

Abstract
We extend and apply theories of filled foam elasticity and failure to recently available data on foods. The predictions of elastic modulus and failure mode dependence on internal pressure and on wall integrity are borne out by photographic evidence of distortion and failure under compressive loading and under the localized stress applied by a knife blade, and by mechanical data on vegetables differing only in their turgor pressure. We calculate the dry modulus of plate-like cellular solids and the cross over between dry-like and fully fluid-filled elastic response. The bulk elastic properties of limp and aging cellular solids are calculated for model systems and compared with our mechanical data, which also show two regimes of response. The mechanics of an aged, limp beam is calculated, thus offering a practical procedure for comparing experiment and theory. This investigation also thereby offers explanations of the connection between turgor pressure and crispness and limpness of cellular materials.
Cellular solids (solid foams) are the subject of wide study [see, for instance, the book by Gibson and Ashby (1)] because of (i) their superior mechanical properties (for example wood), (ii) their use in sophisticated applications in which weight and strength are important (e.g., helicopter blades), and (iii) their widespread use as energy absorbers and insulators (against both sound and heat). They occur widely in nature: for instance, in all parts of plants, in bones, feathers, tissues, etc. Most foodstuffs are foams, either geometrically open, such as bread and cake, or closed. We shall be concerned with geometrically closed cell, fluid-filled, solid foams, most particularly fruits and vegetables. The structure of solid foams formed from a liquid phase is determined by surface tension, which often serves to concentrate material into the edges of cells. Other cellular materials without a liquid state genesis also often have material concentrated into the edges rather than on the surfaces dividing neighboring cells.
Even when the cells remain geometrically closed, the removal of material from the faces to the edges makes the foam function elastically as if it were a skeleton of interconnecting rods—effectively an extremely open structure. In cellular structures from nature, there is a partial concentration of material to the common edges (rather than faces) of meeting cells: for instance, in iris leaves (1). A carrot too has this feature, to a more limited extent (see Fig. 1.) Henceforth, “open” and “closed” refer to the elastic rather than geometrical character of the solid foam.
Figure 1.

(a) A micrograph, taken in an environmental scanning electron microscope under conditions of saturated vapor pressure, of the cellular structure of a fresh carrot. (For further details, see Experimental section). The structure has a relatively low volume fraction of solid, in the manner of a classical foam. The concentration of material into edges rather than faces is not very pronounced. (b) The same region under 60% nominal compression. Cells are not distorted uniformly, as the weakest ones burst first and collapse. The partial pressure of water vapor in the specimen chamber was lowered slightly to drive off the excess fluid released from bursting cells, as this would obscure the image.
We shall review dry and then wet foam properties and then model the elasticity of foams: in particular, how it is modified by filling with liquid (2). In fact, above a critical strain of the solid, the filling liquid while it is trapped in a cell of a cellular solid forces the walls to stretch rather than to bend as they do when typical dry cellular solids are deformed. The internal pressure in cells then rises and determines the elasticity and failure modes. The constitutive relation between stress and strain for a partially filled cellular solid is derived. It is compared with experiments on vegetables with differing internal (turgor) pressures that confirm the two regimes of response predicted. We illustrate the use of the complex elasticity in an applied macroscopic problem by solving the elastic problem of how a limp beam bends when clamped at one end and deflection force is applied at the other end. It shows how, with such a constitutive relation, there can be a sharp change in response that presumably governs the perception of crispness in aging vegetables.
Dry Foams
The elasticity and failure of dry foams are governed by a number of remarkably universal scaling laws. We discuss these because they underpin the understanding of filled foams. In Extension of Theory and Application to Cellular Food Materials, we extend the Gibson and Ashby model (1) to the case in which there is not concentration of material into edges, but in which the wall strength is that attributable to plates rather than beams. However, we shall focus on filled foams, and it will turn out that the precise extent of this elastic predominance of the edges is not essential. Discriminating between the concentration of material into edges or faces is only vital in dry foams, where it determines the most characteristic elasticity. The critical advance of Gibson and Ashby (1) was to represent a dry foam by a structure of connected rods (see Fig. 2), the rods representing the elastically active parts of the previous figures. The regularity of the representation is not critical, as the accuracy of the resulting scaling laws over six orders of magnitude testifies. All that is important is that rods characteristically bend as beams. For elastic purposes, we ignore all of the other regions of the cell.
