

Abstract
We previously provided evidence that the protein encoded by the highly conserved skb1 gene is a putative regulator of Shk1, a p21Cdc42/Rac-activated kinase (PAK) homolog in the fission yeast Schizosaccharomyces pombe. skb1 null mutants are viable and competent for mating but less elongate than wild-type S. pombe cells, whereas cells that overexpress skb1 are hyperelongated. These phenotypes suggest a possible role for Skb1 as a mitotic inhibitor. Here we show genetic interactions of both skb1 and shk1 with genes encoding key mitotic regulators in S. pombe. Our results indicate that Skb1 negatively regulates mitosis by a mechanism that is independent of the Cdc2-activating phosphatase Cdc25 but that is at least partially dependent on Shk1 and the Cdc2 inhibitory kinase Wee1. We provide biochemical evidence for association of Skb1 and Shk1 with Cdc2 in S. pombe, suggesting that Skb1 and Shk1 inhibit mitosis through interaction with the Cdc2 complex, rather than by an indirect mechanism. These results provide evidence of a previously undescribed role for PAK-related protein kinases as mitotic inhibitors. We also describe the cloning of a human homolog of skb1, SKB1Hs, and show that it can functionally replace skb1 in S. pombe. Thus, the molecular functions of Skb1-related proteins have likely been substantially conserved through evolution.
p21Cdc42/Rac-activated kinases (PAKs) comprise a recently recognized family of protein kinases that are highly conserved in eukaryotes (1). PAKs bind to, and in many cases have been shown to be activated by, the small GTPases Cdc42 and Rac, but do not bind to other small GTPases such as Ras or Rho. Studies in model systems ranging from yeasts to mammalian cells have demonstrated diverse roles for PAKs, including regulation of mitogen-activated protein kinase cascades (2–6), cytoskeletal and morphological regulation (3, 7–9), and induction of apoptosis (10). In the fission yeast Schizosaccharomyces pombe, the PAK homolog Shk1 (also called Pak1) is essential for cell viability and is implicated by genetic analyses to play key roles in morphological control and mating response pathways regulated by Ras and Cdc42 GTPases (3, 7). Cdc42 binds to a sequence in the Shk1 regulatory domain that is highly homologous to the Cdc42/Rac binding sites found in the regulatory domains of mammalian PAKs (11, 12). The terminal phenotypes of cdc42 and shk1 null mutants are similar, with each resulting in growth arrest as small round cells (3, 7, 13). Whereas genetic experiments suggest that Shk1 is a key effector for Cdc42 in fission yeast (3, 7), overexpression of shk1 does not suppress the lethality resulting from the cdc42 null mutation (7), raising the possibility that Cdc42 may have at least one other essential effector besides Shk1 (7). The molecular targets of Shk1 have not yet been described.
Previously, we reported on the characterization of a novel gene, skb1, which was identified by a two-hybrid screen for cDNAs encoding proteins that form complexes with Shk1 (11). The Skb1 protein interacts with a regulatory subdomain of Shk1 positioned between its Cdc42-binding and kinase domains, thus implicating Skb1 as a Shk1 regulator. Unlike cdc42 and shk1, the skb1 gene is not essential for viability or mating of S. pombe cells (11). However, skb1 null (skb1Δ) mutants are less elongate and exhibit a slightly slower growth rate than wild-type cells. In contrast, cells overexpressing skb1 are hyperelongated in morphology. Overexpression of shk1 restores elongate morphology to the skb1Δ mutant, suggesting that Shk1 functions downstream of Skb1 (11). Overexpression of skb1 partially restores elongate morphology to fission yeast cells carrying a ras1 null mutation, and this effect is markedly increased by cooverexpressing skb1 with shk1 (11). Cumulatively, these data suggest that Skb1 functions to positively modulate Shk1 function in vivo.
Interestingly, the cell length defects resulting from loss and gain of skb1 function bear resemblance to defects resulting from loss and gain of function of certain genes, such as wee1, encoding dosage-dependent mitotic inhibitors in S. pombe (14–16). As in higher organisms, the initiation of mitosis in fission yeast is dependent on the activity of a cyclin-dependent kinase, Cdc2, and an associated B-type cyclin, Cdc13 (15, 17). Cdc13 is destroyed after completion of mitosis and accumulates during interphase. Cdc2⋅Cdc13 activity is repressed during interphase as a result of inhibitory phosphorylation of Cdc2 on tyrosine 15 by the related protein kinases Wee1 and Mik1 (18, 19). S. pombe cells carrying a wee1 deletion are viable but exit prematurely from G2 at a reduced size, whereas overexpression of wee1 delays G2 to M progression, causing cells to divide at significantly greater than normal lengths (14–16). The mik1Δ mutation has little effect on the timing of mitosis, but deletion of both wee1 and mik1 is synthetically lethal and results in a condition termed lethal mitosis, in which cells divide at a very small size and exhibit abnormal patterns of chromosome segregation (19). Transition from G2 to M phase requires that Cdc2 undergo phosphorylation on threonine 167 by a Cdc2-activating kinase, or CAK (20, 21) and dephosphorylation of tyrosine 15, which occurs primarily through the activity of the Cdc2-activating phosphatase Cdc25 (22, 23). Loss of Cdc25 function causes cells to become blocked at G2 while continuing to accumulate mass and becoming extremely elongated, whereas overexpression of Cdc25, like loss of Wee1, causes cells to exit from G2 at a reduced size (15, 22). Thus Cdc25, like Wee1, functions in a dosage-dependent fashion.
