

Abstract
The neural basis for syntactic processing was studied using event-related fMRI to determine the locations of BOLD signal increases in the contrast of syntactically complex sentences with center-embedded, object-extracted relative clauses and syntactically simple sentences with right-branching, subject-extracted relative clauses in a group of 15 participants in three tasks. In a sentence verification task, participants saw a target sentence in one of these two syntactic forms, followed by a probe in a simple active form, and determined whether the probe expressed a proposition in the target. In a plausibility judgment task, participants determined whether a sentence in one of these two syntactic forms was plausible or implausible. Finally, in a non-word detection task, participants determined whether a sentence in one of these two syntactic forms contained only real words or a non-word. BOLD signal associated with the syntactic contrast increased in the left posterior inferior frontal gyrus in non-word detection and in a widespread set of areas in the other two tasks. We conclude that the BOLD activity in the left posterior inferior frontal gyrus reflects syntactic processing independent of concurrent cognitive operations and the more widespread areas of activation reflect the use of strategies and the use of the products of syntactic processing to accomplish tasks.
Keywords: fMRI syntax, Task effects
1. Introduction
The syntactic structure of a sentence relates the meanings of individual words to one another and thus allows sentences to convey information about who accomplishes and receives an action (“thematic roles”), the properties of items, and other “relational” information. This “propositional” information contributes to knowledge in semantic memory, and is necessary for reasoning, planning, and other human functions.
Functional neuroimaging has become the principle source of information regarding the neural basis of syntactic processing. These studies have led many researchers to articulate models in which one or another aspect of parsing or interpretation is localized in the left posterior inferior frontal gyrus (Broca’s area), or in smaller portions of this region (Ben-Shachar et al., 2003, 2004; Bornkessel et al., 2005; Cooke et al., 2001; Dapretto and Bookheimer, 1999; Embick et al., 2000; Fiebach et al., 2001, 2004, 2005; Friederici, 2002, 2004; Friederici et al., 2000, 2003; Grewe et al., 2005; Hashimoto and Sakai, 2002; Homae et al., 2002; Indefrey et al., 2001; Just et al., 1996; Keller et al., 2001; Meyer et al., 2000; Moro et al., 2001; Newman et al., 2003; Ni et al., 2000; Roder et al., 2002; Ruschemeyer et al., 2005; Sakai et al., 2001, 2003; Stromswold et al., 1996). This area has also been said to be the locus of the ability to learn new syntactic rules (Opitz and Friederici, 2004); Tettamanti et al. (2002) and Musso et al. (2003) have argued the innate capacity that underlies the ability to acquire the syntax of natural language is localized in this region. Whether these functions are localized in this area is thus of interest to cognitive neuroscience, as well as clinically for its implications regarding deficits expected following particular lesions.
However, methodological and design issues make many studies difficult to interpret. Studies that contrast well-formed and ungrammatical sentences or that compare neurovascular responses to judgments about syntactic wellformedness or propositional meaning and judgments about other aspects of language (such as phonological features of words) are extremely common in this literature (e.g., Friederici et al., 2000, 2003; Homae et al., 2003; Embick et al., 2000). Designs of this sort necessarily involve differences in detecting ill-formedness and in response selection across experimental and baseline conditions that cannot be separated from the differences in parsing and interpretation that are the focus of study. Other features of experiments relinquish control over critical aspects of processing. For instance, the use of sentences with phonologically and orthographically legal non-words (“Jabberwocky”) is designed to eliminate semantic processing from consideration (Indefrey et al., 2001; Roder et al., 2002), but it actually allows each participant to arrive at his/her interpretation of non-words and thematic roles.1 Since thematic role assignment interacts with parsing to affect neurovascular responses (Chen et al., 2006; Bornkessel et al., 2005), this approach allows uncontrolled variables to affect results. Perhaps not surprisingly, studies using all these designs have shown multiple areas of activation associated with syntactic processing.
Designs that vary the syntactic operations required to structure experimental and baseline stimuli within a single task are more likely to reveal the neurovascular responses associated with syntactic processing. However, even studies using such designs have also produced contradictory results.2 For instance, the contrast between sentences with object-extracted noun phrases, illustrated in (1), and sentences with subject-extracted noun phrases, illustrated in (2) and (3), has been widely-used to investigate syntactic processing.3
(1) Object-extracted structure (subject–object (SO) sentence): The juice that the boy spilled stained the rug
(2) Subject-extracted structure (object–subject (OS) sentence): The boy spilled the juice that stained the rug
(3) Subject-extracted structure (subject–subject (SS) sentence): The boy who spilled the juice stained the rug.
Assigning syntactic structure and thematic roles in sentences with object-extracted noun phrases (1) makes greater demands on memory and computational functions than assigning syntactic structure and thematic roles in sentences with subject-extracted noun phrases ((2) and (3)) (Gibson, 1998). These greater demands are expected to lead to increases in neurovascular responses to (1) compared to (2) or (3) in brain regions that assign structure and meaning.4
Studies contrasting these structures (or sentences that vary similar aspects of syntactic structure) in a single task have shown numerous patterns of activations, including isolated activation of a single left perisylvian gyrus that differed in different studies (posterior inferior frontal: Stromswold et al., 1996; superior temporal: Caplan et al., 2002), activation of several left perisylvian areas (posterior inferior frontal and superior temporal sulcus: Ben-Shachar et al., 2003, 2004), bilateral activation of perisylvian gyri (posterior inferior frontal and superior temporal: Just et al., 1996; Caplan et al., 2003; Waters et al., 2003), bilateral activation of non-perisylvian gyri (inferior temporal: Cooke et al., 2001), and no activation at all (Indefrey et al., 2001; Fiebach et al., 2005).
Many factors are likely to have affected neurovascular responses differently in these studies, including technology used (positron emission tomography vs magnetic resonance (MR)), stimulus presentation conditions (blocked vs event related), normalization methods (into Talariach vs spherical space), hemodynamic response models employed (finite impulse response (FIR)) vs γ fit methods of determination of significance of activation (fixed vs random effects analyses), areas selected for analysis (omnibus voxel-wise vs preselected region of interest analyses), languages of stimuli (English, German, Dutch, Hebrew, Japanese), and subtle aspects of sentence structure (relative clauses vs indirect questions). However, none of these factors except the use of weak statistical criteria could have led to areas being activated if they do not differ across the sentence conditions. If the results are not due to weak statistical criteria, they suggest that the parsing and interpretive operations and resource demands that differ between object- and subject-extracted structures are not invariantly localized in the same area. They might be distributed, multifocally implemented, or subject to individual variability in localization. However, two considerations complicate the interpretation of these studies.
First, the tasks that have been used tolerate and lead to the use of ancillary cognitive operations (which may called “strategies”). For instance, in the commonly used sentence verification task, in which a target sentence that varies in syntactic structure is presented followed by a probe sentence in a standard simple syntactic form, the syntactic structure of the target often affects accuracy and/or reaction times for responses to probes (e.g., Just et al., 1996). There are many ways that this could happen: subjects may encode the target phonologically, recall its visual form, or engage in other operations that keep the form of a sentence available. Processes such as these are likely to occur more often when participants process complex sentences, and therefore the areas activated when more and less demanding sentences are contrasted in verification tasks may reflect differential use of strategies in different sentence types, not task-independent parsing and sentence interpretation.
Second, the tasks that have been used interact with parsing and sentence interpretation on-line (Tanenhaus et al., 1995; Spivey et al., 2005). Consider plausibility judgment, for example. There is strong evidence that subjects assess the plausibility of thematic role assignments as syntactic constituents are constructed and interpreted (Trueswell et al., 1994; Pearlmutter and MacDonald, 1995) and that these assessments are used to make plausibility judgments while a sentence is being presented (Garnsey et al., 1989). This affects how plausibility judgments are made in SO and OS sentences. Three sets of thematic roles can be assigned at the embedded verb of object-extracted sentences – the roles determined by the combination of the subject and the verb, those determined by the combination of the object and the verb, and those determined by the combination of the subject, the object, and the verb – and each of these sets of thematic roles can be assessed for its plausibility. For instance, in (1), the plausibility of boy being the agent of spill, of juice being the theme of spill, and of boy being the agent spilling juice, can all be assessed. In contrast, in a subject-extracted sentence, a maximum of two thematic roles can be assigned – and assessed for plausibility – at any point in the sentence (e.g., in (2), one thematic role – that of boy as agent of spill – can be assigned at spilled and two sets of thematic roles – that of juice as theme of spill and that of boy as agent of spill and juice as its theme – can be assigned at juice). Since more plausibility judgments occur at the same points as more thematic roles are assigned, BOLD signal differences between object-and subject-extracted sentences might reflect different interim plausibility judgments, not different demands of parsing and interpretation, in these two sentence types.
To determine the effects of ancillary cognitive operations and of the interaction of syntactic processing and task performance on neurovascular effects of syntactic processing, we studied syntactic processing in the same 15 participants in three separate tasks – sentence verification, plausibility judgment, and non-word detection (see Section 2). In all three tasks, we presented syntactically complex sentences with object-extracted structures (“subject object (SO)” sentences), such as (1), and syntactically simple sentences with subject-extracted structures (“object subject (OS)” sentences), such as (2).
In verification, a target sentence in which all nouns were animate and could accomplish the actions of all verbs was presented in one of these forms, followed by a probe presented in a syntactically simple active form. Participants indicated as quickly and as accurately as possible whether the probe expressed thematic roles in the target sentence or not. In plausibility judgment, nouns were animate or inanimate and implausible sentences violated the requirement of a verb that a noun phrase in a particular grammatical position have certain semantic features, usually that it be animate (e.g., The boy that the rug stained spilled the juice). Participants indicated as quickly and as accurately as possible whether the sentence was plausible. In non-word detection, participants viewed the same plausible sentences as were presented in the plausibility judgment task, but the foils consisted of an equal number of sentences of each syntactic type that contained a non-word. Participants indicated as quickly and as accurately as possible whether the sentence contained a non-word.
As discussed above, in sentence verification, ancillary cognitive mechanisms that encode and retain information may be utilized more frequently for syntactically complex than for syntactically simple targets, and in plausibility judgment, task performance may interact with the assignment of syntactic structure and propositional meaning. In contrast, in non-word detection, the task does not require the assignment of syntactic structure or meaning, and any neurovascular effects of the syntactic structure of the sentences are more easily attributable to the process of assigning syntactic structure per se.
