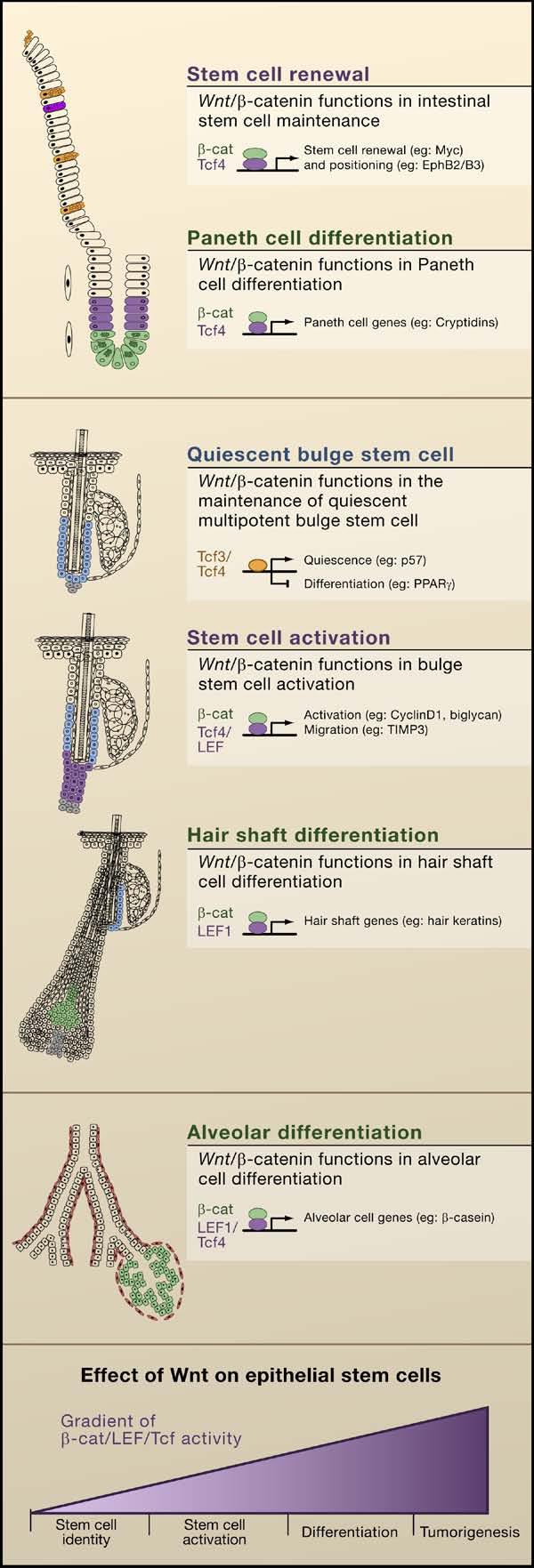
(A) Model summarizing the functions of Wnt/β-catenin signaling in the intestine that regulate stem cell renewal and Paneth cell differentiation. β-catenin and Tcf4 appear to regulate c-Myc and EphB2/B3 during renewal, whereas these transcriptional complexes regulate Paneth cell genes during differentiation. (B) Model summarizing functions that have been suggested for Wnt signaling in stem cell maintenance, activation, and hair shaft differentiation in the follicle. Tcf3/4 regulate genes implicated in stem cell quiescence and differentiation, whereas β-catenin and Tcf/Lef complexes regulate genes likely involved in stem cell activation and migration. β-catenin and Lef1 complexes in precortical cells regulate expression of hair shaft genes. (C) Model summarizing putative functions for Wnt signaling in the mammary gland in cancer development and alveolar differentiation. In combination with Tcf4 and Lef1, β-catenin regulates genes involved in alveolar differentiation. (D) General model depicting these different outcomes: stem cell self-renewal, activation, differentiation, or tumorigenesis, depending upon the level of effective Wnt signaling (that is, β-catenin/Lef/Tcf activity) that an epithelial stem cell receives.
The stem cells of the intestinal crypt appear to be in a constant Wnt-active proliferative state, and when a Tcf4 protein unable to bind β-catenin is expressed in colon cancer cells, it suppresses proliferation and promotes activation of differentiation genes (van de Wetering et al., 2002). By contrast, bulge stem cells undergo a resting phase, during which time they appear to reside in a Wnt restricted environment (DasGupta and Fuchs, 1999). During the transition between quiescent and growing phases of the hair cycle, nuclear β-catenin and Lef1/β-catenin reporter expression can be detected at the bulge base, where the new hair follicle begins to emerge (DasGupta and Fuchs, 1999; Lowry et al., 2005; Merrill et al., 2001). Consistent with the role of Wnt signaling in stem cell activation, transgenic mice expressing a stabilized form of β-catenin exhibit de novo formation of hair follicles and increased follicle density (Gat et al., 1998) as well as precocious re-entry into the regenerative phase of hair growth (Lo Celso et al., 2004; Lowry et al., 2005; Van Mater et al., 2003). Similarly in the mammary gland, precocious bud formation occurs with targeted activation of β-catenin (Teuliere et al., 2005).
Some Tcfs may function in stem cells even when the levels of cytoplasmic/nuclear β-catenin are low and/or absent. It was recently found that Tcf3 is expressed in adult follicle stem cells at all stages of the hair cycle as well as in multipotent embryonic skin progenitors. Moreover, when Tcf3 expression is maintained postnatally, all three skin differentiation lineages are repressed (Nguyen et al., 2006), suggesting that Tcf3 may function to maintain skin stem cells in an undifferentiated state, and when β-catenin is stabilized, Tcf3 repression is relieved, and stem cells become activated to proliferate and embark upon hair-follicle lineage differentiation.
Given a role for Tcf/β-catenin in stem cell activation, it is perhaps not surprising to find that constitutively stabilizing mutations in β-catenin are associated with a variety of human epithelial cancers. Such mutations were first discovered in adenocarcinomas of the colon (Reya and Clevers, 2005) and have since been identified in pilomatricomas, which are tumors of hair shaft cells (Chan et al., 1999; Gat et al., 1998), and in mammary gland adenocarcinomas (Teuliere et al., 2005; Tsukamoto et al., 1988). An interesting feature of epithelial tumors generated by excessive Wnt signaling is that they often display an increased frequency of cells with stem and progenitor properties, in contrast to tumors from mice expressing other oncogenes. In this regard, the Wnt pathway appears to be special if not unique in its ability to target stem cell and/or progenitor cells for transformation.
Another feature of cancers arising from excessive Wnt signaling is that they often selectively display the features of one specific stem cell lineage. This is best explained by the fact that additional Wnt signals are involved at later steps in the differentiation of these select stem cell lineages, e.g., the hair shaft (Merrill et al., 2001), the intestinal Paneth cells (van Es et al., 2005a), and lobulo-alveolar differentiation in mammary epithelium (Teuliere et al., 2005; Tsukamoto et al., 1988). Notably, hair keratin genes are transcriptionally regulated by Lef1/β-catenin (Merrill et al., 2001) and expression of genes specific to Paneth cells, such as cryptidin/defensins, is lost upon deletion of Tcf4 or Wnt receptor Frizzled 5 (van Es et al., 2005a).
