

Abstract
Protein lysine deacetylases play a pivotal role in numerous biological processes and are divided into the Rpd3/Hda1 and sirtuin families, each having members in diverse organisms including prokaryotes. In vertebrates, the Rpd3/Hda1 family contains eleven members, traditionally referred to as histone deacetylases (HDACs) 1–11 and grouped into distinct classes. While most class I HDACs are subunits of multiprotein nuclear complexes crucial for transcriptional repression and epigenetic landscaping, class II members regulate cytoplasmic processes or act like signal transducers trafficking between the cytoplasm and the nucleus. This review focuses on recent findings related to the Rpd3/Hda1 family with an emphasis on evolutionarily conserved domain organization, protein complex formation, regulation, and biological function.
Introduction
Acetylation of the ε-amino group of lysine residues was initially identified on histones four decades ago1 and is now known to occur in different organisms for regulating diverse cellular processes (Box 1). This modification is reversed by lysine deacetylases (KDACs, Box 1). Since histones were first identified as the substrates in 1969 (ref. 2), eukaryotic KDACs have been referred to as histone deacetylases (HDACs). In 1978, butyrate was discovered to be a non-specific HDAC inhibitor3–5, and then the fungal antibiotic Trichostatin A was identified in 1990 as the first specific inhibitor6. Using a similar inhibitor as an affinity tag, Taunton et al.7 purified in 1996 the first HDAC (now known as HDAC1) from cow protein extracts and discovered it to be an orthologue of yeast Rpd3 (reduced potassium dependency 3, Fig. 1a). Because yeast genetics had already established Rpd3 as a global gene regulator8, the unexpected sequence similarity strengthened the link of histone acetylation to gene regulation. Two independent studies confirmed the multiplicity of HDACs9,10 and, together with the identification of HDAC1, set the stage for the establishment of the general notion that these enzymes can function as transcriptional corepressors and spurred the search for additional members of the Rpd3/Hda1 (histone deacetylase 1) family11 (Fig. 1; see also Supplementary information S1 (figure)).
Box 1 Nε-acetylation as a common posttranslational modification
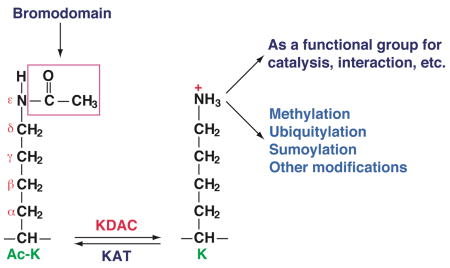
Unlike Nα-acetylation, Nε-acetylation is reversible and is dynamically controlled by opposing actions of lysine acetyltransferases (KATs) and lysine deacetylases (KDACs). This modification has been extensively studied in the past decade. It is now known that Nε-acetylation modifies not only histones and transcription factors (over 60 of them), but also proteins involved in DNA repair and replication, metabolism, cytoskeletal dynamics, apoptosis, protein folding, and cellular signalling131–133. In particular, many mitochondrial proteins appear to be acetylated125,134 (see also Box 2). This modification occurs even in bacteria (Box 2), suggesting that it may evolutionarily precede protein phosphorylation.
Nε-acetylation results in ‘gain-of-function’ and ‘loss-of-function’ effects. In the former case, acetyllysine-containing regions may be recognized by bromodomains in a way that is conceptually similar to the recognition of phosphopeptides by various protein modules135. The ‘loss-of-function’ modification prevents the use of the amino group for different functional purposes. For example, acetylation of a key lysine residue inactivates acetyl-CoA synthetase (Box 2). Acetylation also blocks the amino group from undergoing other modifications such as methylation, ubiquitylation, and sumoylation131. Furthermore, acetylation and neighbouring modifications may counteract or cooperate with one another131.
Figure 1. Domain organization of classical HDACs from yeast and humans.

The deacetylases are grouped into different classes according to sequence similarity to yeast prototypes. In mammals, class I members (Rpd3-like) include HDAC1, 2, 3 and 8; class II members are HDAC4, 5, 6, 7, 9 and 10 (similar to Hda1); and class IV is made up of HDAC11 (Fig. 1). Class II is further divided into two subclasses, IIa (HDAC4, 5, 7 and 9) and IIb (HDAC6 and 10). The total number of amino acid residues in each deacetylase is shown to the right of the figure. Many of the deacetylases have isoforms that result from alternative splicing. For simplicity, the number refers to the longest isoform. The deacetylase (DAC) domain is depicted by a rectangle containing a yellow (class I, (a)), clear (class II, (b)), or light green (class IV, (c)) centre flanked by orange shading, and the percentage amino acid sequence identity/similarity to that of Rpd3 (for class I) or Hda1 (class II/IV) is shown. The sequence identity/similarity of Hda1 and HDAC11 to Rpd3 is given in brackets. The C-terminal domains (shaded rectangles) of Hda1 and Clr3 are homologous (identity/similarity, 26/57%). Dark bars represent similar N-terminal domains and C-terminal tails of class IIa histone deacetylases (HDACs). Myocyte enhancer factor 2 (MEF2)-binding motifs are depicted as small green boxes, whereas 14-3-3 binding motifs are indicated by small boxes labelled with the letter “S” (for serine). SE14, Ser-Glu-containing tetradecapeptide repeats; ZnF, ubiquitin-binding zinc finger.
In 2000, the Sir2 (silent information regulator 2) or sirtuin (Sir two (“tu”)-like protein) family of NAD+-dependent deacetylases was discovered12, which led to the designation of the zinc-dependent Rpd3/Hda1 family as the “classical family”13. HDACs can also be grouped according to phylogenetic analysis and sequence homology14–16. In this classification system, the sirtuin family constitutes class III, whereas the classical family is grouped into class I, II and IV (Fig. 1), with class II being further divided into two subclasses (IIa and IIb); such subclass division is also necessary to accommodate classification of members from C. elegans, D. melanogaster and zebrafish (Supplementary information S1 (figure)).
The HDAC field has been expanding at an amazing pace, owing largely to the following three factors. First, chromatin research has exploded over the past 10–15 years, and HDACs have a key role in deacetylating nucleosomes, interacting with other chromatin regulators and shaping epigenetic landscapes17,18. Second, in addition to histones, these enzymes deacetylate numerous transcription factors and many other proteins that are important for a multitude of cellular processes in eukaryotic and prokaryotic systems (Boxes 1 & 2). Finally, HDAC inhibitors have great potential for treating cancer and other diseases19,20. A majority of compounds that are currently under clinical evaluation target the Rpd3/Hda1 family.
Box 2 Acetylation inhibits acetyl-CoA synthetase from bacteria to humans
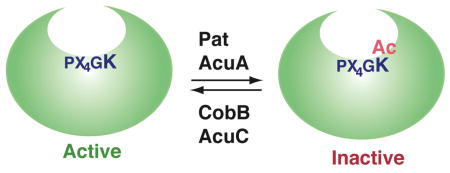
Acetyl-CoA synthetase (Acs) is a ubiquitous metabolic enzyme conserved from bacteria to humans. It is responsible for the conversion of acetate to acetyl-CoA. In 2002, the Acs from Salmonella enterica was discovered to be acetylated on K609, a key catalytic residue136. The modification inhibits the enzymatic activity, thereby regulating the growth of S. enteric on propionate and low concentration of acetate as carbon and energy sources. K609 is evolutionarily conserved in related enzymes, implying that this regulatory mechanism is important in different organisms. Indeed, the Acs orthologues inBacillus subtili and mammals are similarly modified137–139.
Two acetylation–deacetylation systems are known to regulate acetylation in bacteria: Pat (protein acetyltransferase) and CobB (cobalamin biosynthetic gene product B, a Sir2 homologue) inS. enteric, and the acetoin utilization A (AcuA) and AcuC proteins inB. subtili (see figure)137. AcuA and AcuC show some sequence similarity to yeast Gcn5 and Rpd3, respectively137. In addition to AcuC, other bacterial proteins show sequence similarity to members of the Hda1/Rpd3 family16. Although the functional significance of this similarity remains to be addressed, it implies that this family plays a role even in prokaryotes.
This review covers mainly the recent findings relating to this family of deacetylases. We begin with an overview of domain organization and tertiary structure of the mammalian members and compare this organization to that of homologues in yeast and other model organisms. We then describe multisubunit complexes of class I/II HDACs, roles of class IIa members in signal transduction, and cytoplasmic functions of HDAC6. After that, we discuss how these molecular and cellular studies are related to genetic analyses in mice, C. elegans and D. melanogaster, as well as the implications for pathology and drug development. Two themes link different sections of this review: first, how amino acid sequences of the deacetylases determine the structure, function and regulation; and, second, how a simple catalytic domain co-evolves with different motifs and modules to connect functions in diverse cellular processes from various organisms. Both themes are fundamental issues relevant to many other protein families.
Domain organization of class I/II/IV HDACs
Domains and motifs of class I HDACs
As a founding member of class I, Rpd3 from budding yeast comprises an N-terminal deacetylase domain and a C-terminal tail8,10 (Fig. 1a). Both regions are conserved in Clr6 (cryptic locus regulator 6) from Schizosaccharomyces pombe21, but only the catalytic domain is conserved in the bacterial deacetylase AcuC (Box 2) and in class I members from higher eukaryotes13. Mammalian HDAC1 and HDAC2 are almost identical, and in addition to the catalytic domains, they are homologous in the C-terminal tail, which harbors tandem casein kinase 2 (CK2) phosphorylation sites22. The tail of HDAC1 also contains a sumoylation motif22. There are three orthologues in C. elegans, but only one in D. melanogaster or zebrafish (Supplementary information S1 (figure)). Unlike HDAC1, HDAC3 is only conserved from flies to mammals (Fig. 1; and Supplementary information S1 (figure)). Slightly different from HDAC1 and HDAC2, HDAC3 possesses one CK2 phosphorylation site23. HDAC8 is present in sea urchin, but not in C. elegans or D. melanogaster, suggesting that this deacetylase is deuterostome-specific. Instead of CK2 sites, this deacetylase has a conserved motif (Supplementary information S2a (figure)) for protein kinase A phophorylation and negative regulation of deacetylase activity24,25.
Domains and motifs of class II HDACs
Hda1 consists of an N-terminal deacetylase domain and a long C-terminal extension of unknown function (Fig. 1b). This domain organization is conserved in Clr3, a class II member from S. pombe. By contrast, only the deacetylase domain is shared by metazoan homologues (Fig. 1b)14,15,26. In addition to the deacetylase domain, a long N-terminal extension is conserved among the class IIa members in mammals (HDAC4, 5, 7, and 9). This N-terminal extension contains binding sites for myocyte enhancer factor 2 (MEF2) and 14-3-3 proteins. The MEF2-binding motif is conserved from C. elegans to mammals (Supplementary information S1 (figure))26 and displays resemblance to sites in other MEF2-binding proteins27. Only one or two of the 14-3-3 binding sites are conserved in C. elegans and D.melanogaster (Supplementary information S1 (figure)), but three are present in an orthologue from sea urchin (Supplementary information S2b (figure)). Consistent with the sequence conservation, MEF2 and 14-3-3 are major partners of class IIa HDACs.
HDAC3 was shown to be responsible for deacetylase activities associated with HDAC4 and HDAC7 (ref. 28). Consistent with this, mammalian class IIa members possess low deacetylase activity as a result of a Tyr-His substitution29. This substitution occurs in zebrafish, but not in C. elegans, D. melanogaster or sea urchin (Supplementary information S2b (figure)), which implies that it is vertebrate-specific. Interestingly, an equivalent tyrosine residue is dispensable for the esterase activity of a bacterial HDAC-like protein30. It will be important to determine how the substitution is reflected with regard to the function and regulation of class IIa deacetylases in vertebrates.