2. Methods
2.1. Participants
Fifteen participants (11 female, 4 male; mean age 21.8 years, range 19–26; all college undergraduates) took part in the research. The study was conducted with the approval of the Human Research Committee at the Massachusetts General Hospital and informed consent was obtained for all participants. All participants were right-handed, native speakers of English and naïve as to the purposes of the study. Participants were paid for their involvement.
2.2. Psychological procedures
Examples of the experimental items are shown in Table 1.
Table 1.
Examples of sentence types
| Verification | |
| Targets | |
| SO: The fireman who the deputy called saved the sailor | |
| OS: The deputy called the fireman who saved the sailor | |
| Probes | |
| True: A deputy called a fireman | |
| False: A fireman called a deputy | |
| Plausibility judgment | |
| Plausible sentences | |
| SO – incongruent noun animacy order | The deputy that the newspaper identified chased the mugger |
| OS – incongruent noun animacy order | The newspaper identified the deputy that chased the mugger |
| SO – congruent noun animacy order | The wood that the man chopped heated the cabin |
| OS – congruent noun animacy order | The man chopped the wood that heated the cabin |
| Implausible sentences | |
| SO – incongruent noun animacy order | The plumber that the hair extracted clogged the sink |
| OS – incongruent noun animacy order | The hair extracted the plumber that clogged the sink |
| SO – congruent noun animacy order | The bill that the activist angered organized the march |
| OS – congruent noun animacy order | The activist angered the bill that organized the march |
| Non-word detection | |
| Sentences with only real words | |
| Sentences with real words: same as in plausibility judgment | |
| Sentences with non-words | |
| Based on SO – incongruent noun animacy order | The deputy that the haberfelt identified chased the mugger |
| Based on OS – incongruent noun animacy order | The newspaper identified deputy that the chorried the mugger |
| Based on SO – congruent noun animacy order | The wood that the man dribed heated the cabin |
| Based on OS – congruent noun animacy order | The man chopped wood that the heated the gert |
2.3. Sentence verification
The experimental items consisted of 72 pairs of synonymous SO and OS target sentences, exemplified in (4) and (5), for a total of 144 items.
(4) The fireman who the deputy called saved the sailor. (SO)
(5) The deputy called the fireman who saved the sailor. (OS)
Each matching pair of sentences had the same identical lexical items. All noun phrases were singular, common, and definite to ensure that participants would not be influenced by the referential assumptions made by the noun phrases in different ways in the two conditions. Sentences were based on scenarios. There were a total of 72 scenarios (such as the scenario involving a deputy calling a fireman who saved a sailor). Each scenario appeared once as an SO and once as an OS sentence, and the same words were used in each syntactic form of the scenario.
Verification probe sentences were always presented as simple active sentences and either conveyed one set of thematic roles in the target sentence (e.g., a deputy calling a fireman) for the “true” condition or reversed the thematic roles played by the noun phrases around the verbs (e.g., a fireman calling a deputy) or paired nouns with the wrong verb (e.g., a deputy calling a sailor) for the “false” condition. Each target was followed by one probe. An equal number of true and false verification sentences was presented for each sentence type, and both true and false probes were related to the main and subordinate clauses in the target sentences equally often. The task for the participants during the experimental trial was to indicate whether the verification probe sentence conveyed a set of thematic roles that has been conveyed in the target sentence. Response time (RT) and accuracy were measured.
2.4. Plausibility judgment
The experimental items consisted of 144 pairs of SO and OS sentences (see Table 1). Sentences were based on scenarios, each of which appeared once as an SO and once as an OS sentence. Each matching pair of sentences had identical lexical items. All noun phrases were singular, common, and definite to ensure that subjects would not be influenced by the referential assumptions made by the noun phrases in a sentence in different ways in the two conditions. Half the items of each structure in each condition were plausible and the other half were implausible. The violations in plausibility were the result of mismatches in semantic features required by a verb and the animacy of the subject or object noun of the clause. The animacy of subject and object noun phrases and the plausibility of the sentences was systematically varied within each sentence type. Sentences in which the first noun is animate and the second noun inanimate support the interpretation derived from the syntax in OS sentences (congruent animacy order) and conflict with that interpretation in SO sentences (incongruent animacy order) (Traxler et al., 2002; Chen et al., 2006). Implausible sentences became implausible at various points in the relative clauses and the main clauses, to ensure that subjects had to read each sentence in its entirety before they could decide if it was plausible. Overall, the point at which SO sentences became implausible was earlier than the point at which OS sentences became implausible, reducing the possibility that subjects could decide on strategic grounds that an OS sentence was plausible at an earlier point than was possible in a SO sentence.
The task for the participants was to read the sentence and judge the plausibility of the presented item as quickly and accurately as possible. A plausible sentence was described as a sentence that had a meaning the participant could imagine happening in the real world. RTs and accuracy were recorded.
2.5. Non-word detection
The experimental items consisted of the 144 pairs of SO and OS sentences from the plausibility judgment task with changes to the implausible sentences to render them plausible. Half the pairs were altered to include a phonologically and orthographically legal non-word, as illustrated in Table 1. Non-words were located in positions occupied by both the nouns and verbs in both the relative and the main clauses.
The stimulus presentation was identical to that in the plausibility judgment tasks, except that subjects were told to detect the presence of a non-word as accurately and quickly as possible.
2.6. MR procedures
2.6.1. MR imaging parameters
Participants were scanned in three separate sessions, one devoted to each task, on separate days, with the order of tasks counterbalanced across participants. In the initial session, in addition to the functional imaging, two sets of high-resolution anatomical images were acquired in a 3.0T whole body Siemens Trio scanner (Siemens Medical Systems, Iselin, NJ) using a T1-weighted MP-RAGE sequence (TR = 2530 msec, TE = 3 msec, and flip angle = 7°). Volumes consisted of 128 sagittal slices with an effective thickness of 1.33 mm. The in-plane resolution was 1.0 mm × 1.0 mm (256 × 256 matrix, 256 mm Field of View (FOV)).
The functional sessions utilized the same scanner. The functional volume acquisitions utilized a T2*-weighted gradient-echo pulse sequence (TR = 2000 msec, TE = 30 msec, and flip angle = 90°). The volume was comprised of 30 transverse slices aligned along the same AC-PC plane as the registration volume. The interleaved slices were effectively 3 mm thick with a distance of .9 mm between slices. The in-plane resolution was 3.13 × 3.13 mm (64 × 64 matrix, 200 mm FOV). Each run consisted of 200 such volume acquisitions for a total of 6000 images. By definition, the 30 slices of a single volume took the entire TR (2s) to be fully acquired and a new volume was initiated every TR. An initial 8 sec (4 TR equivalent) buffer of RF pulse activations, during which no stimulus items were presented and no functional volumes were acquired, was employed to ensure maximal signal during the length of the functional run.
2.7. Item presentation
In all experiments, each stimulus (target or probe) was visually displayed in its entirety on a single line in the center of the screen. The sentences were projected to the back of the scanner using a Sharp LCD projector and viewed by the participants as a reflection in a mirror attached to the head coil. Responses were recorded via a custom-designed, magnet compatible button box. A Dell Inspiron 4000 computer running a proprietary software package was used to both present the stimuli and record the accuracy and reaction times.
A pseudo-randomized item presentation order for the event-related design was determined by a computer program developed to randomize trial types and vary the duration of inter-stimulus fixation trials for optimum efficiency in the deconvolution and estimation of the hemodynamic response (Burock et al., 1998; Dale, 1999; Dale and Buckner, 1997). In the verification task, 0–12 sec fixation trials (a centered +) were randomly interspersed between sentence trials. In the plausibility judgment and non-word detection tasks, fixation trials were randomly interspersed between each 6 sec sentence trial.
In the verification task, the 144 stimulus trials’ items interspersed with fixation trials were divided into six runs. No pair of matched SO and OS sentences were presented in the same run. A given experimental trial consisted of a fixation cross presented for 300 msec, a 100 msec blank screen, the target sentence presented for 4 sec, a blank screen for 1700 msec, the verification probe sentence presented for 4 sec, and a blank screen for 1900 msec, for a total trial length of 12 sec. In the plausibility judgment and non-word detection tasks, the 288 stimulus items interspersed with the fixation trials were divided into eight runs. No pair of matched SO and OS sentences were presented in the same run. A given experimental trial consisted of a brief 300 msec fixation cross, a 100 msec blank screen, the sentence presented for 5 sec, and a final 600 msec blank screen, for a total trial length of 6 sec. Participants were given a short break after each run.
2.8. Cortical surface reconstruction
The high-resolution anatomical MP-RAGE scans were used to construct a model of each participant’s cortical surface. An average of the two structural scans was used to maximize the signal-to-noise ratio. The cortical reconstruction procedure involved: (1) segmentation of the cortical white matter; (2) tessellation of the estimated border between gray and white matter, providing a geometrical representation for the cortical surface of each participant; and (3) inflation of the folded surface tessellation to unfold cortical sulci, allowing visualization of cortical activation in both the gyri and sulci simultaneously (Dale et al., 1999; Fischl et al., 1999a, 2001).
For purposes of inter-subject averaging, the reconstructed surface for each participant was morphed onto an average spherical representation. This procedure optimally aligns sulcal and gyral features across participants, while minimizing metric distortion, and establishes a spherical-based coordinate system onto which the selective averages and variances of each participant’s functional data can be resampled (Fischl et al., 1999a, 1999b).
2.9. Functional pre-processing
Pre-processing and statistical analysis of the functional MRI data were performed using the FreeSurfer Functional Analysis Stream (FS-FAST) developed at the Martinos Center, Charlestown, MA (Burock and Dale, 2000). For each participant, the acquired native functional volumes were first corrected for potential motion of the participant using the AFNI algorithm (Cox, 1996). Next, the functional volumes were spatially smoothed using a 3-D Gaussian filter with a full-width half-max (FWHM) of 6 mm. Global intensity variations across runs and participants were removed by rescaling all voxels and time points of each run such that the mean in a brain intensity was fixed at an arbitrary value of 1000.
The functional images for each participant were analyzed with a General Linear Model (GLM) using a FIR model of the event-related hemodynamic response (Burock and Dale, 2000). The FIR gives an estimate of the hemodynamic response average at each TR within a pre-stimulus window. The FIR does not make any assumption about the shape of the hemodynamic response. Mean offset and linear trend regressors were included to remove low-frequency drift. The autocorrelation function of the residual error, averaged across all brain voxels, was used to compute a global whitening filter in order to account for the intrinsic serial autocorrelation in fMRI noise. The GLM parameter estimates and residual error variances of each participant’s functional data were resampled onto his or her inflated cortical surface and into the spherical coordinate system using the surface transforms described above. Each participant’s data were then smoothed on the surface tessellation using an iterative nearest-neighbor averaging procedure equivalent to applying a two-dimensional Gaussian smoothing kernel with a FWHM of approximately 8.5 mm. Because this smoothing procedure was restricted to the cortical surface, averaging data across sulci or outside gray matter was avoided.
