

Abstract
Delivery of factors capable of promoting oligodendrocyte precursor cell (OPC) survival and differentiation in vivo is an important therapeutic strategy for a variety of pathologies in which demyelination is a component, including multiple sclerosis and spinal cord injury. Ciliary neurotrophic factor (CNTF) is a neuropoietic cytokine that promotes both survival and maturation of a variety of neuronal and glial cell populations, including oligodendrocytes. Present results suggest that although CNTF has a potent survival and differentiation promoting effect in vitro on OPCs isolated from the adult spinal cord, CNTF administration in vivo is not sufficient to promote oligodendrocyte remyelination in the glial-depleted environment of unilateral ethidium bromide (EB) lesions.
Keywords: CNTF, demyelination, remyelination, oligodendrocyte precursor cell
Identification of signals responsible for regulating the process of remyelination in vivo is critical to the development of therapies aimed at promoting remyelination. The EB model of chemical demyelination is an excellent model to study the potential of exogenously administered growth factors to directly promote oligodendrocyte survival and remyelination (Blakemore et al., 1989; Woodruff et al., 1999). Despite the successful recruitment of Nkx2.2+/Olig2+ OPCs within EB lesions, spontaneous oligodendrocyte remyelination by endogenous OPCs is not efficient (Talbott et al., 2005). Similarly, remyelination failure by endogenous OPCs is observed in chronic MS lesions (Chang et al., 2000; Chang et al., 2002). The identification and delivery of factors that can promote the survival and differentiation of endogenous OPCs in vivo will potentially lead to therapies for patients suffering from demyelinating diseases.
CNTF isolated from the chick eye was originally shown to promote the survival of ciliary ganglionic neurons (Barbin et al., 1984). Subsequent studies demonstrated a protective effect for CNTF on a variety of CNS and PNS neural subtypes (Hagg et al., 1993; Naumann et al., 2003). In vitro, CNTF has been shown to promote the differentiation and survival of peri-natal OPCs, suggesting a potential therapeutic role for CNTF in promoting remyelination by endogenous oligodendrocytes (Louis et al., 1993; Mayer et al., 1994; Stankoff et al., 2002). However, OPCs derived from peri-natal and adult animals demonstrate fundamental differences in rates of division, differentiation, migration and response to growth factors (Wolswijk et al., 1989; Chan et al., 1990; Engel et al., 1996). Therefore, it is important to directly test the effects of CNTF on adult OPCs as these cells are the primary target for CNTF therapy in vivo.
To determine if CNTF could promote the survival of OPCs derived from the adult rat spinal cord, adult OPC cultures were generated as previously described (Talbott et al., 2006). Adult OPCs were induced to differentiate into O1 oligodendrocytes by withdrawal of FGF2 (Chemicon, Temecula, CA) and PDGF-AA (Sigma, St. Louis, MO ) for two days. Then, the differentiated oligodendrocytes continued to grow in basal medium (DMEM/F12 + 1 × N2 + 1 × B27 + 0.1% BSA) alone or basal media with CNTF (10 ng/ml; Sigma, St. Louis, MO) for five more days. Cell survival was assessed using an MTT colorimetric assay according to the manufacturer’s instructions (Chemicon, Temecula, CA).
To measure OPC differentiation in vitro with CNTF, adult OPCs were cultured in either basal media + 5 ng/ml FGF2 (control) or basal media + 5ng/ml FGF2 + 10 ng/ml CNTF. FGF2 was added to both groups to minimize spontaneous OPC differentiation. After 5 days of treatment, cell preparations were fixed and stained with antibodies for the mature oligodendrocyte marker O1 (Undiluted-hybridoma; ATCC, Manassas, VA). For in vitro quantification, ten fields at 20X power were randomly selected by a blinded observer from each of at least 3 different experimental trials. The mean cell counts from each of the three trials were then averaged and independent t-tests were performed to analyze for statistical significance.
For all animal experiments, surgical interventions and peri-operative care were provided in strict accordance with the Public Health Service Policy on Humane Care and Use of Laboratory Animals and were approved by the University of Louisville Institutional Animal Care and Use Committee. For a summary of experimental animal groups see table 1.
Table 1.