In the present study, we have investigated the cell-cycle regulatory properties of Skb1 and Shk1. We show genetic interactions of the skb1 and shk1 genes with genes encoding key mitotic regulators in S. pombe that demonstrate a previously undescribed role for the Skb1 and Shk1 proteins as dosage-dependent mitotic inhibitors. We also show that Cdc2 coprecipitates with Skb1 and Shk1 proteins isolated from S. pombe cell lysates, suggesting that Skb1 and Shk1 inhibit mitosis through interaction with the Cdc2 complex rather that by an indirect mechanism. Finally, we provide evidence that Skb1 protein function has been substantially conserved through evolution by showing that a human homolog of skb1, SKB1Hs, can substitute for skb1 in S. pombe.
MATERIALS AND METHODS
Yeast Strains, Media, and Plasmids.
Wild-type S. pombe strains used in this study were CHP428 (h+ ade6-M210 his7–366 leu1–32 ura4-D18) (E. Chang, New York University) and SP870 (h90 ade6-M210 leu1–32 ura4-D18) (D. Beach, Cold Spring Harbor Laboratory). Mutant S. pombe strains used were 167 (h+ leu1–32 ura4-D18 cdc25–22), 137 (h− leu1–32 ura4-D18 wee1∷ura4), and 41 (h− ade6–216 leu1–32 cdc2–3w) (S. Sazer, Baylor College of Medicine); 606 (h− leu1–32 ura4-D18 cdc2–1w) (K. Gould, Vanderbilt University); GL192 (h+ leu1–32 ura4-D18 cdc2–3w cdc25∷ura4) (P. Russell, Scripps Research Institute); SMPY101 and SMMG100 (h+ ade6–210 his7–366 leu1–32 ura4-D18 skb1∷ura4), SMMG102 (h+ ade6–216 his7–366 leu1–32 ura4-D18 cdc25–22 skb1∷ura4), SMMG103 (h90 ade6-M216 leu1–32 ura4-D18 cdc25–22), SMMG115 (h− ade6-M210 leu1–32 ura4-D18 wee1∷ura4∷ADE2), and the cdc25–22 nmt-shk1 strain SMMG127 (h90 ade6-M210 leu1–32 ura4-D18 cdc25–22 shk1∷ura4∷ADE2 pREP81Shk1[XS]). Cell lengths at division measurements were determined as described (24), by using a phototube micrometer attached to an Olympus B-Max 60 microscope (Olympus, New Hyde Park, NY). At least 20 cells from each culture were measured. Standard yeast genetic methods were performed for construction of S. pombe mutants for this study (25). Cultures were grown on either rich medium (YEA) or synthetic minimal medium (EMM) with appropriate auxotrophic supplements (25).
pREP1 (26), pREP81 (27), pREP1Skb1 (for overexpression of skb1 from the thiamine-repressible nmt1 promoter) (11), pAAUCM (3), pAAUCMSkb1 (for overexpression of skb1 from the adh1 promoter) (11), and pART1CM (28) have been described. pAAUCMShk1 and pART1CMShk1 were constructed by PCR-mediated cloning of a BamHI-KpnI fragment of the full-length shk1 coding sequence into pAAUCM (ura4-based) and pART1CM (LEU2-based) plasmids, respectively, and allow for expression of shk1 as a c-myc epitope-tagged protein from the strong S. pombe adh1 promoter. pREP81Shk1[XS] was constructed by cloning a XhoI-SspI fragment of the shk1 gene into the plasmid pREP81, a LEU2-based plasmid containing a mutationally attenuated version of the nmt1 promoter (27). The XhoI-SspI fragment of shk1 is deleted of the 5′ end of the shk1 coding sequence and codes for a Shk1 protein truncated by the deletion of the N-terminal 118 amino acids of the full-length Shk1 protein. This N-terminal truncated Shk1 mutant is nonetheless capable of fully complementing the shk1 null mutation when expressed from pREP81. pAAUGST was constructed by ligating a 0.7-kb SalI/PstI-digested, PCR-amplified glutathione S-transferase (GST) gene fragment to the corresponding sites of pAAU, a ura4-based plasmid similar to pART1 (29), which allows for expression of sequences from the S. pombe adh1 promoter. pAAUGSTSkb1 was constructed by ligating a BamHI-KpnI skb1 fragment from pGADSkb1ΔN24 into pAAUGST. pAAUGSTShk1 was constructed by PCR-mediated subcloning of the full-length shk1 coding sequence into pAAUGST.
Preparation of Cell Lysates and Coprecipitation Experiments.
S. pombe CHP428 transformants expressing GST fusion proteins were grown to about 107 cells per ml in EMM, harvested by centrifugation, resuspended with yeast lysis buffer [20 mM Hepes (pH 7.6)/200 mM KCl/2 mM EGTA/2 mM EDTA/10 mM sodium molybdate/50 mM sodium fluoride/2 mM sodium pyrophosphate/1 mM sodium vanadate/0.1% Nonidet P-40/10% glycerol/10 μM E64/100 μM leupeptin/1 μM pepstatin A/1 mM phenylmethanesulfonyl fluoride/aprotinin (2 μg/ml)] and ground with glass beads. Crude lysates were clarified twice by centrifugation at 16,000 × g for 15 min and the supernatant fractions were aliquoted and quick-frozen in liquid nitrogen before being stored at −80°C. The supernatant fractions (1 mg of protein) were incubated with 50 μl of a 50% slurry of glutathione agarose beads in a volume of 1 ml yeast lysis buffer for 2 hr at 4°C on a rotary rocker. The beads were washed four times with yeast lysis buffer and bound proteins were separated by SDS/PAGE and transferred to nitrocellulose membranes. Western blot analyses were performed by using anti-Cdc2 PSTAIRE (Santa Cruz Biotechnology), anti-GST (Pierce), and anti-Skb1 (to be described elsewhere) polyclonal antibodies.