Unlike class IIa members, HDAC6 contains tandem deacetylase domains and a C-terminal zinc finger (Fig. 1b & Supplementary information S1 (figure)), and has emerged as a major cytoplasmic deacetylase. The zinc finger can bind ubiquitin, and its core sequence is similar to ZnF-UBP (ubiquitin carboxyl-terminal hydrolase-like zinc finger)26,31. An SE14-repeat domain that is found in human HDAC6 (ref. 26) is almost identical in macaques (XP_001101619) and highly conserved in horses (GenBank Accession No., XP_001493915), but is only partial in mice, implying that this domain is specific to high mammals.
The other class IIb member, HDAC10, has an N-terminal half that is highly similar to the first deacetylase domain of HDAC6, whereas the C-terminal half is leucine-rich (Fig. 1b). Although there are no invertebrate orthologues, a distant relative exists in zebrafish (Supplementary information S2c (figure)), which indicates that HDAC10 might have a vertebrate-specific role. However, very little is known about its real biological function.
HDAC11 as a highly conserved deacetylase
HDAC11 shows sequence similarity to class I and II HDACs (Fig. 1c)32. It is highly conserved from C. elegans and D. melanogaster to humans (Supplementary information S1 (figure)), and there are also related proteins in bacteria16 and plants (atHDA2, NP_568480). HDAC11 is sometimes referred to as a class I member, but phylogenetic analysis indicated that this deacetylase and its homologues belong to a separate class16. Little is known about its function or regulation, but its evolutionary conservation implies a fundamental role in diverse organisms.
Structures of Zn2+-dependent deacetylase domains
Crystal structures have been solved for the catalytic domains of HDAC8 (refs 25,33), HDAC7 (PDB code, 2NVR) and two bacterial deacetylases34,35. The structures are very similar: a single α/β domain composed of an eight-stranded β-sheet sandwiched between over a dozen α-helices (also see Supplementary information S3 (figure)). The catalytic centre contains a zinc ion chelated by side chains of histidine and aspartate residues. These structures indicate that the deacetylase domains from these family members share not only sequence similarity but also overall tertiary structure. Additional domains/motifs and sequence divergence within the deacetylase domains form the molecular basis for the unique subunit associations, and biological function, of each individual family member (see below).
Diversity of class I/II HDAC complexes
With the exception of HDAC8, all class I members can be catalytic subunits of multiprotein complexes (Table 1 and Supplementary information S4 (table)). In mammals, HDAC1 and HDAC2 interact with one another and form the catalytic core of several multisubunit complexes11. This HDAC1-2 core is present in mammalian Sin3, nucleosome remodeling deacetylase (NuRD) and corepressor of REST (repressor element 1 silencing transcription factor (CoREST)) complexes (Table 1 and Supplementary information S5 (table)). As discussed below, one emerging theme is that SANT domain-containing subunits stimulate deacetylase activities36. Another theme is that the subunits with which HDACs associate possess chromatin-remodelling or chromatin-binding ability, which provides an important platform for coordinating deacetylase functions with other chromatin-regulating mechanisms.
Table 1.
Class I HDAC complexes from different eukaryotic organisms
| Complex | Mammal | Drosophila | C. elegans | S. cerevisiae | S. pombe | Arabidopsis | Protein domain | ||
|---|---|---|---|---|---|---|---|---|---|
| Rpd3L | Rpd3S | I | II | ||||||
| Sin3 | HDAC1/2 | dRpd3 | ceHda-1 | Rpd3 | Rpd3 | Clr6 | Clr6 | Rpd3 | Class I deacetylase |
| RbAp46/48 | p55 | Rba-1, Lin-53 | Ume1 | Ume1 | Prw1 | Prw1 | WD40 repeat | ||
| Sin3A | dSin3 | Sin3 | Sin3 | Sin3 | Pst1 | Pst2 | AtSin3 | PAH motifs | |
| Sds3/BRMS1 | Sds3 | Sds3 | Sds3 | Coiled coil | |||||
| RBP1 | |||||||||
| SAP30 | Sap30 | ||||||||
| SAP18 | AtSap18 | Ubiquitin-fold | |||||||
| ING1/2 | Pho23 | Png2 | PHD finger | ||||||
| Rco1 | PHD finger | ||||||||
| Eaf3 | Chromodomain | ||||||||
| Cph1 | PHD finger | ||||||||
| Cph2 | PHD finger | ||||||||
| Alp13 | Chromodomain | ||||||||
| Mi2/NuRD | HDAC1/2 | dRpd3 | ceHda-1 | Rpd3 | Rpd3 | Clr6 | Clr6 | Rpd3 | Class I deacetylase |
| RbAp46/48 | p55 | Rba-1, Lin-53 | WD40 repeat | ||||||
| Mi-2α/β | dMi-2 | Chd-3/-4 | Pickle | Helicase | |||||
| MTA1/2/3 | Egl-1, Egl-27 | SANT domain | |||||||
| MBD3 | Methyl CpG binding | ||||||||
| p66α/β | p66 | ||||||||
| CoREST | HDAC1/2 | dRpd3 | ceHda-1 | Rpd3 | Rpd3 | Clr6 | Clr6 | Rpd3 | Class I deacetylase |
| CoREST | dCoREST | Spr-1 | SANT domain | ||||||
| LSD1 | Su(Var)3-3 | Spr-5 | SWIRM, K-demethylase | ||||||
| BHC80 | PHD finger | ||||||||
| CtBP1 | Dehydrogenase | ||||||||
| N-CoR/SMRT | HDAC3 | dHDAC3 | Class I deacetylase | ||||||
| N-CoR/SMRT | SMRTER | SANT domain | |||||||
| TBL1/TBLR1 | WD40 repeat | ||||||||
| GPS2 | |||||||||
| JMJD2A | Jumonji demethylase | ||||||||
| Kaiso | Methyl CpG binding | ||||||||
Notes: For the mammalian and yeast complexes, only those subject to biochemical purification are listed here. For complexes in Drosophila, C. elegans and Arabidopsis, little biochemical purification has been performed, so the listed subunits are based on sequence and functional similarity with counterparts in yeast and mammals. In the CoREST complex, there is only a sub-stoichiometric amount of CtBP1, and so are JMJD2A and Kaiso in N-CoR/SMRT complexes.
Abbreviations used (in an alphabetical order): BHC80, an 80-kDa subunit of the BRAF-histone deacetylase complex; BRMS1, breast cancer metastasis suppressor 1, similar to SDS3; ceHda-1, C. elegans HDAC1-related protein; CoREST, corepressor of REST (repressor element 1 silencing transcription factor); CtBP1, E1A C-terminal binding protein 1; Eaf3, Esa1-associated factor 3; Hda1, histone deacetylase 1; ING1/2, inhibitor of growth 1 or 2; JMJD2A, jumonji demethylase D group member 2A; LSD1, lysine-specific demethylase 1; MBD3, methyl CpG-binding domain 3; Mit1, Mi2-like protein interacting with Clr three 1; MTA1, metastasis-associated protein 1; Mi2, dermatomyositis-specific autoantigen; NuRD, nucleosome remodeling deacetylase complex; PHD finger, plant homeodomain-linked zinc finger; PAH motif, paired amphipathic helix motif; Rba-1, RbAp48-related protein; Pst1, S. pombe SIN three 1; RBP1, Rb-binding protein 1; SANT domain: Swi3, Ada2, N-CoR, and TFIIIB homologous domain; SAP30, Sin3-associated polypeptide of 30 kDa; SDS3, suppressor of defective silencing 3; N-CoR, nuclear receptor co-repressor; Sin3, SWI-independent 3 (also known as Rpd1, for reduced potassium dependency 1); SMRT, silencing mediator of retinoic acid and thyroid hormone receptors; Su(var)3-3, suppressor of position-effect variegation 3-3; SWIRM domain, Swi3p, Rsc8p and Moira homologous domain; TBL1, transducin β-like protein 1; WD40 repeat, tryptophan-aspartate 40 repeat.
Sin3 complexes from fungi and plants to mammals
S. cerevisiae Rpd3 forms large and small Sin3 complexes, known as Rpd3L and Rpd3S, respectively (Table 1 and Supplementary information S4 (table))10,37–40. The former typically possesses the subunits Ume1, Sds3, Sap30, and Pho23 (an ING (inhibitor of growth) protein). DNA-binding proteins, such as the zinc-finger repressor Ume6, target it to specific loci for transcriptional repression41. Rpd3S shares Sin3 and Ume1 with Rpd3L, but also contains two unique subunits, Eaf3 and Rco138,39. Eaf3 contains a chromodomain for recognizing K36-methylated histone H3 and recruits Rpd3S to nucleosomes that have been methylated by the Set2 histone methyltransferase38,39,42. The recognition efficiency depends on the plant homeodomain-linked (PHD) finger of Rco143. Whereas Rpd3L deacetylates histones at promoter regions, Rpd3S targets transcribed regions to suppress intragenic transcription initiation38,39.
S. pombe has three Sin3 orthologues, two of which have been shown to form deacetylase complexes. One associates with Clr6 to form two slightly different complexes21, both of which share two other subunits, Sds3 and Prw1; however, only one of the complexes contains the ING protein Png2. These complexes silence mating-type genes and are required for cell viability. The second Sin3 orthologue, Pst2, forms a third Clr6 complex with Prw1 and two PHD finger-containing proteins, one of which possesses a helicase domain. This complex reverses K9 acetylation of histone H3 and blocks antisense transcription21.
Mammals express two Sin3 proteins, Sin3A and Sin3B. The Sin3A complex contains HDAC1-2 catalytic core and RbAp46/48 (Table 1 and Supplementary information S5 (table))44, as well as RBP1 (Rb-binding protein 1), SAP180 (similar to RBP1), SDS3, BRMS1 (breast cancer metastasis suppressor 1, similar to SDS3), SAP30, SAP18, and ING1/2. As some of the subunits have different isoforms, there are several distinct Sin3A complexes. Interestingly, ING2 recognizes K4-methylated histone H3 and targets the Sin3A complex to deacetylate nucleosomes in a methylation-dependent manner45. The Sin3B complex shares some similarity to the Sin3A complex but might also contain distinct subunits46. Sin3 homologues are present in C. elegans, D. melanogaster and Arabidopsis thaliana (Table 1), which indicates that similar complexes might exist in various eukaryotes.
Metazoan and plant Mi-2 and NuRD
NuRD contains a catalytic core of HDAC1, HDAC2, RbAp46, and RbAp48 (Table 1 and Supplementary information S5 (table))44,47. In addition, this complex contains MTA (metastasis-associated) proteins, the nucleosome-remodelling ATPase Mi-2 (dermatomyositis-specific autoantigen), MBD3 (methyl CpG-binding domain 3), and p66. Through MBD3, this complex is recruited for DNA methylation-dependent gene silencing. It also associates with MeCP1 (methyl CpG-binding protein 1) and MeCP2 to provide an intimate connection with DNA methylation47,48. Three mammalian MTA isoforms (MTA1, 2 and 3) have been identified47. They possess SANT domains that are crucial for the integrity and deacetylase activity of NuRD. Mi-2 and p66 have α and β isoforms, enabling the formation of distinct complexes. NuRD subunits have been identified in C. elegans, D. melanogaster and A. thaliana (Table 1). Similar complexes probably exist, therefore, in animal and plant kingdoms.