Experimental groups.
| Group 1 | Group 2 | Group 3 | |
|---|---|---|---|
| Day 0 | EB injection | EB injection | EB injection |
| Days 2–5 | BrdU Pulse | ||
| Day 7 | Nanosphere injection | Nanosphere injection | Nanosphere injection |
| Day 9 | Sacrifice | ||
| Day 14 | Sacrifice | ||
| Day 35 | Sacrifice |
Group 1: Animals evaluated for astrogliosis in contralateral VLF following CNTF (n=4) and blank (n=4) nanosphere injection.
Group 2: Animals evaluated for survival of endogenous BrdU-pulsed OPCs following CNTF (n=4) and blank (n=4) nanosphere injection.
Group 3: Animals evaluated for presence of mature oligodendrocytes following CNTF (n=4) and blank (n=4) nanosphere injection into EB lesions.
Group 1
Seven days after induction of focal, unilateral EB demyelinating lesions in the ventrolateral funiculus (VLF) of the rat thoracic spinal cord (for methods for VLF lesion induction see Talbott et al., 2005), human CNTF (R&D, Minneapolis, MN) was delivered via CNTF-loaded poly-lactide-co-glycolide (PLGA) nanospheres made from a spontaneous emulsion technique (Fu et al., 2003). A total of 6 μL of CNTF-loaded nanospheres (25 mg/mL) in sterile saline were stereotactically injected directly into EB lesions with coordinates identical to those used for EB injection. For controls, equal concentrations of BSA-containing blank nanospheres were identically injected into EB lesions. Nine days after EB injection, animals were sacrificed. To assess in vivo bioactivity of nanosphere-derived CNTF, GFAP immunoreactivity was measured in the gray matter contralateral to transplanted EB lesions. Spinal cord specimens for immunohistochemistry were prepared as previously described (Talbott et al., 2005). Image Pro Plus software (Media Cybernetics, MO) was used as previously described (Loy et al., 2002) to quantify GFAP immunoreactivity in a predefined region of interest (ROI) in the contralateral gray matter (ROI approximated in Fig 1D). All images for quantification were obtained with identical capture parameters using the Nikon Eclipse TE300 microscope and photographed with a Spot RT Color CCD camera. The ROI for GFAP quantification was defined by a vertical 250 μm × 300 μm rectangle with the ventral-most border centered at the most ventral region of the contralateral ventral horn. For all in vivo quantitative immunohistochemical analysis, at least 4 epicenter-derived sections (approximately 60 μm apart) from each of 4 animals were analyzed.
Figure 1.
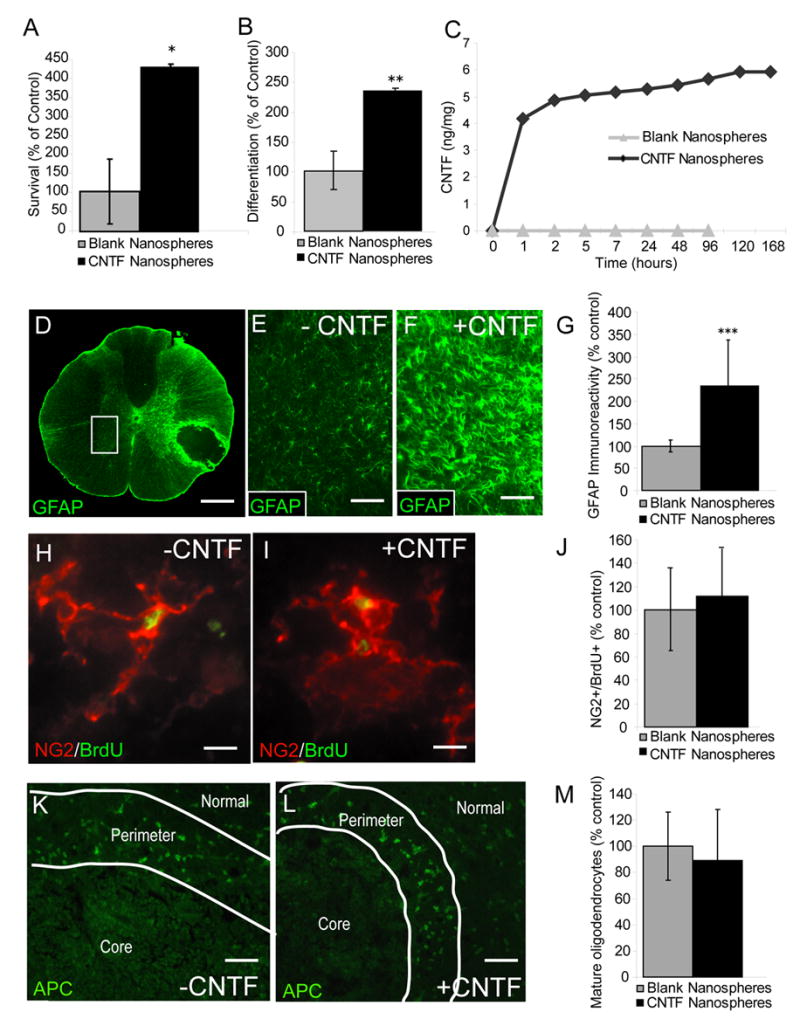