Figure 2.

The skeletal structure of a z–x section of a three-dimensional cellular solid. The cell size is a, and the thickness of elastic members is to. A force F is transmitted along elements such as DB, causing deflection of AB and BC by an amount, the strained configurations being shown in heavier lines. The stretch, w, is required in a distorted filled cell in order to preserve volume.
If the thickness of the cell members is to and the cell lineal dimension is a, then the relative density, ρ, for elastically open cells is ρ ∼ (to/a)2. Gibson and Ashby (1) give structures and the evidence for the elastically open and closed classification. The relative density for solid foams that deform by bending lies in the range 0.01–0.3.
The elasticity of foams is dominated by the fact that a given strain can be more easily obtained by applied stresses causing elastic members to deflect, rather than to stretch or compress. Gibson and Ashby (1) use scaling arguments, relying on experiment to determine a single constant of proportionality. We continue in this spirit here when calculating the modulus of filled foams, and the coupling between elasticity and flow.
The deflection, δ, of a beam with a force F applied transversely is
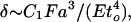 |
1 |
where E is the Young's modulus of the wall material and to is the beam thickness. The constant of proportionality is order unity and depends on the precise beam geometry.
In Fig. 2, F is applied along DB to ABC, causing the deflection. For the foam as a whole, the strain, γ, is δ/a, and from the force per unit area, the stress, σ, is ∼F/a2, whence the modulus Ef is
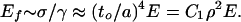 |
2 |
The modulus, if strains in a member were purely compressional or extensional, would be much higher because then the forces, F′ say, give stresses along the beam of F′/to2 ∼ (δ/a)E, whence the modulus of the foam would be Ea,
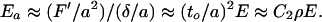 |
3 |
The modulus Ea is now (a/to)Ef, that is, Ea >> Ef, and scales with the area fraction (to/a)2 of the foam that is elastically active. This will be relevant for filled foams. Note that, in Fig. 2, it is the component F that is perpendicular to ABC that is important. For more realistic foams, not all of the force is directed perpendicularly to beams, and there will be some averaged geometrical factor absorbed into the constant C2. One can neglect longitudinal components until they approach the Euler value for the buckling instability of an end-loaded column. Gibson and Ashby (1) examine this question of the elastic (buckling) failure of foams. Elastic buckling failure is perhaps most important in foods, such as when extreme deformations are applied to breads and sponge cakes (3). It will not be important for filled foams. There exist many non-elastic failure modes such as plastic yield, tensile fracture, and brittle fracture. It will turn out below that tensile fracture is most important for the failure of filled foams, so we review the parallel result (1) for dry foams.
When a beam of length a and sectional dimension to is deflected, the maximum stress developed is near the surface and has the value σsurf ∼ (δ/a)(to/a)E. When this stress is equal to the fracture stress σf for the wall material, there is failure in the foam because then the elastic members fail under the applied moments. When the strain is δ/a, the macroscopic stress, using the analysis leading to Eq. 2, is σmac ∼ (δ/a)(to/a)4E, whence the macroscopic failure stress is
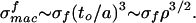 |
4 |
It is much less than the material failure stress because of the concentrating effect of conducting stress through elastic members of the open structure.
A Model for the Elasticity of Filled Foams
If a filled, closed foam is stressed, the resultant strains set up must be such that the incompressibility of a normal liquid is respected. The liquid incompressibility is relative to the comparative ease of stretching wall material. In the event the fluid volume is accommodated by the wall deforming, the elastic scaling laws above are modified. We review the analysis of Warner and Edwards (2) of this problem of elasticity and failure.