CoREST complex
Initially identified as a corepressor of the transcriptional repressor REST49, CoREST is a component of an HDAC1-2 complex referred to as a neuronal corepressor complex (Table 1 and Supplementary information S5 (table))50–52. One subunit, KIAA0601/BHC110, possesses an FAD-binding domain with sequence homology to maize polyamine oxidase51. This subunit was recently shown to be a demethylase for K4-methylated histone H3 and therefore renamed lysine-specific demethylase 1 (LSD1)53. The CoREST complex promotes nucleosomal deacetylation through its SANT domains54. Interestingly, histone demethylation stimulates the deacetylase activity, which, in turn, is required for optimal demethylation, indicating that the two activities are interdependent55. CoREST subunits have been found in C. elegans and D. melanogaster (Table 1), but not in fungi or plants. This complex might, therefore, be metazoan-specific, which is consistent with its primary role in repressing neuronal gene expression.
HDAC3 complexes: from flies to men
Biochemical purification of HDAC3 uncovered an unexpected link to two homologous nuclear receptor corepressors, nuclear receptor co-repressor (N-CoR) and silencing mediator of retinoic acid and thyroid hormone receptors (SMRT) (Table 1 and Supplementary information S5 (table))56–58. Conversely, HDAC3 was identified through affinity purification of both corepressors59–61. The N-CoR and SMRT corepressors stimulate the deacetylase activity of HDAC3 through their SANT domains62, which are highly conserved in SMRTER, the D. melanogaster homologue of SMRT and N-CoR. An HDAC3 orthologue exists in D. melanogaster, suggesting that SMRTER might form a functional deacetylase complex. Similar proteins are absent from C. elegans and plants, so HDAC3 complexes might only be conserved from flies to mammals.
Class II HDAC complexes
Yeast Hda1 self-associates and forms a tetrameric complex with Hda2 and Hda3, in which both are important for the deacetylase activity63. Hda2 and Hda3 homologues are present in Candida albicans, but not in S. pombe or higher organisms. Instead, Clr3 forms a quartet core complex in S. pombe64. One subunit, Mit1, contains a PHD finger and a helicase domain, thus conferring both deacetylase and chromatin-remodelling activities within one complex64 (somewhat reminiscent of the NuRD complex in metazoans). The Clr3 complex is recruited to euchromatic and heterochromatic loci for transcriptional gene silencing through Mit1, a heterochromatin protein 1 (HP1) homologue and a telomere-binding protein64.
Non-catalytic subunits of Hda1 and Clr3 complexes are not conserved in metazoans, so these complexes might be fungi-specific. This is consistent with the fact that metazoan class II HDACs possess unique domains and motifs absent in Hda1 and Clr3 (Fig. 1 and Supplementary information S1 (figure)). Indeed, affinity purification of mammalian class IIa HDAC complexes identified 14-3-3 proteins65 and HSP70 (E.S. & X.J.Y., unpublished results), whereas HDAC6 was found to associate with cytoskeletal proteins and chaperones66,67. As discussed below, 14-3-3 proteins play a major role in regulating the trafficking of class IIa HDACs, and the association partners of HDAC6 shed important light on its important functions in the cytoplasm.
Class IIa HDACs as signal transducers
Nucleocytoplasmic trafficking mediated by 14-3-3
Depending on the cellular context, mammalian class IIa members (HDAC4, 5, 7, 9) can exist in both the nucleus and the cytoplasm14,15,26. They possess intrinsic nuclear import and export signals for dynamic nucleocytoplasmic trafficking, so the association of partners might alter this equilibrium and the subsequent subcellular distribution. MEF2 binds to class IIa HDACs and promotes their nuclear localization. By contrast, 14-3-3 proteins stimulate their cytoplasmic retention in a phosphorylation-dependent manner15,26. HDAC4 contains three 14-3-3-binding sites, S246, S467 and S632, all of which are conserved in HDAC5, 7 and 9 (Fig. 1b). Only the former two sites seem to be functional in HDAC5 and HDAC968,69, while a fourth site might be important in HDAC7 (Fig. 1b)70. Mutational analysis revealed that the S246 site of HDAC4 is the most important15,26. This site is conserved in both C. elegans and D. melanogaster (Fig. 1b and Supplementary information S1b (figure)).
Both 14-3-3 binding and the presence of a nuclear export signal are necessary for the cytoplasmic localization of HDAC4 and paralogues in mammals. The mechanistic impact of 14-3-3 binding is multifaceted (Fig. 2). First, the S246 site is adjacent to the nuclear localization signal (NLS, Fig. 2a), so 14-3-3 might impede access of the nuclear import receptor importin α/β to the NLS. Second, 14-3-3 binding might provide a signal in trans to promote the nuclear export of class IIa HDACs. Third, 14-3-3 binding affects the subnuclear distribution of class IIa HDACs. For example, upon binding to 14-3-3 proteins, an HDAC9 isoform lacking the deacetylase domain and nuclear export signal disperses from nuclear punctae into a uniformly nuclear pattern71. Similarly, 14-3-3 binding to C. elegans HDAC4 (Supplementary information S1a (figure)) does not seem to promote cytoplasmic localization72, perhaps due to the lack of a nuclear export signal in this deacetylase. Fourth, 14-3-3 association protects HDAC7 from proteasomal degradation73,74, which contributes to its cytoplasmic accumulation. Finally, 14-3-3 binding sites are subject to reversible phosphorylation. By binding to phosphorylated residues, 14-3-3 binding might occlude phosphatases, and, therefore, inhibit dephosphorylation, which might function as a feed-forward reinforcing mechanism70.
Figure 2. HDAC4 and homologues as signal transducers.

a Multiple kinases (calcium/calmodulin-dependent protein kinase (CaMK), protein kinase D (PKD), microtubule affinity-regulating kinase (MARK), salt-inducible kinase (SIK), checkpoint kinase 1 (CHK1) and other kinases) mediate specific phosphorylation of human HDAC4 on three 14-3-3-binding sites. Myosin phosphatase-targeting subunit-1 (MYPT1)/protein phosphatase (PP)1β and PP2A can act on these sites. The association of 14-3-3 proteins with HDAC4 retains it in the cytoplasm and prevents its interaction with transcription factors such as myocyte enhancer factor 2 (MEF2), thereby releasing them for transcriptional activation. This model can also be extended to other class IIa members, although some of the kinases and phosphatases have only been shown for HDAC4, 5, 7 and/or 9 (see text for references). At the bottom of the panel, the sequence surronding the major 14-3-3 binding site (S246) of HDAC4 is compared with the corresponding regions of its paralogues (HDAC5, 7 and 9) as well as orthologues from D. melanogaster and C. elegans. The key residues crucial for phosphorylation and 14-3-3 binding are highlighted. This 14-3-3 site constitutes a sequence conservation island140. The nuclear localization signal, which can be impeded by 14-3-3 binding, is also shown. b Signal-responsive transcriptional repression by HDAC4 (left) is analogous to transcriptional activation by transforming growth factor-β (TGFβ)-stimulated SMADs (middle) and cytokine-activated signal transducer and activator of transcription (STAT) proteins (right). 14-3-3 proteins bind to HDAC4 and retain it in the cytoplasm, thereby inhibiting its corepressor activity; upon dephosphorylation, HDAC4 translocates to the nucleus for transcriptional repression. The letter P circled in red denotes phosphorylation. For simplicity, only two 14-3-3 molecules are shown. This schematic also applies to HDAC5, 7 and 9.
Kinases and phosphatases for class IIa HDACs
Five groups of kinases have been shown to phosphorylate the 14-3-3-binding sites on HDACs: Ca2+/calmodulin-dependent kinases (CaMKs)68,75, protein kinase D76–78, microtubule affinity-regulating kinases70,79, salt-inducible kinases72,80, and checkpoint kinase-1 (CHK1) (Fig. 2a)81. Conversely, two phosphatases have been identified, myosin phosphatase-targeting subunit-1 (MYPT1)/protein phosphatase (PP)1β82 and PP2A70,83,84. Available data indicate that most of the kinases phosphorylate all class IIa members in mammals. The S246 site is conserved from C. elegans and D. melanogaster to mammals (Supplementary information S1 (figure)), and most of the kinases have orthologues in invertebrates, indicating that the regulatory mechanisms might be evolutionarily conserved. Consistent with this, the salt-inducible kinase orthologue Kin-29 in C. elegans phosphorylates HDAC4 (Supplementary information S1 (figure)) and promotes 14-3-3 binding72. The nuclear function of class IIa HDACs seems to be regulated by different signalling pathways through reversible phosphorylation at 14-3-3 binding sites. In this regard, these deacetylases function as novel signal transducers, very similar to the STAT and SMAD signal-responsive transcription factors (Fig. 2b). Mapping out events upstream of the kinases (Fig. 2a) will be necessary to determine how the regulation is integrated in cellular signalling networks under various physiological contexts.
Other regulatory mechanisms for class IIa HDACs
In addition to phosphorylation, class IIa HDACs are subject to ubiquitylation and sumoylation73,85,86. Ubiquitylation promotes protein degradation73,74, but the functional consequence of sumoylation is less clear85,87. HDAC4 sumoylation is very efficient, which makes it a potentially important modification. One possibility is that this modification generates a transcriptional repression domain and increases the transcriptional repression potential of HDAC4. In addition, this deacetylase is subject to non-reversible processing. For example, caspase cleavage produces an HDAC4 fragment corresponding to the N-terminal 289 residues88,89, within which there is a transcriptional repression domain26. Moreover, alternative splicing of HDAC9 mRNA produces isoforms that contain the N-terminal repression domain86. The HDAC4 fragment and some of the HDAC9 isoforms possess an intact MEF2-binding site and can still function as transcriptional corepressors, supporting that class IIa HDACs can repress transcription independent of their cryptic deacetylase activity28,29. Regulating the enzymatic activity is a potentially important mechanism. Furthermore, while HDAC4 expression is controlled by microRNA90, transcription of HDAC9 mRNA is regulated by a negative-feedback loop in which HDAC9 represses its own promoter91. Therefore, class IIa members are signal-responsive transcriptional corepressors subject to regulation by multiple mechanisms. Although the nuclear function of class IIa members is well established, their role in the cytoplasm (if any) is unclear. By comparison, the class IIb member HDAC6 is functionally important in this compartment.
HDAC6 as a key cytoplasmic regulator
Unlike many other HDACs, HDAC6 is predominantly (if not exclusively) cytoplasmic31. This finding has led to the simple hypothesis that HDAC6 possesses functions that are unrelated to histone modification, and available results convincingly support this theory. Indeed, multiple cytoplasmic processes seem to be regulated by HDAC6 (Fig. 3).
Figure 3. HDAC6 is a major deacetylase in the cytoplasm.

Through its tandem deacetylase (DAC) domains, HDAC6 deacetylates α-tubulin, cortactin and HSP90 to regulate cell motility, cilium assembly, maturation of the glucocorticoid receptor (GR), and activation of some protein kinases. HDAC6 may also deacetylate the type I interferon receptor (IFNαR)133. Additional cytoplasmic substrates are likely to be identified. Through its ubiquitin-binding zinc finger (ZnF), HDAC6 binds to ubiquitin and regulates aggresome formation, autophagy, activation of heat-shock factor 1 (HSF1), and epidermal growth factor receptor (EGFR)-induced macropinocytosis. It remains to be established whether HDAC6 has a direct role in the nucleus.