2.10. Voxel-wise analysis (or statistical activation maps)
Group statistical activation maps were constructed for contrasts of interest using a t statistic. Contrasts of interest were tested at each voxel on the spherical surface across the group using a random effects model of the cross-participant variance of the FIR parameter estimates. Contrasts were constructed over a window of post-stimulus delays in the FIR model corresponding to the delays at which vascular responses were expected to peak. BOLD signal changes follow electrophysiological events associated with elementary sensory stimuli and simple motor functions by as little as 2 sec, with an established response by 4–6 sec (Bandettini et al., 1993; Turner, 1997). For the verification task, we analyzed BOLD signal in an “early” time window from 6 to 12 sec after the onset of a trial, during which BOLD signal associated with processing the target sentence would be expected to peak, and a “late” time window from 12 to 18 sec after the onset of a trial, during which BOLD signal associated with processing the verification probe sentence would be expected to peak. For the plausibility judgment and non-word detection tasks, we analyzed the hemodynamic response from 6 to 12 sec after the onset of the trial.
To correct for multiple spatial comparisons, we identified significant clusters of activated voxels on the basis of a Monte Carlo simulation (Doherty et al., 2004). A volume of Gaussian distributed numbers was generated for each subject, and was processed in the same manner as the real data, including volumetric smoothing, resampling onto the sphere, smoothing on the spherical surface, random effects analysis, and activation map generation. A clustering program was run on these maps to extract clusters of voxels whose members each exceeded a specified voxel-level p-value threshold and whose area was equal to or greater than a specified size. This process was repeated 3500 times, allowing us to compute the likelihood of one or more clusters of a given size and voxel-level threshold occurring under the null hypothesis. The real data were then subjected to the same clustering procedure as applied to the simulated data using a cluster size threshold of 200 mm2 and threshold for rejection of the null hypothesis at p < .05. These functional activations were displayed on a map of the average folding patterns of the cortical surface, derived using the surface-based morphing procedure (Fischl et al., 1999a, 1999b). The accompanying Talairach coordinates correspond to the vertices within each cluster with the minimum local p-value (i.e., the voxel with the greatest significance level).
3. Results
3.1. Behavioral results
Analyses of variance were performed over participant and item means for accuracy and RTs that had been trimmed for outliers ± 3 sd from the condition mean for each participant. Significant terms of significant interactions were identified using Tukey’s test.
There were effects of syntactic structure in all three tasks (Fig. 1).
Fig. 1.

Accuracy and RTs for different sentence conditions in verification, plausibility judgment (plausible sentences) and non-word detection (sentences with real words). SO: subject object structure; OS: object subject structure. Incongruent: order of animacy of noun phrases biases against syntactically determined meaning; congruent: order of animacy of noun phrases biases towards syntactically determined meaning.
In verification, there was an effect of structure in accuracy (F1(1, 14) = 11.8, p < .01; F2(1, 70) = 16.0, p < .001) and RTs (F1(1, 14) = 27.2, p < .001; F2(1, 35) = 31.3, p < .001). Participants were more accurate and faster in responding to OS than to SO sentences. The fact that accuracy was higher for SO than OS sentences could be due to difficulties assigning structure and meaning to SO sentences, but the fact that RTs were faster for correct responses to SO sentences than for correct responses to OS sentences indicates that cognitive mechanisms that maintained the syntactic structure of a sentence in short term memory, or that allowed it to be reconstructed from another representation, were operational in this study. The effect of probe type was significant in the RT data only (F1(1, 14) = 43.3, p < .001; F2(1, 35) = 29.2, p < .001), as was the interaction of these factors (F1(1, 14) = 10.1, p < .01; F2(1, 35) = 4.5, p < .05). Responses were longer to false verification statements.
In plausibility judgment, for plausible sentences, there were effects of syntactic structure (for accuracy, F1(1, 14) = 18.2, p < .001; F2(1, 136) = 38.4, p < .001; for RTs, F1(1, 14) = 63.8, p < .001; F2(1, 136) = 41.9, p < .001) and an interaction of syntactic structure with noun phrase animacy order (for accuracy, F1(1, 14) = 36.3, p < .001; F2(1, 136) = 55.8, p < .001; for RTs, F1(1, 14) = 49.6, p < .001; F2(1, 136) = 97.7, p < .001). As noted above and in Section 4, the order of animacy of noun phrases biased either towards (was congruent with) or against (was incongruent with) the syntactically determined meaning of a sentence (Traxler et al., 2002). Responses were more accurate and faster for the less complex OS sentences than for the more complex SO sentences for sentences in which the order of the animacy of the noun phrases biased against (was incongruent with) the syntactically determined meaning of the sentences. There was an interaction between the position of implausibility and sentence type (F(1.14) = 5.8, p < .05). RTs were longer for SO than OS sentences that became implausible at V1 but not at V2.
In the non-word detection task, for sentences containing only real words, there were no effects of structure or noun phrase animacy order in the accuracy data, due to ceiling effects. In the RT data, there was an effect of structure (F1(1, 14) = 20.0, p < .001; F2(1, 108) = 4.3, p < .05) and an interaction of structure with order of animacy of nouns (F1(1, 14) = 35.0, p < .001; F2(1, 108) = 6.2, p < .01). Responses were faster for OS than for SO sentences in which the order of the animacy of the noun phrases biased against the syntactically determined meaning of the sentences.
The fact that the interaction of syntactic structure and noun phrase animacy order in the non-word detection task was significant indicates that participants processed stimuli to the level of syntactic structure and sentence-level meaning in that task. However, despite the fact that this interaction was significant by both subjects and items, some participants may have processed only a few stimuli to the sentence level. Three other aspects of the results in the non-word detection task were considered to see if this was likely to have been the case.
First, RTs to sentences with non-words were analyzed for the effect of the position of the non-word. If participants treated the stimuli as sentences, one would expect an effect of position, with longer RTs to sentences in which the non-word appeared further from the start of the sentence. If participants treated the stimuli as strings of words, they might be expected to scan the string for a non-word as in a visual search experiment with closely matched foils, and an effect of position of the non-word might not be expected. The data are shown in Fig. 2. RTs increased as the non-word appeared further into the sentence (F1(1, 56) = 63.9, p < .001; F2(1, 125) = 31.8, p < .001), and the differences between all positions were significant by Tukey’s test.
Fig. 2.

Mean RT for correct responses in non-word detection for sentences containing non-words, showing increased RTs as a function of non-word location and longer RTs for object- than for subject- extracted sentences with non-words at the position of the embedded verb (V1).
Second, if participants processed the stimuli in the non-word detection study as sentences, one might expect that the effect of position of the non-word on RTs would be affected by sentence type. Reaction times might be longer for complex sentences than for simple sentences when the non-word appeared in the syntactically demanding position of the complex sentence, and not otherwise. The data are shown in Fig. 2. Reaction times for correct responses to subject–object sentences with non-words in embedded verb position (2287 msec) were significantly longer than those for object–subject sentences with non-words in either main verb or object positions (1994 msec and 1982 msec, respectively) while reaction times for sentences with non-words in other positions did not differ (for the interaction of location of non-word and sentence type, F1(1, 56) = 6.9, p < .001; F2(1, 125) = 3.2, p < .01). This strongly suggests that subjects assigned aspects of structure and meaning even to the sentences with non-words.
Third, if participants processed some sentences deeply to the level of meaning and others only superficially to the level of word form, one might expect that the distribution of RTs would be bimodal. Histogams of RTs by deciles are shown in Fig. 3. These showed unimodal distributions for all sentence types (all bimodality coefficients (b) < .555).
Fig. 3.

Histogram of RTs for responses in non-word detection. Top panels: subject-extracted sentences. Blue: sentences with real words; red: sentences with non-words. Bottom panels: object-extracted sentences. Pink: sentences with real words; green: sentences with non-words.
Comparisons of responses across tasks were examined for cues regarding how participants processed syntactic structures. Because the stimuli in the verification task differed from those in the other two tasks and responses in the verification task were made to probes not a target sentence, a comparison of performance in the verification and the other two tasks is not easily interpretable. However, the same participants saw the same plausible sentences containing only real words in the plausibility and non-word detection tasks; the differences in these tasks consisted in what aspects of these stimuli were relevant to the responses. Differences in performances between the two experiments were therefore of interest and were compared.
For the plausible sentences containing only real words, analysis of variances (ANOVAs) showed a significant effect of task in accuracy (F1(1, 14) = 9.3, p < .01; F2(1, 37) = 17.0, p < .001) and RTs (F1(1, 14) = 8.9, p < .01; F2(1, 37) = 53.6, p < .001). Accuracy was higher and RTs were faster in non-word detection than in plausibility judgment. This indicates that some operation in plausibility judgment was more demanding than an operation in non-word detection; making the plausibility judgment as opposed to determining that there was a non-word in the sentence is the obvious candidate for such an operation.
There was also an interaction of task X structure for RTs (F1(1, 14) = 21.0, p < .001; F2(1, 37) = 13.5, p < .001). As noted, there was a significant difference in RTs for SO and OS sentences in both tasks, but the difference in RTs was greater in plausibility judgment (389 msec) than in non-word detection (158 msec) (t = 3.8, p (one tailed) < .001). This indicates that the effect of syntactic complexity was greater on some operation that took place in plausibility judgment than it was on some operation that took place in non-word detection. A reasonable candidate for this operation would be use of the plausibility of a sentence to perform the task, if, as we indicated above, determination of the plausibility of a sentence takes place on-line and is more demanding at the point of syntactic complexity in the complex SO sentences, and this information is used incrementally to perform the judgment task.
3.2. fMRI results
3.2.1. Sentence verification
The areas of significant difference in BOLD signal responses to SO and OS sentences in the “early” and “late” TR periods are shown in Fig. 4 and summarized in Table 2.5
Fig. 4.

Areas of BOLD signal differences between stimuli in the verification task. Color overlays represent p-values of the contrast such that the color threshold (red) corresponds to p = .01 and ceilings (yellow) at p = .001. Blue overlays represent areas in which BOLD signal was reduced. Each area has a minimum area of 200 mm2 and a false-positive p < .05. The number label for each cluster corresponds to the cluster with the number contrast in Table 2
Left top: early TR interval (6–12 sec post-stimulus onset), SO–OS contrast.