Elasticity: it is clear from Fig. 2 that, in bending, a cell is reduced in volume by roughly a2δ. The cell volume is then preserved (see the filled cell) by stretching the cell wall elements such as AB and BC by w, where w ∼ δ + O(δ2). Now the fluid compels a higher energy process (stretch) to occur, and the foam modulus is increased. The force in AB, f, is f/to2 ∼ (w/a)E. This force in members such as AB is spread over a face of a cell of area a2, and the pressure in the cell accordingly rises by an amount p ∼ f/a2 ∼ (w/a)(to/a)2E. Members DF remain straight, unless they are on the boundary of the foam, because the pressure rise in neighboring cells is the same. Taking a slab of foam with external stresses applied on the two large flat faces, the total stress σt applied to a cell is σt ∼ p + F/a2. The modulus, on identifying w with δ before flow takes place and thus identifying w/a as the strain γ, is
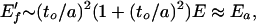 |
5 |
the first contribution being the stretch, the second being the bend found in dry foams. The prime on E′f indicates the modulus is modified by filling. The incompressibility of the fluid causes the modulus of the foam to rise to order Ea, the value found in Eq. 3 appropriate to all of the wall material being elastically active. Changes in volume attributable to F compressing members such as BD is minimal because F is much smaller than the stretching force f.
Failure in filled foams will be amended by the incompressibility of the filling fluid for the same reasons that the modulus was changed. The analysis leading to Eq. 5 and the formula for the maximum stress, σsurf, resulting from bending show that the wall stresses associated with stretch and bend are (δ/a)E and (δ/a)(to/a)E, respectively (for both elastically open and elastically closed foams). Because to/a is very small, we ignore the latter, bend-induced wall stresses when considering rupture. In a filled foam, the macroscopic stress is ∼(δ/a)(to/a)2E. Setting (δ/a)E = σf, the rupture stress characteristic of the wall material, we obtain for the filled foam failure stress:
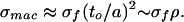 |
6 |
This replaces the empty foam mechanism and failure stress (Eq. 4) for failure.
Extension of Theory and Application to Cellular Food Materials
We modify the dry elasticity of foams to the case in which material is not concentrated into beams but only into plates. We then discuss the elasticity and failure in vegetables and fruits.
Elasticity.
The dry modulus, that attributable to the framework without the intervention of the filling fluid, would appear to be of little practical significance to vegetables. We check here that it is indeed small. It will play a role in determining the constitutive relation for aging food. If the cell walls are elastically plates of thickness to and area a2, rather than being beams, then they will form a much weaker system than rods at the same volume fraction ρ. This is because the stiffness against bending scales as the cube of the thickness, and, in a plate structure, material is spread more thinly. The plate equivalent of Fig. 2 gives deflections δ ∝ Fa3/to3a, whereupon the stress σ = F/a2 and strain γ = δ/a are related as
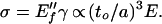 |
7 |
The foam modulus is then E″f = C3ρ3E because the solid volume fraction in a system of plates scales as ρ ∼ to/a. The double prime on E" denotes a modulus caused by plate bending. One must be careful with this result. If plates are curved and constrained (the curved “plates” of an eggshell are a classic example), then the elasticity of their deflections can be subtle for geometrical reasons and they can assume a rigidity greater than that suggested by the above bending analysis. We are assuming that a significant number of plates in our structure are not thus geometrically constrained. In fact, Gibson and Ashby (1) discuss cellular solids composed of plate-like members. They analyze the case in which walls bending cause associated walls to stretch as plates (even in the absence of a filling fluid) and hence to dominate the elasticity to give a modulus E ∝ ρ. We are assuming here that cell walls in deflated cells are floppy and only bend, unless after some deformation they are ultimately constrained by the incompressibility of the filling fluid, whereupon only then do they begin to stretch.