CNTF promotes the survival and differentiation of adult OPCs in vitro but not in vivo. A, The survival of newly differentiated adult OPCs was assessed using the MTT assay. Adult OPCs were differentiated for two days by withdrawal of FGF2 and PDGF-AAaa, then continued to grow in basal medium with or without CNTF (10 ng/ml) for five more days. Quantitative analysis of surviving MTT+ cells demonstrated an approximate 300% increase in survival for cells treated with CNTF relative to control (*p = 0.03, t = 4.3, df = 2). B, Adult OPCs were cultured in either basal media + 5 ng/ml FGF2 (control) or basal media + 5 ng/ml FGF2 + 10 ng/ml CNTF. FGF2 was added to both groups to minimize spontaneous OPC differentiation. As measured by expression of the more mature oligodendrocyte marker O1, CNTF treated cultures demonstrate significantly increased differentiation relative to control (**p = 0.003, t = 2.7, df = 4). Data are expressed as the mean ± standard deviation. C, CNTF-loaded nanospheres (black diamonds) release CNTF in a sustained fashion in vitro for up to 168 hours (latest time point tested) while blank nanospheres (gray triangles) release no detectable CNTF. At each time point indicated, media was completely removed and protein quantity was assessed using an ELISA kit specific for recombinant human CNTF (rhCNTF) (expressed as ng of CNTF protein per mg of CNTF-loaded nanospheres). D, GFAP immunoreactivity was assessed in the gray matter (white box) contralateral to transplanted EB lesions 7 days after nanosphere injection (6 μl of nanospheres at 25 μg/μl). Compared to blank nanospheres (E), CNTF-nanosphere (F) injection into EB lesions of the ventrolateral funiculus (VLF) dramatically increases GFAP immunoreactivity in the contralateral gray matter. The area shown in E and F corresponds to a high magnification view of the region delineated by the white box in D. G, Quantitative pixel counts of GFAP immunoreactivity 9 days after nanosphere injection demonstrates a 130% increase in the contralateral gray matter of CNTF-nanosphere treated lesions compared to blank nanospheres (*** p < 0.001, t = 2.1, df = 20). EB lesions treated with blank (H) or CNTF-loaded (I) nanospheres (6 μl of nanospheres at 25 μg/μl) 5 days after EB injection contain similar densities of NG2+/BrdU+ cells at 14 days after EB injection (J, p = 0.54, t = 0.62, df = 16). BrdU pulse labeling (100 mg/kg every twelve hours) was performed during days 2, 3, and 4 post EB injection. K-M, Quantitative analysis of APC/CC-1+ profiles in the perimeter of (K) blank and (L) CNTF-nanosphere treated lesions was performed 4 weeks after nanosphere injection (M, p = 0.45, t = 2.1, df = 18). A,B,F,I,L, The mean ± standard deviation are expressed as the percentage of control where the average density of positive profiles in blank nanosphere-injected lesions equals 100%. Independent t-tests were performed for all statistical analysis. Scale bar = 300 μm in (D); 75 μm in (E,F); 65 μm in (H,I).
