

Abstract
Aspergillus nidulans contains two functionally distinct fatty acid synthases (FASs): one required for primary fatty acid metabolism (FAS) and the other required for secondary metabolism (sFAS). FAS mutants require long-chain fatty acids for growth, whereas sFAS mutants grow normally but cannot synthesize sterigmatocystin (ST), a carcinogenic secondary metabolite structurally and biosynthetically related to aflatoxin. sFAS mutants regain the ability to synthesize ST when provided with hexanoic acid, supporting the model that the ST polyketide synthase uses this short-chain fatty acid as a starter unit. The characterization of both the polyketide synthase and FAS may provide novel means for modifying secondary metabolites.
Keywords: sterigmatocystin, aflatoxin, mycotoxin, polyketide, hexanoic acid
Fatty acids (FAs) and polyketides (PKs) are structurally dissimilar molecules that are synthesized by the evolutionarily related fatty acid synthase (FAS) and polyketide synthase (PKS), respectively (1). Both types of enzymes facilitate the reiterative condensation of simple carboxylic acids; typically, acetyl-CoA serves as a starter unit and malonyl-CoA serves as an extender unit. In the case of FASs, the β-keto group generated by the condensation is always fully reduced by a cycle of reactions (keto reduction, dehydration, and enoyl reduction) before the next condensation reaction. Some or all of this reduction cycle can be omitted by the PKS and accounts for much of the structural diversity found in this class of natural compounds.
The long-chain FAs produced by known FASs have a primary function in the cell where the vast majority are found incorporated as lipids into membranes or storage bodies. In contrast, many PKs produced by PKSs have obscure or unknown functions in the producing organism and are typically classified as secondary metabolites. However, many PKs have tremendous importance to man in that they display a broad range of useful antibiotic and immunosuppressant activities as well as less desirable phyto- and mycotoxic activities.
Perhaps the most infamous PK is the mycotoxin aflatoxin (AF), which is among the most highly toxic, carcinogenic natural products known (2). AF was first characterized in 1960 following the death of more than a hundred thousand poultry in England that had ingested AF-contaminated peanut meal (3). This discovery quickly led to legislation regulating trade of AF-contaminated agricultural commodities at both the national and international levels. Compliance to these regulations causes the loss of millions of dollars in produce in the United States each year; trade sanctions and health effects likely triple these losses. In Third-World nations that do not have the ways, means, or luxury to destroy contaminated foods, a high incidence of human liver cancers is correlated in certain geographic areas with the ingestion of AFs (4). In fact, it has been shown that AF-induced liver cancer is linked to a specific mutation in the tumor suppressor gene p53 (5–7).
While only a few fungal species synthesize AF, at least 20 species (including Aspergillus nidulans) produce the biosynthetically-related compound sterigmatocystin (ST; Fig. 1). The PK precursor of ST and AF is norsolorinic acid, and was originally assumed to be produced by a PKS catalyzing 10 condensations and two reduction cycles (2, 8). However, the presence of an aliphatic moiety adjacent to a keto group in norsolorinic acid could also be explained by a novel PK assembly mechanism in which the ST/AF PKS used a short-chain FA (hexanoyl-CoA) as the starter unit instead of acetyl-CoA (Fig. 1) (9). This second hypothesis is supported by two lines of evidence. First, feeding studies demonstrated that C6 chains (via hexanoic acid) can be incorporated intact into ST/AF pathway intermediates (9, 10). Second, recent genetic studies showed that genes encoding a FAS homolog are linked to genes previously shown to be required for ST/AF biosynthesis (11, 12). The work presented here demonstrates that the FAS homolog linked to the A. nidulans ST gene cluster is indeed necessary for ST biosynthesis. In addition, we describe a second pair of linked genes in A. nidulans encoding a different FAS homolog and show that they are required for synthesis of long-chain FAs for primary metabolism but are not needed for ST production.
Figure 1.