HDAC6 regulates cytoskeletal dynamics
HDAC6 has been shown to function as an α-tubulin deacetylase92–94, and it associates with microtubules and colocalizes with a microtubule motor complex92. Exogenous expression of HDAC6 leads to global deacetylation of α-tubulin and RNA interference-mediated knockdown of HDAC6 increases α-tubulin acetylation. Ectopic expression of HDAC6 also promotes chemotaxis92, which indicates that HDAC6-mediated deacetylation probably regulates microtubule-dependent cell mobility. In addition, HDAC6 might regulate cell adhesion95, the formation of the immune synapse96, and motor-mediated cargo transport along microtubules97,98. HDAC6 also has an unexpected role in the reabsorption of primary cilia of mammalian cells99. Like most HDACs, HDAC6 is a phosphoprotein100. Phosphorylation by Aurora A kinase activates the tubulin deacetylase activity of HDAC6 and promotes ciliary disassembly99. HDAC6 thus has roles in various microtubule-dependent cytoplasmic processes.
The filamentous (F)-actin-binding protein cortactin also interacts with HDAC6 and functions as its substrate67. As shown for α-tubulin, exogenous expression of HDAC6 leads to cortactin hypoacetylation, whereas inhibition of the deacetylase activity results in hyperacetylation. Independently, cortactin was identified as an acetylated protein in a proteomic study (M. Yoshida, personal communication). Interestingly, HDAC6 alters the ability of cortactin to bind F-actin by modulating the charge in the repeat region of cortactin and, in this way, modifies cell movement67. Therefore, in addition to its role in controlling microtubule-dependent cell motility, HDAC6 also seems to influence actin-dependent cell motility.
HDAC6 deacetylates HSP90
The heat shock chaperone protein HSP90 also associates with HDAC6 and is a bona fide cytoplasmic substrate101,102. HDAC6 is required for HSP90-dependent activation of the glucocorticoid receptor (GR). Treatment of cells with an HDAC6-sensitive deacetylase inhibitor or small interfering RNA-mediated knockdown of HDAC6 causes HSP90 to dissociate from p23 (a co-chaperone) and GR101. Studies in a cell-free system showed that HDAC6 promotes the assembly of stable HSP90–GR heterocomplexes but does not affect the intrinsic properties of GR103. Moreover, depletion of HDAC6 induces HSP90 acetylation and reduces its association with the leukemic tyrosine kinase Bcr-Abl102. HSP90 acetylation also regulates kinase activity of CHK1 (ref. 104). As a large number of other proteins, including many protein kinases involved in oncogenesis, require HSP90 for their maturation and function, it will be exciting to examine whether HSP90 deacetylation is important for their regulation. Although the list of cytoplasmic substrates of HDAC6 will probably continue to grow, the issue whether this deacetylase has a nuclear role awaits further investigation31.
HDAC6 and ubiquitin-dependent pathways
HDAC6 from mouse testes associates with the mammalian homologue of yeast UFD3 (ubiquitin fusion degradation protein 3) and p97/VCP31,66, a member of the AAA+ family of ATPases. Strikingly, the C-terminal zinc finger of HDAC6 efficiently interacts with ubiquitin, which causes HDAC6 to dissociate from the protein complex31. These results indicate an unexpected role for HDAC6 in ubiquitin-dependent pathways.
The expression of misfolded proteins in cells often leads to the formation of aggresomes105, a process that is facilitated by the delivery of misfolded proteins to these inclusion bodies via dynein-dependent transport on microtubules105. HDAC6 is a component of the aggresome, and treatment of cells with a proteasome inhibitor results in the accumulation of HDAC6 in polyubiquitin-containing aggresomes106. Mechanistically, HDAC6 associates with dynein motors and functions as an adaptor to link these motors to ubiquitylated misfolded proteins for delivery to aggresomes and degradation by proteasomes106. p97/VCP disrupts HDAC6–ubiquitin complexes and counteracts the ability of HDAC6 to promote the accumulation of polyubiquitylated proteins107. While the deacetylase activity of HDAC6 was found to be required for aggresome formation106, two recent studies showed that tubulin acetylation or HDAC6 inhibition accelerates dynein- and kinesin-mediated transport97,98.
Further support for the proposal that HDAC6 is essential for efficient management of cytotoxic proteins is provided by two recent studies suggesting that HDAC6 increases clearance of various substrates via autophagy108,109, a lysosome-mediated degradation pathway. Impairment of the ubiquitin–proteasome system induces autophagy, and this induction rescues neurodegeneration associated with such impairment108. Exogenous expression of HDAC6 alone compensates for an impaired ubiquitin–proteasome system and rescues neurodegeneration by autophagy108. Exactly how HDAC6 accelerates the turnover of misfolded proteins by autophagy is not known. Whatever the mechanism, it is clear that HDAC6 has a key role in the cellular clearance of misfolded proteins by different pathways and might function as a potential therapeutic target for neurodegenerative diseases. By associating with HSP90 and binding to ubiquitin, HDAC6 functions as a sensor for ubiquitylated protein aggregates to regulate the activation of heat-shock factor 1 (HSF1) during stress responses110. Moreover, HDAC6 is an essential component of stress granules111. This deacetylase, therefore, has key roles in different levels of cellular responses.
HDAC6 might also regulate cytoplasmic processes, such as macropinocytosis112. The mechanism by which the deacetylase and ubiquitin-binding activities are coordinated remains to be addressed. Together, recent studies have established HDAC6 as a major cytoplasmic regulator (Fig. 3) with properties that are not shared by other deacetylase, which is consistent with its unique domain organization (Fig. 1 and Supplementary information S1 (figure)).
Class I/II/IV HDACs in metazoan development
Genetic analyses of some class I/II members in C. elegans, D. melanogaster and mice have not only revealed important roles for HDAC1, 2 and 3, and class IIa members, in different developmental processes, but have also raised the important issue how to correlate these developmental functions with results from biochemical and cell-based assays. The findings from the genetic analyses are summarized in Table 2 and Supplementary information S6 (table), but some important points are noteworthy. First, HDAC1, 2 and 3 are catalytic subunits of multiprotein proteins (Table 1), so in addition to the deacetylases themselves, genetic analyses of associated subunits are very informative about functions of different deacetylase complexes (Table 2). Second, global deletion experiments have revealed that the functions of mouse HDAC1 and HDAC2 are not identical during development, and that they function differently in cultured tumour cells, suggesting that these two deacetylases do not always remain together as a catalytic core. Third, deletion of Sds3 or Sin3A affects K9 methylation of histone H3 and the subsequent distribution of HP1 nuclear foci113,114, indicating that K9 deacetylation is a prerequisite for subsequent methylation. This notion is well established in S. pombe21,64, and provides further support for an intimate relationship between histone deacetylation and methylation. And finally, all four members of class IIa have been subject to gene targeting in mice, with resultant phenotypes linked to MEF2 (Table 2), thereby indicating that the MEF2–HDAC axis (Fig. 2b) operates in many tissues aside from muscle115.
Table 2.
Class I/II/IV HDACs and associated subunits in human disease and mouse development
| Class | Member | Associated subunit | Map position | Human disease | Effect of mouse gene deletion |
|---|---|---|---|---|---|
| I | HDAC1 | 1p34.1 | Embryonic lethal by E9.5 (refs. 141, 142) | ||
| Proliferation defects in ES cells141,143 | |||||
| I | HDAC2 | 6q21 | Colon cancer | Perinatal death from cardiac defects142 | |
| Cardiomyopathy from heart-specific deletion of both HDAC1 and HDAC2 (ref. 142) | |||||
| Sin3A | 15q24.2 | Embryonic lethal by E6.5; splenomegaly & glomerulopathy in heterozygotes114,144 | |||
| Sds3 | 12q24.23 | Embryonic lethal by E6.5 | |||
| Defects in heterochromatin formation & chromosome segregation113 | |||||
| Mi2β | 12p13 | Defects in T cell development & CD4 gene expression145 | |||
| MBD3 | 19p13.3 | Pre- or peri-natal death146 | |||
| Defects in ES cell pluripotency | |||||
| p66α | 19p13.11 | Embryonic lethal by E9.5 (ref. 147) | |||
| LSD1 | 1p36.12 | Embryonic lethal by E7.5 (ref. 148) | |||
| I | HDAC3 | 5q31.3 | Embryonic lethality with vascular defects (R. Mongomery and E. Olson, pers. commun.) | ||
| Liver hypertrophy (S. Hiebert, pers. commun.) | |||||
| N-CoR | 17p11.2 | Embryonic lethal by E15.5 & defects in neuronal and hematopoitic differentiation149 | |||
| I | HDAC8 | Xq13 | Craniofacial abnormalities (M. Haberland and E. Olson, pers. commun.) | ||
| IIa | HDAC4 | 2q37.2 | Osteodystrophy? | Small size, chondrocyte hypertrophy | |
| Exencephaly150 | |||||
| IIa | HDAC5 | 17q21 | Cardiac hypertrophy151 | ||
| IIa | HDAC7 | 12q13.1 | Cardiovascular defects152 | ||
| IIa | HDAC9 | 7p12.1 | Cardiac hypertrophy153 | ||
| IIb | HDAC6 | Xp11.22 | Neurodegeneration? | Viable with massive increase of α-tubulin acetylation (P. Matthias, pers. commun.) | |
| IIb | HDAC10 | 22q13.3 | ? | ||
| IV | HDAC11 | 3p25.2 | ? |
Note: For associated subunits, only those subject to gene inactivation analysis in mice are listed here.
Implications for disease and therapy
As discussed above, studies in cell-based and animal models shed important light on the potential involvement of the Rpd3/Hda1 family members in the pathophysiology of human diseases. Related to this, HDAC2 has been linked to tumourigenesis in colorectal cancers and increased expression of this deacetylase has been found in a majority of human colon cancer explants116. In addition, a frame shift mutation, which causes loss of HDAC2 and confers resistance to HDAC inhibition, has been detected in some human cancers117.
Members of the family have also been implicated in neuropathological conditions. Accumulating data indicate a key role for HDAC6 in neurodegeneration. It is required for aggresome formation and present in Lewy bodies associated with neurodegenerative disorders like Parkinson’s diseases and dementia106. Impairment of autophagy resulting from decreased HDAC6 activities might predispose individuals to neurodegeneration108. On the other hand, HDAC6 inhibition promotes tubulin acetylation and facilitates microtubule-based vesicle transport, thereby compensating transport deficit in Huntington’s disease98. Thus, functional roles of HDAC6 may vary in different pathological states. Overall, compared to the rapidly accumulating knowledge about the molecular and developmental roles of the Rpd3/Hda1family of deacetylases, much less is known about their direct and specific roles in human pathology. Additional studies are also required to extrapolate accurately results from animal studies to humans and to determine how different HDACs might be altered or mutated in different types or stages of diseases.
Classical HDAC inhibitors have the potential to be therapeutic and chemo-preventive agents for cancer19,118, neurodegenerative disorders20, heart disease69, muscular dystrophy119, spinal muscular atrophy120, and transplantation intolerance121. Attesting to this potential, one such inhibitor was recently approved for treating cutaneous T-cell lymphoma19. Most of the inhibitors currently available display limited selectivity towards Rpd3/Hda1 family members in humans, raising the important question about the identity of the deacetylase(s) actually targeted in vivo. It might not any of the class IIa members since knockout studies suggest that their activation may be beneficial (Table 2). HDAC6 plays an important role in the management of misfolded proteins, so its activation of HDAC6 may also be beneficial (Fig. 3). However, it positively regulates cell motility and the activation of some oncogenic kinases, and its inhibition accelerates microtubule-based vesicle transport, so this deacetylase could be one in vivo target. Other candidates include class I members. If HDAC1 or 2, then the question is which deacetylase complex is the authentic target in patients. Of relevance, RNA interference-mediated knockdown of HDAC1 in D. melanogaster and mammalian cells recapitulates some effects of trichostatin A on cell cycle progression122,123.