Group 2
BrdU pulse-labeling and dual-label immunohistochemistry were performed to determine the effect of CNTF on endogenous OPCs. To label proliferating OPCs, rats were pulse-labeled with BrdU (100 mg/kg in sterile saline every 12 hours) for 72 hrs beginning two days after ethidium bromide injection. This is the time point at which maximal OPC proliferation was previously seen after EB spinal cord lesions (Talbott et al., 2005). Two days following the BrdU pulse, EB lesions were injected with either CNTF-loaded or blank nanospheres. Two weeks after EB injection the number of surviving BrdU+ and BrdU+/NG2+ cells within CNTF-nanosphere and control injected lesions was determined. Manual identification of NG2+/BrdU+ cells for quantification was performed as previously described (McTigue et al., 2001, Talbott et al., 2005).
Group 3
To determine if CNTF could enhance endogenous oligodendrocyte remyelination in vivo, CNTF-loaded or blank nanospheres were injected into EB lesions 7 days after lesion induction and animals were sacrificed four weeks later. APC+ cell density was manually determined in the periphery of EB lesions using methods as described for group 2.
For both in vitro and in vivo studies, protein quantity was assessed using an ELISA kit (R&D Systems, Minneapolis, MN) for human CNTF. Results are expressed as ng of CNTF protein per mg of CNTF-loaded nanospheres for in vitro studies and ng of CNTF per mg of total protein for in vivo studies. In vitro, spheres release 1–5 ng CNTF/mg spheres/hr for up to 168 hrs (the longest time point tested, Fig 1C). To quantify CNTF release in vivo at seven days after CNTF injection, spinal cords were collected following perfusion with 200 ml of chilled saline. A five mm segment of spinal cord containing the injection site was dissected and processed with ELISA. Total protein concentrations from spinal samples were measured with a BCA protein assay kit (Pierce, Rockford, IL).
For immunohistochemical analysis, the following primary antibodies were used; NG2 (1:200; Chemicon), BrdU (1:100; Biodesign), GFAP (1:200; Dako), APC/CC-1 (1:100; Oncogene), and ED-1 (1:800; Chemicon). Appropriate secondary antibodies were applied for 90 minutes at room temperature. The following species-specific secondary antibodies were used: donkey anti-mouse fluoroisothiocyanate (FITC)-conjugated (1:100), donkey anti-rabbit rhodamine red (1:200) and FITC-conjugated Fab’ fragments (1:100), and donkey anti-sheep FITC-conjugated IgG (1:100). All secondary antibodies were supplied by Jackson Immunoresearch Lab (Baltimore, MD).
CNTF-treatment in vitro resulted in a significant increase in the number of surviving oligodendrocytes as assayed by MTT reactivity (p = 0.03, t = 4.3, df = 2; Fig. 1A). The resulting increase in surviving cells following CNTF treatment did not result from increased proliferation as evidenced by a lack of BrdU incorporation by differentiated cells (data not shown). In addition to promoting adult OPC survival, CNTF administration to adult OPC cultures promotes adult OPC differentiation as evidenced by the significantly increased number of more differentiated, O1+ oligodendrocytes in CNTF-treated cultures as compared to untreated control cultures (p = 0.003, t = 2.7, df = 4; Fig 1B). Thus, as with peri-natal OPCs, CNTF has a robust survival and differentiation-promoting effect on adult spinal cord-derived OPCs in vitro.
CNTF has been previously shown to induce robust astrogliosis (Winter et al., 1995; Levison et al., 1998). CNTF-nanosphere injection resulted in a dramatic astrogliosis extending to the contralateral gray matter as evidenced by a significant 130% increase in GFAP immunoreactivity compared to blank nanospheres (p < 0.001, t = 2.1, df = 20; Fig. 2D–G). Astrogliosis was not due to an increased inflammatory response to CNTF-loaded nanospheres as evidenced by a similar density of ED-1+ macrophages/activated microglia within CNTF and blank nanosphere-injected lesions (94 ± 20% of ED-1 immunoreactivity in controls versus CNTF injected lesions, p = 0.49, t = 2.1, df = 18). These results provide evidence for specific bioactivity by nanosphere-derived CNTF in vivo. In CNTF nanosphere-injected lesions, levels of CNTF were 0.141 ± 0.014 ng CNTF per mg of protein (n=4), compared to 0.040 ± 0.013 ng in the control animals (n=2), further confirming CNTF release by engrafted nanospheres.