SKB1Hs cDNA Cloning.
The oligonucleotide primers 5′-GAATGGGATCCCCAAGTGACCGTAGTCTCATCA and 5′-AGATGAATTCCACCTTCTGTACTACAGGAGCAG were used in PCRs to amplify a fragment of SKB1Hs from a HeLa cell cDNA library constructed in the plasmid pEBS7 (kindly provided by R. Legerski, University of Texas M. D. Anderson Cancer Center, Houston, TX). The partial SKB1Hs cDNA was used as a probe to screen for the full-length cDNA from the pEBS7 HeLa cell library. Two potential full-length SKB1Hs cDNAs were isolated from a screen of about 1.5 million bacterial transformants. A search of the expressed sequence tag databases revealed a clone (GenBank accession no. AA417623) containing an in-frame stop codon 5′ of our predicted SKB1Hs translational initiation codon. PCR was used to facilitate cloning of the full-length SKB1Hs cDNA into the pREP1 and pAAUGST plasmids for expression in S. pombe.
RESULTS
Skb1 Is a Dosage-Dependent Mitotic Inhibitor.
Cell length measurements showed that S. pombe skb1Δ cells divide at about <90% the length of wild-type cells, whereas cells overexpressing skb1 divide at ≈50% greater than normal length (Table 1). These observations suggested the possibility that Skb1 is a dosage-dependent mitotic inhibitor. To explore further this possibility, we tested for genetic interactions between the skb1 gene and genes encoding key mitotic regulators in S. pombe. The temperature-sensitive (t.s.) cdc25–22 mutation causes S. pombe cells to arrest at G2 of the cell cycle when they are shifted to the restrictive temperature of 36°C (14, 30). We predicted that if Skb1 is indeed a mitotic inhibitor, then loss of skb1 gene function might suppress loss of cdc25 function. We constructed a cdc25–22 skb1Δ mutant and compared its growth characteristics to that of the cdc25–22 mutant at the normally nonpermissive temperature of 36°C and at the semipermissive temperature of 33°C. The cdc25–22 mutant produced no visible colonies on YEA plates incubated at 36°C, whereas the cdc25–22 skb1Δ mutant was capable of growing at 36°C (Fig. 1A, Top). At 33°C, the cdc25–22 skb1Δ mutant produced substantially larger colonies than the cdc25–22 mutant (Fig. 1A, Bottom). These results demonstrate that the t.s. growth defect of the cdc25–22 mutant can be suppressed by the skb1Δ mutation. At 36°C, the cdc25–22 skb1Δ mutant grew more slowly than a cdc25–22 wee1Δ mutant (data not shown). We conclude that the skb1Δ mutation is not as potent a suppressor of the cdc25–22 t.s. mutation as the wee1Δ mutation. This finding is consistent with the observation that skb1Δ mutants, while shorter than wild-type cells at division, are substantially longer than wee1Δ cells (Table 1).
Table 1.
Cell size at division
| Strain | Overexpressed gene | Length at division, μm |
|---|---|---|
| Wild type | none | 15.2 ± 0.2 |
| skb1 | 23.4 ± 0.6 | |
| shk1 | 17.4 ± 0.5 | |
| skb1Δ | none | 13.3 ± 0.4 |
| cdc2-3w cdc25Δ | none | 18.3 ± 0.5 |
| skb1 | 35.8 ± 1.9 | |
| shk1 | 27.6 ± 1.0 | |
| wee1Δ | none | 7.7 ± 0.3 |
| skb1 | 7.6 ± 0.2 | |
| cdc2-1w | none | 9.0 ± 0.2 |
| skb1 | 9.0 ± 0.2 | |
| nmt-shk1 (33°C) | none | 10.9 ± 0.3 |
| cdc25-22 (33°C) | none | 38.8 ± 1.3 |
| cdc25-22 nmt-shk1 (33°C) | none | 26.5 ± 0.9 |
All cultures were grown to log phase at 28°C, unless otherwise indicated. skb1 was overexpressed from the plasmid pREP1 (nmt1 promoter). shk1 was overexpressed from pART1CM (adh1 promoter).
Figure 1.

Genetic interactions of skb1 with genes encoding key mitotic regulators in S. pombe. (A) The skb1 null mutation partially suppresses the t.s. growth defect of the cdc25–22 mutant. cdc25–22 and cdc25–22 skb1Δ S. pombe strains were streaked on YEA plates and incubated at either 36°C (Top) or 33°C (Bottom). The cdc25–22 skb1Δ mutant strain produced visible colonies at 36°C, whereas the cdc25–22 strain did not. At 33°C, the cdc25–22 skb1Δ strain grew significantly faster than the cdc25–22 mutant. (B) Overexpression of skb1 exacerbates the cdc25–22 t.s. growth defect. The cdc25–22 mutant was transformed with a plasmid for overexpression of skb1 (pREP1Skb1) or with a control plasmid (pREP1). Transformants were streaked onto EMM plates and were assayed for growth at 30°C (Top) or 33°C (Bottom). Overexpression of skb1 retarded the growth of the cdc25–22 mutant at 30°C and completely inhibited growth at 33°C. (C) cdc25–22 pREP1- and pREP1Skb1-transformed cells were grown at 26°C in liquid EMM to a density of about 106 cells per ml then shifted to either 33°C or 36°C and incubated for 5 hr before 4′,6-diamidino-2-phenylindole (DAPI) staining and microscopic analysis to determine the percentage of binucleated and septated cells in each culture. (D) skb1 overexpression is inhibitory to the growth of cdc2–3w cdc25Δ, but not cdc2–3w cells. pREP1- and pREP1Skb1-transformed cdc2–3w cells (Top) and cdc2–3w cdc25Δ cells (Bottom) were assayed for growth by being streaked on EMM plates and incubated at 30°C. (E) pREP1- and pREP1Skb1-transformed wild-type cells (CHP428) were streaked onto EMM and grown at either 30°C (Top) or 36°C (Bottom). (F) skb1 overexpression inhibits growth of the wee1Δ mutant at 36°C. pREP1- and pREP1Skb1-transformed wee1Δ cells were streaked onto EMM and grown at either 30°C (Top) or 36°C (Bottom).