In terms of designing new HDAC inhibitors, are isoform-specific compounds preferable to pan-HDAC inhibitors? With regard to isoform-specific intervention, would it be more reasonable to target the signalling pathways that control HDAC functions? Each HDAC has multiple substrates (such as those for HDAC6; Fig. 3) or binding partners (such as MEF2 and 14-3-3 for class IIa HDACs; Fig. 2), so the interaction interface might be a suitable intervention point for designing selective inhibitors.
It is important to note that the real in vivo target may vary in various diseases and different pathological stages. As inactivation of some classical HDACs is detrimental under certain conditions, small-molecule activators may have therapeutic potential26. Because many of the deacetylase are evolutionarily conserved (Fig. 1 and Supplementary information S1 (figure)), different model organisms provide economical alternatives for screening HDAC modulators and testing novel strategies of therapy and prevention.
Conclusions and perspectives
Since identification of the first HDACs in 1996, additional members of the classical family have been found in various organisms (Fig. 1, Supplementary information S1 (figure) and Box 2). There are 11 members in humans, and with the exception of HDAC8, all class I members form multisubunit complexes (Table 1). These complexes show different patterns of evolutionary conservation and carry out key roles in deacetylating histones, interacting with other chromatin regulators and shaping epigenetic programmes. Genome-wide localization maps have been generated for classical HDACs and components of their multisubunit complexes in yeast18,43,64 and should function as valuable guides for further identification and characterization of similar complexes in multicellular organisms.
Class I deacetylases also target non-histone substrates (Box 1), but further characterization is needed to address how the deacetylase complexes are involved. Members of class IIa function as signal transducers to repress transcription in a phosphorylation-dependent manner (Fig. 2). They are at a central interface between signalling and gene regulatory networks. The class IIb member HDAC6 is a major deacetylase in the cytoplasm, using its ubiquitin-binding domain as a stress sensor (Fig. 3). HDAC8 and HDAC10 appear to be specific to deuterostomes and vertebrates, respectively, whereas HDAC11 is conserved even in invertebrates and plants. Compared to other mammalian HDACs, much less is known about functions of these three deacetylases.
Classical HDACs and some of their associated subunits have been subjected to genetic analyses in mice (Table 2), as well as in C. elegans, D. melanogaster and zebrafish (supplementary information S6 (table)). One challenge is to correlate the resulting phenotypes with results from biochemical, molecular and cellular studies. A coherent and integrated picture from a variety of studies will not only provide a valuable guide for identifying and characterizing related human diseases, but will also form an important basis for rational drug design. Global or tissue-specific gene knockout in mice tends to provide information related to early development, whereas human diseases often develop in the late stages of life. Therefore, time-specific gene inactivation or knockdown is expected to greatly extend our understanding of human pathology and to efficiently mimic effects of HDAC inhibition to identify the best in vivo targets for developing new generations of HDAC inhibitors.
Recent proteomic studies have identified a diverse array of Nε-acetylated proteins124–126, but the responsible enzymes are largely unknown. There are also acetylation-like modifications such as formylation127 and propionylation128,129. Moreover, protein O-acetylation has just been reported130, and classical HDACs display intrinsic esterase activity30. These exciting discoveries make us wonder whether we are still at the dawn of a new era for protein acetylation research. How the Rpd3/Hda1 family is involved in deacetylating newly-identified Nε-acetylated proteins, controlling O-acetylation and lysine acetylation-like modifications, and interacting with other post-translational modifications, such as methylation, phosphorylation, ubiquitylation and sumoylation, is an exciting question awaiting further investigation. Answers to this question will provide rich and novel insights into the fundamental issue how members of the Rpd3/Hda1 family are involved in shaping ‘histone code-like’ regulatory programmes that have evolved from bacteria and yeasts to mice and men.
Supplementary Material
Acknowledgments
Owing to strict space constraints and the wealth of literature, we have used seminal reviews and supplemental figures/tables to cover some of the primary findings from this field. Research in our laboratories has been supported by grants from the NIH, AHA, and the Kaul Foundation (to E.S.) and from the Canadian funding agencies NCIC, CIHR, NSERC and CFI (to X.J.Y.).
Glossary terms
- 14-3-3 proteins
Members of a family of acidic proteins conserved from yeast and plants to humans. They are highly abundant, ~30 kDa in size and often recognize target proteins with the sequence motif RXXpS/TXP or RXXXpS/TXP, where X is any residue and pS/T denotes phosphoserine or phosphothreonine
- AAA+ ATPASES
ATPases that are associated wit h a variety of cellular activities. A superfamily of proteins with one or two nucleotide-binding domains (‘AAA modules’), which often form ring-like oligomers and function as chaperones in diverse cellular processes
- ASTROCYTE
A star-shaped glial cell that supports the tissue of the central nervous system
- AUTOPHAGY
A pathway for the recycling of cellular contents, in which materials inside the cell are packaged into vesicles and are then targeted to the vacuole or lysosome for bulk turnover
- BASAL BODY
The structure found at the base of eukaryotic cilia and flagella that consists of an array of nine microtubule triplets, as well as other proteins, and which is involved in the organization and assembly of the ciliary axoneme
- BROMODOMAIN
A conserved protein module first described for the Drosophila melanogaster homeotic gene regulator Brahma (means ‘creator’ in Hindu). This domain is present in many gene and/or chromatin regulators and has the ability to recognize acetyllysine motifs
- CHAPERONE
Protein that mediates assembly of another polypeptide-containing structure, but does not form part of the completed structure, or participate in its biological function
- CHEMOTAXIS
A type of migration that is stimulated by a gradient of a chemical stimulant or chemoattractant
- CHONDROCYTE
A differentiated cell of cartilage tissue
- CHROMODOMAIN
Originally identified as a 37-residue chromobox shared by the heterochromatin protein HP1 and the polycomb protein Pc2. Subsequently found in many other chromatin regulators and recognizes methyllysine protein motifs
- DEUTEROSTOME
An animal in which the anus develops from the first opening of the embryo, and the mouth is formed later. These include echinoderms and chordates
- DYNEIN
Microtubule-based molecular motor that moves towards the minus end of microtubules
- EUCHROMATIC
DNA that contains most of the structural genes, changes structure during the cell cycle and undergoes transcriptional regulation
- F-ACTIN
(Filamentous actin). A flexible, helical polymer of G-actin (globular actin) monomers that is 5–9 nm in diameter
- HOMOLOGUES
Genes (or their products) that are descended from a common ancestral gene
- IMMUNE SYNAPSE
A junction that forms at the contact region between a T cell and its target cells. T-cell activation occurs here
- INCLUSION BODIES
Insoluble aggregates of misfolded proteins; inclusion bodies are common in prokaryotes and the brains of patients affected by triplet-repeat diseases
- KINESIN
Microtubule-based molecular motor, most often directed towards the plus end of microtubules
- MACROPINOCYTOSIS
refers to a series of events initiated by extensive plasma membrane reorganization or ruffling to form an external macropinocytic structure that is then enclosed and internalized
- MATING TYPE GENES
Genes in the yeast chromosome that control the sexual fate of the yeast cell
- microRNA
A small RNA of ~21 nucleotides for regulating expression of mRNAs to which it is complementary in sequence
- MICROTUBULES
Hollow tubes, 25 nm in diameter, formed by the lateral association of 13 protofilaments, which are themselves polymers of α- and β-tubulin subunits
- MYOCYTE ENHANCER FACTOR 2 (MEF2)
An evolutionarily conserved transcription factor, the specific DNA-binding activity of which is mediated by an N-terminal MADS box and an adjacent MEF2-specific domain. Although initially identified as a muscle-specific transcription factor, different MEF2 isoforms are important in various tissues
- NUCLEOSOME
The basic structural subunits of chromatin, which consist of ~200 base pairs of DNA wrapped around an octamer of histones
- ORTHOLOGUE
product of a gene in two or more species that has evolved from a common ancestor
- PARALOGUES
Sequences, or genes, that have originated from a common ancestral sequence, or gene, by a duplication event
- PHD FINGER
A plant homeodomain-linked (PHD) zinc finger that chelates double zinc ions. This type of zinc finger is present in many chromatin regulators and was recently shown to bind the N-terminal tails of core histones in a methylation-dependent or -independent manner
- SANT DOMAIN
A 60-residue module initially identified in Swi3, Ada2, N-CoR, and TFIIIB as a putative DNA-binding domain. Present in various transcriptional and chromatin regulators and now considered as a histone-binding module
- SCAFFOLD PROTEIN
A protein that serves as a platform for the assembly of other proteins
- SE14-REPEAT
Ser-Glu-containing tetradecapeptide repeats found in HDAC6 proteins from high mammals
- ZnF-UBP
A ubiquitin-binding zinc finger present in HDAC6 and several ubiquitin-specific proteases
Footnotes
Weblinks
Rpd3: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=417699
Hda1: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=6324307
HDAC1: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=13128860
HDAC2: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=68068066
HDAC3: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=13128862
HDAC4: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=153085395
HDAC5: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=62750347
HDAC6: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=13128864
HDAC7: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=13259522
HDAC8: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=8132878
HDAC9: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=17158039
HDAC10: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=20070354
HDAC11: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=13376228
References
- 1.Gershey EL, Vidali G, Allfrey VG. Chemical studies of histone acetylation. The occurrence of epsilon-N-acetyllysine in the f2a1 histone. J Biol Chem. 1968;243:5018–5022. [PubMed] [Google Scholar]