Endogenous OPCs proliferate maximally between 2 and 5 days after induction of EB lesions (Talbott et al., 2005). Both CNTF and blank nanosphere-injected lesions contained BrdU-labeled cells after two weeks. CNTF-nanosphere injection did not result in a significant change in the number of BrdU-labeled cells relative to blank nanosphere injected controls (83 ± 33% of control; p = 0.28, t = 1.12, df = 14). Further, dual-label immunohistochemistry for BrdU and the OPC marker NG2 showed that CNTF-nanosphere injection did not result in a relative increase in the number of surviving NG2+/BrdU+ cells after 2 weeks (112 ± 40% of control; p = 0.54, t = 0.62, df = 16; Fig. 2H–J).
Immunohistochemical analysis for the mature oligodendrocyte marker APC/CC-1 revealed only occasional oligodendrocytes in the central core of both blank and CNTF-treated lesions four weeks after nanosphere injection. Quantification of oligodendrocyte density in the perimeter of EB lesions did not reveal a difference between CNTF and control groups (89 ± 26% of control; p = 0.45, t = 2.1, df = 18; Fig. 2K–M).
Despite positive in vitro results and the confirmed delivery of bioactive CNTF into EB demyelinating lesions, neither survival nor differentiation of endogenous OPCs was enhanced in vivo by exogenously administered CNTF. Translating positive in vitro results with single growth factors into the complex in vivo environment of the demyelinated spinal cord has proven difficult. Previous studies have demonstrated that, in isolation, the exogenous delivery of neuregulin (Penderis et al., 2003), insulin-like growth factor 1 (IGF-1) (O’Leary et al., 2002), and platelet derived growth factor (PDGF) (Woodruff et al., 2004) fails to promote remyelination following EB-induced demyelination. Linker et al. (2002) has shown that oligodendrocyte pathology is severely exacerbated in CNTF−/− mice following myelin oligodendrocyte glycoprotein (MOG)-induced experimental autoimmune encephalomyelitis (EAE). This phenotype in CNTF−/− mice could be reversed by treatment with antiserum to TNF-α, suggesting that the protective effects of CNTF were mediated through modulation of the inflammatory response as opposed to direct effects of CNTF on oligodendrocyte survival and maturation. Likewise, Kuhlmann et al. (2006) recently showed that systemic administration of CNTF ameliorates EAE pathology primarily by abrogating central nervous system (CNS) inflammation and blood brain barrier breakdown. In that study, the potential neuroprotective and pro-remyelinating properties of CNTF were not directly addressed, but the complete absence of inflammation in high dose CNTF treated animals suggests that the protective mechanism was primarily mediated by CNTF’s suppression of T-cell infiltration into the CNS rather than direct effects by CNTF on OPC survival and remyelination. Using the EB model of demyelination, Penderis et al. (2003) has shown that the previously demonstrated pro-remyelinating effect of neuregulin (GGF-2) in EAE (Cannella et al., 1998) is not replicated in the non-immune mediated EB model of demyelination. Likewise, present results suggest that direct promotion of oligodendrocyte remyelination by local CNTF delivery in vivo does not occur in the absence of a primary neuroinflammatory insult. Collectively, these results suggest that the beneficial effect of CNTF on in vivo demyelinating injury models are not attributable to its direct promotion of OPC survival and differentiation despite the demonstration of these properties in vitro. Multiple studies have revealed that single growth factor delivery has little effect on altering remyelination in vivo. Numerous factors interact in a coordinated manner to orchestrate the complex process of remyelination. Thus, future attempts at promoting remyelination will likely depend upon timely combinatorial therapies and/or identification of molecules that regulate many downstream factors responsible for successful remyelination.