To understand how β-catenin/Tcf signaling controls epithelial stem cells, researchers have focused on identifying genes that are regulated upon Wnt signaling. In recent years, transcriptional profiling has been used to determine the genes downstream of β-catenin signaling in epithelial stem cells (Batlle et al., 2002; Lowry et al., 2005; Nguyen et al., 2006; Silva-Vargas et al., 2005; van de Wetering et al., 2002). Among the interesting new Wnt targets that have surfaced are ephrin receptors EphB2 and EphB3, which are expressed in the intestinal crypts in response to Wnt signaling (Batlle et al., 2002). By contrast, the EphrinB1 ligand is expressed in the cells above the crypts. Compound mutations of EphB2 and EphB3 result in the mispositioning of cells along the crypt-villus axis, suggesting that ephrins and their ligands establish a boundary between proliferative and differentiating compartments. In addition to participating in the proper positioning of the cells within the intestine, EphBs seem to regulate cell proliferation within the crypts (Holmberg et al., 2006), and EphB2 expression is lost during progression from a benign adenoma to aggressive carcinoma (Batlle et al., 2005). Ephrins and their receptors exhibit differential expression patterns in other epithelia, but their significance awaits further investigation.
In the hair follicle, the stabilization of β-catenin stimulates resting bulge stem cells to proliferate and regenerate hair follicles. Through transcriptional profiling of purified bulge stem cells during resting and active phases of the hair cycle, it was discovered that during stem cell activation a number of genes associated with cell-cycle progression are transcribed (Lowry et al., 2005). When the levels of β-catenin are transgenically elevated in quiescent stem cells, the transcription of some of these genes is elevated, providing insights into how Wnt signaling may promote stem cell activation. Notably, Wnt signaling in bulge stem cells did not upregulate the hair keratin genes that are induced at later stages of hair-follicle differentiation (Lowry et al., 2005). Thus as stem cells are activated and progress along a particular differentiation lineage, they appear to respond to different environmental cues and activate different sets of Wnt target genes. The multiplicity of transcriptional partners for β-catenin, coupled with the complex regulation of its stability, are likely to contribute to the effect and potency of β-catenin/Lef1/Tcf activity in cells. These regulatory mechanisms of Wnt signaling are likely to impact on whether and when the stem cells will self-renew, become activated to proliferate, or embark upon a terminal differentiation program.
Notch Signaling and Epithelial Stem Cells
Tissue homeostasis and wound repair requires epithelial stem cells to execute different programs of terminal differentiation. To achieve these feats, stem cells must be instructed by their microenvironment to selectively adopt a particular cell lineage. Wnt signaling is not the only signal transduction pathway that instructs stem cells. Like Wnt signaling, Notch signaling controls selective cell-fate determination in variety of different tissues (Artavanis-Tsakonas et al., 1999). Conserved throughout the eukaryotic kingdom, the canonical Notch signaling pathway regulates cell-fate decisions through close-range, cell-cell interactions.
In mammals, there are four Notch transmembrane receptors (Notch1–4). Notch ligands are also transmembrane proteins comprising three different sub-families, each containing several members. Upon ligand engagement, Notch receptors are sequentially cleaved by metalloproteinases and γ-secretases (presenillins 1 and 2). Once released, Notch's intracellular domain (NICD) is free to translocate to the nucleus, where it can associate with the DNA-binding protein RBP-J to convert it into a transcriptional activator. The best characterized Notch target genes belong to the hairy enhancer of split (Hes) complex and consist of the b-HLH transcription factors Hes (1–7) and Hey (1–3) (Iso et al., 2003). In the intestine, Notch signaling regulates cell-fate determination in the crypt, where Notch1, several Notch ligands, and Hes1, 6, and 7 are preferentially expressed (Schroder and Gossler, 2002). The first evidence that Notch signaling controls stem cell-fate determination within the intestinal epithelium came from the analysis of the knockout mice for Hes1 and one of its repressed target genes, Math1 (Figure 4). Mice deficient for the Hes1 gene possess an increase of neuroendocrine cells, Paneth cells, and goblet cells at the expense of intestinal enterocytes (Jensen et al., 2000; Suzuki et al., 2005) whereas the knockout of Math1 or gain-of-function mutations in Notch leads to the opposite phenotype (Fre et al., 2005; Stanger et al., 2005; Yang et al., 2001). Additionally, inhibition of γ-secretase (Searfoss et al., 2003), or conditional targeting of RBP-J (van Es et al., 2005b), results in a dramatic expansion of the goblet cell population within the mouse intestine. Notch signaling can also control TA cell proliferation. Crypt cell proliferation is grossly impaired upon RBPJ ablation or following γ-secretase inhibition (van Es et al., 2005b), whereas proliferation is enhanced by overactivation of Notch signaling (Fre et al., 2005). Remarkably, by chemically blocking γ-secretase, intestinal tumors in APC mutant mice differentiate into goblet cells, illuminating the potential of using chemical inhibitors of Notch signaling as possible therapeutic tools in the treatment of intestinal cancers (van Es et al., 2005b). Together, these results suggest a model in which Notch signaling acts on intestinal stem cells both to maintain their proliferative progenitor state and also to restrict the fate of neuroendocrine, goblet, and Paneth cells.
Figure 4. Notch Signaling Regulates Lineages Determination of Epithelial Stem Cells.
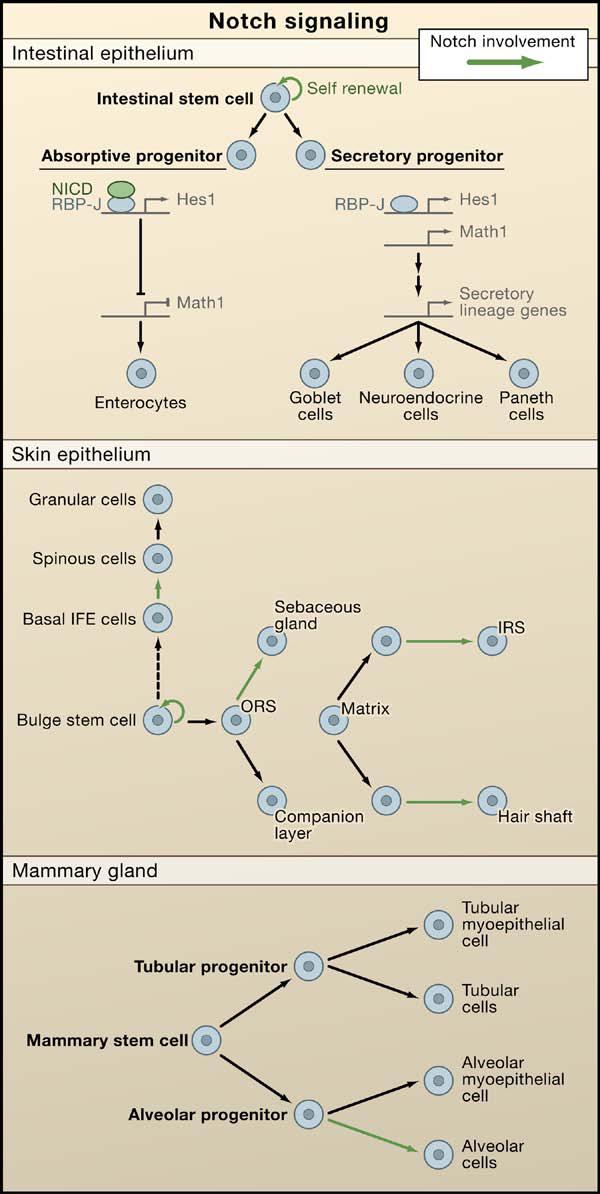
In canonical Notch signaling, upon binding to Notch ligands, Notch receptors are cleaved and the intracellular domain translocates to the nucleus to act in concert with an RBP-J transcription factor to regulate gene expression. The schematics illustrate the putative roles of Notch signaling in three different epithelial tissues. (Top) A role for Notch in intestinal stem cell self-renewal and differentiation. Notch/RBP-J restricts the fate of secretory lineages in the intestine (Goblet, Paneth, and enteroendocrine cells). In the absorptive progenitors, Notch activity leads to the expression of Hes1 and subsequent repression of Math1, leading to enterocyte differentiation. In the absence of Notch signaling, Math1 expression is not repressed and allows secretory progenitors to differentiate into goblet, neuroendrocrine, and Paneth cells. (Middle) A role for Notch in stem cell maintenance and differentiation in the epidermis and hair follicle. Notch promotes spinous cell fate in the interfollicular epidermis (IFE) and regulates differentiation of the sebaceous gland, the hair shaft, and the inner root sheath cells of the hair follicle. (Bottom) A role for Notch signaling in alveolar differentiation in the mammary gland. Mammary stem cells form alveolar and ductal progenitors that each form a myoepithelial cell in addition to tubular and alveolar cells, respectively. Notch signaling regulates the differentiation of alveolar progenitors into alveolar cells.