- 2.Inoue A, Fujimoto D. Enzymatic deacetylation of histone. Biochem Biophys Res Commun. 1969;36:146–150. doi: 10.1016/0006-291x(69)90661-5. [DOI] [PubMed] [Google Scholar]
- 3.Candido EP, Reeves R, Davie JR. Sodium butyrate inhibits histone deacetylation in cultured cells. Cell. 1978;14:105–113. doi: 10.1016/0092-8674(78)90305-7. [DOI] [PubMed] [Google Scholar]
- 4.Sealy L, Chalkley R. The effect of sodium butyrate on histone modification. Cell. 1978;14:115–121. doi: 10.1016/0092-8674(78)90306-9. [DOI] [PubMed] [Google Scholar]
- 5.Vidali G, Boffa LC, Bradbury EM, Allfrey VG. Butyrate suppression of histone deacetylation leads to accumulation of multiacetylated forms of histones H3 and H4 and increased DNase I sensitivity of the associated DNA sequences. Proc Natl Acad Sci USA. 1978;75:2239–2243. doi: 10.1073/pnas.75.5.2239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Yoshida M, Kijima M, Akita M, Beppu T. Potent and specific inhibition of mammalian histone deacetylase both in vivo and in vitro by trichostatin A. J Biol Chem. 1990;265:17174–17179. Identified the first specific HDAC inhibitor and linked HDAC inhibition to cell growth arrest. [PubMed] [Google Scholar]
- 7.Taunton J, Hassig CA, Schreiber SL. A mammalian histone deacetylase related to the yeast transcriptional regulator Rpd3p. Science. 1996;272:408–411. doi: 10.1126/science.272.5260.408. Affinity-purification and identification of the first HDAC, supporting the link of gene silencing to histone deacetylation. [DOI] [PubMed] [Google Scholar]
- 8.Vidal M, Gaber RF. RPD3 encodes a second factor required to achieve maximum positive and negative transcriptional states in Saccharomyces cerevisiae. Mol Cell Biol. 1991;11:6317–6327. doi: 10.1128/mcb.11.12.6317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yang WM, Inouye C, Zeng Y, Bearss D, Seto E. Transcriptional repression by YY1 is mediated by interaction with a mammalian homolog of the yeast global regulator RPD3. Proc Natl Acad Sci USA. 1996;93:12845–12850. doi: 10.1073/pnas.93.23.12845. First report on the direct physical link of Rpd3 to a sequence-specific transcriptional repressor. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rundlett SE, et al. HDA1 and RPD3 are members of distinct yeast histone deacetylase complexes that regulate silencing and transcription. Proc Natl Acad Sci USA. 1996;93:14503–14508. doi: 10.1073/pnas.93.25.14503. Discovery of Hda1 and Rpd3 as deacetylase subunits of two multiprotein complexes, and identification of related proteins in bacteria. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Grozinger CM, Schreiber SL. Deacetylase enzymes: biological functions and the use of small-molecule inhibitors. Chem Biol. 2002;9:3–16. doi: 10.1016/s1074-5521(02)00092-3. [DOI] [PubMed] [Google Scholar]
- 12.Haigis MC, Guarente LP. Mammalian sirtuins--emerging roles in physiology, aging, and calorie restriction. Genes Dev. 2006;20:2913–2921. doi: 10.1101/gad.1467506. [DOI] [PubMed] [Google Scholar]
- 13.de Ruijter AJ, van Gennip AH, Caron HN, Kemp S, van Kuilenburg AB. Histone deacetylases (HDACs): characterization of the classical HDAC family. Biochem J. 2003;370:737–749. doi: 10.1042/BJ20021321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Khochbin S, Verdel A, Lemercier C, Seigneurin-Berny D. Functional significance of histone deacetylase diversity. Curr Opin Genet Dev. 2001;11:162–166. doi: 10.1016/s0959-437x(00)00174-x. [DOI] [PubMed] [Google Scholar]
- 15.Verdin E, Dequiedt F, Kasler HG. Class II histone deacetylases: versatile regulators. Trends Genet. 2003;19:286–293. doi: 10.1016/S0168-9525(03)00073-8. [DOI] [PubMed] [Google Scholar]
- 16.Gregoretti IV, Lee YM, Goodson HV. Molecular evolution of the histone deacetylase family: functional implications of phylogenetic analysis. J Mol Biol. 2004;338:17–31. doi: 10.1016/j.jmb.2004.02.006. An elegant phylogenetic analysis about different groups of histone deacetylases. [DOI] [PubMed] [Google Scholar]
- 17.Goldberg AD, Allis CD, Bernstein E. Epigenetics: a landscape takes shape. Cell. 2007;128:635–638. doi: 10.1016/j.cell.2007.02.006. [DOI] [PubMed] [Google Scholar]
- 18.Millar CB, Grunstein M. Genome-wide patterns of histone modifications in yeast. Nat Rev Mol Cell Biol. 2006;7:657–666. doi: 10.1038/nrm1986. [DOI] [PubMed] [Google Scholar]
- 19.Marks PA, Breslow R. Dimethyl sulfoxide to vorinostat: development of this histone deacetylase inhibitor as an anticancer drug. Nat Biotechnol. 2007;25:84–90. doi: 10.1038/nbt1272. [DOI] [PubMed] [Google Scholar]
- 20.Butler R, Bates GP. Histone deacetylase inhibitors as therapeutics for polyglutamine disorders. Nat Rev Neurosci. 2006;7:784–796. doi: 10.1038/nrn1989. [DOI] [PubMed] [Google Scholar]
- 21.Nicolas E, et al. Distinct roles of HDAC complexes in promoter silencing, antisense suppression and DNA damage protection. Nat Struct Mol Biol. 2007;14:372–380. doi: 10.1038/nsmb1239. [DOI] [PubMed] [Google Scholar]
- 22.Sengupta N, Seto E. Regulation of histone deacetylase activities. J Cell Biochem. 2004;93:57–67. doi: 10.1002/jcb.20179. [DOI] [PubMed] [Google Scholar]
- 23.Zhang X, et al. Histone deacetylase 3 (HDAC3) activity is regulated by interaction with protein serine/threonine phosphatase 4. Genes Dev. 2005;19:827–839. doi: 10.1101/gad.1286005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lee H, Rezai-Zadeh N, Seto E. Negative regulation of histone deacetylase 8 activity by cyclic AMP-dependent protein kinase A. Mol Cell Biol. 2004;24:765–773. doi: 10.1128/MCB.24.2.765-773.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Vannini A, et al. Crystal structure of a eukaryotic zinc-dependent histone deacetylase, human HDAC8, complexed with a hydroxamic acid inhibitor. Proc Natl Acad Sci USA. 2004;101:15064–15069. doi: 10.1073/pnas.0404603101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yang XJ, Grégoire S. Class II histone deacetylases: From sequence to function, regulation and clinical implication. Mol Cell Biol. 2005;25:2873–2884. doi: 10.1128/MCB.25.8.2873-2884.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Han A, He J, Wu Y, Liu JO, Chen L. Mechanism of recruitment of class II histone deacetylases by myocyte enhancer factor-2. J Mol Biol. 2005;345:91–102. doi: 10.1016/j.jmb.2004.10.033. [DOI] [PubMed] [Google Scholar]
- 28.Fischle W, et al. Enzymatic activity associated with class II HDACs is dependent on a multiprotein complex containing HDAC3 and SMRT/N-CoR. Mol Cell. 2002;9:45–57. doi: 10.1016/s1097-2765(01)00429-4. [DOI] [PubMed] [Google Scholar]
- 29.Lamb N, et al. Unraveling the hidden catalytic activity of vertebrate class IIa histone deacetylases. Proc Natl Acad Sci USA. 2007;104:17355–17340. doi: 10.1073/pnas.0706487104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Moreth K, et al. An active site tyrosine residue is essential for amidohydrolase but not for esterase activity of a class 2 histone deacetylase-like bacterial enzyme. Biochem J. 2007;401:659–665. doi: 10.1042/BJ20061239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Boyault C, Sadoul K, Pabion M, Khochbin S. HDAC6, at the crossroads between cytoskeleton and cell signaling by acetylation and ubiquitination. Oncogene. 2007;26:5468–5476. doi: 10.1038/sj.onc.1210614. [DOI] [PubMed] [Google Scholar]
- 32.Gao L, Cueto MA, Asselbergs F, Atadja P. Cloning and functional characterization of HDAC11, a novel member of the human histone deacetylase family. J Biol Chem. 2002;277:25748–25755. doi: 10.1074/jbc.M111871200. [DOI] [PubMed] [Google Scholar]
- 33.Somoza JR, et al. Structural snapshots of human HDAC8 provide insights into the class I histone deacetylases. Structure. 2004;12:1325–1334. doi: 10.1016/j.str.2004.04.012. [DOI] [PubMed] [Google Scholar]
- 34.Finnin MS, et al. Structures of a histone deacetylase homologue bound to the TSA and SAHA inhibitors. Nature. 1999;401:188–193. doi: 10.1038/43710. [DOI] [PubMed] [Google Scholar]
- 35.Nielsen TK, Hildmann C, Dickmanns A, Schwienhorst A, Ficner R. Crystal structure of a bacterial class 2 histone deacetylase homologue. J Mol Biol. 2005;354:107–120. doi: 10.1016/j.jmb.2005.09.065. [DOI] [PubMed] [Google Scholar]
- 36.Boyer LA, Latek RR, Peterson CL. The SANT domain: a unique histone-tail-binding module? Nat Rev Mol Cell Biol. 2004;5:158–163. doi: 10.1038/nrm1314. [DOI] [PubMed] [Google Scholar]
- 37.Kasten MM, Dorland S, Stillman DJ. A large protein complex containing the yeast Sin3p and Rpd3p transcriptional regulators. Mol Cell Biol. 1997;17:4852–4858. doi: 10.1128/mcb.17.8.4852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Keogh MC, et al. Cotranscriptional Set2 methylation of histone H3 lysine 36 recruits a repressive Rpd3 complex. Cell. 2005;123:593–605. doi: 10.1016/j.cell.2005.10.025. [DOI] [PubMed] [Google Scholar]
- 39.Carrozza MJ, et al. Histone H3 methylation by Set2 directs deacetylation of coding regions by Rpd3S to suppress spurious intragenic transcription. Cell. 2005;123:581–592. doi: 10.1016/j.cell.2005.10.023. [DOI] [PubMed] [Google Scholar]
- 40.Colina AR, Young D. Raf60, a novel component of the Rpd3 histone deacetylase complex required for Rpd3 activity in Saccharomyces cerevisiae. J Biol Chem. 2005;280:42552–42556. doi: 10.1074/jbc.M511561200. [DOI] [PubMed] [Google Scholar]
- 41.Kadosh D, Struhl K. Repression by Ume6 involves recruitment of a complex containing Sin3 corepressor and Rpd3 histone deacetylase to target promoters. Cell. 1997;89:365–371. doi: 10.1016/s0092-8674(00)80217-2. [DOI] [PubMed] [Google Scholar]
- 42.Joshi AA, Struhl K. Eaf3 chromodomain interaction with methylated H3-K36 links histone deacetylation to Pol II elongation. Mol Cell. 2005;20:971–978. doi: 10.1016/j.molcel.2005.11.021. [DOI] [PubMed] [Google Scholar]
- 43.Li B, et al. Combined action of PHD and chromodomains directs the Rpd3S HDAC to transcribed chromatin. Science. 2007;316:1050–1054. doi: 10.1126/science.1139004. [DOI] [PubMed] [Google Scholar]
- 44.Ahringer J. NuRD and SIN3 histone deacetylase complexes in development. Trends Genet. 2000;16:351–356. doi: 10.1016/s0168-9525(00)02066-7. [DOI] [PubMed] [Google Scholar]
- 45.Shi X, et al. ING2 PHD domain links histone H3 lysine 4 methylation to active gene repression. Nature. 2006;442:96–99. doi: 10.1038/nature04835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Le Guezennec X, Vermeulen M, Stunnenberg HG. Molecular characterization of Sin3 PAH-domain interactor specificity and identification of PAH partners. Nucleic Acids Res. 2006;34:3929–3937. doi: 10.1093/nar/gkl537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Denslow SA, Wade PA. The human Mi-2/NuRD complex and gene regulation. Oncogene. 2007;26:5433–5438. doi: 10.1038/sj.onc.1210611. [DOI] [PubMed] [Google Scholar]
- 48.Klose RJ, Bird AP. Genomic DNA methylation: the mark and its mediators. Trends Biochem Sci. 2006;31:89–97. doi: 10.1016/j.tibs.2005.12.008. [DOI] [PubMed] [Google Scholar]