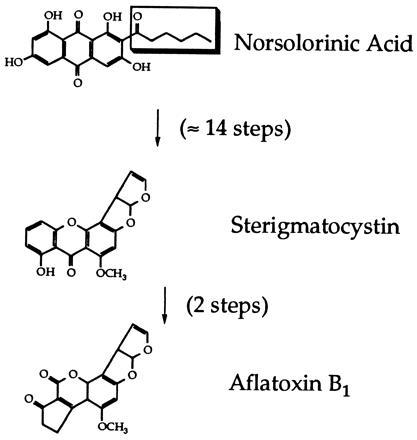
Sterigmatocystin/AF pathway. The aliphatic moiety of norsolorinic acid, the first intermediate in the pathway, is enclosed in a box.
MATERIALS AND METHODS
Fungal Strains and Growth Conditions.
All A. nidulans strains used in this study are described in Table 1. Conidial suspensions (1 × 106 spores per ml) were generated after growth on minimal media (MM) with appropriate supplements (13). FA-requiring strains were grown on MM plus 2 mM myristic acid (n-tetradecanoic acid, Sigma), 0.1% Tween 40 (Sigma), and biotin or MM plus 3.0% oatmeal (Quaker Oats, Chicago) (MMO) and biotin.
Table 1.
A. nidulans strains used in this study
| Strains | Genotype | Source |
|---|---|---|
| FGSC237 | pabaA1, yA2, trpC801, veA1 | FGSC* |
| FGSC89 | argB2, biA1, veA1 | FGSC |
| PW1 | biA1, argB2, methG1, veA1 | Ref. 35 |
| TTAARG | biA1, methG1, veA1 | Ref. 35 |
| TTA11 | pabaA1, yA2, veA1 | Ref. 36 |
| TDB1 | stcJ, biA1, methG1, veA1 | This study |
| TDB5 | stcK, pabaA1, yA2, veA1 | This study |
| TDB51.1 | fasA, biA1, veA1 | This study |
| TDB57.1 | fasB, biA1, veA1 | This study |
| JYP1-13 | biA1, veA1, stcA | Ref. 27 |
Fungal Genetics Stock Center (Kansas City, MO).
Nucleic Acid Manipulations.
Standard molecular techniques were used to manipulate DNA in vitro (14). DNA probes were generated by labeling fragments with 32P by random primer extension as described by Sambrook et al. (14).
ST Production and Detection.
A. nidulans ST production was induced by inoculating 40 ml MMO shake cultures (300 rpm at 30°C) containing appropriate supplements for auxotrophic requirements (Table 1). No FA supplementation was required for fasA− and fasB− mutant strains grown on this medium. For feeding studies, n-hexanoic acid, n-octanoic acid, or n-decanoic acid (20 μg, Sigma) was added to the medium at 24, 48, 72, and/or 96 hr after inoculation. After 120 hr, the cultures were extracted with equal volume acetone followed by equal volume chloroform as described (15). Extracts from the strains indicated were spotted on a precoated TLC plate (Analtech) along with 100 ng of ST standard (Sigma). Samples were resolved using toluene/ethyl acetate/glacial acetic acid (80:10:10 vol/vol/vol). The plates were then dried, sprayed with 20% aluminum chloride in 95% ethanol, and baked at 95°C for 10 min. Although ST could be readily observed as a bright yellow color under fluorescent light, additional UV-absorbing material comigrated with ST. Therefore, the region from each lane having the equivalent retention factor to ST was recovered from the plate, eluted with chloroform, and spotted onto a second TLC plate to reduce the number of comigrating compounds. These samples were resolved with benzene/glacial acetic acid (95:5 vol/vol).
Sequence Analysis.
Approximately 25 restriction endonuclease fragments of the chromosome VIII specific cosmid pW19E9 were inserted into the vectors pBluescript KS (Stratagene) and pK19 (16, 17). DNA sequencing of both strands was performed using synthetic primers and the SequiTherm Cycle Sequencing Kit (Epicentre Technology, Madison, WI) according to the manufacturer’s directions as well as the ABI PRISM DNA Sequencing Kits (Perkin–Elmer). Nucleotide sequence was translated in all six reading frames and compared with the protein data bases (Swiss-PROT and GenBank/European Molecular Biology Laboratory) by the blastx 1.4 program (18, 19). Amino acid sequences were analyzed by the gap, pileup, and pretty programs of the Genetics Computer Group (Madison, WI) sequence analysis software package (20). The GenBank accession no. for fasA and fasB is U75347U75347 and for stcJ and stcK is U34740U34740 (coordinates 25134 to 36275).