The weakness, manifested by the even higher power, ρ3, than in the Gibson and Ashby case (1), is perhaps a matter of everyday experience—when vegetables lose their internal pressure, they are limp because they presumably derive negligible strength from their own cellular structure. That is, they exhibit a weak, dry-like response because the filling fluid is no longer sufficient to make their walls taut. This is the first stage of elasticity, up to a strain say of γo. We shall speculate that this picture survives until the strain changes the volume sufficiently that the walls again become taut and modulus rises. Then the filled foam analysis of A Model for the Elasticity of Filled Foams is applicable, cell wall stretching giving an enhancement of the modulus from ρ2 or ρ3 to ρ. This is the second regime of elasticity.
If a cell requires a strain γo before the walls have to stretch to accommodate the fluid volume, the stress–strain relation at any given point for such a material must be
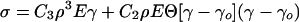 |
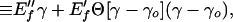 |
8 |
where, until γ = γo, we have the very weak first stage elasticity of the plate assembly (the E″f term) and, for γ ≥ γo, we have, in addition to the bending elasticity, the more dominant ρ elasticity (the second stage, E′f term), where all of the cell material is participating in the stretch. The step function†, Θ, implements the onset of the latter contribution. The threshold strain is a measure of how slack the cell walls are. In the “Modulus” section we compare this relation to simple strain experiment. We used the idea of stretching in the failure analysis (Eq. 6), and this is followed up in the experimental analysis of failure in the “Failure” section.
The Mechanics of a Limp Beam.
Limp solids are seldom deformed in a simple strain geometry that would expose the constitutive relation (Eq. 8) directly as the uniaxial compression experiments in the “Modulus” section do. Rather, the perception of limpness will depend on how a macroscopic object responds to a complex deformation. Nonhomogeneous strains will dictate that different regions of the solid will deform with different moduli, either E″f or E′f. As strains proceed, the balance of the two regions will change, leading to a most unusual macroscopic response. One might imagine, for instance, that a limp carrot being bent will suffer this mixed and changing elasticity. Perception of its state will depend on the balance of the responses. We wish to model this situation.
One can now pose and solve the problem of how a beam, composed of the material with the constitutive relation (Eq. 8), deforms. Take a beam of length L, width a, and thickness T, as in Fig. 3. Near the clamp, y = 0, is where the curvature is greatest, R smallest, and is thus where the crossover to ρ-elasticity starts first. At the deflection end, the torques are smallest and therefore 1/R the least. Thus, the ρ term is absent at this end. For very small deflections, the whole beam deforms weakly with ρ3.
Figure 3.

Bending of a beam. The deflection is Δ, the thickness T, the width a, and the length of the beam L. The transverse deflecting force, F, is applied to one end of the beam. The position, y, is measured from the other, clamped end. For any cross section at a given y, the strain will vary with position x away from the neutral line, being zero on the line itself. Further away from the line, the strain may exceed γo, and the response will be hard. The sections closer to the clamp, at small y, are more distorted, and the crossover γo will be closer to the neutral axis. It is this variation of elastic response across the beam, on going down the beam, that makes the problem of the overall response nontrivial.
We give an analysis for moderate deflections (the full problem reduces to elliptic integrals). In any section, a distance y from the clamped end of the beam, there are extensions and compressions above or below the neutral line that give forces and hence torques balancing that arising from the deflection force F:
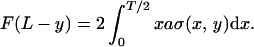 |
9 |
Locally, the strain is γ(x, y) = x/R(y), where 1/R(y) is the curvature of the beam at y along its length, that is d2Δ/dy2 = 1/R(y), with Δ the deflection. We assume for simplicity in Eq. 9 that the material responds similarly in compression and extension.