We tested next the effects of skb1 overexpression in the cdc25–22 mutant. Overexpression of skb1 was not inhibitory to the growth of wild-type cells and was only slightly inhibitory to the growth of cdc25–22 cells at the permissive temperature of 26°C (data not shown). At 30°C, cdc25–22 cells that overexpressed skb1 from a high-copy plasmid grew more slowly than cells transformed with a control plasmid (Fig. 1B, Top). At 33°C, overexpression of skb1 caused cdc25–22 cells to become growth arrested, whereas cdc25–22 cells transformed with a control plasmid were competent for growth (Fig. 1B, Bottom). These results show that skb1 overexpression exacerbates the t.s. growth defect of the cdc25–22 mutant. We observed further microscopically that both skb1-overexpressing cdc25–22 cells incubated at 33°C and cdc25–22 control cells incubated at 36°C for 5 hr consisted primarily of nonseptated cells containing a single nucleus (Fig. 1D). In addition, fluorescence-activated cell-sorting analysis showed virtually identical profiles for skb1-overexpressing cdc25–22 cultures incubated at 33°C and cdc25–22 control cells incubated at 36°C (data not shown). Taken together, the above-described data suggest that Skb1 functions as a dosage-dependent mitotic inhibitor in S. pombe.
To address whether Skb1 inhibits mitosis in a Cdc25-dependent fashion, we examined the effect of skb1 overexpression in cdc2–3w and cdc2–3w cdc25Δ mutants. cdc2–3w mutants exhibit reduced responsiveness to the dosage of Cdc25 and retain viability even when the cdc25 gene is deleted (31). Overexpression of skb1 had no deleterious effect on the growth of cdc2–3w cells (Fig. 1D, Top). However, skb1 overexpression was highly inhibitory to the growth of cdc2–3w cdc25Δ cells (Fig. 1D, Bottom) and also caused these cells to divide at about twice the length at which they normally divide (Table 1). These results demonstrate that Skb1 inhibits mitosis by a Cdc25-independent mechanism.
We asked next whether Skb1 inhibits mitosis in a Wee1-dependent fashion. Whereas skb1 overexpression caused both wild-type and Cdc25-deficient cells to divide at a significantly greater length, it had no effect on the size at which wee1Δ cells divided (Table 1). In addition, skb1 overexpression did not inhibit mitosis in the cdc2–1w strain (Table 1), a mutant that is largely unresponsive to wee1 dosage (14–16). These results suggest that Skb1 inhibits mitosis by a Wee1-dependent mechanism. However, further analysis of skb1 overexpression in wee1 mutant strains complicated this conclusion. In addition to analyzing the effects of skb1 overexpression in the wee1 null mutant, we also overexpressed skb1 in the wee1–50 mutant, which carries a t.s. allele of wee1 (i.e., the wee1–50 mutant is viable, but exhibits a wee phenotype at 36°C). Surprisingly, we found that skb1 overexpression caused the wee1–50 mutant to be inhibited severely for growth at 36°C (data not shown). We then assayed skb1-overexpressing wee1Δ cells for t.s. growth and found that, like the wee1–50 mutant, skb1 overexpression caused the wee1Δ mutant to be severely inhibited for growth at 36°C (Fig. 1F). Overexpression of skb1 had no effect on the growth of wild-type cells at 36°C (Fig. 1E). Microscopic analysis indicated that skb1-overexpressing wee1Δ cells were approximately the same length as wee1Δ control cells (data not shown). Thus, skb1 overexpression does not affect the size at which wee1 mutant cells divide when incubated at 36°C.
Shk1 Is Required for Skb1-Dependent Mitotic Inhibitory Function.
Experiments were performed next to determine whether Skb1 mitotic inhibitory function depends on Shk1. We asked first whether overexpression of skb1 exacerbates the cdc25–22 t.s. growth defect in shk1-deficient cells. Since shk1 is an essential gene, we constructed a cdc25–22 mutant, SMMG127 (cdc25–22 nmt1-shk1), in which shk1 expression is under the control of a mutationally attenuated allele of the thiamine-repressible nmt1 promoter (Methods). Whereas skb1 overexpression caused cdc25–22 shk1+ cells to undergo growth arrest at 33°C (Figs. 1B and Fig. 2A, Top), it did not affect the growth of cdc25–22 nmt1-shk1 cells at 33°C (Fig. 2A, Bottom). This result suggests that the mitotic inhibitory function of Skb1 is at least partially dependent on Shk1.
Figure 2.