Gene Disruption Vectors and Transformation.
Each gene was disrupted by replacing an internal portion of the coding region with a selectable marker using standard transformation procedures (21). The disruption vector pNK17 (targeting stcJ) was created by exchanging a 1.3-kb BamHI fragment in pNK16 with a BamHI fragment containing argB gene from pSalArgB. This results in a truncated StcJ at amino acid 430, effectively removing the ketoreductase and ketosynthase domains. pNK16 is an EcoRI/XbaI subclone of cosmid pL11C09. The disruption vector pDB14 (targeting stcK) was created by exchanging a 2.4-kb XhoI fragment in pDB13 with XhoI fragment containing trpC gene from pTL11. This results in a truncated StcK at amino acid 713, effectively removing the enoyl reductase, dehydrase, and malonyl/palmitoyl transferase domains. pDB13 is a KpnI subclone of cosmid pL11C09.
The disruption vector pDB51 (targeting fasA) was created by using argB to ligate together two plasmids (pDB36 and pDB26) containing distal ends of the coding sequence of fasA such that a 2.2-kb fragment of fasA was replaced with argB. This results in a truncated FasA at amino acid 100, effectively removing all three active site domains (acyl carrier protein, ketoreductase, and ketosynthase domains). The disruption vector pDB57 (targeting fasB) was created in a similar manner replacing a 1.3 kb fragment of fasB with argB. This results in a truncated FasB at amino acid 1390, effectively removing the dehydrase and malonyl/palmitoyl transferase domains.
Identification of Essential FAS Genes.
Highly conserved regions of the α or β subunits of fungal FAS peptides were identified by sequence alignments (to date, all fungal FASs are comprised of α and β subunits). The α and β subunits aligned were from Candida albicans (22, 23), Saccharomyces cerevisiae (24, 25), Penicillium patulum (28), and Yarrowia lipolytica (26). Inosine was substituted at highly degenerate positions within each oligo in some instances to facilitate annealing. The FAS α primers were 5′-GARGGITGYGTIGARATGGCITGGATHATGGG-3′ and 5′-CATCCAIGCNCCIGCNGCICCRTTIGGYTGICC-3′. The predicted PCR product, based on the S. cerevisiae FAS2 gene (α subunit), is 980 bp and encompasses the β-keto acyl synthase domain. The FAS β primers were 5′-CKIRGIACIGCIACYTGCATIGTCATICC-3′ and 5′-GGIWSICARGARCARGGIATGGGIATGG-3′. The predicted PCR product, based on the S. cerevisiae FAS1 (β subunit), is 530 bp and encompasses the malonyl/palmitoyl transferase domain. The PCR conditions for amplification were: 35 cycles of 1 min at 94°C, 30 s at 50°C, and 1 min at 72°C. PCR products were radiolabeled and used to screen a pWE15 cosmid library to identify putative FAS-containing cosmids (17).
RESULTS
stcJ and stcK Are Required for ST Synthesis but Not Growth.
To test the hypothesis that the FAS genes (stcJ and stcK) located within the A. nidulans ST gene cluster (12) functioned specifically in ST biosynthesis, we constructed A. nidulans strains in which each gene had been disrupted. As shown in Fig. 2, stcJ- and stcK-disrupted strains grew without exogenous long-chain FAs, indicating that neither gene is required for normal FA metabolism. In addition, the stcJ− and stcK− strains were morphologically identical to isogenic wild-type strains but failed to produce ST under conditions that supported ST production by wild-type (Fig. 3).
Figure 2.