Left bottom: early TR interval (6–12 sec post-stimulus onset), false–true contrast.
Right top: late TR interval (12–18 sec post-stimulus onset), SO–OS contrast.
Right bottom: late TR interval (12–18 sec post-stimulus onset), false–true contrast.
Table 2.
Talairach coordinates, gyri and Brodmann area localizations of peak activation, and cluster size, for clusters of activation in the early TR intervals, corresponding to the target sentence, and the late TR intervals, corresponding to the probe sentence, in the verification task
| Contrast | Left hemisphere
|
Right hemisphere
|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cluster | Region | BA | Talairach (x, y, z) | Size (mm2) | p-Value | Cluster | Region | BA | Talairach (x, y, z) | Size (mm2) | p-Value | |
| Early TR interval of sentence verification: effect of syntactic structure or probe type | ||||||||||||
| Syntactic structure | 1 | Superior temporal gyrus | 42 | (−52, −24, 10) | 272 | −.000043 | 2 | Precentral gyrus | 4 | (17, −16, 57) | 266 | −.000022 |
| 3 | Precentral gyrus | 4 | (31, −11, 46) | 320 | −.000193 | |||||||
| 4 | Superior temporal gyrus | 41/42 | (49, −25, 15) | 221 | −.000004 | |||||||
| Probe type | 1 | Inferior frontal gyrus | 45/44 | (−44, 13, 21) | 230 | .001282 | 6 | Supramarginal gyrus | 40 | (50, −46, 44) | 360 | .001462 |
| 2 | Middle frontal gyrus | 9 | (−43, 8, 38) | 399 | .000175 | |||||||
| 3 | Supramarginal gyrus | 40 | (−49, −38, 36) | 525 | .000031 | |||||||
| 4 | Angular gyrus | 39/40 | (−45, −49, 28) | 356 | .000101 | |||||||
| 5 | Precuneus | 7 | (−14, −57, 40) | 460 | .000002 | |||||||
| Late TR interval of sentence verification: effect of syntactic structure or probe type | ||||||||||||
| Syntactic structure | 1 | Inferior frontal gyrus | 44 | (−52, 13, 9) | 763 | .000065 | 10 | Supramarginal gyrus | 40 | (38, −56, 40) | 330 | .000237 |
| 2 | Middle frontal gyrus | 9 | (−55, 8, 38) | 526 | .000010 | 11 | Supramarginal gyrus | 40 | (33, −44, 42) | 720 | .000179 | |
| 3 | Superior temporal gyrus | 39 | (−39, −58, 29) | 1277 | .000174 | 12 | Middle frontal gyrus | 8/9 | (33, 13, 32) | 1236 | .000030 | |
| 4 | Middle temporal gyrus | 21 | (−48, −35, −1) | 354 | .000003 | 13 | Superior frontal gyrus | 8 | (−1, 31, 41) | 427 | .000515 | |
| 5 | Superior temporal gyrus | 22 | (−55, −16, 3) | 399 | −.000337 | |||||||
| 6 | Precuneus | 31 | (−16, −46, 40) | 559 | .000290 | |||||||
| 7 | Superior frontal gyrus | 6 | (−19, 8, 57) | 390 | .000830 | |||||||
| 8 | Medial frontal gyrus | 10 | (−18, 59, 2) | 232 | −.000723 | |||||||
| 9 | Anterior cingulate gyrus | 24/32 | (−9, 36, 1) | 254 | −.000305 | |||||||
| Probe type | 1a | Inferior frontal gyrus | 44/45 | (−61, 18, 6) | 3489 | .000001 | 7 | Occipito-parietal sulcus | 19/40 | (37, −63, 39) | 792 | .000889 |
| 1b | Middle frontal gyrus | 9 | (−53, 6, 39) | 500 | .000028 | 8 | Angular gyrus | 40/39 | (49, −50, 33) | 222 | .000877 | |
| 2 | Angular gyrus | 39/40 | (−43, −52, 33) | 1530 | .000001 | 9 | Supramarginal gyrus | 40 | (36, −40, 34) | 250 | .000036 | |
| 3a | Superior temporal gyrus | 22 | (−53, −45, 18) | 2684 | .000001 | 10 | Middle frontal gyrus | 9 | (26, 36, 29) | 2573 | .000005 | |
| 3b | Middle temporal gyrus | 21 | (−57, −25, −3) | 625 | .000005 | 11 | Insula | (25, 25, 0) | 314 | .000139 | ||
| 4 | Precuneus | 7 | (−11, −54, 41) | 865 | .000005 | 12 | Middle temporal gyrus | 21 | (49, −15, −8) | 297 | .000518 | |
| 5 | Superior frontal gyrus | 6 | (−14, 14, 58) | 996 | .000037 | 13 | Superior frontal gyrus | 8 | (−1, 27, 49) | 225 | .004227 | |
| 6 | Gyrus rectus | 11 | (−6, 37, −23) | 362 | .000001 | 14 | Precuneus | 7 | (4, −62, 46) | 542 | .000032 | |
All localizations are derived from estimates in Talairach and Tournoux (1988) atlas for the voxel with the minimum p-value (the voxel with the most significant effect) in each cluster. The gyrus and Brodmann area in which the most significant voxel is localized, or the gyrus and Brodmann area closest to a maximum in white matter, is listed. The cluster numbers correspond to clusters in Fig. 4. Note that areas colored in blue in Fig. 4 are ones in which BOLD signal was greater for OS than for SO sentences.
In the early temporal interval period from 6 to 12 sec after the onset of a trial, there were a few areas of activation in which BOLD signal was greater for OS than for SO sentences (the left superior temporal gyrus, and the right precentral and superior temporal gyri). In this temporal interval, there were also several areas in which BOLD signal was greater for sentences with probes that were false than for sentences with probes that were true (the left inferior and middle frontal gyri, the left angular gyrus, the supramarginal gyri bilaterally, and the medial aspect of the left superior parietal lobule (the medial precuneus)).
In the late temporal interval period from 12 to 18 sec after the onset of a trial, associated with the presentation of the probe, there were multiple areas in which BOLD signal was greater for SO than for OS sentences. The areas activated on the lateral surface of the left hemisphere – the inferior and middle frontal gyri, an arc of tissue in the inferior parietal lobule whose most active voxel was located in the superior temporal gyrus, and the middle temporal gyrus – are located in or adjacent to the perisylvian association cortex of the left hemisphere. On the medial surface of the left hemisphere, there was increased BOLD signal in the superior frontal gyrus and the precuneus. There were also three areas in which BOLD signal was higher for OS than for SO sentences – one on the lateral surface in the superior temporal gyrus and two on the medial surface, in the anterior cingulate and medial frontal gyri. In the right hemisphere, there was increased BOLD signal for SO compared to OS sentences in two areas in the supramarginal gyrus, in the middle frontal gyrus, and in an area on the medial surface of the superior frontal gyrus. All differences in BOLD activity as a function of sentence type occurred in association with true probes (i.e., there were no significant areas of activation for SO compared to OS sentences for false probes).
There were 14 areas in which BOLD signal was higher for false probes than for true probes. These areas were fairly symmetrical on the lateral hemispheric surfaces, and were found in frontal, temporal and parietal lobes. The extent and magnitude of these areas were greater in this temporal period than in the earlier temporal period.
3.2.2. Plausibility judgment
As noted, the sentences used in plausibility judgment varied in both syntactic structure and in whether the order of animacy of noun phrases was congruent with the meaning of the sentence as determined by its syntax. Fig. 5 displays the statistical activation maps of the contrasts for plausible SO and OS sentences that attained statistical significance, as a function of the fit between the order of animacy of noun phrases and the meaning of the sentence as determined by its syntax. The areas in which there was significant BOLD signal activity in each contrast are summarized in Table 3.
Fig. 5.

Areas of BOLD signal differences between plausible stimuli in the plausibility judgment task. Color overlays represent p-values as in Fig. 4. The number label for each cluster corresponds to the cluster with the number contrast in Table 3.
Left panel: SO–OS contrast for sentences in which noun animacy order biases towards the meaning of the sentence (“congruent” stimuli).
Right panel: SO–OS contrast for sentences in which noun animacy order biases against the meaning of the sentence (“incongruent” stimuli).
Table 3.
Talairach coordinates, gyri and Brodmann area localizations of peak activation, and cluster size, for clusters of activation in the comparison of plausible SO and OS sentences types in plausibility judgment
| Contrast | Left hemisphere
|
Right hemisphere
|
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cluster | Region | BA | Talairach (x, y, z) | Size (mm2) | p-Value | Cluster | Region | BA | Talairach (x, y, z) | Size (mm2) | p-Value | ||||||||
| Plausibility judgment: effect of syntactic structure | |||||||||||||||||||
| Congruent | 1 | Middle frontal gyrus | 6 | (−42, 3, 43) | 345 | .000013 | 8 | Precuneus | 7 | (22, −50, 51) | 1301 | .000009 | |||||||
| 2 | Middle temporal gyrus | 37 | (−63, −55, −2) | 576 | .000081 | 9 | Insula | (25, 17, −6) | 350 | .000033 | |||||||||
| 3 | Inferior temporal gyrus | 19 | (−55, −78, 7) | 240 | .003155 | 10 | Fusiform gyrus | 36/37 | (40, −39, −9) | 483 | .000002 | ||||||||
| 4 | Inferior temporal gyrus | 18 | (−29, −83, −5) | 2681 | .000018 | 11 | Lingual gyrus | 8 | (11, −83, 2) | 2823 | .000001 | ||||||||
| 5 | Inferior temporal gyrus | 20 | (−56, −44, −18) | 276 | .000331 | 12 | Precuneus | 7 | (3, −49, 49) | 488 | .000274 | ||||||||
| 6 | Precuneus | 7 | (−16, −56, 48) | 397 | .000005 | 13 | Precuneus | 7 | (18, −67, 43) | 381 | .000566 | ||||||||
| 7 | Posterior cingulate and cuneus | 31 | (−17, −64, 16) | 237 | .001545 | 14 | Lingual gyrus | 19 | (21, −54, 6) | 567 | .000536 | ||||||||
| Incongruent | 1 | Inferior frontal gyrus | 45/47 | (−41, 29, 2) | 518 | .000167 | 8 | Superior frontal gyrus | 6 | (13, 2, 47) | 475 | .000007 | |||||||
| 2a | Inferior frontal gyrus | 44 | (−49, 12, 20) | 4605 | .000165 | 9 | Middle frontal gyrus | 6 | (29, 6, 40) | 220 | .001208 | ||||||||
| 2b | Middle frontal gyrus | 6 | (−38, −1, 50) | – | .000008 | 10 | Inferior frontal gyrus | 45 | (35, 34, 14) | 1066 | .000389 | ||||||||
| 2c | Precentral gyrus | 6 | (−52, 1, 24) | – | .000139 | 11 | Inferior frontal gyrus | 45 | (43, 33, 2) | 436 | .000818 | ||||||||
| 2d | Inferior frontal gyrus | 44 | (−56, 10, 10) | – | .000004 | 12 | Superior frontal gyrus | 6 | (0, 19, 43) | 363 | .000294 | ||||||||
| 2e | Superior frontal gyrus | 6 | (−15, 12, 49) | – | .000035 | ||||||||||||||
| 3 | Supramarginal gyrus | 40 | (−43, −41, 34) | 407 | .002037 | ||||||||||||||
| 4 | Precuneus | 7 | (−35, −62, 39) | 668 | .000130 | ||||||||||||||
| 5 | Middle temporal gyrus | 21 | (−56, −46, 4) | 1313 | .000002 | ||||||||||||||
| 6 | Fusiform gyrus | 37 | (−49, −51, −19) | 1931 | .000055 | ||||||||||||||
| 7 | Fusiform gyrus | 19 | (−32, −64, 4) | 298 | .000067 | ||||||||||||||
The cluster numbers correspond to clusters in Fig. 5.