Genetic evidence that Shk1 has a mitotic inhibitory function. (A) Exacerbation of the cdc25–22 t.s. growth defect caused by skb1 overexpression is Shk1-dependent. PAAUCM- and pAAUCMSkb1-transformed cdc25–22 (Top) and cdc25–22 nmt-shk1 (Bottom) cells were streaked onto EMM containing 15 μM thiamine and incubated at 33°C for 3 days. (B) Overexpression of shk1 exacerbates the cdc25–22 t.s. growth defect. The cdc25–22 mutant was transformed with a plasmid for overexpression of shk1 (pAAUCMShk1) or with a control plasmid (pAAUCM). Transformants were streaked onto EMM plates and assayed for growth at 30°C (Top) and 33°C (Bottom). Overexpression of shk1 retarded the growth of the cdc25–22 mutant at 30°C and completely inhibited growth at 33°C. (C) Cooverexpression of skb1 and shk1 is only slightly inhibitory to growth of wild-type (CHP428) S. pombe cells. (D) Cooverexpression of skb1 and shk1 more severely exacerbates the cdc25–22 t.s. growth defect than overexpression of either gene alone. The cdc25–22 mutant was transformed with pREP1Skb1 (Skb1) and/or pAAUCMShk1 (Shk1) or with the control plasmids pREP1 and/or pAAUCM. Primary transformants were streaked onto EMM plates and incubated at 30°C. Cooverexpression of skb1 and shk1 completely inhibited growth of the cdc25–22 mutant at 30°C, whereas overexpression of either gene alone only retarded growth.
Interestingly, even though shk1-deficient (nmt-shk1) cells are substantially shorter than skb1Δ cells (Table 1), the shk1-deficient cdc25–22 strain exhibited the same t.s. growth characteristics as the cdc25–22 shk1+ strain (data not shown). Thus, unlike the skb1Δ mutation, shk1 deficiency does not suppress the cdc25–22 t.s. growth defect. However, overexpression of shk1, like skb1, exacerbated the t.s. growth defect of the cdc25–22 mutant (Fig. 2B) and was highly inhibitory to the growth of cdc2–3w cdc25Δ mutant (data not shown). Furthermore, cooverexpression of skb1 and shk1, which was only moderately inhibitory to the growth of wild-type S. pombe cells (Fig. 2C), completely inhibited growth of the cdc25–22 strain at 30°C (Fig. 2D), whereas overexpression of either gene alone, as already noted, resulted in only a retardation of growth of the cdc25–22 mutant at the same temperature. Thus, cooverexpression of skb1 and shk1 is more toxic to the cdc25–22 mutant than overexpression of either gene alone. shk1 overexpression, like overexpression of skb1, also caused the wee1Δ mutant to exhibit a t.s. growth defect (data not shown). Taken together, these various data suggest that Shk1 has a mitotic inhibitory function and that this function is required for Skb1-dependent mitotic inhibition. However, the failure of shk1 deficiency to suppress the cdc25–22 t.s. growth defect, in contrast to loss of skb1 function, raises the possibility that Skb1 may act on other proteins in addition to Shk1 to inhibit mitosis.
Skb1 and Shk1 Associate with Cdc2 in S. pombe.
Biochemical experiments were performed to address whether Skb1 and Shk1 are associated with a Cdc2 complex in S. pombe. Lysates were prepared from wild-type S. pombe cells expressing Skb1 and Shk1 as GST fusion proteins. The supernatant fractions of the cell lysates were incubated with glutathione agarose beads to purify GST fusion proteins and the resulting complexes were resolved by SDS/PAGE and were subjected to immunoblot analysis. As shown in Fig. 3A, we detected association of Cdc2 with both GST-Skb1 and GST-Shk1, but not with GST. These results provide evidence that Skb1 and Shk1 associate with a Cdc2 complex in S. pombe, suggesting that they inhibit mitosis by a mechanism that involves interaction with a component(s) of that complex, rather than by an indirect mechanism. Using this same strategy, we were able also to coprecipitate Skb1 with a GST fusion of an N-terminally truncated Shk1 protein (Shk1ΔN) lacking the first 116 amino acids of the full length Shk1 sequence, but only from cells in which Skb1 was overexpressed (Fig. 3B). This result provides biochemical confirmation of the Skb1–Shk1 two-hybrid interaction, which we described in a previous report (11). We were unable to assay for coprecipitation of Skb1 with GST-full length Shk1 because cooverexpression of the two proteins was toxic to S. pombe cells (data not shown).
Figure 3.
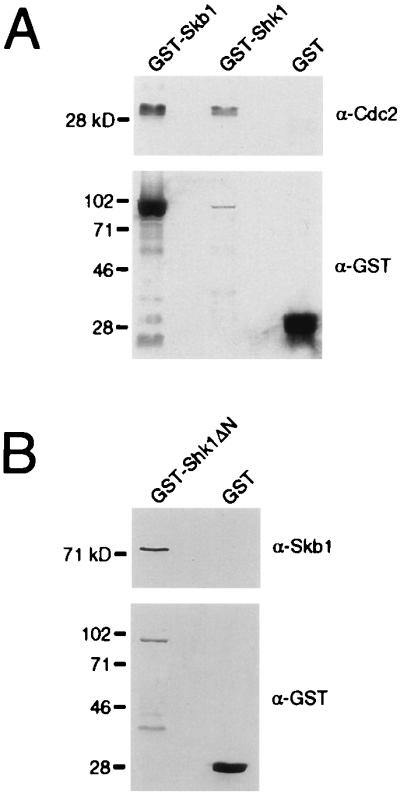
Skb1 and Shk1 associate with Cdc2 and with each other in fission yeast. (A) S. pombe strain CHP428 was transformed with plasmids for expression of GST (pAAUGST), GST-Skb1 (pAAUGSTSkb1), or GST-Shk1 (pAAUGSTShk1) fusion proteins. Transformants were grown to about 107 cells per ml in EMM, then lysed and GST complexes precipitated as described in Methods. Complexes were resolved by SDS/PAGE and transferred to nitrocellulose. Immunoblotting was performed by using anti-Cdc2 PSTAIRE antibody (Top). Membranes were then stripped and reprobed with anti-GST antibody (Bottom). Cdc2 protein was associated with GST-Skb1 and GST-Shk1, but not with GST. (B) Shk1 and Skb1 proteins associate in S. pombe cells. Lysates were prepared from CHP428 cells overexpressing skb1 and either GST or GST-Shk1ΔN, as described for A. GST and GST-Shk1ΔN complexes were resolved by SDS/PAGE and subjected to immunoblot analysis by using anti-Skb1 antibody (Top). Blots were subsequently stripped and probed with anti-GST antibody (Bottom). Skb1 protein was associated with GST-Shk1ΔN, but not with GST.