- 49.Andres ME, et al. CoREST: a functional corepressor required for regulation of neural-specific gene expression. Proc Natl Acad Sci USA. 1999;96:9873–9878. doi: 10.1073/pnas.96.17.9873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.You A, Tong JK, Grozinger CM, Schreiber SL. CoREST is an integral component of the CoREST- human histone deacetylase complex. Proc Natl Acad Sci USA. 2001;98:1454–1458. doi: 10.1073/pnas.98.4.1454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Humphrey GW, et al. Stable histone deacetylase complexes distinguished by the presence of SANT domain proteins CoREST/kiaa0071 and Mta-L1. J Biol Chem. 2001;276:6812–6824. doi: 10.1074/jbc.M007372200. [DOI] [PubMed] [Google Scholar]
- 52.Hakimi MA, et al. A core-BRAF35 complex containing histone deacetylase mediates repression of neuronal-specific genes. Proc Natl Acad Sci USA. 2002;99:7420–7425. doi: 10.1073/pnas.112008599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Shi Y, et al. Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. Cell. 2004;119:941–953. doi: 10.1016/j.cell.2004.12.012. [DOI] [PubMed] [Google Scholar]
- 54.Lee MG, Wynder C, Cooch N, Shiekhattar R. An essential role for CoREST in nucleosomal histone 3 lysine 4 demethylation. Nature. 2005;437:432–435. doi: 10.1038/nature04021. [DOI] [PubMed] [Google Scholar]
- 55.Lee MG, et al. Functional interplay between histone demethylase and deacetylase enzymes. Mol Cell Biol. 2006;26:6395–6402. doi: 10.1128/MCB.00723-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wen YD, et al. The histone deacetylase-3 complex contains nuclear receptor corepressors. Proc Natl Acad Sci USA. 2000;97:7202–7207. doi: 10.1073/pnas.97.13.7202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Zhang J, Kalkum M, Chait BT, Roeder RG. The N-CoR-HDAC3 nuclear receptor corepressor complex inhibits the JNK pathway through the integral subunit GPS2. Mol Cell. 2002;9:611–623. doi: 10.1016/s1097-2765(02)00468-9. [DOI] [PubMed] [Google Scholar]
- 58.Karagianni P, Wong J. HDAC3: taking the SMRT-N-CoRrect road to repression. Oncogene. 2007;26:5439–5449. doi: 10.1038/sj.onc.1210612. [DOI] [PubMed] [Google Scholar]
- 59.Guenther MG, et al. A core SMRT corepressor complex containing HDAC3 and TBL1, a WD40-repeat protein linked to deafness. Genes Dev. 2000;14:1048–1057. [PMC free article] [PubMed] [Google Scholar]
- 60.Li J, et al. Both corepressor proteins SMRT and N-CoR exist in large protein complexes containing HDAC3. EMBO J. 2000;19:4342–4350. doi: 10.1093/emboj/19.16.4342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Underhill C, Qutob MS, Yee SP, Torchia J. A novel nuclear receptor corepressor complex, N-CoR, contains components of the mammalian SWI/SNF complex and the corepressor KAP-1. J Biol Chem. 2000;275:40463–40470. doi: 10.1074/jbc.M007864200. [DOI] [PubMed] [Google Scholar]
- 62.Guenther MG, Barak O, Lazar MA. The SMRT and N-CoR corepressors are activating cofactors for histone deacetylase 3. Mol Cell Biol. 2001;21:6091–6101. doi: 10.1128/MCB.21.18.6091-6101.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wu J, Carmen AA, Kobayashi R, Suka N, Grunstein M. HDA2 and HDA3 are related proteins that interact with and are essential for the activity of the yeast histone deacetylase HDA1. Proc Natl Acad Sci USA. 2001;98:4391–4396. doi: 10.1073/pnas.081560698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sugiyama T, et al. SHREC, an effector complex for heterochromatic transcriptional silencing. Cell. 2007;128:491–504. doi: 10.1016/j.cell.2006.12.035. [DOI] [PubMed] [Google Scholar]
- 65.Grozinger CM, Schreiber SL. Regulation of histone deacetylase 4 and 5 transcriptional activity by 14-3-3-dependent cellular localization. Proc Natl Acad Sci USA. 2000;97:7835–7840. doi: 10.1073/pnas.140199597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Seigneurin-Berny D, et al. Identification of components of the murine histone deacetylase 6 complex: link between acetylation and ubiquitination signaling pathways. Mol Cell Biol. 2001;21:8035–8044. doi: 10.1128/MCB.21.23.8035-8044.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Zhang X, et al. HDAC6 modulates cell motility by altering the acetylation level of cortactin. Mol Cell. 2007;27:197–213. doi: 10.1016/j.molcel.2007.05.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.McKinsey TA, Zhang CL, Lu J, Olson EN. Signal-dependent nuclear export of a histone deacetylase regulates muscle differentiation. Nature. 2000;408:106–111. doi: 10.1038/35040593. First report on the kinase-mediated nuclear export of class IIa HDACs. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.McKinsey TA, Olson EN. Cardiac histone acetylation--therapeutic opportunities abound. Trends Genet. 2004;20:206–213. doi: 10.1016/j.tig.2004.02.002. [DOI] [PubMed] [Google Scholar]
- 70.Martin M, Kettmann R, Dequiedt F. Class IIa histone deacetylases: regulating the regulators. Oncogene. 2007;26:5450–5467. doi: 10.1038/sj.onc.1210613. [DOI] [PubMed] [Google Scholar]
- 71.Zhang CL, McKinsey TA, Olson EN. The transcriptional corepressor MITR is a signal-responsive inhibitor of myogenesis. Proc Natl Acad Sci USA. 2001;98:7354–7359. doi: 10.1073/pnas.131198498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.van der Linden AM, Nolan KM, Sengupta P. KIN-29 SIK regulates chemoreceptor gene expression via an MEF2 transcription factor and a class II HDAC. EMBO J. 2007;26:358–370. doi: 10.1038/sj.emboj.7601479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Li X, Song S, Liu Y, Ko SH, Kao HY. Phosphorylation of the histone deacetylase 7 modulates its stability and association with 14-3-3 proteins. J Biol Chem. 2004;279:34201–34208. doi: 10.1074/jbc.M405179200. [DOI] [PubMed] [Google Scholar]
- 74.Potthoff MJ, et al. Histone deacetylase degradation and MEF2 activation promote the formation of slow-twitch myofibers. J Clin Invest. 2007;117:2459–2467. doi: 10.1172/JCI31960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Linseman DA, et al. Inactivation of the myocyte enhancer factor-2 repressor histone deacetylase-5 by endogenous Ca(2+)/calmodulin-dependent kinase II promotes depolarization-mediated cerebellar granule neuron survival. J Biol Chem. 2003;278:41472–41481. doi: 10.1074/jbc.M307245200. [DOI] [PubMed] [Google Scholar]
- 76.Vega RB, et al. Protein kinases C and D mediate agonist-dependent cardiac hypertrophy through nuclear export of histone deacetylase 5. Mol Cell Biol. 2004;24:8374–8385. doi: 10.1128/MCB.24.19.8374-8385.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Parra M, Kasler H, McKinsey TA, Olson EN, Verdin E. Protein kinase D1 phosphorylates HDAC7 and Induces its nuclear export after TCR activation. J Biol Chem. 2005;280:13762–13770. doi: 10.1074/jbc.M413396200. [DOI] [PubMed] [Google Scholar]
- 78.Dequiedt F, et al. Phosphorylation of histone deacetylase 7 by protein kinase D mediates T cell receptor-induced Nur77 expression and apoptosis. J Exp Med. 2005;201:793–804. doi: 10.1084/jem.20042034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Chang S, Bezprozvannaya S, Li S, Olson EN. An expression screen reveals modulators of class II histone deacetylase phosphorylation. Proc Natl Acad Sci USA. 2005;102:8120–8125. doi: 10.1073/pnas.0503275102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Berdeaux R, et al. SIK1 is a class II HDAC kinase that promotes survival of skeletal myocytes. Nat Med. 2007;13:597–603. doi: 10.1038/nm1573. [DOI] [PubMed] [Google Scholar]
- 81.Kim MA, et al. Identification of novel substrates for human checkpoint kinase Chk1 and Chk2 through genome-wide screening using a consensus Chk phosphorylation motif. Exp Mol Med. 2007;39:205–212. doi: 10.1038/emm.2007.23. [DOI] [PubMed] [Google Scholar]
- 82.Parra M, Mahmoudi T, Verdin E. Myosin phosphatase dephosphorylates HDAC7, controls its nucleocytoplasmic shuttling, and inhibits apoptosis in thymocytes. Genes Dev. 2007;21:638–643. doi: 10.1101/gad.1513107. Shows that a PP1 phosphatase complex is directly involved in controlling the nucleocytoplasmic trafficking of HDAC7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Sucharov CC, Langer S, Bristow M, Leinwand L. Shuttling of HDAC5 in H9C2 cells regulates YY1 function through CaMKIV/PKD and PP2A. Am J Physiol Cell Physiol. 2006;291:C1029–1037. doi: 10.1152/ajpcell.00059.2006. [DOI] [PubMed] [Google Scholar]
- 84.Illi B, et al. Nitric Oxide Modulates Chromatin Folding in Human Endothelial Cells via PP2A Activation and Class II HDACs Nuclear Shuttling. Circ Res. 2007 doi: 10.1161/CIRCRESAHA.107.157305. Epub. [DOI] [PubMed] [Google Scholar]
- 85.Kirsh O, et al. The SUMO E3 ligase RanBP2 promotes modification of the HDAC4 deacetylase. EMBO J. 2002;21:2682–2691. doi: 10.1093/emboj/21.11.2682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Petrie K, et al. The histone deacetylase 9 gene encodes multiple protein isoforms. J Biol Chem. 2003;278:16059–16072. doi: 10.1074/jbc.M212935200. [DOI] [PubMed] [Google Scholar]
- 87.Tatham MH, et al. Polymeric chains of SUMO-2 and SUMO-3 are conjugated to protein substrates by SAE1/SAE2 and Ubc9. J Biol Chem. 2001;276:35368–35674. doi: 10.1074/jbc.M104214200. [DOI] [PubMed] [Google Scholar]
- 88.Paroni G, et al. Caspase-dependent regulation of histone deacetylase 4 nuclear-cytoplasmic shuttling promotes apoptosis. Mol Biol Cell. 2004;15:2804–2818. doi: 10.1091/mbc.E03-08-0624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Liu F, Dowling M, Yang XJ, Kao GD. Caspase-mediated specific cleavage of human histone deacetylase 4. J Biol Chem. 2004;279:34537–34546. doi: 10.1074/jbc.M402475200. [DOI] [PubMed] [Google Scholar]
- 90.Chen JF, et al. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat Genet. 2006;38:228–233. doi: 10.1038/ng1725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Haberland M, et al. Regulation of HDAC9 gene expression by MEF2 establishes a negative-feedback loop in the transcriptional circuitry of muscle differentiation. Mol Cell Biol. 2007;27:518–525. doi: 10.1128/MCB.01415-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Hubbert C, et al. HDAC6 is a microtubule-associated deacetylase. Nature. 2002;417:455–458. doi: 10.1038/417455a. First report on the unexpected α-tubulin deacetylase activity of HDAC6. [DOI] [PubMed] [Google Scholar]
- 93.Matsuyama A, et al. In vivo destabilization of dynamic microtubules by HDAC6-mediated deacetylation. EMBO J. 2002;21:6820–6831. doi: 10.1093/emboj/cdf682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Zhang Y, et al. HDAC-6 interacts with and deacetylates tubulin and microtubules in vivo. EMBO J. 2003;22:1168–1179. doi: 10.1093/emboj/cdg115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Tran AD, et al. HDAC6 deacetylation of tubulin modulates dynamics of cellular adhesions. J Cell Sci. 2007;120:1469–1479. doi: 10.1242/jcs.03431. [DOI] [PubMed] [Google Scholar]
- 96.Serrador JM, et al. HDAC6 deacetylase activity links the tubulin cytoskeleton with immune synapse organization. Immunity. 2004;20:417–428. doi: 10.1016/s1074-7613(04)00078-0. [DOI] [PubMed] [Google Scholar]