The shape of the limp beam is now given by the solution R(y) of the equation:
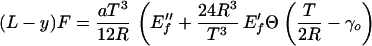 |
10 |
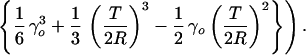 |
Although apparently complicated by the step function Θ, the equation is straightforward to analyze. The strains in a given cross section of a beam are maximal at the surfaces, x = ±T/2. The beam crosses over from a purely ρ3 behavior to a mixture of ρ3 and ρ1 within one cross section when the curvature at this cross section is such that T/2R = γo. Along a beam, this crossover at the surface first occurs at a critical position, yo, given by
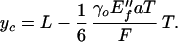 |
11 |
(Note that the combination γoE″f is a characteristic stress that, when multiplied by the cross section aT, yields a characteristic force against which applied forces F are compared. The surviving length in Eq. 11 is T, which sets the scale for crossovers.) At small forces F < 1/6 aTγoE″f T/L such that the crossover is nowhere achieved, the whole beam is classical, being described with the dry, plate-bending foam modulus E″f:
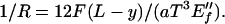 |
12 |
For larger F, the beam profile is described by two equations, the solutions of which are matched at y = yc; that is, by Eq. 12 for y > yc and by
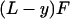 |
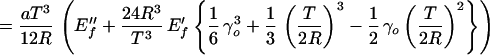 |
13 |
for y < yc. This is a straightforward cubic for R(F). The deflection Δ(y) can be simply derived by integration because Δ(y) = ∫0y dy′ ∫0y′ dy"(1/R(y")).
In applying these predictions of bulk response, one must be careful to consider the possibility of gradients in turgor pressure in limp vegetables, here manifesting themselves as a spatially dependent γo.
Force-Deflection Characteristics of Partially Filled Cellular Solids.
The above analysis, although giving the complete shape of a clamped, limp beam suffering a deflection force while obeying the constitutive relation (Eq. 8), is not explicit enough to offer a practical means of comparison with mechanical experiments on vegetables. We develop an explicit method of proceeding in practice. First, reduce the lengths L, Δ, and R by the thickness T and reduce the maximum strain in a cross section, T/2R, by the transition strain γo to give a reduced strain Γ = T/(2R)/γo. Extract a factor of the hard modulus E′f from the brackets in Eq. 13 and call the reduced modulus E″f/E′f = E. Reduce the force to the dimensionless F̃ = 6F/(aTE′fγo); that is, the force is reduced by 1/6 of the force in a hard beam (E′f) of sectional area aT strained to γo. The moment equation (Eq. 13) for the strain takes the universal form (with L and y now reduced lengths):
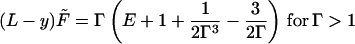 |
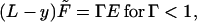 |
14 |
from the solutions of which any system is described because particular systems are recovered by a reversal of the reduction procedure above. Instead of solving the cubic and being faced with numerical integrals of the complicated resulting solutions, one can attack the deflection of the end, Δ(y = L), indirectly.
The reduced form of the deflection is Δ = 2γo ∫0L dy ∫0y dy′Γ(y′). We can change independent variable from y to Γ:
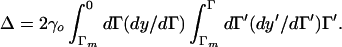 |
15 |
The strain Γm is the maximal occurring for the beam as a whole: that is, at the surface of the section of the beam at the clamp y = 0, where the curvature is the greatest. The Jacobeans of the transformation depend on whether one is below or above the crossover strain, Γ = 1:
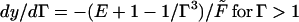 |
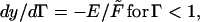 |
16 |
whereupon the intervals of integration must be broken down to accommodate the differing forms (physically where the elasticity is either ρ3 or ρ).
When the force is sufficiently small that the maximum strain in the beam, Γm, is less than the crossover Γ = 1, then the beam is simple, with a reduced deflection Δ = 2γoL3F̃/3E [giving the classical result in unreduced variables of Δ = 4L3F̃/(E″faT3)].
When the force is sufficiently large that the maximum strain in the beam, Γm, is greater than the crossover Γ = 1, one must substitute Eq. 16 into Eq. 15 with some care. One obtains
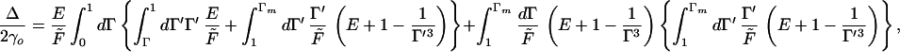 |
17 |
which is simply integrated to give:
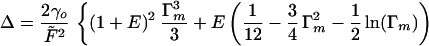 |
18 |
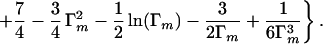 |
When the strain Γm is large, the dominant term in Δ is Γm3, which is again of the classical form, this time for a beam uniformly of modulus E′f—the weak response has been taken out and the hard response that starts in the beam's skin (which is in any case elastically dominant) dictates the response.