Functional Homology of S. pombe and Human Skb1 Proteins.
In our original report on the cloning and characterization of skb1, we described a partial sequence for a human homolog of skb1, SKB1Hs, which we derived from overlapping EST sequences identified from a blast search of the nucleic acid databases (11). We have since cloned and sequenced a probable full-length SKB1Hs cDNA (GenBank accession no. AF015913) (Materials and Methods). The predicted Skb1Hs protein is 39% identical to S. pombe Skb1 (henceforth Skb1Sp) and 34% identical to a predicted Skb1-related protein, Skb1Ce, encoded by the uncharacterized Caenorhabditis elegans ORF CE34E10.5 (Fig. 4). Human genomic DNA digested with either BamHI or EcoRI was subjected to Southern blot analysis by using the SKB1Hs cDNA as a probe (Fig. 4B). Single hybridizing bands were produced from each digest. No additional bands were detected under low-stringency hybridization conditions (data not shown), suggesting that there are no other genes closely related to SKB1Hs at the nucleic acid sequence level.
Figure 4.

(A) Alignment of human (Skb1Hs), S. pombe (Skb1Sp), and C. elegans (Skb1Ce) Skb1 homologs. Alignments were generated by using the lasergene megalign program (DNAstar, Madison, WI). Identical amino acid residues are indicated by black boxes. (B) Southern blot of BamHI- (Left) and EcoRI- (Right) digested human genomic DNA by using the SKB1Hs cDNA as a probe.
A plasmid was constructed for expressing SKB1Hs in S. pombe from the nmt1 promoter. Expression of SKB1Hs not only caused wild-type S. pombe cells to divide at a significantly greater length, but also restored greater than wild-type length to skb1Δ cells (Fig. 5A). Furthermore, expression of SKB1Hs exacerbated the t.s. growth defect of both cdc25–22 (not shown) and cdc25–22 skb1Δ (Fig. 5B) mutants and was highly inhibitory to the growth of cdc2–3w cdc25Δ cells (Fig. 5C). In addition, Cdc2 coprecipitated with Skb1Hs GST fusion protein in S. pombe cells (Fig. 5D). These results demonstrate that the Skb1Sp and SKB1Hs proteins are conserved substantially in function.
Figure 5.

Skb1Hs functions as a mitotic inhibitor in S. pombe. (A) Cell division size data for wild-type and skb1Δ cells transformed with pREP1Skb1Hs, for expression of SKB1Hs from the S. pombe nmt1 promoter, or the control plasmid pREP1. SKB1Hs expression causes wild-type and skb1Δ cells to divide at a substantially greater length. (B) SKB1Hs expression exacerbates the t.s. growth defect of a cdc25–22 skb1Δ mutant (Bottom), but does not inhibit the growth of wild-type S. pombe cells (Top). (C) SKB1Hs expression is inhibitory to the growth of cdc2–3w cdc25Δ mutant (Bottom) but not the cdc2–3w mutant (Top). (D) Cdc2 associates with GST-Skb1Hs protein in fission yeast. Cells expressing either GST or GST-Skb1Hs were grown to log phase, lysed, and GST complexes isolated and subjected to immunoblot analysis with anti-Cdc2 and anti-GST antibody, as described in Fig. 3.
DISCUSSION
In conclusion, we have provided evidence that the protein encoded by the highly conserved skb1 gene negatively regulates mitosis in S. pombe by a mechanism that is at least partially dependent on the PAK homolog Shk1 and the Cdc2 inhibitory kinase Wee1. Our results provide evidence of a previously undescribed role for PAK-related protein kinases as mitotic inhibitors. We have provided biochemical evidence that Skb1Sp and Shk1 associate with Cdc2 in S. pombe, suggesting that the mitotic inhibitory activities of Skb1Sp and Shk1 occur through interaction with the Cdc2 complex, rather than by an indirect mechanism. The simplest model accounting for our results would place Skb1Sp and Shk1 function upstream of Wee1. However, our results do not rule out the possibility that Skb1Sp and Shk1 act in a codependent parallel pathway that converges on Cdc2 with the Wee1 pathway. Indeed, our finding that overexpression of either skb1Sp or shk1 causes wee1 mutants to exhibit t.s. growth defects may be an indication that Skb1Sp and Shk1 do, in fact, function in a pathway parallel to that of Wee1. We have found that purified recombinant Skb1Sp and Shk1 proteins do not inhibit Cdc2⋅Cdc13 activity in vitro (R.P. and S.M., unpublished results), suggesting that Skb1 and Shk1 do not likely act directly on Cdc2.
