

Abstract
Molecular phylogenetic analyses, based mainly on ribosomal RNA, show that three amitochondriate protist lineages, diplomonads, microsporidia, and trichomonads, emerge consistently at the base of the eukaryotic tree before groups having mitochondria. This suggests that these groups could have diverged before the mitochondrial endosymbiosis. Nevertheless, since all these organisms live in anaerobic environments, the absence of mitochondria might be due to secondary loss, as demonstrated for the later emerging eukaryote Entamoeba histolytica. We have now isolated from Trichomonas vaginalis a gene encoding a chaperone protein (HSP70) that in other lineages is addressed to the mitochondrial compartment. The phylogenetic reconstruction unambiguously located this HSP70 within a large set of mitochondrial sequences, itself a sister-group of α-purple bacteria. In addition, the T. vaginalis protein exhibits the GDAWV sequence signature, so far exclusively found in mitochondrial HSP70 and in proteobacterial dnaK. Thus mitochondrial endosymbiosis could have occurred earlier than previously assumed. The trichomonad double membrane-bounded organelles, the hydrogenosomes, could have evolved from mitochondria.
Keywords: mitochondrial origin, hydrogenosome, phylogeny, trichomonads
Trichomonads (phylum Parabasala) are flagellated protists that possess a well developed flagellar apparatus, a large cytoskeleton, and a Golgi apparatus (1). Most of them are parasites. They live under anaerobic conditions or in a low oxygen level habitats. They lack mitochondria and peroxisomes but have hydrogenosomes. Hydrogenosomes are double membrane-bounded organelles, lacking DNA, which have no tricarboxylic acid cycle enzymes and cytochromes and do not carry out oxidative phosphorylation (2, 3). Similarly to mitochondria, however, these organelles use pyruvate as a major substrate and oxidize it to acetyl-CoA. Ferredoxin and succinate thiokinase are similar to mitochondrial ones. But pyruvate:ferredoxin oxidoreductase (4) and hydrogenase (5) are specific to the hydrogenosomes. The electrons produced in this process reduce protons and are eliminated as molecular hydrogen. Some hydrogenosomal enzymes are similar to mitochondrial ones, but some others are specific (3). Thus two main hypotheses have been raised for the evolutionary origin of hydrogenosomes (3, 6, 7). They are either modified mitochondria or descendants of an anaerobic bacterium that evolved toward another type of symbiotic organelle.
In the eukaryotic tree based on small subunit rRNA comparison, trichomonads, together with microsporidia and diplomonads, represent one of the early branching groups (8, 9, 10). The fact that these three groups emerging at the base of the eukaryotic tree are amitochondriate obviously suggests that mitochondrial symbiosis may have occurred at a later stage of eukaryotic evolution. These phyla that lack mitochondria and peroxisomes were previously classified into a kingdom named Archaezoa (11). More recently, Cavalier-Smith (12, 13) proposed to exclude Parabasala from Archaezoa because they show typical stacked dictyosomes of Golgi apparatus.
Absence of mitochondria in trichomonads could be explained by a primitive absence (14, 15), a secondary modification toward hydrogenosome or a loss. After an endosymbiotic event, a part of the genetic material of the symbiont is transferred to the nucleus and could remain in the genome even after a secondary loss. We decided thus to search for a gene that could be such a marker helping to choose between these hypotheses.
The 70-kDa heat shock proteins (HSP70s) are among the most highly conserved proteins present in all organisms, Archaebacteria, Eubacteria, and Eukaryota (16). They are involved in the correct folding of mature proteins and in the translocation of newly synthesized polypeptides across membranes (17). In eukaryotes, they constitute a multigenic family, the members of which are located in cytosol, endoplasmic reticulum, mitochondria, and chloroplasts. It has been demonstrated that a duplication that occurred at the beginning of the eukaryotic evolution has given rise to the cytosolic and endoplasmic reticulum genes, while the mitochondrial and chloroplast genes are the result of a transfer from the organelle DNA to the nucleus (18). In addition to typical eukaryotic genes, we have found in Trichomonas vaginalis two genes of eubacterial affinity. A detailed phylogenetic analysis demonstrated that these genes consistently emerged within the mitochondrial group. This suggested that the mitochondrial endosymbiotic event could have occurred before the emergence of T. vaginalis.