Acknowledgments
This research was supported by NS38665, RR15576, the Kentucky Spinal Cord and Head Injury Research Trust, Norton Healthcare, and the Commonwealth of Kentucky Research Challenge for Excellence Trust Fund.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Reference List
- Barbin G, Manthorpe M, Varon S. Purification of the chick eye ciliary neuronotrophic factor. J Neurochem. 1984;43:1468–1478. doi: 10.1111/j.1471-4159.1984.tb05410.x. [DOI] [PubMed] [Google Scholar]
- Blakemore WF, Crang AJ. The relationship between type-1 astrocytes, Schwann cells and oligodendrocytes following transplantation of glial cell cultures into demyelinating lesions in the adult rat spinal cord. J Neurocytol. 1989;18:519–528. doi: 10.1007/BF01474547. [DOI] [PubMed] [Google Scholar]
- Cannella B, Hoban CJ, Gao YL, Garcia-Arenas R, Lawson D, Marchionni M, Gwynne D, Raine CS. The neuregulin, glial growth factor 2, diminishes autoimmune demyelination and enhances remyelination in a chronic relapsing model for multiple sclerosis. Proc Natl Acad Sci U S A. 1998;95:10100–10105. doi: 10.1073/pnas.95.17.10100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan CL, Wigley CB, Berry M. Oligodendrocyte-type 2 astrocyte (O-2A) progenitor cells from neonatal and adult rat optic nerve differ in their responsiveness to platelet-derived growth factor. Brain Res Dev Brain Res. 1990;55:275–282. doi: 10.1016/0165-3806(90)90209-h. [DOI] [PubMed] [Google Scholar]
- Chang A, Nishiyama A, Peterson J, Prineas J, Trapp BD. NG2-positive oligodendrocyte progenitor cells in adult human brain and multiple sclerosis lesions. J Neurosci. 2000;20:6404–6412. doi: 10.1523/JNEUROSCI.20-17-06404.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang A, Tourtellotte WW, Rudick R, Trapp BD. Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. N Engl J Med. 2002;346:165–173. doi: 10.1056/NEJMoa010994. [DOI] [PubMed] [Google Scholar]
- Engel U, Wolswijk G. Oligodendrocyte-type-2 astrocyte (O-2A) progenitor cells derived from adult rat spinal cord: in vitro characteristics and response to PDGF, bFGF and NT-3. Glia. 1996;16:16–26. doi: 10.1002/(SICI)1098-1136(199601)16:1<16::AID-GLIA3>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- Fu K, Harrell R, Zinski K, Um C, Jaklenec A, Frazier J, Lotan N, Burke P, Klibanov AM, Langer R. A potential approach for decreasing the burst effect of protein from PLGA micropsheres. J Pharm Sci. 2003;92:1582–1591. doi: 10.1002/jps.10414. [DOI] [PubMed] [Google Scholar]
- Hagg T, Varon S. Ciliary neurotrophic factor prevents degeneration of adult rat substantia nigra dopaminergic neurons in vivo. Proc Natl Acad Sci U S A. 1993;90:6315–6319. doi: 10.1073/pnas.90.13.6315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuhlmann T, Remington L, Cognet I, Bourbonniere L, Zehntner S, Guilhot F, Herman A, Guay-Giroux A, Antel JP, Owens T, Gauchat JF. Continued administration of ciliary neurotrophic factor protects mice from inflammatory pathology in experimental autoimmune encephalomyelitis. Am J Pathol. 2006;169:584–598. doi: 10.2353/ajpath.2006.051086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levison SW, Hudgins SN, Crawford JL. Ciliary neurotrophic factor stimulates nuclear hypertrophy and increases the GFAP content of cultured astrocytes. Brain Res. 1998;803:189–193. doi: 10.1016/s0006-8993(98)00594-0. [DOI] [PubMed] [Google Scholar]
- Linker RA, Maurer M, Gaupp S, Martini R, Holtmann B, Giess R, Rieckmann P, Lassmann H, Toyka KV, Sendtner M, Gold R. CNTF is a major protective factor in demyelinating CNS disease: a neurotrophic cytokine as modulator in neuroinflammation. Nat Med. 2002;8:620–624. doi: 10.1038/nm0602-620. [DOI] [PubMed] [Google Scholar]
- Louis JC, Magal E, Takayama S, Varon S. CNTF protection of oligodendrocytes against natural and tumor necrosis factor-induced death. Science. 1993;259:689–692. doi: 10.1126/science.8430320. [DOI] [PubMed] [Google Scholar]