Skb1Sp is a member of a novel, but highly conserved, family of proteins that are apparently ubiquitous in eukaryotic organisms. We have shown here that a human homolog of Skb1Sp, Skb1Hs, can functionally substitute for Skb1Sp in S. pombe. This result suggests that Skb1 protein function has been remarkably conserved through evolution. It will be interesting to determine whether Skb1-related proteins, together with PAK-related kinases, function as mitotic regulators in higher organisms. Interestingly, Pollack and colleagues have cloned SKB1Hs from a two-hybrid screen for Janus kinase (JAK)2-interacting proteins (B. Pollack and S. Pestka, personal communication). This interaction suggests the possibility that Skb1Hs could link JAK2 to a PAK signaling pathway in mammalian cells, a hypothesis consistent with the observation that certain ligands that activate JAKs also activate PAKs (32). Moreover, since JAKs are activated both by ligands that have proliferative effects, such as erythropoietin, and by ligands with antiproliferative effects, such as interferon (33), the possibility exists that Skb1 proteins could link JAK-mediated signaling pathways to cell-cycle control complexes.
An skb1-related gene, HSL7, has also been cloned from S. cerevisiae as a result of a genetic screen for mutations that are lethal in combination with a deletion of the histone H3 amino terminus (34). Interestingly, S. cerevisiae cells carrying an hsl7 null mutation exhibit a G2 delay. This phenotype is in striking contrast to that of S. pombe skb1Sp null mutants, which exit prematurely from G2. Based on these results, it would appear that Skb1Sp and Hsl7 have opposite functions, with Skb1Sp being a mitotic inhibitor in S. pombe and Hsl7 possibly functioning as a mitotic inducer in S. cerevisiae. The mitotic delay caused by the hsl7 mutation is suppressed by deletion of the swe1 gene, which encodes the single S. cerevisiae homolog of Wee1 (34). Thus, Hsl7 interacts genetically with Swe1, although in a fashion opposite to the Skb1–Wee1 interaction in S. pombe. Unlike skb1Sp, no gain-of-function phenotype(s) have yet been described for HSL7, and such phenotypes would be highly useful for further clarifying the role of Hsl7 during mitosis in S. cerevisiae. We have found that, unlike human Skb1Hs, Hsl7 cannot substitute for Skb1Sp in S. pombe (P.Y. and S.M., unpublished results), suggesting that Skb1Sp and Skb1Hs are more closely related in function than either is to Hsl7.
Genetic data suggest that Shk1 performs at least two distinct functions in S. pombe. One function is required for normal mating response, which is not essential for cell viability or normal cell shape, and which involves interaction between Shk1 and a Ras-dependent mitogen-activated protein kinase pathway (3, 7, 35). The second Shk1-dependent function is essential for cell viability and, possibly, normal morphology (3, 7). At this time, it is unclear whether the cell-cycle regulatory functions of Shk1 are distinct from its function(s) as a morphological regulator. It also remains to be established whether the essential function(s) of Shk1 are related to its role(s) as a mitotic and morphological regulator. Further elucidation of Shk1-mediated cellular functions will be greatly facilitated by the identification of potential downstream targets for Shk1, and we are presently conducting screens to identify such targets.
Acknowledgments
We thank Anjana Kundu for technical assistance; Kathy Gould, Vicki Huff, Randy Legerski, Nick Rhind, and Shelley Sazer for strains and materials; and Jenny Henkel, Nick Rhind, and Anthony Polverino for comments on the manuscript. M.G. was supported by a National Institutes of Health predoctoral training grant. This work was supported by National Institutes of Health Grant R01GM53239 (to S.M.).
ABBREVIATIONS
- PAKS
p21Cdc42/Rac-activated kinases
- GST
glutathione S-transferase
- t.s.
temperature sensitive
- JAK
Janus kinase
Note Added in Proof
Krapivinsky et al. (36) have identified Skb1Hs as a protein that binds to a potential chloride channel protein called pICln.