MATERIALS AND METHODS
DNA Isolation.
DNA was extracted, as described (19), from an axenic culture of a T. vaginalis strain (from Homo sapiens, isolated by Guy Brugerolle, University Clermont–Ferrand 2, Aubiere, France).
Cloning of T. vaginalis HSP70 Genes.
We amplified 700-bp fragments using degenerate primers corresponding to the 5′ terminal end of HSP70 gene. Primers were based on highly conserved regions of the protein, GIDLGT (5′-CCCGGGIATHGAYYTIGGNAC-3′, sense primer) and GGEDFE (5′-GAATTCYAAYTCRTCICCNCC-3′, antisense primer), respectively located at the positions 6 and 223 of the Agrobacterium tumefaciens sequence (20). PCR was carrying out according to the standard conditions (Biotaq, Bioprobe International, Richmond, CA; Trio-Thermoblock, Eurogentec, Brussels) with 30 cycles of 94°C for 45 sec, 55°C for 1 min, and 72°C for 1 min. A single band of the expected size was obtained and cloned into the phagemid pBluescript II SK− (Stratagene). Among all sequenced clones (corresponding to eight different HSP70s) using the T7 sequencing kit (Pharmacia), we chose one of the two that showed close phylogenetic relationship with mitochondrial sequences. Then, this sequence was used as a probe to screen a λZap II cDNA library. One of the isolated clones contained an insert of ≈2 kb and was sequenced using primer walking.
Southern Blotting.
T. vaginalis DNA was digested with various restriction enzymes (HindIII, SalI, SspI, and EcoRV). The fragments obtained were separated by agarose gel electrophoresis, transferred to nitrocellulose membrane and hybridized to the 0.7-kb PCR product already used for library screening.
Phylogenetic Analyses.
The amino acid sequence of T. vaginalis was manually entered into an extensive alignment of HSP70 homologs using the must package (21). Only 499 unambiguously aligned positions were used. Phylogenetic trees were constructed using maximum likelihood, maximum parsimony, and distance methods performed by, respectively, the protml program (22) version 2.2, the paup program (23) version 3.1, and the must package (21) version 1.0.
Maximum parsimony trees were obtained by 100 random addition heuristic search replicates and maximum likelihood trees by the quick add operational taxonomic units search, using the Jones–Taylor–Thornton model of amino acid substitution and retaining the 300 top ranking trees (options -jf -q -n 300). Bootstrap proportions were calculated by analysis of 100 replicates for maximum parsimony analysis and 1000 for neighbor joining analysis (24). For maximum likelihood analysis, bootstrap proportions are computed by using the resampling of estimated likelihood method (25) because of computing time limitations.
RESULTS
Fragments of about 700 bp corresponding to eight different HSP70 genes have been amplified by PCR from whole genomic DNA of T. vaginalis. Among them, five were of endoplasmic reticulum type, one of cytosolic type (data not shown) and two exhibited eubacterial features. Preliminary phylogenetic analysis demonstrated that the latter two sequences emerged together within the cluster of mitochondrial sequences. To make sure that these two fragments are from T. vaginalis, they were hybridized under high stringent conditions to Southern blots of genomic DNA (data not shown). Thus, we decided to obtain the complete sequence of one of them by screening a cDNA library. The insert was 1865 bp and showed an ORF of 609 amino acids. The codon usage is of the same pattern as for β-tubulin (26) and ferredoxin (27) genes of T. vaginalis. The 3′-flanking region of the cDNA clone was A+T rich and short, as already noted (28, 29). The 5′ end unfortunately was truncated so the clone did not contain more than nine nucleotides upstream from the first methionine. They might be coding for a part of the processed sequence as for hydrogenosomal enzymes (29).