A similar role for Notch signaling in promoting proliferation and differentiation was also found in mammary epithelium. Overexpression of the active form of Notch4 leads to mammary tumorigenesis (Hu et al., 2006), and although ablation of RBP-J does not disturb initial mammary gland morphogenesis, alveolar differentiation is severely impaired during pregnancy (Buono et al., 2006) (Figure 4). Thus, it came as a surprise when Notch was implicated as a tumor suppressor in the cornea and epidermis, as judged by the fact that loss of Notch1 in adult epidermis and cornea leads to epithelial hyperthickening accompanied by expansion of basal layers (Rangarajan et al., 2001). Additional support came from studies showing that Notch1-deficient skin displays a higher susceptibility of developing chemically induced cancer (Nicolas et al., 2003).
At first glance, it might appear as though Notch signaling promotes maintenance and self-renewal in some epithelial stem cells while inhibiting these processes in others. However, multiple Notch receptors are expressed in the skin, and when all canonical Notch signaling is ablated in embryonic epidermis, proliferation is reduced and differentiation is impaired (Blanpain et al., 2006). Whether the hyperproliferation seen upon Notch1 ablation arises from alterations in noncanonical Notch signaling or as an indirect consequence of epidermal barrier defects is not yet clear. However, in the absence of RBP-J, follicle stem cells cannot be maintained, drawing parallels in the action of canonical Notch signaling on epithelial stem cells (Blanpain et al., 2006; Yamamoto et al., 2003).
Like Wnt signaling, Notch signaling appears to exert its effects at multiple steps along lineage pathways. For example, in the hair follicle, NICD1 is also expressed in the matrix cells of both the hair shaft and its channel or inner root sheath (Pan et al., 2004), and conditional ablation of γ-secretase, or combined with the loss of either Notch receptor genes or RBPJ, results in defects in sebaceous gland development and terminal differentiation of the inner root sheath and hair shaft (Blanpain et al., 2006; Pan et al., 2004; Vauclair et al., 2005; Yamamoto et al., 2003) (Figure 4). Taken together, these studies reveal multiple roles for Notch signaling not only in stem cells but also later, in their lineages.
Other Signaling Pathways and Stem Cell Function
Although Wnt and Notch signaling cascades have been the most extensively studied in epithelial stem cells, Bone Morphogenetic Protein (BMP) signaling appears to influence the activation of multiple different types of epithelial stem cells. Thus, inhibition of BMP signaling by overexpression of Noggin results in induction of hair placode formation as well as de novo formation of the intestinal crypt (Botchkarev et al., 2001; Haramis et al., 2004; He et al., 2004). Mutations that affect BMP signaling account for almost half of the cases of juvenile polyposis syndrome, a condition that can lead to intestinal cancer (Sancho et al., 2004). Similarly, conditional ablation of the Bmpr1a gene, encoding a BMP receptor protein, results in the continued activation of hair-follicle stem cells and the eventual formation of follicular tumors (Andl et al., 2004). BMP signaling is also required for proper stem cell differentiation, as targeted ablation of Bmpr1a results in the accretion of undifferentiated hair-follicle and intestinal epithelial cells (Andl et al., 2004; Kobielak et al., 2003; Sancho et al., 2004).
The roles of other signaling pathways in epithelial stem cell biology are less clear. Embryos lacking Sonic hedgehog (Shh), for instance, display a severely impaired hair-follicle morphogenesis, whereas the development of the intestine and mammary gland are relatively normal. Conversely, activating mutations of Shh signaling or inactivating mutations of its negative regulator (Patched) are found in familial and sporadic basal cell carcinomas, a frequent skin cancer, and in several other types of epithelial cancers including pancreatic carcinoma (Beachy et al., 2004). Unexpectedly, reducing hedgehog signaling even seems to enhance cell proliferation in neonatal and adult intestine (Madison et al., 2005; van den Brink et al., 2004). Together, these data suggest that hedgehog signaling may promote proliferation in some epithelial cells types whereas in other epithelial cells it inhibits proliferation or has no effect.
Epithelial Stem Cells and Regenerative Medicine
Pioneering studies by H. Green and coworkers led to successful culturing of human epidermal keratinocytes, thereby opening the door for subsequent clinical applications of stratified epithelial stem cells in regenerative medicine (Rheinwald and Green, 1975; Figure 5). Not all cells from freshly isolated human epidermis displayed a fountain of youth potential, but some keratinocytes generated large colonies (“holoclones”), which could be passaged for several hundred generations (Barrandon and Green, 1987). Green and colleagues realized the clinical potential of their findings and applied them to the successful treatment of burn patients (O'Connor et al., 1981). The use of fibrin matrices has since accelerated keratinocyte growth and improved long-term skin engraftments (Pellegrini et al., 1999; Ronfard et al., 2000; Figure 5). These procedures now save the lives of severe burn patients throughout the world. The methods also hold promise when combined with ex vivo gene therapies to correct a variety of life-threatening genetic skin disorders, including blistering conditions (Mavilio et al., 2006). The recent discovery of methods to isolate and purify cells of the dermal papillae (Rendl et al., 2005; Richardson et al., 2005) offers the prospect for coaxing bulge stem cells to regenerate hair follicles in a clinical setting.
Figure 5. Regenerative Potential of Epidermal and Corneal Epithelium.