The comparison of SO and OS sentences in which the order of animacy of noun phrases was congruent with the syntactically derived meaning of the sentence showed increases in BOLD signal in the middle frontal, middle and inferior temporal gyri, the medial aspect of the precuneus and adjacent portions of the cuneus of the left hemisphere, and the precuneus, the insula, and the fusiform and lingual gyri of the right hemisphere.
The comparison of SO and OS sentences in which the order of animacy of noun phrases was incongruent with the syntactically derived meaning of the sentence showed extensive increases in BOLD signal in the left hemisphere in the inferior, middle, and superior frontal gyri, the inferior and superior parietal lobe, the middle and fusiform temporal gyri, and the precentral gyrus. In the right hemisphere, the contrast showed increases in BOLD activity in the three frontal gyri.
3.2.3. Non-word detection
Fig. 6 displays the statistical maps of the contrasts for plausible sentences that attained statistical significance. Table 4 is a listing of the Talairach coordinate locations of the local minimum p-value within each cluster.
Fig. 6.
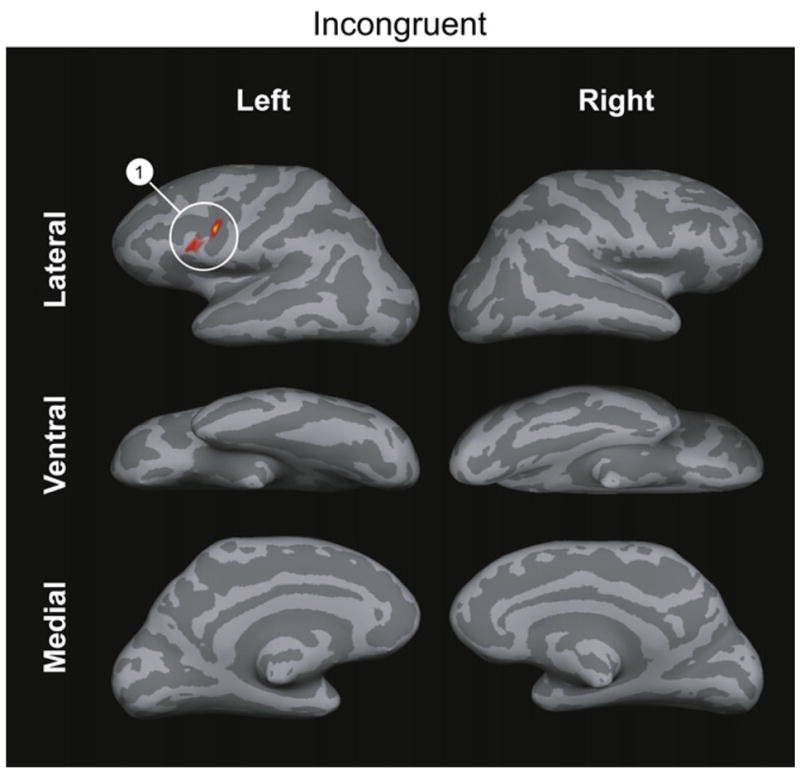
Areas of BOLD signal differences between SO and OS sentences containing real words in which noun animacy order biases against the meaning of the sentence (“incongruent” stimuli) in the non-word judgment task. Color overlays represent p-values as in Fig. 4. The number label for each cluster corresponds to the cluster with the number contrast in Table 4.
Table 4.
Talairach coordinates, gyri andBrodmann area locationof peak activation, and cluster size, for comparison of SOand OS sentences types with real words only in which the order of animacy of noun phrases biased against the syntactically determined meaning in non-word detection
| Contrast | Left hemisphere
|
|||||
|---|---|---|---|---|---|---|
| Cluster | Region | BA | Talairach (x, y, z) | Size (mm2) | p-Value | |
| Non-word detection: effect of syntactic structure | ||||||
| Incongruent | 1 | Inferior frontal gyrus | 44 | (−44,6,15) | 453 | .000170 |
The cluster numbers correspond to clusters in Fig. 6.
The comparison of SO and OS sentences in which the order of animacy of noun phrases was congruent with the syntactically derived meaning of the sentence showed no areas of activation. The comparison of SO and OS sentences in which the order of animacy of noun was incongruent with the syntactically derived meaning of the sentence showed significant BOLD signal in the left posterior inferior frontal gyrus (BA 44).
4. Discussion
The BOLD signal effects can be interpreted in light of the behavioral results.6 With respect to the verification task, the behavioral data provide evidence that participants utilized one or more ancillary cognitive mechanism. With respect to the plausibility judgment task, they show that participants performed an operation in plausibility judgment that was more difficult than any in non-word detection and that operation was more difficult when a syntactically complex SO sentence was being processed than when a syntactically simple OS sentence was being processed. An obvious candidate for this operation is the on-line use of the plausibility of the sentence to perform the task. With respect to the non-word detection task, they show that, though the task did not require them to do so, participants reliably processed sentences to the level of syntactic structure and meaning.
The BOLD signal effects of the syntactic contrast used here were much more widespread in the verification and plausibility judgment tasks than in the non-word detection task, where they were limited to the left posterior inferior frontal gyrus. A possible explanation of the differences in the areas activated in these tasks is that the activity in the left posterior inferior frontal gyrus reflects the demands and operations associated with the process of constructing the syntactic structure of the more complex compared to the less complex sentences and assigning the meaning of that sentence in the face of a bias against that meaning due to the order of noun animacy, and that the activity in other areas reflects the greater use of ancillary cognitive operations in the more complex compared to the less complex sentences in the verification task and the greater demands associated with using the products of syntactic and interpretive processes to accomplish the demands of the task on-line in the more complex than the less complex sentences in the plausibility judgment task. We shall first discuss several considerations that arise on the assumption that this interpretation of the data is correct, and then turn to a discussion of alternative interpretations of the main results of these experiments.
The principal finding in the verification task was that increases in BOLD signal arose in association with verifying the truth of statements referring to previously presented more complex SO syntactic structures compared to less complex OS syntactic structures, and did so in a late time period associated with the presentation of the probe and not in an early time period associated with the presentation of the target sentences. The effect of structure in the late time period could be due to very delayed neurovascular effects of processing associated with the comprehension of the targets. This is somewhat unlikely given the usual temporal relation of BOLD signal to stimulus presentation (Bandettini et al., 1993; Turner, 1997), but the fact that the target and probe were not presented as separate events and that the hemodynamic responses to them cannot be modeled separately does not allow us to rule out this possibility. However, as noted, the behavioral data show that something more than just the meaning of the target sentence was available and processed at the point of the probe. Even if the initial syntactic processing of the target sentence contributed to the BOLD signal responses seen in the late time periods, it is highly likely that other processes did so as well.
What might these other representations and processes be? We have mentioned that phonological representations may be stored in memory (McCutchen and Perfetti, 1982) and used to reconstruct syntactic structure and thence propositional meaning, and that orthographic representations may be stored and used either to construct phonological representations and/or to access semantic and syntactic lexical information from which the propositional content of the target can be reconstructed. Another possibility is that participants maintained a richer semantic representation of the target in memory than is required for the task, one that included the discourse-level information that the first mentioned noun is the focus of the discourse. This information distinguishes SO and OS sentences that convey the same thematic roles and also makes SO sentences more complex than OS sentences, since the focus of the discourse in an SO sentence is the theme of the verb of the relative clause, which is a relatively infrequent combination of semantic values (Van Dijk and Kintsch, 1983; Ericsson and Kintsch, 1995). A third possibility is that the syntactic structure that had been computed during comprehension of the target was retained (Traxler and Pickering, 2004) and re-used during verification.
Increases in BOLD signal associated with SO compared to OS sentences occurred in two broad regions: brain areas related to language (the left inferior, and possibly middle, frontal, inferior parietal, and the left and possibly the right inferior parietal gyri, can be considered to be possibly language-related cortex) and brain areas related to cognitive control (more anterior and superior bilateral frontal regions and the left cingulate). The exact mechanisms that are responsible for these activations can only be determined by more detailed studies, but we may speculate that the activity in the first of these sets of areas may reflect increased rehearsal and phonological storage of the target SO sentences (Smith et al., 1998) and that the activity in the second of these sets of areas may reflect aspects of cognitive control that differ in these sentence types, such as deciding when enough evidence has accrued to allow a response to be made (Menon et al., 2001).