The general phylogeny of Eubacteria (Fig. 1), obtained using HSP70 sequences and maximum likelihood method, was in agreement with phylogenies based on other molecular markers (30–32), demonstrating that HSP70 was an accurate phylogenetic indicator (16). For example, both high G+C and low G+C Gram-positive bacteria were monophyletic. Mitochondrial and chloroplastic sequences are sister-groups of purple bacterial and cyanobacterial ones, respectively.
Figure 1.
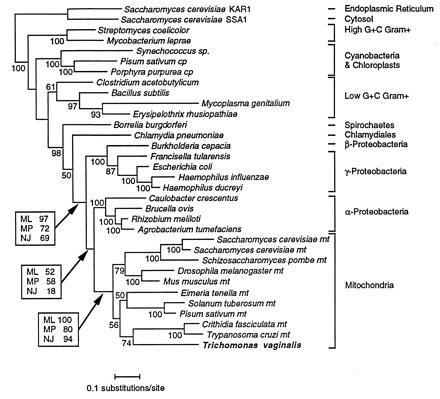
Phylogenetic tree based on comparison of major eubacterial dnaK and organellar HSP70. The outgroup is represented by two eukaryotic sequences of HSP70 from Saccharomyces cerevisiae, an endoplasmic reticulum resident form (KAR1) and a cytosolic resident form (SSA1). The tree was constructed using the maximum likelihood method using the Jones–Taylor–Thornton model of amino acid substitutions (lnL = −16059.6). The reliability of nodes is given as bootstrap values, shown below the considered nodes. For the three nodes of main interest, the bootstrap proportions computed by the three different tree reconstruction methods are given in boxes.
Phylogenetic trees were also constructed using maximum parsimony method (giving 11 equally parsimonious trees of length 2796) and using neighbor joining method. The topologies are similar except those concerning the relationships between high G+C Gram-positive and cyanobacteria on one hand and between spirochetes and chlamydiales on other hand. Since species sampling (33) can influence the results, we have confirmed that this result was obtained irrespective of the species sampling, particularly when using all 60 available prokaryotic sequences (data not shown).
Position of T. vaginalis within this mitochondrial group was supported by high bootstrap values, whatever the tree reconstruction method used: maximum likelihood, 100%; maximum parsimony, 80%; and neighbor joining, 94%. In this subtree, T. vaginalis emerged within mitochondrial HSP70 forming a sister-group with Euglenozoa, with a bootstrap value of 74%.
The mitochondrial group belongs to the purple bacteria (30, 34, 35) with bootstrap values of 97% for maximum likelihoood, 72% for maximum parsimony, and 69% for neighbor joining. More precisely, mitochondria appeared to be a sister-group of the α subdivision of purple bacteria, but not included within them, probably because of the absence, in our sample, of the rickettsiales, the assumed closest relatives of mitochondria (30).
In addition, the alignment exhibited two strong signatures (Fig. 2). Firstly, T. vaginalis shared with mitochondria and purple bacteria the GDAWV sequence. Borrelia burgdorferi and Chlamydia pneumoniae also possessed a part of this signature, GDA, confirming the phylogenetic affinity between spirochetes, chlamydiales and purple bacteria (30). The other signature, the YSPSQI sequence, was shared by all mitochondrial and α subdivision of proteobacterial sequences, but with a few conservative substitutions. This confirms the monophyly of mitochondria and of α-Proteobacteria.
Figure 2.