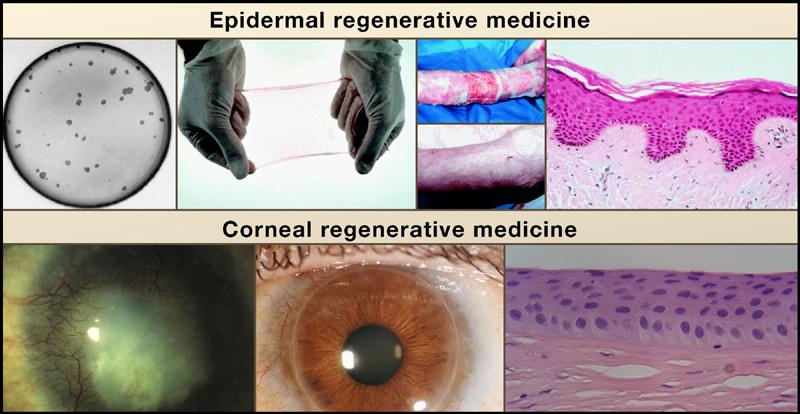
Top, from left to right: Colonies of human keratinocytes grown on a fibrin matrix can be used to form an epithelial sheet. These sheets were used to treat a patient with burns on over 95% of his body (top image). After 3.5 years, the transplantation has completely healed the patient's skin (bottom image). Histologically, the long-term transplant was normal and contained all of the normal epidermal layers and architecture (reprinted with permission from Lippincott, Williams, and Wilkins; Ronfard et al., 2000). Bottom, from left to right: Long-term potential of the cornea from patient with alkali burns to the cornea causing stromal scarring and neovascularization (Pellegrini, et al., 1997). Cultures of limbal stem cells isolated from a biopsy of the contralateral eye were transplanted and were able to repair the damaged cornea and produce a healthy cornea, allowing 100% recovery of visual acuity. A biopsy from the regenerated cornea demonstrated that the tissue organization was normal (Courtesy of Drs. Graziella Pellegrini, Paolo Rama, and Michelle DeLuca).
Although the epidermis has been the front-runner in the epithelial arena of regenerative medicine, the importance of developing applications to generate corneal cells for treating certain forms of blindness has garnered the focus of clinical researchers. The permanent loss of corneal epithelium results in an atypical re-epithelialization by the bulbar conjunctival cells, and this in turn stimulates neovascularization and inflammation. The limbus is the putative corneal stem cell niche, and limbal transplantations have been used to successfully restore corneal epithelium and eyesight (Kenyon and Tseng, 1989). More recently, cases of eye damage have been successfully treated by first culturing limbal stem cells and then transplanting the autologous corneal epithelial sheets (Pellegrini et al., 1997). The successes achieved in epidermal and corneal regeneration using cultured epithelial stem cells serve as graphic illustrations of how exciting advances in the laboratory can be translated into useful therapies. These results are also encouraging for the development of new methods for the repair or regeneration of other epithelia, damaged through injury or disease.
One intriguing issue still unaddressed is the extent to which a multipotent stem cell within an epithelium can be used to generate other epithelial tissues when exposed to different mesenchymes. Embryonic epithelial cells have this capacity—what about adult epithelial stem cells? If it is possible to coax a bulge stem cell to make corneal epithelium when exposed to corneal mesenchyme rather than dermal papillae, this would greatly expand the clinical applications of this readily accessible source of adult stem cells.
Conclusions
In this Review, we have limited our focus to comparative features of epithelial stem cells and what we might gain from such comparisons. There are many other signaling pathways and programs of gene expression that have been elegantly demonstrated to impact on the behavior of specific types of epithelial stem cells. There is a strong likelihood that as we learn more about epithelial stem cells, additional parallels will emerge. This seems especially likely in view of increasing evidence that Wnt and Notch signaling are likely to integrate with other signals such as BMP, Shh, and growth factors emanating from stem cells, their immediate progeny, and their microenvironment. Taken together, these signals converge to generate the distinct features of stem cells, including self-renewal, proliferation and survival, and suppression of cell-fate commitment and lineage determination.
There are still only a few cases where multipotent epithelial stem cells have been isolated and purified. With the recent isolation of bulge and mammary epithelial stem cells, this should set the stage for rapid advances in the future. Both bulge and mammary epithelial stem cells exist as relatively slow-cycling progenitors, both are positive for the markers keratin 14 and 5, and both express high levels of integrins. Given the existing methods to isolate cells from transgenic mice on the basis of their ability to express keratin 5 and integrins and their infrequent cycling (Tumbar et al., 2004), it should be possible to isolate stem cells from a variety of epithelia including lung, reproductive tracts, tongue, cornea, esophagus, and sweat glands. Although internal epithelia express keratins 8 and 18 instead of 5 and 14 and undergo turnover more rapidly than surface and glandular epithelia, methods to purify these cells are likely to be forthcoming in the near future.
As characteristics of surface, glandular, and internal epithelial stem cells emerge, it will be interesting to ascertain what features they will share that might account for their common properties of self-renewal and repression of differentiation fates. It will also be exciting to unearth the features that uniquely define them, such as their different degrees of quiescence and long-term survival or their unipotency versus multipotency. What is the impact of an epithelial stem cell's native niche on its behavior? Which traits are cell autonomous? How similar are corneal, epidermal, and esophageal stem cells? To what extent do eccrine sweat and salivary gland stem cells resemble those of a mammary gland? What prompts them to secrete the tissue-specific products, sweat, saliva, or milk? Defining the mechanisms that govern epithelial stem cell behavior is not only a rapidly progressing field of science but is also an increasingly important biomedical terrain for devising new and improved treatments of damaged and/or cancerous epithelial tissues. Biologists, clinicians, and patients will watch with interest as the mysteries underlying the fascinating properties of epithelial stem cells continue to emerge.
ACKNOWLEDGMENTS
We thank Y. Barrandon, H. Clevers, G. Cotsarelis, M. De Luca, H. Varmus, and J. Visvader for providing images. C.B. is supported by HFSPO and FNRS, V.H. is a Robert Black Fellow of the Damon Runyon Cancer Research Foundation (DRG-1802-04), and E.F. is an HHMI investigator and receives funding from the NIH and the Stem Cell Research Foundation.