- Loy DN, Crawford CH, Darnall JB, Burke DA, Onifer SM, Whittemore SR. Temporal progression of angiogenesis and basal lamina deposition after contusive spinal cord injury in the adult rat. J Comp Neurol. 2002;445:308–324. doi: 10.1002/cne.10168. [DOI] [PubMed] [Google Scholar]
- McTigue DM, Wei P, Stokes BT. Proliferation of NG2-positive cells and altered oligodendrocyte numbers in the contused rat spinal cord. J Neurosci. 2001;21:3392–3400. doi: 10.1523/JNEUROSCI.21-10-03392.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer M, Bhakoo K, Noble M. Ciliary neurotrophic factor and leukemia inhibitory factor promote the generation, maturation and survival of oligodendrocytes in vitro. Development. 1994;120:143–153. doi: 10.1242/dev.120.1.143. [DOI] [PubMed] [Google Scholar]
- Naumann T, Schnell O, Zhi Q, Kirsch M, Schubert KO, Sendtner M, Hofmann HD. Endogenous ciliary neurotrophic factor protects GABAergic, but not cholinergic, septohippocampal neurons following fimbria-fornix transection. Brain Pathol. 2003;13:309–321. doi: 10.1111/j.1750-3639.2003.tb00030.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Leary MT, Hinks GL, Charlton HM, Franklin RJ. Increasing local levels of IGF-I mRNA expression using adenoviral vectors does not alter oligodendrocyte remyelination in the CNS of aged rats. Mol Cell Neurosci. 2002;19:32–42. doi: 10.1006/mcne.2001.1062. [DOI] [PubMed] [Google Scholar]
- Penderis J, Woodruff RH, Lakatos A, Li WW, Dunning MD, Zhao C, Marchionni M, Franklin RJ. Increasing local levels of neuregulin (glial growth factor-2) by direct infusion into areas of demyelination does not alter remyelination in the rat CNS. Eur J Neurosci. 2003;18:2253–2264. doi: 10.1046/j.1460-9568.2003.02969.x. [DOI] [PubMed] [Google Scholar]
- Stankoff B, Aigrot MS, Noel F, Wattilliaux A, Zalc B, Lubetzki C. Ciliary neurotrophic factor (CNTF) enhances myelin formation: a novel role for CNTF and CNTF-related molecules. J Neurosci. 2002;22:9221–9227. doi: 10.1523/JNEUROSCI.22-21-09221.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talbott JF, Loy DN, Liu Y, Qiu MS, Bunge MB, Rao MS, Whittemore SR. Endogenous Nkx2.2+/Olig2+ oligodendrocyte precursor cells fail to remyelinate the demyelinated adult rat spinal cord in the absence of astrocytes. Exp Neurol. 2005;192:11–24. doi: 10.1016/j.expneurol.2004.05.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talbott JF, Cao Q, Enzmann GU, Benton RL, Achim V, Cheng XX, Mills MD, Rao MS, Whittemore SR. Schwann cell-like differentiation by adult oligodendrocyte precursor cells following engraftment into the demyelinated spinal cord is BMP-dependent. Glia. 2006;54:147–159. doi: 10.1002/glia.20369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winter CG, Saotome Y, Levison SW, Hirsh D. A role for ciliary neurotrophic factor as an inducer of reactive gliosis, the glial response to central nervous system injury. Proc Natl Acad Sci. 1995;92:5865–5869. doi: 10.1073/pnas.92.13.5865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolswijk G, Noble M. Identification of an adult-specific glial progenitor cell. Development. 1989;105:387–400. doi: 10.1242/dev.105.2.387. [DOI] [PubMed] [Google Scholar]
- Woodruff RH, Franklin RJ. Demyelination and remyelination of the caudal cerebellar peduncle of adult rats following stereotaxic injections of lysolecithin, ethidium bromide, and complement/anti-galactocerebroside: a comparative study. Glia. 1999;25:216–228. doi: 10.1002/(sici)1098-1136(19990201)25:3<216::aid-glia2>3.0.co;2-l. [DOI] [PubMed] [Google Scholar]
- Woodruff RH, Fruttiger M, Richardson WD, Franklin RJ. Platelet-derived growth factor regulates oligodendrocyte progenitor numbers in adult CNS and their response following CNS demyelination. Mol Cell Neurosci. 2004;25:252–262. doi: 10.1016/j.mcn.2003.10.014. [DOI] [PubMed] [Google Scholar]