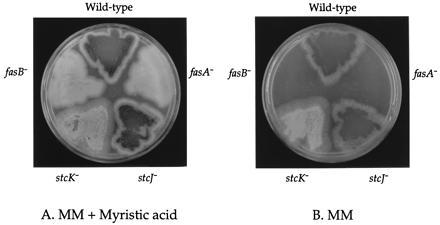
Growth requirements of the strains disrupted in the two FASs. From the top clockwise: TTAARG (wild type), TDB57.1 (fasB−), TDB5 (stcK−), TDB1 (stcJ−), and TDB51.1 (fasA−). Minimal media (B) and minimal media plus 2 mM myristic acid (n-tetradecanoic acid, Sigma) (A).
Figure 3.
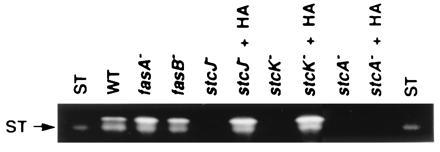
ST analysis of oatmeal culture extracts by TLC. Approximately 20 times as much material from stcJ− and stcK− mutants was loaded as compared with wild type and samples were resolved with benzene/glacial acetic acid (95:5 vol/vol). The nature of the band above ST is unknown.
Addition of Hexanoic Acid Restores ST Biosynthesis.
To further test the hypothesis that stcJ and stcK were required for synthesis of a FA needed for norsolorinic acid synthesis, we added C6 straight-chain FA (hexanoic acid) to stcJ− and stcK− mutant strains grown under ST-producing conditions. As shown in Fig. 3, ST synthesis was restored upon the addition of hexanoic acid to the culture media. In contrast, hexanoic acid failed to restore ST biosynthesis in a stcA− (PKS disrupted) strain (27). This confirmed the need of both FAS and PKS enzymes for ST synthesis.
An alternative source of the C6 carbon-chain (versus de novo synthesis) could be β-oxidation of long-chain FAs. This appears not to be the case as the addition of neither C8 (n-octanoic acid) nor C10 (n-decanoic acid) restored ST synthesis in stcJ− and stcK− strains (data not shown). In addition, we did not observe the appearance of any new metabolites suggesting that the PKS cannot use these FAs, or does so at an efficiency too small to detect the products by TLC.
fasA and fasB Are Required for Growth but Not ST Synthesis.
Our finding that StcJ and StcK [encoding a FAS involved in secondary metabolism (sFAS)] were only required for production of a PK and not required for primary FA biosynthesis led us to investigate the possibility that a different FAS was responsible for long-chain FA synthesis in A. nidulans. The two sets of degenerate FAS primers (specific for the gene encoding either the FAS α or β subunit) generated predicted size fragments from A. nidulans genomic DNA. Direct sequencing of these fragments indicated that they were distinct from analogous stcJ and stcK fragments. The chromosome VIII specific cosmid pW19E9 hybridized to both the FAS α and β specific probes and in fact was found to contain full copies of each gene, which were named fasA and fasB, respectively. Fig. 2 shows that disruption of either fasA or fasB resulted in strains that were unable to grow on minimal medium unless supplemented with myristic acid (C14 straight chain FA). FAs of less than C12 were unable to complement these mutations (data not shown). In contrast to stcJ− and stcK− strains, fasA− and fasB− strains were able to produce ST (Fig. 3).
Comparison of the Essential FAS with the sFAS.
The FAS2 (α subunit) and FAS1 (β subunit) peptide sequences from Penicillium, Candida, Saccharomyces, Yarrowia, and A. nidulans FasA and FasB share between 60% to 70% identity, respectively (Fig. 4). In contrast, the highest identity shared by StcJ (sFAS α) is 44% with Penicillium and the highest identity shared by StcK (sFAS β) is 37% with Yarrowia (12). Nevertheless, the eight active site domains (three on the α subunit and five on the β subunit) required for constitutive FA synthesis in S. cerevisiae were present in both A. nidulans FAS sets and were arranged in the same colinear manner as reported for all fungal FASs. In addition, both sets of genes are divergently transcribed from a shared promoter region; approximately 550 nucleotides separate the putative stcJ and stcK start codons and approximately 1700 nucleotides separate the putative fasA and fasB start codons.
Figure 4.