REFERENCES
- Andl T, Ahn K, Kairo A, Chu EY, Wine-Lee L, Reddy ST, Croft NJ, Cebra-Thomas JA, Metzger D, Chambon P, et al. Epithelial Bmpr1a regulates differentiation and proliferation in postnatal hair follicles and is essential for tooth development. Development. 2004;131:2257–2268. doi: 10.1242/dev.01125. [DOI] [PubMed] [Google Scholar]
- Andl T, Reddy ST, Gaddapara T, Millar SE. WNT signals are required for the initiation of hair follicle development. Dev. Cell. 2002;2:643–653. doi: 10.1016/s1534-5807(02)00167-3. [DOI] [PubMed] [Google Scholar]
- Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch signaling: cell fate control and signal integration in development. Science. 1999;284:770–776. doi: 10.1126/science.284.5415.770. [DOI] [PubMed] [Google Scholar]
- Barker N, Huls G, Korinek V, Clevers H. Restricted high level expression of Tcf-4 protein in intestinal and mammary gland epithelium. Am. J. Pathol. 1999;154:29–35. doi: 10.1016/S0002-9440(10)65247-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrandon Y, Green H. Three clonal types of keratinocyte with different capacities for multiplication. Proc. Natl. Acad. Sci. USA. 1987;84:2302–2306. doi: 10.1073/pnas.84.8.2302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Batlle E, Henderson JT, Beghtel H, van den Born MM, Sancho E, Huls G, Meeldijk J, Robertson J, van de Wetering M, Pawson T, Clevers H. Beta-catenin and TCF mediate cell positioning in the intestinal epithelium by controlling the expression of EphB/ephrinB. Cell. 2002;111:251–263. doi: 10.1016/s0092-8674(02)01015-2. [DOI] [PubMed] [Google Scholar]
- Batlle E, Bacani J, Begthel H, Jonkheer S, Gregorieff A, van de Born M, Malats N, Sancho E, Boon E, Pawson T, et al. EphB receptor activity suppresses colorectal cancer progression. Nature. 2005;435:1126–1130. doi: 10.1038/nature03626. [DOI] [PubMed] [Google Scholar]
- Beachy PA, Karhadkar SS, Berman DM. Tissue repair and stem cell renewal in carcinogenesis. Nature. 2004;432:324–331. doi: 10.1038/nature03100. [DOI] [PubMed] [Google Scholar]
- Bickenbach JR. Identification and behavior of label-retaining cells in oral mucosa and skin. J. Dent. Res. 1981;60 Spec No C:1611–1620. doi: 10.1177/002203458106000311011. [DOI] [PubMed] [Google Scholar]
- Birchmeier C, Birchmeier W. Molecular aspects of mesenchymal-epithelial interactions. Annu. Rev. Cell Biol. 1993;9:511–540. doi: 10.1146/annurev.cb.09.110193.002455. [DOI] [PubMed] [Google Scholar]
- Bjerknes M, Cheng H. Clonal analysis of mouse intestinal epithelial progenitors. Gastroenterology. 1999;116:7–14. doi: 10.1016/s0016-5085(99)70222-2. [DOI] [PubMed] [Google Scholar]
- Blanpain C, Fuchs E. Epidermal stem cells of the skin. Annu. Rev. Cell Dev. Biol. 2006;22:339–373. doi: 10.1146/annurev.cellbio.22.010305.104357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanpain C, Lowry WE, Geoghegan A, Polak L, Fuchs E. Self-renewal, multipotency, and the existence of two cell populations within an epithelial stem cell niche. Cell. 2004;118:635–648. doi: 10.1016/j.cell.2004.08.012. [DOI] [PubMed] [Google Scholar]
- Blanpain C, Lowry WE, Pasolli HA, Fuchs E. Canonical notch signaling functions as a commitment switch in the epidermal lineage. Genes Dev. 2006;20:3022–3035. doi: 10.1101/gad.1477606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Botchkarev VA, Botchkareva NV, Nakamura M, Huber O, Funa K, Lauster R, Paus R, Gilchrest BA. Noggin is required for induction of the hair follicle growth phase in postnatal skin. FASEB J. 2001;15:2205–2214. doi: 10.1096/fj.01-0207com. [DOI] [PubMed] [Google Scholar]
- Buono KD, Robinson GW, Martin C, Shi S, Stanley P, Tanigaki K, Honjo T, Hennighausen L. The canonical Notch/RBPJ signaling pathway controls the balance of cell lineages in mammary epithelium during pregnancy. Dev. Biol. 2006;293:565–580. doi: 10.1016/j.ydbio.2006.02.043. [DOI] [PubMed] [Google Scholar]
- Chan EF, Gat U, McNiff JM, Fuchs E. A common human skin tumour is caused by activating mutations in beta-catenin. Nat. Genet. 1999;21:410–413. doi: 10.1038/7747. [DOI] [PubMed] [Google Scholar]
- Chu EY, Hens J, Andl T, Kairo A, Yamaguchi TP, Brisken C, Glick A, Wysolmerski JJ, Millar SE. Canonical WNT signaling promotes mammary placode development and is essential for initiation of mammary gland morphogenesis. Development. 2004;131:4819–4829. doi: 10.1242/dev.01347. [DOI] [PubMed] [Google Scholar]
- Claudinot S, Nicolas M, Oshima H, Rochat A, Barrandon Y. Long-term renewal of hair follicles from clonogenic multipotent stem cells. Proc. Natl. Acad. Sci. USA. 2005;102:14677–14682. doi: 10.1073/pnas.0507250102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cotsarelis G, Cheng SZ, Dong G, Sun TT, Lavker RM. Existence of slow-cycling limbal epithelial basal cells that can be preferentially stimulated to proliferate: implications on epithelial stem cells. Cell. 1989;57:201–209. doi: 10.1016/0092-8674(89)90958-6. [DOI] [PubMed] [Google Scholar]
- Cotsarelis G, Sun TT, Lavker RM. Label-retaining cells reside in the bulge area of pilosebaceous unit: implications for follicular stem cells, hair cycle, and skin carcinogenesis. Cell. 1990;61:1329–1337. doi: 10.1016/0092-8674(90)90696-c. [DOI] [PubMed] [Google Scholar]
- Cunha GR, Young P, Christov K, Guzman R, Nandi S, Talamantes F, Thordarson G. Mammary phenotypic expression induced in epidermal cells by embryonic mammary mesenchyme. Acta Anat. (Basel) 1995;152:195–204. doi: 10.1159/000147698. [DOI] [PubMed] [Google Scholar]
- Dasgupta R, Fuchs E. Multiple roles for activated LEF/TCF transcription complexes during hair follicle development and differentiation. Development. 1999;126:4557–4568. doi: 10.1242/dev.126.20.4557. [DOI] [PubMed] [Google Scholar]
- Dor Y, Brown J, Martinez OI, Melton DA. Adult pancreatic beta-cells are formed by self-duplication rather than stem-cell differentiation. Nature. 2004;429:41–46. doi: 10.1038/nature02520. [DOI] [PubMed] [Google Scholar]
- Ferraris C, Chevalier G, Favier B, Jahoda CA, Dhouailly D. Adult corneal epithelium basal cells possess the capacity to activate epidermal, pilosebaceous and sweat gland genetic programs in response to embryonic dermal stimuli. Development. 2000;127:5487–5495. doi: 10.1242/dev.127.24.5487. [DOI] [PubMed] [Google Scholar]
- Fre S, Huyghe M, Mourikis P, Robine S, Louvard D, Artavanis-Tsakonas S. Notch signals control the fate of immature progenitor cells in the intestine. Nature. 2005;435:964–968. doi: 10.1038/nature03589. [DOI] [PubMed] [Google Scholar]