In connection with this latter possibility, a finding worth noting in the verification task is that there was greater BOLD signal associated with false probes, consistent with the longer RTs to those probes. Greater BOLD signal and longer RTs to false probes may occur because, on average, more checking of propositions in the probe and target sentence is likely to occur in false than in true probes. For instance, if participants randomly select one set of the thematic roles derived from the target to match against those of the probe, on half the true trials a positive match will occur after checking the first proposition, allowing the “yes” response to be selected, while on the false trials, it will always be necessary to check both propositions to determine the correct response. If this is the case, areas in which false probes produced greater BOLD signal than true probes may be ones in which matching the meaning of one proposition against that of another takes place. Alternatively, some of the activation in this contrast may be due to an “error” signal associated with recognizing that one proposition does not correspond to another. Again, we may speculate about which cognitive processes might be responsible for neurovascular effects in different areas. Following the considerations outlined above, matching propositional content of the probe to the target – and the use of rehearsal and phonological storage in this effort – is likely to have activated perisylvian cortical areas involved in language processing (the left inferior and possibly the middle frontal gyri and the left and possibly the right inferior parietal and middle and superior temporal areas), while neurovascular responses are more likely to have been generated in response to error signals in the orbital and superior frontal gyri. The reason for other increases in BOLD signal in this contrast – such as those in the superior parietal lobe – is unclear at present.
In the plausibility judgment task, the behavioral data replicate previously documented results (Chen et al., 2006; Caplan and Chen, 2006) and parallel results in eye fixation measures in a verification task (Traxler et al., 2002). They show effects of syntactic structure, which are more extensive when the order of animacy of nouns is incongruent with the syntactically derived meaning of the sentence. Some of this activity is likely to be due to the increased processing demands made by the combination of the need to construct a more complex syntactic structure and to assign thematic roles in that structure in the face of competing possibilities for thematic roles derived from heuristics based upon the order of animacy of nouns. However, even considering just this task in isolation, the fact that RTs were disproportionately increased in implausible SO sentences in which the embedded verb created the implausibility suggests that some part of the BOLD signal associated with the contrast of the syntactically more and less complex sentences is likely to also be due to task-related process. We have said that these task-related processes involve the incremental use of plausibility ratings of thematic roles to perform the task. More specifically, we envisage this process to consist of the incremental transfer of plausibility ratings of thematic roles to a response selection process and the incremental weighting of that process on the basis of these ratings.
We have a clue as to what part of the BOLD signal effect in the plausibility judgment task is not due to these sentence type-task performance interactions from the results in the non-word detection task, because the use of the products of syntactic and interpretive processes were unlikely to have been used to perform the task. In that task, effects of syntactic structure in sentences with incongruent noun animacy order were found only in the left posterior inferior frontal gyrus. Because the smaller extent of BOLD signal in the syntactic contrasts in the non-word detection task is not likely to be due to participants only rarely processing sentences deeply, we may tentatively conclude that this activation reflects the extent of the area that supports the syntactic and interpretive processing that differs between these sentence types in this task. As we have emphasized, this processing occurs incidentally in this task, and is therefore not likely to have been accompanied by ancillary cognitive operations whose purpose would be to keep the syntactic structure or semantic meaning of the sentences available, or by operations that use this information to accomplish the task. The left posterior inferior frontal gyrus thus emerges as a candidate for the area in which these syntactic and interpretive operations take place, leaving all the other areas activated in the SO–OS contrasts in the verification and plausibility judgment experiments as ones in which concurrent ancillary cognitive operations occur more often in SO than in OS sentences and in which assigning the structure and meaning of these sentences interacts with accomplishing tasks.
Turning to the parsing and interpretive operations that differ in the contrasts we used, several models of the syntactic and interpretive operations that occur in this contrast have been proposed. One of the most explicit has been developed by Traxler et al. (2002), who studied eye fixation patterns in similar sentences (SO and SS sentences – (1) and (3) above). Traxler et al. (2002) found longer first-pass eye fixation durations in SO than in SS sentences regardless of noun animacy and thematic role plausibility, and concluded that the first operations participants undertook were purely syntactic and that these operations were more demanding for the initial structuring of the object-extracted clause in SO sentences than for the subject-extracted clause in SS sentences. Upon encountering these demands of parsing, participants re-inspected the sentence-initial noun. At this point and going forward, syntax and semantics interacted. All fixation measures after first-pass eye fixations on the relative clause and main verb were longer in the SO sentences with animate sentence-initial nouns than in all other sentences, and were the same in all other sentences. Traxler et al. concluded that, in SO sentences with animate sentence-initial nouns, the animacy of the sentence-initial noun biased against the object-relativized structure, leading to increased processing load at a second stage of processing. The BOLD signal effects in the non-word detection task would be due to these increased demands, if we relate them to this model.
What alternatives are there to concluding that the BOLD signal effect in the non-word detection study reflects some aspect of task-independent, strategy-free syntactic and interpretive processing, and that the more widespread effects in the other studies reflect other, co-ocurring, cognitive operations? Answers to this question fall into two categories: (1) the suggestion that parts of the BOLD signal effect of the SO–OS contrast found in the verification and plausibility tasks that fall outside the area activated in the non-word detection task result from task- and strategy-free syntactic and interpretive operations, and (2) the suggestion that the BOLD signal effect of the SO–OS contrast in the non-word detection task are not pure reflections of task- and strategy-free syntactic and interpretive operations.
The first of these suggestions could be expanded as follows. As noted above, syntactic and semantic operations interact in the assignment of the meaning of the sentences used here. Different areas could be activated in different contrasts in the verification and plausibility judgment tasks because of differences in these interactions. Specifically, in the verification task, the sentences were all “equibiased” in the sense that all nouns were roughly equally plausible as agents and themes of all verbs. In contrast, in the plausibility judgment task, the sentences were all “biased” in the sense that the animacy order of the nouns reinforced or contradicted the meaning conveyed by the syntax. One possibility is that the different areas that were activated in the SO–OS contrast in these tasks are associated with the different interactions of the plausibility of different sets of thematic roles based on noun animacy order and syntactically derived meanings.
This argument, however, runs into the same problem encountered above. The contrast between SO and OS sentences in which noun animacy order biased against the meaning of the sentence activated only one area – the left posterior inferior frontal gyrus – in both the tasks in which this contrast occurred (plausibility judgment and non-word detection). If the area activated by this contrast in non-word detection is the one in which the interaction of that structure with the use of the particular noun animacy order found in those sentences takes place, the other areas activated by this contrast in plausibility judgment cannot be supporting these operations (unless these operations are supported by different areas in different individuals). It is possible – though unlikely, in our opinion – that other areas are activated in plausibility judgment and verification by comprehension processes associated with the interaction of the syntax and sets of thematic roles derived from other noun animacy orders. This question could be pursued by analyses of these thematic and syntactic contrasts in multiple tasks, including tasks in which syntactic processing is incidental.
This brings us to the second possibility suggested above –that the BOLD signal effect of the SO–OS contrast in the non-word detection task is itself not a pure reflection of task- and strategy-free syntactic and interpretive operations. There are two reasons this might be the case.
First, although we have said that sentence-level processing in this task occurs incidentally and is not likely to have engendered the use of ancillary cognitive operations, we cannot be sure of this analysis of how participants performed the task. Ancillary cognitive operations might be associated with parsing and interpretation even if parsing and interpretation are incidental. For instance, a reviewer of an earlier version of this paper suggested that the activity in the left posterior inferior frontal gyrus may be due to the involvement of this area in eye movement control. Similarly, we cannot know from this study alone whether participants rehearsed the sentences in the non-word detection task, and whether they did so to a greater extent for object- than for subject-extracted sentences. Neither of these possible ancillary operations seem likely to us to be the source of the BOLD signal effect in the left inferior posterior frontal gyrus. With respect to eye movement control, this area is quite distant from the frontal eye fields. With respect to rehearsal, the reaction times are sufficiently fast that it is unlikely that participants engaged in much sub-vocal rehearsal. However, this study does not rule out these, or other, operations as possible sources of the BOLD signal effects found in this task. Other protocols, such as ones that include measuring eye fixation or the use of articulatory suppression, could address these possibilities.
The second reason that the BOLD signal effect of the SO–OS contrast in the non-word detection task may not be a pure reflection of task- and strategy-free syntactic and interpretive operations is that the task may not avoid the “contamination” of syntactic analysis and interpretation by incremental task performance. Non-word detection does not require a response based on the structure and meaning of a sentence, but this property of the task, while a step towards eliminating task effects, does not go far enough. What is required to ensure that the BOLD signal associated with a syntactic contrast is not partially a result of the use of syntactic analysis and interpretation to perform a task is not a task that does not require these aspects of processing but a task that cannot be performed on the basis of a syntactic structure and its interpretation. Non-word detection does not meet this stronger requirement: it can be accomplished on the basis of the semantic meaning of the sentence. Participants can decide if a stimulus contains a non-word if the structure of the sentence or its propositional meaning is lexically incomplete.
There are tasks that cannot be accomplished on the basis of the structure or meaning of the sentence, such as a font change detection task in which object- and subject-extracted sentences are presented, half with all words in the same font and half with one word in a similar but different font. Whether one sees syntactic and interpretive effects in such tasks depends on many factors that affect the time rise of information relevant to the response and information about the structure of the sentence. Systematic study of syntactic and semantic variables on BOLD signal in such tasks may help identify the areas that support various aspects of task-independent syntactic and associated interpretive processing more convincingly.
To summarize, the present study provides evidence that certain aspects of syntactic processing and closely related use of semantic information to assign thematic roles in object- compared to subject-extracted sentences is supported by a small brain area, the left posterior inferior frontal gyrus, and that a large number of brain regions are activated by this contrast in many studies because individuals utilize strategies and use the products of syntactic processing and interpretation to perform tasks.
This conclusion, which will require additional studies to be confirmed, raises a basic question about the nature of the syntactic operations that are independent of task. Until very recently, the standard view was that, although the assignment of syntactic structure may not be as independent from the use of information derived from statistical regularities of the language, plausibility and context as highly modular theories (e.g., Frazier and Clifton, 1986) had postulated, parsing and sentence interpretation were independent of perception of visual context, planning actions and encoding meanings derived from linguistic stimuli into memory. This view has been seriously challenged by recent work. Tanenhaus et al. (2000) summarized a large body of research using visual world tracking as follows: “It is becoming increasing clear that even the earliest moments of linguistic processing are modulated by context… call[ing] into question the long-standing belief that perceptual systems create context-independent perceptual representations (p. 577).”
This poses two very difficult challenges. First, if Tanenhaus is correct, there is likely to be considerable variability in cognitive systems that become activated during comprehension, and this variability may occur across individuals performing the same task, across tasks within an individual, and even within an individual performing the same task on different occasions. Second, this perspective leads to the question of which, if any, linguistic operations are task-independent. Despite the very impressive evidence cited by Tanenhaus pointing to the early interaction of visual analysis and motor planning with syntactic analysis, it remains possible – indeed likely, in our view – that some syntactic operations are unaffected by tasks (e.g., assignment of very local structures such as prepositional phrases), and that the parser provides a range of possible structures that are quickly narrowed down by task context. Confronting these challenges seems to us to be essential to making progress towards understanding the neural basis for syntactic processing.