Amino acid alignments of two representative FAS domains. (A) FAS β-keto acyl synthase domain of the α subunit. Ca, C. albicans (23); Sc, S. cerevisiae (25); Pp, P. patulum (28); and FasA and StcJ, A. nidulans. The constitutive FAS a peptides share 78% identity within this domain in contrast to StcJ that shares only 57% identity. (B) FAS malonyl/palmitoyl transferase domain of the β subunit. Ca, C. albicans (22); Sc, S. cerevisiae (24); Yl, Y. lipolytica (26); and FasB and StcK, A. nidulans. The constitutive FAS β peptides share 72% identity in contrast to StcK that shares 44% identity.
DISCUSSION
The finding that the ST/AF PKs are synthesized through the activity of both a PKS and a FAS defines a novel route for the synthesis of a secondary metabolite. There are several examples in the literature of secondary metabolites derived from products of de novo FA biosynthesis, but the FA source has not been determined and has been assumed to involve FAs from primary metabolism (ref. 29 and references therein). In most cases, these secondary metabolites are not PKs, but it has been suggested that the PKS involved in benastanin (a Streptomyces metabolite) synthesis may use hexanoic acid as a starter unit (29). This hypothesis has not been tested and no information about the possible source of the putative FA starter is available. The finding that a distinct FAS is dedicated to ST/AF metabolite production is novel but is probably not specific to this pathway. For example, it has recently been shown that a FAS with significant identities to fungal FAS1 (encoding the β subunit) is required for production of HC toxin, a tetrapeptide produced by the fungus Cochliobolus carbonum (ref. 30 and J. D. Walton, personal communication). To date, the precise role of the FAS β in HC toxin synthesis has not been demonstrated, but it does not affect growth and is proposed to function in the synthesis of the decanoic acid moiety attached to an amino acid in the tetrapeptide (J. D. Walton, personal communication).
It is interesting that Aspergillus has a unique FAS that is apparently dedicated to the synthesis of a short-chain FA used in secondary metabolism as well as an essential FAS that synthesizes long-chain FAs required for primary growth. The synthesis of short-chain FAs has been best characterized in plants where it occurs through modified branched-chain amino acid metabolism (31). These short-chain FAs have never been shown to be involved in PK synthesis, but are commonly found as components of esters that have poorly characterized functions (31). The synthesis of the hexanoate moiety that likely primes ST PKS clearly occurs by a different mechanism that is more akin to long-chain FA synthesis. Nevertheless, it remains possible that the sFAS is able to synthesize long-chain FAs but fails to complement fasA and fasB mutations because it is only active late in the life cycle. In any case, the sFAS must also have an unprecedented ability to synthesize short-chain FAs.
Considerable efforts have been expended to understand how organisms control or regulate FA chain length. In general, constitutive FA synthesis results in the production of long-chain FAs (>C12). In plants and bacteria, the proportion of different length FAs generated by the type II FAS complex is controlled by the activities of the ketosynthase and acyl-acyl carrier protein thioesterase domains (32). The mechanism regulating FA chain length for the mammalian type I FAS is not clear but it can involve tissue specific control by a distinct thioesterase activity (33). As yet, the mechanism controlling FA chain length in fungi has not been addressed but no known fungal FAS has an obvious thioesterase domain. Thus regulation of FA chain length in fungi is likely controlled by some aspect of the ketosynthase domain or by an uncharacterized activity. Understanding the mechanism allowing sFAS to synthesize short-chain FAs while the essential FAS synthesize long-chain FAs is likely to help define the complex program controlling FA chain length by differing FASs.
Finally, the demonstration of an FA-primed PKS has important implications to molecular engineering of PKs. The ability of the PKS to synthesize ST from exogenous hexanoic acid in sFAS mutants implies that it is able to load this unique length starter without a functional sFAS and then initiate the cycles of condensation reactions leading to norsolorinic acid. Watanabe et al. (34) suggested that the Aspergillus parasiticus ST/AF PKS must interact directly with the ST/AF FAS homologs of StcJ and StcK for efficient incorporation of hexanoate into norsolorinic acid. In support of this hypothesis, stcJ− and stcK− mutant strains grown with hexanoic acid accumulated approximately 20-fold less ST than did wild type grown under the same conditions. While this decreased accumulation of ST could be due to a need for sFAS/PKS interactions, there are other explanations and the clear resolution of this question will require in vitro experiments measuring the activity of all three enzymes. In any case, understanding the mechanism by which the ST/AF PKS and FAS cooperate in synthesizing ST/AF may provide additional tools for engineering PK diversity.