- Fuchs E, Raghavan S. Getting under the skin of epidermal morphogenesis. Nat. Rev. Genet. 2002;3:199–209. doi: 10.1038/nrg758. [DOI] [PubMed] [Google Scholar]
- Gat U, Dasgupta R, Degenstein L, Fuchs E. De Novo hair follicle morphogenesis and hair tumors in mice expressing a truncated beta-catenin in skin. Cell. 1998;95:605–614. doi: 10.1016/s0092-8674(00)81631-1. [DOI] [PubMed] [Google Scholar]
- Ghazizadeh S, Taichman LB. Multiple classes of stem cells in cutaneous epithelium: a lineage analysis of adult mouse skin. EMBO J. 2001;20:1215–1222. doi: 10.1093/emboj/20.6.1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giancotti FG, Tarone G. Positional control of cell fate through joint integrin/receptor protein kinase signaling. Annu. Rev. Cell Dev. Biol. 2003;19:173–206. doi: 10.1146/annurev.cellbio.19.031103.133334. [DOI] [PubMed] [Google Scholar]
- Giannakis M, Stappenbeck TS, Mills JC, Leip DG, Lovett M, Clifton SW, Ippolito JE, Glasscock JI, Arumugam M, Brent MR, Gordon JI. Molecular properties of adult mouse gastric and intestinal epithelial progenitors in their niches. J. Biol. Chem. 2006;281:11292–11300. doi: 10.1074/jbc.M512118200. [DOI] [PubMed] [Google Scholar]
- Haramis AP, Begthel H, van den Born M, van Es J, Jonkheer S, Offerhaus GJ, Clevers H. De novo crypt formation and juvenile polyposis on BMP inhibition in mouse intestine. Science. 2004;303:1684–1686. doi: 10.1126/science.1093587. [DOI] [PubMed] [Google Scholar]
- Hardy MH. The secret life of the hair follicle. Trends Genet. 1992;8:55–61. doi: 10.1016/0168-9525(92)90350-d. [DOI] [PubMed] [Google Scholar]
- He X. Unwinding a path to nuclear beta-catenin. Cell. 2006;127:139–155. doi: 10.1016/j.cell.2006.09.016. [DOI] [PubMed] [Google Scholar]
- He XC, Zhang J, Tong WG, Tawfik O, Ross J, Scoville DH, Tian Q, Zeng X, He X, Wiedemann LM, et al. BMP signaling inhibits intestinal stem cell self-renewal through suppression of Wnt-beta-catenin signaling. Nat. Genet. 2004;36:1117–1121. doi: 10.1038/ng1430. [DOI] [PubMed] [Google Scholar]
- Hennighausen L, Robinson GW. Information networks in the mammary gland. Nat. Rev. Mol. Cell Biol. 2005;6:715–725. doi: 10.1038/nrm1714. [DOI] [PubMed] [Google Scholar]
- Holmberg J, Genander M, Halford MM, Anneren C, Sondell M, Chumley MJ, Silvany RE, Henkemeyer M, Frisen J. EphB receptors coordinate migration and proliferation in the intestinal stem cell niche. Cell. 2006;125:1151–1163. doi: 10.1016/j.cell.2006.04.030. [DOI] [PubMed] [Google Scholar]
- Horsley V, O'Carroll D, Tooze R, Ohinata Y, Saitou M, Obukhanych T, Nussenzweig M, Tarakhovsky A, Fuchs E. Blimp1 defines a progenitor population that governs cellular input to the sebaceous gland. Cell. 2006;126:597–609. doi: 10.1016/j.cell.2006.06.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu C, Dievart A, Lupien M, Calvo E, Tremblay G, Jolicoeur P. Overexpression of activated murine Notch1 and Notch3 in transgenic mice blocks mammary gland development and induces mammary tumors. Am. J. Pathol. 2006;168:973–990. doi: 10.2353/ajpath.2006.050416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huelsken J, Vogel R, Erdmann B, Cotsarelis G, Birchmeier W. beta-Catenin controls hair follicle morphogenesis and stem cell differentiation in the skin. Cell. 2001;105:533–545. doi: 10.1016/s0092-8674(01)00336-1. [DOI] [PubMed] [Google Scholar]
- Iso T, Kedes L, Hamamori Y. HES and HERP families: multiple effectors of the Notch signaling pathway. J. Cell. Physiol. 2003;194:237–255. doi: 10.1002/jcp.10208. [DOI] [PubMed] [Google Scholar]
- Ito M, Liu Y, Yang Z, Nguyen J, Liang F, Morris RJ, Cotsarelis G. Stem cells in the hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis. Nat. Med. 2005;11:1351–1354. doi: 10.1038/nm1328. [DOI] [PubMed] [Google Scholar]
- Jensen J, Pedersen EE, Galante P, Hald J, Heller RS, Ishibashi M, Kageyama R, Guillemot F, Serup P, Madsen OD. Control of endodermal endocrine development by Hes-1. Nat. Genet. 2000;24:36–44. doi: 10.1038/71657. [DOI] [PubMed] [Google Scholar]
- Jones PH, Watt FM. Separation of human epidermal stem cells from transit amplifying cells on the basis of differences in integrin function and expression. Cell. 1993;73:713–724. doi: 10.1016/0092-8674(93)90251-k. [DOI] [PubMed] [Google Scholar]
- Kenyon KR, Tseng SC. Limbal autograft transplantation for ocular surface disorders. Ophthalmology. 1989;96:709–722. doi: 10.1016/s0161-6420(89)32833-8. [DOI] [PubMed] [Google Scholar]
- Kobielak K, Pasolli HA, Alonso L, Polak L, Fuchs E. Defining BMP functions in the hair follicle by conditional ablation of BMP receptor IA. J. Cell Biol. 2003;163:609–623. doi: 10.1083/jcb.200309042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolodka TM, Garlick JA, Taichman LB. Evidence for keratinocyte stem cells in vitro: long term engraftment and persistence of transgene expression from retrovirus-transduced keratinocytes. Proc. Natl. Acad. Sci. USA. 1998;95:4356–4361. doi: 10.1073/pnas.95.8.4356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kordon EC, Smith GH. An entire functional mammary gland may comprise the progeny from a single cell. Development. 1998;125:1921–1930. doi: 10.1242/dev.125.10.1921. [DOI] [PubMed] [Google Scholar]
- Korinek V, Barker N, Moerer P, van Donselaar E, Huls G, Peters PJ, Clevers H. Depletion of epithelial stem-cell compartments in the small intestine of mice lacking Tcf-4. Nat. Genet. 1998;19:379–383. doi: 10.1038/1270. [DOI] [PubMed] [Google Scholar]
- Levy V, Lindon C, Harfe BD, Morgan BA. Distinct stem cell populations regenerate the follicle and interfollicular epidermis. Dev. Cell. 2005;9:855–861. doi: 10.1016/j.devcel.2005.11.003. [DOI] [PubMed] [Google Scholar]
- Lo Celso C, Prowse DM, Watt FM. Transient activation of beta-catenin signalling in adult mouse epidermis is sufficient to induce new hair follicles but continuous activation is required to maintain hair follicle tumours. Development. 2004;131:1787–1799. doi: 10.1242/dev.01052. [DOI] [PubMed] [Google Scholar]
- Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu. Rev. Cell Dev. Biol. 2004;20:781–810. doi: 10.1146/annurev.cellbio.20.010403.113126. [DOI] [PubMed] [Google Scholar]