The deflection (Eq. 18) Δ(Γm), and the relation for F̃(Γm) from (Eq. 14) at y = 0:
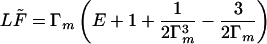 |
19 |
need not be solved for Δ(F̃), which is a complicated procedure. Rather, one can simply plot Δ(Γm) − F̃(Γm) by using Γm > 1 as a parametric variable. An example is shown in Fig. 4, where γo = 0.05, E = 0.05, and L = 10. This is a straightforward procedure, which could be used to compare with experimental examinations of vegetable “beams.” The figure exposes the essential physics of limp beam bending. Until the crossover occurs from all strains in the beam being less than γo to some being greater than γo, that is, at Γm = 1, the beam has F̃ ∝ Δ with a slope reflecting E″f. When parts of the beam have strains sufficient to make these parts respond with E′f, the force required to give deflection rises with Δ faster than linearly because more and more material is responding with E′f. This starts at Δ = 2γoL2/3 (=10/3 for the parameters in Fig. 4). The dotted line is an extrapolation of the weak beam response to illustrate the departure that occurs when γo is exceeded for the first time. After most of the material is reacting with E′f, the limiting strong beam response is attained, and the curve is again linear, but with a much higher slope.
Figure 4.

Reduced force plotted against deflection for a partially filled foam with reduced modulus E = 0.05, transition strain γo = 0.05, and reduced length L = 10. Note the classical linear dependence of deflection force on deflection before the crossover (at Δ = 10/3 for these parameters) and for deflections well beyond where the strain crossover is achieved.
Experimental
All imaging and mechanical testing was performed in an Electroscan Environmental Scanning Electron Microscope, Model 2010. Images were taken with a 15-kV accelerating voltage. An Oxford Instruments custom built tensile stage, capable of cooling the specimen to 3°C, was used for in situ mechanical tests. The specimen chamber was held at 5 torr (1 torr = 133 Pa) of water vapor, thus ensuring that the carrots remained hydrated throughout the experiments. All carrot specimens (Daucus carota) were nominally 6-mm cubes of parenchymal material. Further details on how the mechanical tests were performed can be found in ref. 4.
Modulus.
We present in Fig. 5 stress–strain data for raw carrot and for carrot cooked for 1 minute. The uniform uniaxial compressive strains applied correspond to the situation of the constitutive relation (Eq. 5). At very small strains, both raw and cooked carrot are soft. Raw carrot quickly hardens (a steeper slope to the stress–strain relation) whereas cooked carrot remains softer to larger strains, eventually crossing over to a hard response. It is most noticeable that finally the cooked and raw carrot have the same modulus (equal slopes). This is entirely consistent with the idea that cell walls initially bend and then tighten when the fixed volume of the cell filling causes the cell walls to stretch. The effect of cooking is evidently to reduce the internal filling pressure and, initially at least, leave the walls elastically intact. When stretching dominates and is responsible for the (higher) modulus of the cellular material, the two materials with differing internal pressure (filling) can then take on the same strength (modulus), provided the wall material has not deteriorated. For vegetables cooked longer, or for greater degrees of aging that involve, for instance, degrading the cell walls, this equivalence of the larger strain amplitude moduli is not observed. We believe that Fig. 5 is a qualitative confirmation of our proposed constitutive relation (Eq. 8) for fluid-filled foams. The strains in Fig. 5 are completely reversible. We see from the cyclic compression data of Fig. 8 of ref. 4 that strains of the order of those explored in our Fig. 5 are entirely recoverable.
Figure 5.

Compression stress–strain curves collected during in situ straining of carrot specimens. The cooked carrot specimen was prepared by immersing the 6-mm cube in boiling water for 60 seconds. Note that, at larger extensions, where presumably cell walls are stretching in both cooked and uncooked carrots, the modulus is the same.
Failure.