Alignment of a portion of T. vaginalis HSP70 with eubacterial and organellar homologs showing mitochondrial signatures. On this alignment, the first amino acid corresponds to amino acid 70 on the A. tumefaciens sequence. Our sequence was aligned with mitochondrial homologs, a set of eubacterial dnaK sequences representing the major already sequenced groups and also with a chloroplastic HSP70. Identical amino acids were indicated with dots and deletions with dashes. Numerous mitochondrial signatures have already been pointed out by Falah and Gupta (49). We present the signatures shared by the T. vaginalis sequence, mitochondrial homologs and related eubacterial dnaK. Box A (position 99 on the A. tumefaciens sequence) corresponds to the GDAWV signature specific of mitochondrial HSP70s and proteobacterial dnaK. This signature is hyper-conserved since no substitution is observed within these groups. Box B (position 110 on the A. tumefaciens sequence) shows the YSPSQI signature found in mitochondrial homologs and only in the α subdivision of proteobacterial dnaK. This signature is less conserved in Caulobacter crescentus but most of its substitutions are conservative changes. A 26 amino acids insertion (16) shared by all species and absent in low G+C and high G+C Gram-positive bacteria can be noted (box C).
DISCUSSION
We have shown the existence in the protist T. vaginalis of a typical HSP70 gene, that in other eukaryotes codes for a protein located in mitochondria. Phylogenetic analyses located T. vaginalis sequence inside the mitochondrial group. In addition, this HSP70 presented mitochondrial signatures shared only by α-Proteobacteria or the whole group of purple bacteria. Therefore, the evidence is consistent with this HSP70 sequence being of mitochondrial origin in spite of the fact that T. vaginalis lacks mitochondria. This nuclear sequence could be the result of an ancient gene transfer from mitochondria to nucleus, suggesting that trichomonads have had mitochondria in their early history.
This sequence could also be the result of an horizontal transfer. Since phylogenetic analysis locates it within the mitochondrial group, the donor organism should be a mitochondrion. It thus seems possible to exclude the hypothesis of an horizontal transfer from a bacteria to Trichomonas. Because all the known mitochondrial HSP70s are nuclear-encoded, the donor organism might be an eukaryote. But the horizontal transfer between eukaryotic lineages is known to have a low frequency and so far seems to involve only mobile elements (for review, see ref. 36). In addition, Roger et al. (37) have found another gene of mitochondrial affinity in T. vaginalis, a chaperonin (cpn60). Their phylogenetic analyses based on distance, parsimony, and maximum likelihood methods strongly support the emergence of T. vaginalis protein within the mitochondrial ones. Moreover, a cpn60 is decorated by heterologous antibodies (38) and shows an hydrogenosomal location. Thus, two low probability events are required in the hypothesis of horizontal transfer, a much less parsimonious hypothesis than that of mitochondrial loss.
This result provides a new insight into the origin of hydrogenosomes in parabasalids. Numerous hypotheses have already been proposed, but two of them remain likely. Hydrogenosomes could have originated through endosymbiosis of an anaerobic bacterium that contained most of the enzymes retained by actual hydrogenosomes (39, 40). Alternatively, they could arise by conversion of other organelles such as mitochondria (13, 41).
To elucidate the evolutionary origin of hydrogenosome, others have sequenced a number of its enzymes (29, 42). Analysis of relationships of hydrogenosomal constituents with homologous proteins did not cluster T. vaginalis enzymes with mitochondrial ones. But Hrdý and Müller (29) have shown that such a strong divergence of hydrogenosomal enzymes could be due to the high mutational saturation level of these proteins and to the small number of available sequences, especially from protists. Comparison of the saturation levels (43) suggests that the HSP70 is less affected by multiple substitutions than enzymes of the hydrogenosomal metabolism (not shown) and thus HSP70 is a better phylogenetic marker.