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
References
- 1.Sells M A, Chernoff J. Trends Cell Biol. 1997;7:162–167. doi: 10.1016/S0962-8924(97)01003-9. [DOI] [PubMed] [Google Scholar]
- 2.Polverino A, Frost J, Yang P, Hutchison M, Neiman A M, Cobb M H, Marcus S. J Biol Chem. 1995;270:26067–26070. doi: 10.1074/jbc.270.44.26067. [DOI] [PubMed] [Google Scholar]
- 3.Marcus S, Polverino A, Chang E, Robbins D, Cobb M H, Wigler M H. Proc Natl Acad Sci USA. 1995;92:6180–6184. doi: 10.1073/pnas.92.13.6180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Leberer E, Dignard D, Harcus D, Thomas D Y, Whiteway M. EMBO J. 1992;11:4815–4824. doi: 10.1002/j.1460-2075.1992.tb05587.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zhang S, Han J, Sells M A, Chernoff J, Knaus U G, Ulevitch R J, Bokoch G M. J Biol Chem. 1995;270:23934–23936. doi: 10.1074/jbc.270.41.23934. [DOI] [PubMed] [Google Scholar]
- 6.Frost J A, Xu S, Hutchison M R, Marcus S, Cobb M H. Mol Cell Biol. 1996;16:3707–3713. doi: 10.1128/mcb.16.7.3707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ottilie S, Miller P J, Johnson D I, Creasy C L, Sells M A, Bagrodia S, Forsburg S L, Chernoff J. EMBO J. 1995;14:5908–5919. doi: 10.1002/j.1460-2075.1995.tb00278.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Manser E, Huang H Y, Loo T H, Chen X Q, Dong J M, Leung T, Lim L. Mol Cell Biol. 1997;17:1129–1143. doi: 10.1128/mcb.17.3.1129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sells M A, Knaus U G, Bagrodia S, Ambrose D M, Bokoch G M, Chernoff J. Curr Biol. 1997;7:202–210. doi: 10.1016/s0960-9822(97)70091-5. [DOI] [PubMed] [Google Scholar]
- 10.Rudel T, Bokoch G M. Science. 1997;276:1571–1574. doi: 10.1126/science.276.5318.1571. [DOI] [PubMed] [Google Scholar]
- 11.Gilbreth M, Yang P, Wang D, Frost J, Polverino A, Cobb M H, Marcus S. Proc Natl Acad Sci USA. 1996;93:13802–13807. doi: 10.1073/pnas.93.24.13802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Burbelo P D, Drechsel D, Hall A. J Biol Chem. 1995;270:29071–29074. doi: 10.1074/jbc.270.49.29071. [DOI] [PubMed] [Google Scholar]
- 13.Miller P J, Johnson D I. Mol Cell Biol. 1994;14:1075–1083. doi: 10.1128/mcb.14.2.1075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Russell P, Nurse P. Cell. 1987;49:559–567. doi: 10.1016/0092-8674(87)90458-2. [DOI] [PubMed] [Google Scholar]
- 15.Forsburg S L, Nurse P. Annu Rev Cell Biol. 1991;7:227–256. doi: 10.1146/annurev.cb.07.110191.001303. [DOI] [PubMed] [Google Scholar]
- 16.Wu L, Russell P. EMBO J. 1997;16:1342–1350. doi: 10.1093/emboj/16.6.1342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Okayama H, Nagata A, Jinno S, Murakami H, Tanaka K, Nakashima N. Adv Cancer Res. 1996;69:17–62. doi: 10.1016/s0065-230x(08)60859-3. [DOI] [PubMed] [Google Scholar]
- 18.Parker L L, Atherton-Fessler S, Piwnica-Worms H. Proc Natl Acad Sci USA. 1992;89:2917–2921. doi: 10.1073/pnas.89.7.2917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lundgren K, Walworth N, Booher R, Dembski M, Kirschner M, Beach D. Cell. 1991;64:1111–1122. doi: 10.1016/0092-8674(91)90266-2. [DOI] [PubMed] [Google Scholar]
- 20.Gould K L, Moreno S, Owen D J, Sazer S, Nurse P. EMBO J. 1991;10:3297–3309. doi: 10.1002/j.1460-2075.1991.tb04894.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Buck V, Russell P, Millar J B. EMBO J. 1995;14:6173–6183. doi: 10.1002/j.1460-2075.1995.tb00308.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Russell P, Nurse P. Cell. 1986;45:145–153. doi: 10.1016/0092-8674(86)90546-5. [DOI] [PubMed] [Google Scholar]
- 23.Millar J B, McGowan C H, Lenaers G, Jones R, Russell P. EMBO J. 1991;10:4301–4309. doi: 10.1002/j.1460-2075.1991.tb05008.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Thuriaux P, Nurse P, Carter B. Mol Gen Genet. 1978;161:215–220. doi: 10.1007/BF00274190. [DOI] [PubMed] [Google Scholar]
- 25.Alfa C, Fantes P, Hyams J, McLeod M, Warbrick E. Experiments with Fission Yeast: A Laboratory Course Manual. Plainview, NY: Cold Spring Harbor Lab. Press; 1993. [Google Scholar]
- 26.Maundrell K. J Biol Chem. 1990;265:10857–10864. [PubMed] [Google Scholar]
- 27.Basi G, Schmid E, Maundrell K. Gene. 1993;123:131–136. doi: 10.1016/0378-1119(93)90552-e. [DOI] [PubMed] [Google Scholar]
- 28.Chang E C, Barr M, Wang Y, Jung V, Xu H P, Wigler M H. Cell. 1994;79:131–141. doi: 10.1016/0092-8674(94)90406-5. [DOI] [PubMed] [Google Scholar]
- 29.McLeod M, Stein M, Beach D. EMBO J. 1987;6:729–736. doi: 10.1002/j.1460-2075.1987.tb04814.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Fantes P. Nature (London) 1979;279:428–430. doi: 10.1038/279428a0. [DOI] [PubMed] [Google Scholar]
- 31.Millar J B, Lenaers G, Russell P. EMBO J. 1992;11:4933–4941. doi: 10.1002/j.1460-2075.1992.tb05600.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Teo M, Manser E, Lim L. J Biol Chem. 1995;270:26690–26697. doi: 10.1074/jbc.270.44.26690. [DOI] [PubMed] [Google Scholar]
- 33.Ihle J N. Philos Trans R Soc Lond B. 1996;351:159–166. doi: 10.1098/rstb.1996.0012. [DOI] [PubMed] [Google Scholar]
- 34.Ma X J, Lu Q, Grunstein M. Genes Dev. 1996;10:1327–1340. doi: 10.1101/gad.10.11.1327. [DOI] [PubMed] [Google Scholar]
- 35.Yang P, Kansra S, Pimental R A, Gilbreth M, Marcus S. J Biol Chem. 1998;273:18481–18489. doi: 10.1074/jbc.273.29.18481. [DOI] [PubMed] [Google Scholar]
- 36.Krapivinsky G, Pu W, Wickman K, Krapivinsky L, Clapham D E. J Biol Chem. 1998;273:10811–10814. doi: 10.1074/jbc.273.18.10811. [DOI] [PubMed] [Google Scholar]