The predictions (2) of A Model for the Elasticity of Filled Foams are that, under a compressive load, cell walls in directions perpendicular to the compression axis are in effect subjected to extensional strains induced by the incompressibility constraints. The accompanying stresses lead to failures in walls in this direction. This is clearly seen in carrots. The unstrained sample of Fig. 1 is subjected to compression in the vertical direction. Figs. 6 and 7 show the development of a crack in the vertical direction. The crack is caused by the failure attributable to the overextension of horizontal members; the close up shows the failure of horizontal walls leading to a crack propagating vertically.
Figure 6.

The sample of Fig. 1 is subjected to compressive stress along the vertical axis. Near the center, a vertical crack can be seen to be forming. [Reprinted with permission from ref. 4 (Copyright 1998, Annals of Botany Company).]
Figure 7.

Cellular-level close up of Fig. 6. Overstretched horizontal members are seen to have failed. [Reprinted with permission from ref. 4 (Copyright 1998, Annals of Botany Company).]
Further examples of large but localized strains leading to fracture are shown in Fig. 8, micrographs of slicing carrots that are fresh, 1 week old, and 3 weeks old. The behavior is very different according to the internal pressure. In Fig. 8a, cells rupture in the high strain areas closest to the blade tip. Where the internal pressure is lower and the cells are capable of large, easy distortions (with E″f) before stretching of the cell walls occurs, rupture is delayed. Large sections extending far from the blade are seen to distort. This is reported in more detail by Thiel and Donald (4).
Figure 8.

The strong deformation near the tip of a knife slicing a carrot. (a) Fresh carrot. (b) One-week-old carrot. (c) Three-week-old carrot. The extreme deformations in b and c are only possible after fluid has been lost (see ref. 4 for the original analysis). [Reprinted with permission from ref. 4 (Copyright 1998, Annals of Botany Company).]
We should note that not all failure in stressed vegetables and fruit is caused by overstretching of cell walls. It can be observed in aged apples and pears that failure is attributable to the adhesion between neighboring cell walls failing; that is, the failure mechanism is extracellular. The current work does not address this question.
Conclusions
We have presented a range of deformation mechanisms of closed-cell cellular solids, filled and unfilled, to model elasticity and failure in foods. Bending, up to a deformation where the filling fluid's volume is felt, is proposed as a dry foam-like deformation leading to a low modulus. At higher deformations, walls then must stretch, and the modulus rises. A two-part constitutive relation is derived that has a cross-over strain dependent on fluid filling (turgor pressure in plants). Experiments in a matching simple strain geometry confirm two-step response, a cross-over varying as turgor pressure is changed. Remarkable microscopic investigations can now test these ideas, perhaps leading the way for further analysis. We have suggested a way of modeling limpness in vegetables. This takes the form of a model calculation of a limp beam's response. The underlying constitutive relation manifests itself at the macroscopic level in the sharp cross-over from limpness to crispness as the region of “hard” response spreads in the beam. The method can be straightforwardly extended to large amplitude deformations and failure.
Acknowledgments
We thank Drs. Burrows, Gidley, Lillford, and Ormerod of Unilever for valuable advice and for a critical reading of the paper. M. W. is grateful to Unilever Plc for financial support during the period when this theory of filled foams was carried out. B. L. T. acknowledges the support of the Biotechnology and Biological Sciences Research Council for support while this work was carried out.
Footnotes
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.020501797.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.020501797
The step function has Θ(x) = 0 for x < 0 and Θ(x) = 1 for x ≥ 0.
References
- 1.Gibson L J, Ashby M F. Cellular Solids. Cambridge, U.K.: Cambridge Univ. Press; 1999. [Google Scholar]
- 2.Warner M, Edwards S F. Europhys Lett. 1988;5:623–628. [Google Scholar]
- 3.Attenborongh G J. Cereal Sci. 1989;9:61–70. [Google Scholar]
- 4.Thiel B L, Donald A M. Ann Bot (London) 1998;82:727–733. [Google Scholar]