Our results are consistent with the hypothesis that parabasalid hydrogenosomes are modified mitochondria, although the possibility that mitochondria have been completely lost and that another endosymbiosis occurred cannot be excluded. More generally, secondary cytological disappearance of mitochondria and correlative appearance of hydrogenosomes have occurred repeatedly in bona fide aerobic lineages that have adapted to anaerobic environment such as rumen ciliates and fungi (44). The ciliates that possess hydrogenosomes with double bounding membranes are polyphyletic, as evidenced from small subunit ribosomal rRNA-based phylogeny (45). This supports the idea that these hydrogenosomes could derive from mitochondria. Nevertheless, in the fungus Neocallimastix, hydrogenosomes are single-membrane bounded. This leads one to suppose that, in fungi, they could be derived from peroxisomes rather than from mitochondria (6). Indeed, the finding that proteins of fungal hydrogenosomes have the sexine–lysine–leucine peroxisomal targeting sequences provides support for this hypothesis (46).
It is thus easier to imagine that these numerous cases correspond to a conserved possibility of converting mitochondria or peroxisomes to hydrogenosomes rather than to several independent bacterial symbioses converging toward a functionally similar organelle. Nevertheless, the problem of the origin of the metabolic properties of the hydrogenosome remains an open question. Hrdý and Müller (29) have suggested that in energy metabolism replacement of an enzyme by another or horizontal gene transfer (from prokaryotes to eukaryotes) could occur in an easy manner.
In summary, our data provide evidence that T. vaginalis may have had mitochondria during its evolution as it is the case for other extant amitochondriate protists such as Entamoeba histolytica (47). Secondary loss of mitochondria could be a recurrent phenomenon (48) and it would be interesting to look for mitochondrial molecular markers in the two other deep branching amitochondriate lineages, diplomonads and microsporidia. More generally, the absence of some characteristics in these lineages (for example, peroxisomes) has led others to propose scenarii of progressive construction of the eukaryotic cell (7, 13, 15) by assuming that the groups at the base of the eukaryotic tree could be relics of ancestral states. However, secondary loss of complex characters like mitochondria leads us to suggest that the common ancestor to all the extent eukaryotic species displayed all the major eukaryotic characteristics. Moreover, the mitochondrial endosymbiosis might be an evolutionary key innovation that provided sufficient selective advantages to eliminate all the other eukaryotic ancestral forms.
Acknowledgments
We acknowledge the constructive criticisms of Dr. Miklós Müller and thank Prof. André Adoutte for reading the manuscript. We also thank Dr. Patricia Johnson for providing a sample of the T. vaginalis cDNA library and Dr. Guy Brugerolle for supplying a culture and for support. We are grateful to Karine Budin for her help in sequence management and to Nicolas Puchot for his technical help. This work was supported by a grant from the Groupement de Recherches et d’Études sur les Génomes, décision d’aide no. 94/125. A.G. was supported by a fellowship from the Ministère de l’Éducation Nationale, de l’Enseignement Supérieur, de la Recherche et de l’Insertion Professionnelle.