- Lowry WE, Blanpain C, Nowak JA, Guasch G, Lewis L, Fuchs E. Defining the impact of beta-catenin/Tcf transactivation on epithelial stem cells. Genes Dev. 2005;19:1596–1611. doi: 10.1101/gad.1324905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackenzie IC. Retroviral transduction of murine epidermal stem cells demonstrates clonal units of epidermal structure. J. Invest. Dermatol. 1997;109:377–383. doi: 10.1111/1523-1747.ep12336255. [DOI] [PubMed] [Google Scholar]
- Madison BB, Braunstein K, Kuizon E, Portman K, Qiao XT, Gumucio DL. Epithelial hedgehog signals pattern the intestinal crypt-villus axis. Development. 2005;132:279–289. doi: 10.1242/dev.01576. [DOI] [PubMed] [Google Scholar]
- Mavilio F, Pellegrini G, Ferrari S, Di Nunzio F, Di Iorio E, Recchia A, Maruggi G, Ferrari G, Provasi E, Bonini C, et al. Correction of junctional epidermolysis bullosa by transplantation of genetically modified epidermal stem cells. Nat. Med. 2006;12:1397–1402. doi: 10.1038/nm1504. [DOI] [PubMed] [Google Scholar]
- Merrill BJ, Gat U, Dasgupta R, Fuchs E. Tcf3 and Lef1 regulate lineage differentiation of multipotent stem cells in skin. Genes Dev. 2001;15:1688–1705. doi: 10.1101/gad.891401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore KA, Lemischka IR. Stem cells and their niches. Science. 2006;311:1880–1885. doi: 10.1126/science.1110542. [DOI] [PubMed] [Google Scholar]
- Morris RJ, Liu Y, Marles L, Yang Z, Trempus C, Li S, Lin JS, Sawicki JA, Cotsarelis G. Capturing and profiling adult hair follicle stem cells. Nat. Biotechnol. 2004;22:411–417. doi: 10.1038/nbt950. [DOI] [PubMed] [Google Scholar]
- Nguyen H, Rendl M, Fuchs E. Tcf3 governs stem cell features and represses cell fate determination in skin. Cell. 2006;127:171–183. doi: 10.1016/j.cell.2006.07.036. [DOI] [PubMed] [Google Scholar]
- Nicolas M, Wolfer A, Raj K, Kummer JA, Mill P, van Noort M, Hui CC, Clevers H, Dotto GP, Radtke F. Notch1 functions as a tumor suppressor in mouse skin. Nat. Genet. 2003;33:416–421. doi: 10.1038/ng1099. [DOI] [PubMed] [Google Scholar]
- O'Connor NE, Mulliken JB, Banks-Schlegel S, Kehinde O, Green H. Grafting of burns with cultured epithelium prepared from autologous epidermal cells. Lancet. 1981;1:75–78. [PubMed] [Google Scholar]
- Oshima H, Rochat A, Kedzia C, Kobayashi K, Barrandon Y. Morphogenesis and renewal of hair follicles from adult multipotent stem cells. Cell. 2001;104:233–245. doi: 10.1016/s0092-8674(01)00208-2. [DOI] [PubMed] [Google Scholar]
- Pan Y, Lin MH, Tian X, Cheng HT, Gridley T, Shen J, Kopan R. gamma-secretase functions through Notch signaling to maintain skin appendages but is not required for their patterning or initial morphogenesis. Dev. Cell. 2004;7:731–743. doi: 10.1016/j.devcel.2004.09.014. [DOI] [PubMed] [Google Scholar]
- Pellegrini G, Traverso CE, Franzi AT, Zingirian M, Cancedda R, De Luca M. Long-term restoration of damaged corneal surfaces with autologous cultivated corneal epithelium. Lancet. 1997;349:990–993. doi: 10.1016/S0140-6736(96)11188-0. [DOI] [PubMed] [Google Scholar]
- Pellegrini G, Ranno R, Stracuzzi G, Bondanza S, Guerra L, Zambruno G, Micali G, De Luca M. The control of epidermal stem cells (holoclones) in the treatment of massive full-thickness burns with autologous keratinocytes cultured on fibrin. Transplantation. 1999;68:868–879. doi: 10.1097/00007890-199909270-00021. [DOI] [PubMed] [Google Scholar]
- Pinto D, Gregorieff A, Begthel H, Clevers H. Canonical Wnt signals are essential for homeostasis of the intestinal epithelium. Genes Dev. 2003;17:1709–1713. doi: 10.1101/gad.267103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ponder BA, Schmidt GH, Wilkinson MM, Wood MJ, Monk M, Reid A. Derivation of mouse intestinal crypts from single progenitor cells. Nature. 1985;313:689–691. doi: 10.1038/313689a0. [DOI] [PubMed] [Google Scholar]
- Potten CS. The epidermal proliferative unit: the possible role of the central basal cell. Cell Tissue Kinet. 1974;7:77–88. doi: 10.1111/j.1365-2184.1974.tb00401.x. [DOI] [PubMed] [Google Scholar]
- Potten CS, Owen G, Booth D. Intestinal stem cells protect their genome by selective segregation of template DNA strands. J. Cell Sci. 2002;115:2381–2388. doi: 10.1242/jcs.115.11.2381. [DOI] [PubMed] [Google Scholar]
- Radtke F, Clevers H. Self-renewal and cancer of the gut: two sides of a coin. Science. 2005;307:1904–1909. doi: 10.1126/science.1104815. [DOI] [PubMed] [Google Scholar]
- Rangarajan A, Talora C, Okuyama R, Nicolas M, Mammucari C, Oh H, Aster JC, Krishna S, Metzger D, Chambon P, et al. Notch signaling is a direct determinant of keratinocyte growth arrest and entry into differentiation. EMBO J. 2001;20:3427–3436. doi: 10.1093/emboj/20.13.3427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rendl M, Lewis L, Fuchs E. Molecular dissection of mesenchymal-epithelial interactions in the hair follicle. PLoS Biol. 2005;3:e331. doi: 10.1371/journal.pbio.0030331. 10.1371/journal.pbio.0030331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reya T, Clevers H. Wnt signalling in stem cells and cancer. Nature. 2005;434:843–850. doi: 10.1038/nature03319. [DOI] [PubMed] [Google Scholar]
- Rheinwald JG, Green H. Serial cultivation of strains of human epidermal keratinocytes: the formation of keratinizing colonies from single cells. Cell. 1975;6:331–343. doi: 10.1016/s0092-8674(75)80001-8. [DOI] [PubMed] [Google Scholar]
- Richardson GD, Arnott EC, Whitehouse CJ, Lawrence CM, Reynolds AJ, Hole N, Jahoda CA. Plasticity of rodent and human hair follicle dermal cells: implications for cell therapy and tissue engineering. J. Investig. Dermatol. Symp. Proc. 2005;10:180–183. doi: 10.1111/j.1087-0024.2005.10101.x. [DOI] [PubMed] [Google Scholar]
- Ro S, Rannala B. A stop-EGFP transgenic mouse to detect clonal cell lineages generated by mutation. EMBO Rep. 2004;5:914–920. doi: 10.1038/sj.embor.7400218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ronfard V, Rives JM, Neveux Y, Carsin H, Barrandon Y. Long-term regeneration of human epidermis on third degree burns transplanted with autologous cultured epithelium grown on a fibrin matrix. Transplantation. 2000;70:1588–1598. doi: 10.1097/00007890-200012150-00009. [DOI] [PubMed] [Google Scholar]
- Sancho E, Batlle E, Clevers H. Signaling pathways in intestinal development and cancer. Annu. Rev. Cell Dev. Biol. 2004;20:695–723. doi: 10.1146/annurev.cellbio.20.010403.092805. [DOI] [PubMed] [Google Scholar]
- Schmidt GH, Winton DJ, Ponder BA. Development of the pattern of cell renewal in the crypt-villus unit of chimaeric mouse small intestine. Development. 1988;103:785–790. doi: 10.1242/dev.103.4.785. [DOI] [PubMed] [Google Scholar]