Footnotes
This work was supported by Grant DC02146 from NIH.
It is precisely the attribution of referential and semantic values to the non-words that gives Jabberwocky its power. These values are idiosyncratic. In Through Looking Glass, Lewis Carroll illustrates the phenomenon in the glosses that Humpty Dumpty gives Alice immediately following the recitation of “Jabberwocky.” These include interpretations such as that brillig means “four o’clock in the afternoon – the time when you begin broiling things for dinner” and that toves are “something like badgers, lizards and corkscrews, that make nests under sundials and live on cheese.”
The studies of the neural basis for learning syntax in adults (Tettamanti et al., 2002; Musso et al., 2003; Opitz and Friederici, 2004) have been more consistent, reporting BOLD signal response increases to novel grammars only in left inferior frontal lobe, though here too the exact areas of activation have ranged over a fairly large portion of this area. Other design issues – such as the choice of foils in the grammaticality judgment task in the Tettamanti et al. and Musso et al. studies, which may not have led to participants classifying stimuli as well-formed or ill-formed according to principles of Universal Grammar – complicate the interpretation of some of these studies.
There are two separate ways that the labels “SO” and “OS” have been applied to sentences. We and others have used them to refer to the location of relative clauses and the extraction of NPs in those clauses. In this usage, SO refers to a relative clause located on the subject of the sentence, in which the object of the relative clause has been extracted. In our work, SO sentences have a non-canonical thematic role order when simple transitive verbs taking agent – patient/theme roles are used. In the German literature, the term SO has been used to refer to the order of subjects and objects that correspond to sentences with a canonical subject–object thematic role order. An additional possible source of confusion is that “thematic role order” is sometimes, but not always, called “word order.” We will continue to use the labels SO, SS and OS to refer to relative clause location and NP extraction as we have in previous publications, hoping this consistency will lead to the least confusion.
SO and OS sentences differ along more syntactic dimensions than SO and SS sentences. SO sentences differ from OS sentences in having both an object-extracted relative clause and a center-embedded configuration of clauses and from SS sentences only in the first of these respects. The contrast of SO and SS sentences thus focuses on a more specific aspect of syntactic structure than the contrast between SO and OS does (for extensive discussion of related issues, see Fiebach et al., 2004). However, the greater number of syntactic differences between SO and OS than between SO and SS sentences is an advantage for the goal of determining whether syntactic effects arise in association with the comprehension, encoding, retrieval and/or verification of sentence meaning because it increases the likelihood of finding a BOLD signal effect of syntactic structure. In addition, OS sentences can be made to be synonymous with SO sentences, which further increases the likelihood that any BOLD signal effects are due to syntactic processing compared to the comparison of SO and SS sentences, which cannot be synonymous.
The tables list the gyri and Brodmann areas in which the voxel with the most reliable activation (lowest p-value) was found, or the gyrus and BA immediately above this voxel if the voxel was subcortical. An anonymous reviewer cautioned that these localizations are based on normalizing to the Talairach and Tournoux (1988) atlas and are subject to various inaccuracies. In addition, we note that the localizations listed do not capture the entirety of the activation. For instance, cluster 3 in the contrast of SO and OS sentences in the late TR periods in the verification experiment is largely in the inferior parietal lobe, but extends into the superior temporal gyrus (which is continuous with the angular gyrus) and its most significant voxel is in the temporal, not the parietal, lobe. Similarly, in the plausibility judgment task contrast of SO and OS sentences with congruent noun animacy order (Table 3, Fig. 5), the maximally significant voxel in cluster 7 localizes to the posterior cingulate, but is immediately adjacent to the cuneus. Readers are encouraged to examine the figures themselves to obtain a fuller view of the areas that were activated in particular comparisons.
Several results in the verification task require comment. The effect of probe type in the early interval must reflect processing of the probe, which appeared before the end of the “early” time window (see below for discussion of why false probes should lead to more BOLD signal than true probes). The increased BOLD signal for OS than SO sentences in the early interval may be due to participants matching OS sentences to probes earlier than SO sentences. It is not clear why this effect was found in the late time intervals.
References
- Bandettini PA, Jesmanowicz A, Wong EC, Hyde JS. Processing strategies for time-course data sets in functional MRI of the human brain. Magnetic Resonance in Medicine. 1993;30:161–173. doi: 10.1002/mrm.1910300204. [DOI] [PubMed] [Google Scholar]
- Ben-Shachar M, Hendler T, Kahn I, Ben-Bashat D, Grodzinsky Y. The neural reality of syntactic transoformations: evidence from fMRI. Psychology Science. 2003;14:433–440. doi: 10.1111/1467-9280.01459. [DOI] [PubMed] [Google Scholar]
- Ben-Shachar M, Palti D, Grodsinsky Y. The neural correlates of syntactic movement: converging evidence from two fMRI experiments. Neuroimage. 2004;21:1320–1336. doi: 10.1016/j.neuroimage.2003.11.027. [DOI] [PubMed] [Google Scholar]
- Bornkessel I, Zysset S, Friederici AD, Von Cramon DY, Schlesewsky M. Who did what to whom? The neural basis of argument hierarchies during language comprehension. Neuroimage. 2005;26:221–233. doi: 10.1016/j.neuroimage.2005.01.032. [DOI] [PubMed] [Google Scholar]
- Burock MA, Buckner RL, Woldorff MG, Rosen BR, Dale AM. Randomized event-related experimental designs allow for extremely rapid presentation rates using functional MRI. Neuroreport. 1998;9:3735–3739. doi: 10.1097/00001756-199811160-00030. [DOI] [PubMed] [Google Scholar]
- Burock MA, Dale AM. Estimation and detection of event-related fMRI signals with temporally correlated noise: a statistically efficient and unbiased approach. Human Brain Map. 2000;11:249–260. doi: 10.1002/1097-0193(200012)11:4<249::AID-HBM20>3.0.CO;2-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caplan D, Chen E. Using fMRI to discover cognitive operations. Cortex. 2006;42:393–395. doi: 10.1016/s0010-9452(08)70366-6. [DOI] [PubMed] [Google Scholar]
- Caplan D, Vijayan S, Kuperg G, West C, Waters G, Greve D, Dale AM. Vascular responses to syntactic processing: event-related fMRI study of relative clauses. Human Brain Map. 2002;15:26–38. doi: 10.1002/hbm.1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caplan D, Waters G, Alpert N. Effects of age and speed of processing on rCBF correlates of syntactic processing in sentence comprehension. Human Brain Map. 2003;19:112–131. doi: 10.1002/hbm.10107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen E, West WC, Waters G, Caplan D. Determinants of BOLD signal correlates of processing object-extracted relative clauses. Cortex. 2006;42:591–604. doi: 10.1016/s0010-9452(08)70397-6. [DOI] [PubMed] [Google Scholar]
- Cooke A, Devita C, Gee J, Aslop D, Detre J, Chen W, Grossman M. Neural basis for sentence comprehension: grammatical and short-term memory components. Human Brain Map. 2001;15:80–94. doi: 10.1002/hbm.10006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox RW. AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Computers and Biomedical Research. 1996;29:162–173. doi: 10.1006/cbmr.1996.0014. [DOI] [PubMed] [Google Scholar]
- Dale AM. Optimal experimental design for event-related fMRI. Human Brain Map. 1999;8:109–114. doi: 10.1002/(SICI)1097-0193(1999)8:2/3<109::AID-HBM7>3.0.CO;2-W. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dale AM, Buckner RL. Selective averaging of rapidly presented individual trials using fMRI. Human Brain Map. 1997;5:329–340. doi: 10.1002/(SICI)1097-0193(1997)5:5<329::AID-HBM1>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- Dale AM, Fischl B, Sereno MI. Cortical surface-based analysis –I: Segmentation and surface reconstruction. Neuroimage. 1999;9:179–194. doi: 10.1006/nimg.1998.0395. [DOI] [PubMed] [Google Scholar]
- Dapretto M, Bookheimer SY. Form and content: dissociating syntax and semantics in sentence comprehension. Neuron. 1999;24:427–432. doi: 10.1016/s0896-6273(00)80855-7. [DOI] [PubMed] [Google Scholar]
- Doherty CP, West WC, Dilley LC, Shattuck-Hufnagel S, Caplan D. Question/statement judgments: an fMRI study of intonation processing. Human Brain Mapping. 2004;23:85–98. doi: 10.1002/hbm.20042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Embick D, Marantz A, Miyashita Y, O’Neil W, Sakai KL. A syntactic specialization for Broca’s area. Proceedings of the National Academy of Sciences of the United States of America. 2000;97:6150–6154. doi: 10.1073/pnas.100098897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ericsson KA, Kintsch W. Long-term working memory. Psychological Review. 1995;102:211–245. doi: 10.1037/0033-295x.102.2.211. [DOI] [PubMed] [Google Scholar]
- Fiebach CJ, Schlesewsky M, Lohmann G, Von Cramon DY, Friederici AD. Revisiting the role of Broca’s area in sentence processing: syntactic integration versus syntactic working memory. Human Brain Mapping. 2005;24:79–91. doi: 10.1002/hbm.20070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiebach CJ, Schlesewsky M, Friederici AD. Syntactic working memory and the establishment of filler-gap dependencies: insights from ERPs and fMRI. Journal of Psycholinguistic Research. 2001;30:321–338. doi: 10.1023/a:1010447102554. [DOI] [PubMed] [Google Scholar]