Footnotes
References
- 1.Brugerolle G. Protoplasma. 1991;164:70–90. [Google Scholar]
- 2.Johnson P J, Lahti C J, Bradley P J. J Parasitol. 1993;79:664–670. [PubMed] [Google Scholar]
- 3.Müller M. J Gen Microbiol. 1993;139:2879–2889. doi: 10.1099/00221287-139-12-2879. [DOI] [PubMed] [Google Scholar]
- 4.Hrdý I, Müller M. J Mol Evol. 1995;41:388–396. [PubMed] [Google Scholar]
- 5.Bui E T N, Johnson P J. Mol Biochem Parasitol. 1996;76:305–310. doi: 10.1016/0166-6851(96)02567-4. [DOI] [PubMed] [Google Scholar]
- 6.Cavalier-Smith T. Ann NY Acad Sci. 1987;503:55–71. doi: 10.1111/j.1749-6632.1987.tb40597.x. [DOI] [PubMed] [Google Scholar]
- 7.Hübsmann N, Hausman K. Eur J Protistol. 1994;30:365–371. [Google Scholar]
- 8.Leipe D D, Gunderson J H, Nerad T A, Sogin M L. Mol Biochem Parasitol. 1993;59:41–48. doi: 10.1016/0166-6851(93)90005-i. [DOI] [PubMed] [Google Scholar]
- 9.Hashimoto T, Nakamura Y, Nakamura F, Shirakura T, Adachi J, Goto N, Okamoto K, Hasegawa M. Mol Biol Evol. 1994;11:65–71. doi: 10.1093/oxfordjournals.molbev.a040093. [DOI] [PubMed] [Google Scholar]
- 10.Philippe H, Adoutte A. In: Protistological Actualities: Proceedings of the Second European Congress of Protistology. Brugerolle G, Mignot J P, editors. France: Aubiere; 1995. pp. 17–33. [Google Scholar]
- 11.Cavalier-Smith T. Nature (London) 1987;326:332–333. doi: 10.1038/326332a0. [DOI] [PubMed] [Google Scholar]
- 12.Cavalier-Smith T. McGraw–Hill 1989 Yearbook of Science and Technology. New York: McGraw–Hill; 1988. pp. 175–179. [Google Scholar]
- 13.Cavalier-Smith T. Microbiol Rev. 1993;57:953–994. doi: 10.1128/mr.57.4.953-994.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Schlegel M. Trends Ecol Evol. 1994;9:330–335. doi: 10.1016/0169-5347(94)90153-8. [DOI] [PubMed] [Google Scholar]
- 15.Patterson D J. In: Progress in Protistology. Hausmann K, Hülsmann N, editors. Stuttgart: Fischer; 1994. pp. 1–14. [Google Scholar]
- 16.Gupta R S, Singh B. Curr Biol. 1994;12:1104–1114. doi: 10.1016/s0960-9822(00)00249-9. [DOI] [PubMed] [Google Scholar]
- 17.Hartl F U, Hlodan R, Langer T. Trends Biol Sci. 1994;19:20–25. doi: 10.1016/0968-0004(94)90169-4. [DOI] [PubMed] [Google Scholar]
- 18.Boorstein W R, Ziegelhoffer T, Craig E A. J Mol Evol. 1994;38:1–17. doi: 10.1007/BF00175490. [DOI] [PubMed] [Google Scholar]
- 19.Viscogliosi E, Philippe H, Baroin A, Perasso R, Brugerolle G. J Eukaryotic Microbiol. 1993;40:411–421. doi: 10.1111/j.1550-7408.1993.tb04935.x. [DOI] [PubMed] [Google Scholar]
- 20.Segal G, Ron E Z. J Bacteriol. 1995;177:5952–5958. doi: 10.1128/jb.177.20.5952-5958.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Philippe H. Nucleic Acids Res. 1993;21:5264–5272. doi: 10.1093/nar/21.22.5264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Adachi, J. & Hasegawa, M. (1992) molphy: Programs for Molecular Phylogenetics (Inst. of Stat. Math., Tokyo), Vol. 1.
- 23.Swofford, D. L. (1993) paup: Phylogenetic Analysis Using Parsimony (Illinois Nat. Hist. Survey, Champaign), Version 3.1.