- Schroder N, Gossler A. Expression of Notch pathway components in fetal and adult mouse small intestine. Gene Expr. Patterns. 2002;2:247–250. doi: 10.1016/s1567-133x(02)00060-1. [DOI] [PubMed] [Google Scholar]
- Searfoss GH, Jordan WH, Calligaro DO, Galbreath EJ, Schirtzinger LM, Berridge BR, Gao H, Higgins MA, May PC, Ryan TP. Adipsin, a biomarker of gastrointestinal toxicity mediated by a functional gamma-secretase inhibitor. J. Biol. Chem. 2003;278:46107–46116. doi: 10.1074/jbc.M307757200. [DOI] [PubMed] [Google Scholar]
- Shackleton M, Vaillant F, Simpson KJ, Stingl J, Smyth GK, Asselin-Labat ML, Wu L, Lindeman GJ, Visvader JE. Generation of a functional mammary gland from a single stem cell. Nature. 2006;439:84–88. doi: 10.1038/nature04372. [DOI] [PubMed] [Google Scholar]
- Shin K, Fogg VC, Margolis B. Tight junctions and cell polarity. Annu. Rev. Cell. Dev. Biol. 2006;22:207–235. doi: 10.1146/annurev.cellbio.22.010305.104219. [DOI] [PubMed] [Google Scholar]
- Silva-Vargas V, Lo Celso C, Giangreco A, Ofstad T, Prowse DM, Braun KM, Watt FM. Beta-catenin and Hedgehog signal strength can specify number and location of hair follicles in adult epidermis without recruitment of bulge stem cells. Dev. Cell. 2005;9:121–131. doi: 10.1016/j.devcel.2005.04.013. [DOI] [PubMed] [Google Scholar]
- Stanger BZ, Datar R, Murtaugh LC, Melton DA. Direct regulation of intestinal fate by Notch. Proc. Natl. Acad. Sci. USA. 2005;102:12443–12448. doi: 10.1073/pnas.0505690102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stingl J, Eirew P, Ricketson I, Shackleton M, Vaillant F, Choi D, Li HI, Eaves CJ. Purification and unique properties of mammary epithelial stem cells. Nature. 2006;439:993–997. doi: 10.1038/nature04496. [DOI] [PubMed] [Google Scholar]
- Sun TT, Lavker RM. Corneal epithelial stem cells: past, present, and future. J. Investig. Dermatol. Symp. Proc. 2004;9:202–207. doi: 10.1111/j.1087-0024.2004.09311.x. [DOI] [PubMed] [Google Scholar]
- Suzuki K, Fukui H, Kayahara T, Sawada M, Seno H, Hiai H, Kageyama R, Okano H, Chiba T. Hes1-deficient mice show precocious differentiation of Paneth cells in the small intestine. Biochem. Biophys. Res. Commun. 2005;328:348–352. doi: 10.1016/j.bbrc.2004.12.174. [DOI] [PubMed] [Google Scholar]
- Taub R. Liver regeneration: from myth to mechanism. Nat. Rev. Mol. Cell Biol. 2004;5:836–847. doi: 10.1038/nrm1489. [DOI] [PubMed] [Google Scholar]
- Taylor G, Lehrer MS, Jensen PJ, Sun TT, Lavker RM. Involvement of follicular stem cells in forming not only the follicle but also the epidermis. Cell. 2000;102:451–461. doi: 10.1016/s0092-8674(00)00050-7. [DOI] [PubMed] [Google Scholar]
- Teuliere J, Faraldo MM, Deugnier MA, Shtutman M, Ben-Ze'ev A, Thiery JP, Glukhova MA. Targeted activation of beta-catenin signaling in basal mammary epithelial cells affects mammary development and leads to hyperplasia. Development. 2005;132:267–277. doi: 10.1242/dev.01583. [DOI] [PubMed] [Google Scholar]
- Tsukamoto AS, Grosschedl R, Guzman RC, Parslow T, Varmus HE. Expression of the int-1 gene in transgenic mice is associated with mammary gland hyperplasia and adenocarcinomas in male and female mice. Cell. 1988;55:619–625. doi: 10.1016/0092-8674(88)90220-6. [DOI] [PubMed] [Google Scholar]
- Tumbar T, Guasch G, Greco V, Blanpain C, Lowry WE, Rendl M, Fuchs E. Defining the epithelial stem cell niche in skin. Science. 2004;303:359–363. doi: 10.1126/science.1092436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van de Wetering M, Sancho E, Verweij C, de Lau W, Oving I, Hurlstone A, van der Horn K, Batlle E, Coudreuse D, Haramis AP, et al. The beta-catenin/TCF-4 complex imposes a crypt progenitor phenotype on colorectal cancer cells. Cell. 2002;111:241–250. doi: 10.1016/s0092-8674(02)01014-0. [DOI] [PubMed] [Google Scholar]
- van den Brink GR, Bleuming SA, Hardwick JC, Schepman BL, Offerhaus GJ, Keller JJ, Nielsen C, Gaffield W, van Deventer SJ, Roberts DJ, Peppelenbosch MP. Indian Hedgehog is an antagonist of Wnt signaling in colonic epithelial cell differentiation. Nat. Genet. 2004;36:277–282. doi: 10.1038/ng1304. [DOI] [PubMed] [Google Scholar]
- van Es JH, Jay P, Gregorieff A, van Gijn ME, Jonkheer S, Hatzis P, Thiele A, van den Born M, Begthel H, Brabletz T, et al. Wnt signalling induces maturation of Paneth cells in intestinal crypts. Nat. Cell Biol. 2005a;7:381–386. doi: 10.1038/ncb1240. [DOI] [PubMed] [Google Scholar]
- van Es JH, van Gijn ME, Riccio O, van den Born M, Vooijs M, Begthel H, Cozijnsen M, Robine S, Winton DJ, Radtke F, Clevers H. Notch/gamma-secretase inhibition turns proliferative cells in intestinal crypts and adenomas into goblet cells. Nature. 2005b;435:959–963. doi: 10.1038/nature03659. [DOI] [PubMed] [Google Scholar]
- van Genderen C, Okamura RM, Farinas I, Quo RG, Parslow TG, Bruhn L, Grosschedl R. Development of several organs that require inductive epithelial-mesenchymal interactions is impaired in LEF-1-deficient mice. Genes Dev. 1994;8:2691–2703. doi: 10.1101/gad.8.22.2691. [DOI] [PubMed] [Google Scholar]
- Van Mater D, Kolligs FT, Dlugosz AA, Fearon ER. Transient activation of beta-catenin signaling in cutaneous keratinocytes is sufficient to trigger the active growth phase of the hair cycle in mice. Genes Dev. 2003;17:1219–1224. doi: 10.1101/gad.1076103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vauclair S, Nicolas M, Barrandon Y, Radtke F. Notch1 is essential for postnatal hair follicle development and homeostasis. Dev. Biol. 2005;284:184–193. doi: 10.1016/j.ydbio.2005.05.018. [DOI] [PubMed] [Google Scholar]
- Watt FM. Role of integrins in regulating epidermal adhesion, growth and differentiation. EMBO J. 2002;21:3919–3926. doi: 10.1093/emboj/cdf399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto N, Tanigaki K, Han H, Hiai H, Honjo T. Notch/RBP-J signaling regulates epidermis/hair fate determination of hair follicular stem cells. Curr. Biol. 2003;13:333–338. doi: 10.1016/s0960-9822(03)00081-2. [DOI] [PubMed] [Google Scholar]
- Yang Q, Bermingham NA, Finegold MJ, Zoghbi HY. Requirement of Math1 for secretory cell lineage commitment in the mouse intestine. Science. 2001;294:2155–2158. doi: 10.1126/science.1065718. [DOI] [PubMed] [Google Scholar]
- Zeps N, Bentel JM, Papadimitriou JM, D'Antuono MF, Dawkins HJ. Estrogen receptor-negative epithelial cells in mouse mammary gland development and growth. Differentiation. 1998;62:221–226. doi: 10.1046/j.1432-0436.1998.6250221.x. [DOI] [PubMed] [Google Scholar]