Acknowledgments
We thank Robert Butchko for his assistance with photography and all of our colleagues for many useful discussions. This work was supported by National Research Initiative Competitive Grants Program/U.S. Department of Agriculture Grant 9501256 (D.W.B.), U.S. Department of Agriculture–Cooperative State Research Service Grant 93–37201-9405 (T.H.A. and N.P.K.), Eli Lilly funds (D.W.B., T.H.A., and N.P.K.), and Texas A&M Experiment Station Research Enhancement funds (T.H.A. and N.P.K.).
Footnotes
Abbreviations: FA, fatty acid; FAS, fatty acid synthase; sFAS, FAS related to secondary metabolism; PK, polyketide; PKS, polyketide synthase; AF, aflatoxin; ST, sterigmatocystin.
Data deposition: The sequence reported in this paper has been deposited in the GenBank data base (accession no. U75347U75347).
References
- 1.Hutchinson C R, Fujii I. Annu Rev Microbiol. 1995;49:201–238. doi: 10.1146/annurev.mi.49.100195.001221. [DOI] [PubMed] [Google Scholar]
- 2.Dutton M F. Microbiol Rev. 1988;52:274–295. doi: 10.1128/mr.52.2.274-295.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sargeant K, Sheridan A, O’Kelly J. Nature (London) 1961;192:1095–1097. [Google Scholar]
- 4.Wogan, G. N., Househam, K. C. & Hundt, H. K. (1992) Cancer Res. 52, Suppl., 2114S–2118S. [PubMed]
- 5.Hsu I C, Metcalf R A, Sun T, Welsh J A, Wang N J, Harris C C. Nature (London) 1991;350:427–428. doi: 10.1038/350427a0. [DOI] [PubMed] [Google Scholar]
- 6.Bressac B, Kew M, Wands J, Ozturk M. Nature (London) 1991;350:429–431. doi: 10.1038/350429a0. [DOI] [PubMed] [Google Scholar]
- 7.Coursaget, P., Depril, N., Chabaud, M., Nandi, R., Mayelo, V., LeCann, P. & Yvonnet, B. (1993) Br. J. Cancer 1395–1397. [DOI] [PMC free article] [PubMed]
- 8.Bhatnagar D, Ehrlich K C, Cleveland T E. In: Oxidation-reduction Reactions in Biosynthesis of Secondary Metabolites. Bhatnagar D, Lillehoj E B, Arora D K, editors. Vol. 5. New York: Dekker; 1992. pp. 255–286. [Google Scholar]
- 9.Townsend C A, Christensen S B, Trautwein K. J Am Chem Soc. 1984;106:3868–3869. [Google Scholar]
- 10.Brobst S W, Townsend C A. Can J Chem. 1994;72:200–207. [Google Scholar]
- 11.Mahanti N, Bhatnagar D, Linz J E. Appl Environ Microbiol. 1995;62:191–195. doi: 10.1128/aem.62.1.191-195.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Brown D W, Yu J-H, Kelkar H S, Fernandes M, Nesbitt T C, Keller N P, Adams T H, Leonard T J. Proc Natl Acad Sci USA. 1996;93:1418–1422. doi: 10.1073/pnas.93.4.1418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cove D J. Mol Gen Genet. 1976;146:147–159. doi: 10.1007/BF00268083. [DOI] [PubMed] [Google Scholar]
- 14.Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd Ed. Plainview, NY: Cold Spring Harbor Lab. Press; 1989. [Google Scholar]
- 15.Keller N P, Segner S, Bhatnagar D, Adams T H. Appl Environ Microbiol. 1995;61:3628–3632. doi: 10.1128/aem.61.10.3628-3632.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gibson T J, Coulson A R, Sulston J E, Little P F R. Gene. 1987;53:275–281. doi: 10.1016/0378-1119(87)90016-3. [DOI] [PubMed] [Google Scholar]