- 24.Saitou M, Nei M. Mol Biol Evol. 1987;4:406–425. doi: 10.1093/oxfordjournals.molbev.a040454. [DOI] [PubMed] [Google Scholar]
- 25.Kishino H, Miyata T, Hasegawa M. J Mol Evol. 1990;30:151–160. doi: 10.1007/BF02109497. [DOI] [PubMed] [Google Scholar]
- 26.Katiyar S K, Edlind T D. Mol Biochem Parasitol. 1994;64:33–42. doi: 10.1016/0166-6851(94)90132-5. [DOI] [PubMed] [Google Scholar]
- 27.Johnson P J, d’Oliveira C E, Gorrell T E, Müller M. Proc Natl Acad Sci USA. 1990;87:6097–6101. doi: 10.1073/pnas.87.16.6097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Quon D V, Delgadillo M G, Khachi A, Smale S T, Johnson P J. Proc Natl Acad Sci USA. 1994;91:4579–4583. doi: 10.1073/pnas.91.10.4579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hrdý I, Müller M. J Eukaryotic Microbiol. 1995;42:593–603. doi: 10.1111/j.1550-7408.1995.tb05913.x. [DOI] [PubMed] [Google Scholar]
- 30.Olsen G J, Woese C R, Overbeek R. J Bacteriol. 1994;176:1–6. doi: 10.1128/jb.176.1.1-6.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ludwig W, Neumaier J, Klugbauer N, Brockmann E, Roller C, Jilg S, Reetz K, Schachtner I, Ludvigsen A, Bachleitner M, Fischer U, Schleifer K H. Antonie van Leeuwenhoek. 1993;64:285–305. doi: 10.1007/BF00873088. [DOI] [PubMed] [Google Scholar]
- 32.Eisen J A. J Mol Evol. 1995;41:1105–1123. doi: 10.1007/BF00173192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lecointre G, Philippe H, L, Le Guyader H. Mol Phylogenet Evol. 1993;2:205–224. doi: 10.1006/mpev.1993.1021. [DOI] [PubMed] [Google Scholar]
- 34.Viale A M, Arakaki A K. FEBS Lett. 1994;341:146–151. doi: 10.1016/0014-5793(94)80446-x. [DOI] [PubMed] [Google Scholar]
- 35.Gray M W. Curr Opin Genet Dev. 1993;3:884–890. doi: 10.1016/0959-437x(93)90009-e. [DOI] [PubMed] [Google Scholar]
- 36.Kidwell M G. Annu Rev Genet. 1993;27:235–256. doi: 10.1146/annurev.ge.27.120193.001315. [DOI] [PubMed] [Google Scholar]
- 37.Roger A J, Clark C G, Doolittle W F. Proc Natl Acad Sci USA. 1996;93:14618–14622. doi: 10.1073/pnas.93.25.14618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bozner P. J Parasitol. 1996;82:103–111. [PubMed] [Google Scholar]
- 39.Whatley J M, John P, Whatley F R. Proc R Soc London B. 1979;204:165–187. doi: 10.1098/rspb.1979.0020. [DOI] [PubMed] [Google Scholar]
- 40.Müller M. Symp Soc Gen Microbiol. 1980;30:127–142. [Google Scholar]
- 41.Finlay B J, Fenchel T. FEMS Microbiol Lett. 1989;65:311–314. [Google Scholar]
- 42.Länge S, Rozario C, Müller M. Mol Biol Parasitol. 1994;66:297–308. doi: 10.1016/0166-6851(94)90156-2. [DOI] [PubMed] [Google Scholar]
- 43.Philippe H, Sörhannus U, Baroin A, Perasso R, Gasse F, Adoutte A. J Evol Biol. 1994;7:247–265. [Google Scholar]
- 44.Fenchel T, Finlay B J. Ecology and Evolution in Anoxic Worlds. Oxford: Oxford Univ. Press; 1995. [Google Scholar]
- 45.Embley T M, Finlay B J, Dyal P L, Hirt R P, Wilkinson M, Williams A G. Proc R Soc London B. 1995;262:87–93. doi: 10.1098/rspb.1995.0180. [DOI] [PubMed] [Google Scholar]
- 46.Marvin-Sikkema F D, Kraak M N, Veenhuis M, Gottschal J C, Prins R A. Eur J Cell Biol. 1993;61:86–91. [PubMed] [Google Scholar]
- 47.Clark C G, Roger A J. Proc Natl Acad Sci USA. 1995;92:6518–6521. doi: 10.1073/pnas.92.14.6518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Müller M. In: Energy Metabolism of Amitochondriate Protists, An Evolutionary Puzzle. Schlegel M, Hausmann K, editors. Leipziger Univ., Leipzig, Germany: Ehrenberg-Festschrift; 1996. pp. 63–76. [Google Scholar]
- 49.Falah M, Gupta R S. J Bacteriol. 1994;176:7748–7753. doi: 10.1128/jb.176.24.7748-7753.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]