- Fiebach CJ, Vos SH, Friederici AD. Neural correlates of syntactic ambiguity in sentence comprehension for low and high span readers. Journal of Cognitive Neuroscience. 2004;16:1562–1575. doi: 10.1162/0898929042568479. [DOI] [PubMed] [Google Scholar]
- Fischl B, Sereno MI, Dale AM. Cortical surface-based analysis –II: Inflation, flattening, and a surface-based coordinate system. Neuroimage. 1999a;9:195–207. doi: 10.1006/nimg.1998.0396. [DOI] [PubMed] [Google Scholar]
- Fischl B, Sereno MI, Tootell RB, Dale AM. High-resolution intersubject averaging and a coordinate system for the cortical surface. Human Brain Map. 1999b;8:272–284. doi: 10.1002/(SICI)1097-0193(1999)8:4<272::AID-HBM10>3.0.CO;2-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischl B, Liu A, Dale AM. Automated manifold surgery: constructing geometrically accurate and topologically correct models of the human cerebral cortex. IEEE Transactions on Medical Imaging. 2001;20:70–80. doi: 10.1109/42.906426. [DOI] [PubMed] [Google Scholar]
- Frazier L, Clifton C. The independence of syntactic processing. Journal of Memory and Language. 1986;25:348–368. [Google Scholar]
- Friederici A, Opitz B, Von Cramon DY. Segregating semantic and syntactic aspects of processing in the human brain: an fMRI investigation of different word types. Cerebral Cortex. 2000;10:698–705. doi: 10.1093/cercor/10.7.698. [DOI] [PubMed] [Google Scholar]
- Friederici A, Ruschemeyer SA, Hahne A, Fiebach CJ. The role of left inferior frontal and superior temporal cortex in sentence comprehension: localizing syntactic and semantic processes. Cerebral Cortex. 2003;13:170–177. doi: 10.1093/cercor/13.2.170. [DOI] [PubMed] [Google Scholar]
- Friederici A. Towards a neural basis of auditory sentence processing. Trends in Cognitive Science. 2002;6:78–84. doi: 10.1016/s1364-6613(00)01839-8. [DOI] [PubMed] [Google Scholar]
- Friederici A. Processing local transition versus long-distance syntactic dependencies. Trends in Cognitive Science. 2004;8:245–247. doi: 10.1016/j.tics.2004.04.013. [DOI] [PubMed] [Google Scholar]
- Garnsey SM, Tanenhaus MK, Chapman RM. Evoked potentials and the study of sentence comprehension. Journal of Psycholinguistic Research. 1989;18:51–60. doi: 10.1007/BF01069046. [DOI] [PubMed] [Google Scholar]
- Gibson E. Syntactic complexity: locality of syntactic dependencies. Cognition. 1998;68:1–76. doi: 10.1016/s0010-0277(98)00034-1. [DOI] [PubMed] [Google Scholar]
- Grewe TG, Bornkessel I, Zysset S, Wiese R, Von Cramon DY, Schlesewsky M. The emergence of the unmarked: a new perspective on the language-specific function of Broca’s area. Human Brain Mapping. 2005;26:178–190. doi: 10.1002/hbm.20154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashimoto R, Sakai K. Specialization in the left prefrontal cortex for sentence comprehension. Neuron. 2002;35:589–597. doi: 10.1016/s0896-6273(02)00788-2. [DOI] [PubMed] [Google Scholar]
- Homae F, Yahata N, Sakai KL. Selective enhancement of functional connectivity in the left prefrontal cortex during sentence processing. NeuroImage. 2003;20:578–586. doi: 10.1016/s1053-8119(03)00272-6. [DOI] [PubMed] [Google Scholar]
- Homae F, Hashimoto R, Nakajima K, Miyashita Y, Sakai KL. From perception to sentence comprehension: the convergence of auditory and visual information of language in the left inferior frontal cortex. NeuroImage. 2002;16:883–900. doi: 10.1006/nimg.2002.1138. [DOI] [PubMed] [Google Scholar]
- Indefrey P, Hagoort P, Herzog H, Seitz RJ, Brown CM. Syntactic processing in the left prefrontal cortex in independent of lexical meaning. NeuroImage. 2001;14:546–555. doi: 10.1006/nimg.2001.0867. [DOI] [PubMed] [Google Scholar]
- Just MA, Carpenter PA, Keller TA, Eddy WF, Thulborn KR. Brain activation modulated by sentence comprehension. Science. 1996;274:114–116. doi: 10.1126/science.274.5284.114. [DOI] [PubMed] [Google Scholar]
- Keller TA, Carpenter PA, Just MA. The neural bases of sentence comprehension: a fMRI examination of syntactic and lexical processing. Cerebral Cortex. 2001;11:223–237. doi: 10.1093/cercor/11.3.223. [DOI] [PubMed] [Google Scholar]
- McCutchen D, Perfetti CA. The visual tongue-twister effect: phonological activation in silent reading. Journal of Verbal Learning and Verbal Behavior. 1982;21:672–687. [Google Scholar]
- Menon V, Adleman NE, White CD, Glover GH, Reiss AL. Error-related brain activation during a Go/NoGo response inhibition task. Human Brain Mapping. 2001;12:131–143. doi: 10.1002/1097-0193(200103)12:3<131::AID-HBM1010>3.0.CO;2-C. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer M, Friederici AD, Von Cramon DY. Neurocognition of auditory sentence comprehension: event related fMRI reveals sensitivity to syntactic violations and task demands. Cognitve Brain Research. 2000;9:19–33. doi: 10.1016/s0926-6410(99)00039-7. [DOI] [PubMed] [Google Scholar]
- Moro A, Tettamanti M, Perani D, Donati C, Cappa SF, Fazio F. Syntax and the brain: disentangling grammar by selective anomalies. NeuroImage. 2001;13:110–118. doi: 10.1006/nimg.2000.0668. [DOI] [PubMed] [Google Scholar]
- Musso M, Moro A, Glauche V, Rijntjes M, Reichenbach J, Buchel C, Weiller C. Broca’s area and the language instinct. Nature Neuroscience. 2003;6:774–781. doi: 10.1038/nn1077. [DOI] [PubMed] [Google Scholar]
- Newman S, Just MA, Keller TA, Roth J, Carpenter PA. Differential effects of syntactic and semantic processing on the subregions of Broca’s area. Brain Research. Cognitive Brain Research. 2003;16:297–307. doi: 10.1016/s0926-6410(02)00285-9. [DOI] [PubMed] [Google Scholar]
- Ni W, Constable RT, Mencl WE, Pugh KR, Fulbright RK, Shaywitz BA, Gore JC, Shankweiler D. An event-related neuroimaging study distinguishing form and content in sentence processing. Journal of Cognitive Neuroscience. 2000;12:120–133. doi: 10.1162/08989290051137648. [DOI] [PubMed] [Google Scholar]
- Opitz B, Friederici AD. Brain correlates of language learning: the neuronal dissociation of rule-based versus similarity-based learning. Journal of Neuroscience. 2004;24:8436–8440. doi: 10.1523/JNEUROSCI.2220-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pearlmutter NJ, MacDonald MC. Individual differences and probabalistic constraints in syntactic ambiguity resolution. Journal of Memory and Language. 1995;34:521–542. [Google Scholar]
- Roder B, Stock O, Neville H, Bien S, Rosler F. Brain activation modulated by the comprehension of normals and psedo-word sentencesof different processing demands:afunctionalmagnetic resonance imaging study. NeuroImage. 2002;15:1003–1014. doi: 10.1006/nimg.2001.1026. [DOI] [PubMed] [Google Scholar]
- Ruschemeyer S, Fiebach CJ, Kempe V, Friederici AD. Processing lexical semantic and syntactic information in first and second language: fMRI evidence from German and Russian. Human Brain Mapping. 2005;25:266–286. doi: 10.1002/hbm.20098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakai KL, Hashimoto R, Homae F. Sentence processing in the cerebral cortex. Neuroscience Research. 2001;39:1–10. doi: 10.1016/s0168-0102(00)00205-4. [DOI] [PubMed] [Google Scholar]
- Sakai KL, Homae F, Hashimoto R. Sentence processing is uniquely human. Neuroscience Research. 2003;46:273–279. doi: 10.1016/s0168-0102(03)00122-6. [DOI] [PubMed] [Google Scholar]
- Smith EE, Jonides J, Marshuetz C, Koeppe RA. Components of verbal working memory: evidence from neuroimaging. Proceedings of the National Academy of Sciences of the United States of America. 1998;95:876–882. doi: 10.1073/pnas.95.3.876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spivey MJ, Tanenhaus MK, Eberhard KM, Sedivy JC. Eye movements and spoken language comprehension: effects of visual context on syntactic ambiguity resolution. Cognitive Psychology. 2005;45:447–481. doi: 10.1016/s0010-0285(02)00503-0. [DOI] [PubMed] [Google Scholar]
- Stromswold K, Caplan D, Alpert N, Rauch S. Localization of syntactic comprehension by positron emission tomography. Brain Language. 1996;52:452–473. doi: 10.1006/brln.1996.0024. [DOI] [PubMed] [Google Scholar]
- Talairach J, Tournoux P. Co-planar Stereotaxic Atlas of the Human Brain: 3-Dimensional Proportional System – an Approach to Cerebral Imaging. New York, NY: Thieme Medical Publishers; 1988. [Google Scholar]
- Tanenhaus MK, Spivey-Knowlton MJ, Eberhard KM, Sedivy JC, et al. Integration of visual and linguistic information in spoken language comprehension. Science. 1995;268:1632–1634. doi: 10.1126/science.7777863. [DOI] [PubMed] [Google Scholar]
- Tanenhaus M, Maghuson JS, Dahan D, Chambers C. Eye movements and lexical access in spoken-language comphrension: evaluating a linking hypothesis between fixations and linguistic processing. Journal of Psycholinguistic Research. 2000;29:557–580. doi: 10.1023/a:1026464108329. [DOI] [PubMed] [Google Scholar]
- Tettamanti M, Alkadhi H, Moro A, Perani D, Kollias S, Weniger D. Neural correlates for the acquisition of natural language syntax. NeuroImage. 2002;17:700–709. [PubMed] [Google Scholar]
- Traxler MJ, Pickering MJ. Syntactic priming in comprehension, architectures and mechanisms for language processing conference. Aix en Provence; France: 2004. [Google Scholar]
- Traxler M, Morris R, Seely R. Processing subject and object relative clauses: evidence from eye movements. Journal Memory Language. 2002;47:69–90. [Google Scholar]
- Trueswell JC, Tanenhaus M, Garnsey S. Semantic influences on parsing: use of thematic role information in syntactic ambiguity resolution. Journal Memory Language. 1994;33:285–318. [Google Scholar]
- Turner R. Signal sources in BOLD contrast fMRI. Advance Experiment Medical Biology. 1997;413:19–25. doi: 10.1007/978-1-4899-0056-2_2. [DOI] [PubMed] [Google Scholar]
- Van Dijk TA, Kintsch W. Strategies of Discourse Comprehension. New York: Academic Press; 1983. [Google Scholar]
- Waters G, Caplan D, Alpert N, Stanczak L. Individual differences in rCBF correlates of syntactic processing in sentence comprehension: effects of working memory and speed of processing. NeuroImage. 2003;19:101–112. doi: 10.1016/s1053-8119(03)00007-7. [DOI] [PubMed] [Google Scholar]