- 17.Brody H, Griffith J, Cuticchia A J, Arnold J, Timberlake W E. Nucleic Acids Res. 1991;19:3105–3109. doi: 10.1093/nar/19.11.3105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. J Mol Biol. 1990;215:403–410. doi: 10.1016/S0022-2836(05)80360-2. [DOI] [PubMed] [Google Scholar]
- 19.Gish W, States D J. Nat Genet. 1993;3:266–272. doi: 10.1038/ng0393-266. [DOI] [PubMed] [Google Scholar]
- 20.Devereux J, Haeberli P, Smithies O. Nucleic Acids Res. 1984;12:387–395. doi: 10.1093/nar/12.1part1.387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Miller B, Miller K, Timberlake W. Mol Cell Biol. 1985;5:1714–1721. doi: 10.1128/mcb.5.7.1714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhao X J, Cihlar R L. Gene. 1994;147:119–124. doi: 10.1016/0378-1119(94)90050-7. [DOI] [PubMed] [Google Scholar]
- 23.Southard S B, Cihlar R L. Gene. 1995;1:133–138. doi: 10.1016/0378-1119(95)00033-3. [DOI] [PubMed] [Google Scholar]
- 24.Schweizer M, Roberts L M, Höltke H J, Takabayashi K, Höllerer E, Hoffmann B, Müller G, Köttig H, Schweizer E. Mol Gen Genet. 1986;203:479–486. doi: 10.1007/BF00422073. [DOI] [PubMed] [Google Scholar]
- 25.Mohamed A H, Chirala S S, Mody N H, Huang W, Wakil S J. J Biol Chem. 1988;263:12315–12325. [PubMed] [Google Scholar]
- 26.Schweizer E, Kottig H, Regler R, Rottner G. J Basic Microbiol. 1988;28:283–292. doi: 10.1002/jobm.3620280502. [DOI] [PubMed] [Google Scholar]
- 27.Yu J-H, Leonard T J. J Bacteriol. 1995;177:4792–4800. doi: 10.1128/jb.177.16.4792-4800.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wiesner P, Beck J, Beck N F, Ripka S, Müller G, Lucke S, Schweizer E. Eur J Biochem. 1988;177:69–79. doi: 10.1111/j.1432-1033.1988.tb14346.x. [DOI] [PubMed] [Google Scholar]
- 29.O’Hagan, D. (1995) Nat. Prod. Rep. V12, p1–32.
- 30.Nikolskaya A N, Panaccione D G, Walton J D. Gene. 1995;165:207–211. doi: 10.1016/0378-1119(95)00555-k. [DOI] [PubMed] [Google Scholar]
- 31.Kroumova A B, Xie Z, Wagner G J. Proc Natl Acad Sci USA. 1994;91:11437–11441. doi: 10.1073/pnas.91.24.11437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Jaworski J G, Tai H, Ohlrogge J B, Post-Beittenmiller D. Prog Lipid Res. 1994;33:47–54. doi: 10.1016/0163-7827(94)90008-6. [DOI] [PubMed] [Google Scholar]
- 33.Tai M H, Chirala S S, Wakil S J. Proc Natl Acad Sci USA. 1993;90:1852–1856. doi: 10.1073/pnas.90.5.1852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Watanabe C M H, Wilson D, Linz J E, Townsend C A. Chem Biol. 1996;3:463–469. doi: 10.1016/s1074-5521(96)90094-0. [DOI] [PubMed] [Google Scholar]
- 35.Adams J H, Boylan M T, Timberlake W E. Cell. 1988;54:353–362. doi: 10.1016/0092-8674(88)90198-5. [DOI] [PubMed] [Google Scholar]
- 36.Han S, Navarro J, Greve R A, Adams T H. EMBO J. 1993;12:2449–2457. doi: 10.1002/j.1460-2075.1993.tb05899.x. [DOI] [PMC free article] [PubMed] [Google Scholar]