- 97.Reed NA, et al. Microtubule acetylation promotes kinesin-1 binding and transport. Curr Biol. 2006;16:2166–2172. doi: 10.1016/j.cub.2006.09.014. [DOI] [PubMed] [Google Scholar]
- 98.Dompierre JP, et al. Histone deacetylase 6 inhibition compensates for the transport deficit in Huntington’s disease by increasing tubulin acetylation. J Neurosci. 2007;27:3571–3583. doi: 10.1523/JNEUROSCI.0037-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Pugacheva EN, Jablonski SA, Hartman TR, Henske EP, Golemis EA. HEF1-dependent Aurora A activation induces disassembly of the primary cilium. Cell. 2007;129:1351–1363. doi: 10.1016/j.cell.2007.04.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Linding R, et al. Systematic discovery of in vivo phosphorylation networks. Cell. 2007;129:1415–1426. doi: 10.1016/j.cell.2007.05.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Kovacs JJ, et al. HDAC6 regulates Hsp90 acetylation and chaperone-dependent activation of glucocorticoid receptor. Mol Cell. 2005;18:601–607. doi: 10.1016/j.molcel.2005.04.021. [DOI] [PubMed] [Google Scholar]
- 102.Bali P, et al. Inhibition of histone deacetylase 6 acetylates and disrupts the chaperone function of heat shock protein 90: a novel basis for antileukemia activity of histone deacetylase inhibitors. J Biol Chem. 2005;280:26729–26734. doi: 10.1074/jbc.C500186200. [DOI] [PubMed] [Google Scholar]
- 103.Murphy PJ, Morishima Y, Kovacs JJ, Yao TP, Pratt WB. Regulation of the dynamics of hsp90 action on the glucocorticoid receptor by acetylation/deacetylation of the chaperone. J Biol Chem. 2005;280:33792–33799. doi: 10.1074/jbc.M506997200. [DOI] [PubMed] [Google Scholar]
- 104.Scroggins BT, et al. An acetylation site in the middle domain of Hsp90 regulates chaperone function. Mol Cell. 2007;25:151–159. doi: 10.1016/j.molcel.2006.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Kopito RR. Aggresomes, inclusion bodies and protein aggregation. Trends Cell Biol. 2000;10:524–530. doi: 10.1016/s0962-8924(00)01852-3. Refs. 103–105 demonstrate that HDAC6 binds to and deacetylates Hsp90, thereby regulating its chaperone activity towards client proteins. [DOI] [PubMed] [Google Scholar]
- 106.Kawaguchi Y, et al. The deacetylase HDAC6 regulates aggresome formation and cell viability in response to misfolded protein stress. Cell. 2003;115:727–738. doi: 10.1016/s0092-8674(03)00939-5. Demonstrates that HDAC6 plays a key role in aggresome formation, thereby linking this deacetylase to cellular management of misfolded proteins. [DOI] [PubMed] [Google Scholar]
- 107.Boyault C, et al. HDAC6-p97/VCP controlled polyubiquitin chain turnover. EMBO J. 2006;25:3357–3366. doi: 10.1038/sj.emboj.7601210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Pandey UB, et al. HDAC6 rescues neurodegeneration and provides an essential link between autophagy and the UPS. Nature. 2007;447:859–863. doi: 10.1038/nature05853. [DOI] [PubMed] [Google Scholar]
- 109.Iwata A, Riley BE, Johnston JA, Kopito RR. HDAC6 and microtubules are required for autophagic degradation of aggregated huntingtin. J Biol Chem. 2005;280:40282–40292. doi: 10.1074/jbc.M508786200. Refs 108–109 make the unexpected link of HDAC6 to autophagy. [DOI] [PubMed] [Google Scholar]
- 110.Boyault C, et al. HDAC6 controls major cell response pathways to cytotoxic accumulation of protein aggregates. Genes Dev. 2007;21:2172–2181. doi: 10.1101/gad.436407. An elegant study linking HDAC6 to the activation of the transcription factor HSF1 in response to cytotoxic protein aggreates. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Kwon S, Zhang Y, Matthias P. The deacetylase HDAC6 is a novel essential component of stress granules involved in the stress response. Genes Dev. 2007;21 doi: 10.1101/gad.461107. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Gao YS, et al. Histone deacetylase 6 regulates growth factor-induced actin remodeling and endocytosis. Mol Cell Biol. 2007 doi: 10.1128/MCB.00393-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.David G, Turner GM, Yao Y, Protopopov A, DePinho RA. mSin3-associated protein, mSds3, is essential for pericentric heterochromatin formation and chromosome segregation in mammalian cells. Genes Dev. 2003;17:2396–2405. doi: 10.1101/gad.1109403. Describes mouse knockout studies for a non-catalytic subunit of HDAC complexes. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Dannenberg JH, et al. mSin3A corepressor regulates diverse transcriptional networks governing normal and neoplastic growth and survival. Genes Dev. 2005;19:1581–1595. doi: 10.1101/gad.1286905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Arnold MA, et al. MEF2C transcription factor controls chondrocyte hypertrophy and bone development. Dev Cell. 2007;12:377–389. doi: 10.1016/j.devcel.2007.02.004. [DOI] [PubMed] [Google Scholar]
- 116.Zhu P, et al. Induction of HDAC2 expression upon loss of APC in colorectal tumorigenesis. Cancer Cell. 2004;5:455–463. doi: 10.1016/s1535-6108(04)00114-x. [DOI] [PubMed] [Google Scholar]
- 117.Ropero S, et al. A truncating mutation of HDAC2 in human cancers confers resistance to histone deacetylase inhibition. Nat Genet. 2006;38:566–569. doi: 10.1038/ng1773. [DOI] [PubMed] [Google Scholar]
- 118.Bolden JE, Peart MJ, Johnstone RW. Anticancer activities of histone deacetylase inhibitors. Nat Rev Drug Discov. 2006;5:769–784. doi: 10.1038/nrd2133. [DOI] [PubMed] [Google Scholar]
- 119.Minetti GC, et al. Functional and morphological recovery of dystrophic muscles in mice treated with deacetylase inhibitors. Nat Med. 2006;12:1147–1150. doi: 10.1038/nm1479. [DOI] [PubMed] [Google Scholar]
- 120.Avila AM, et al. Trichostatin A increases SMN expression and survival in a mouse model of spinal muscular atrophy. J Clin Invest. 2007;117:659–671. doi: 10.1172/JCI29562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Tao R, et al. Deacetylase inhibition promotes the generation and function of regulatory T cells. Nat Med. 2007;13:1299–1307. doi: 10.1038/nm1652. [DOI] [PubMed] [Google Scholar]
- 122.Foglietti C, et al. Dissecting the biological functions of Drosophila histone deacetylases by RNA interference and transcriptional profiling. J Biol Chem. 2006;281:17968–17976. doi: 10.1074/jbc.M511945200. [DOI] [PubMed] [Google Scholar]
- 123.Senese S, et al. Role for histone deacetylase 1 in human tumor cell proliferation. Mol Cell Biol. 2007;27:4784–4795. doi: 10.1128/MCB.00494-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Iwabata H, Yoshida M, Komatsu Y. Proteomic analysis of organ-specific post-translational lysine-acetylation and -methylation in mice by use of anti-acetyllysine and -methyllysine mouse monoclonal antibodies. Proteomics. 2005;5:4653–4664. doi: 10.1002/pmic.200500042. [DOI] [PubMed] [Google Scholar]
- 125.Kim SC, et al. Substrate and functional diversity of lysine acetylation revealed by a proteomics survey. Mol Cell. 2006;23:607–618. doi: 10.1016/j.molcel.2006.06.026. [DOI] [PubMed] [Google Scholar]
- 126.Xie H, Bandhakavi S, Roe MR, Griffin TJ. Preparative peptide isoelectric focusing as a tool for improving the identification of lysine-acetylated peptides from complex mixtures. J Proteome Res. 2007;6:2019–2026. doi: 10.1021/pr060691j. [DOI] [PubMed] [Google Scholar]
- 127.Jiang T, Zhou X, Taghizadeh K, Dong M, Dedon PC. N-formylation of lysine in histone proteins as a secondary modification arising from oxidative DNA damage. Proc Natl Acad Sci USA. 2007;104:60–65. doi: 10.1073/pnas.0606775103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Chen Y, et al. Lysine propionylation and butyrylation are novel post-translational modifications in histones. Mol Cell Proteomics. 2007;6:812–819. doi: 10.1074/mcp.M700021-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Garrity J, Gardner JG, Hawse W, Wolberger C, Escalante-Semerena JC. N-lysine propionylation controls the activity of propionyl-CoA synthetase. J Biol Chem. 2007;282:30239–30245. doi: 10.1074/jbc.M704409200. [DOI] [PubMed] [Google Scholar]
- 130.Mukherjee S, Hao YH, Orth K. A newly discovered post-translational modification--the acetylation of serine and threonine residues. Trends Biochem Sci. 2007;32:210–216. doi: 10.1016/j.tibs.2007.03.007. [DOI] [PubMed] [Google Scholar]
- 131.Yang XJ, Grégoire S. Metabolism, cytosketon and cellular signaling in the grip of protein Nepsilon- and O-acetylation. EMBO Rep. 2007;8:556–561. doi: 10.1038/sj.embor.7400977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Mattagajasingh I, et al. SIRT1 promotes endothelium-dependent vascular relaxation by activating endothelial nitric oxide synthase. Proc Natl Acad Sci USA. 2007;104:14855–14860. doi: 10.1073/pnas.0704329104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Tang X, et al. Acetylation-dependent signal transduction for type I interferon receptor. Cell. 2007;31:93–105. doi: 10.1016/j.cell.2007.07.034. [DOI] [PubMed] [Google Scholar]
- 134.Lambard DB, et al. Mammalian Sir2 homolog SIRT3 regulates global mitochondrial lysine acetylation. Mol Cell Biol. 2007 doi: 10.1128/MCB.01636-07. Epub. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Seet BT, Dikic I, Zhou MM, Pawson T. Reading protein modifications with interaction domains. Nat Rev Mol Cell Biol. 2006;7:473–483. doi: 10.1038/nrm1960. [DOI] [PubMed] [Google Scholar]
- 136.Starai VJ, Celic I, Cole RN, Boeke JD, Escalante-Semerena JC. Sir2-dependent activation of acetyl-CoA synthetase by deacetylation of active lysine. Science. 2002;298:2390–2392. doi: 10.1126/science.1077650. [DOI] [PubMed] [Google Scholar]
- 137.Gardner JG, Grundy FJ, Henkin TM, Escalante-Semerena JC. Control of acetyl-Coenzyme A synthetase (AcsA) activity by acetylation/deacetylation without NAD+ involvement in Bacillus subtilis. J Bacteriol. 2006;188:5460–5468. doi: 10.1128/JB.00215-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Hallows WC, Lee S, Denu JM. Sirtuins deacetylate and activate mammalian acetyl-CoA synthetases. Proc Natl Acad Sci USA. 2006;103:10230–10235. doi: 10.1073/pnas.0604392103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Schwer B, Bunkenborg J, Verdin RO, Andersen JS, Verdin E. Reversible lysine acetylation controls the activity of the mitochondrial enzyme acetyl-CoA synthetase 2. Proc Natl Acad Sci USA. 2006;103:10224–10229. doi: 10.1073/pnas.0603968103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Yang XJ, Grégoire S. A recurrent phospho-sumoyl switch in transcriptional repression and beyond. Mol Cell. 2006;23:779–786. doi: 10.1016/j.molcel.2006.08.009. [DOI] [PubMed] [Google Scholar]
- 141.Lagger G, et al. Essential function of histone deacetylase 1 in proliferation control and CDK inhibitor repression. EMBO J. 2002;21:2672–2681. doi: 10.1093/emboj/21.11.2672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Montgomery RL, et al. Histone deacetylases 1 and 2 redundantly regulate cardiac morphogenesis, growth, and contractility. Genes Dev. 2007;21:1790–1802. doi: 10.1101/gad.1563807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Zimmermann S, et al. Reduced body size and decreased intestinal tumor rates in HDAC2-mutant mice. Cancer Res. 2007;67:9047–9054. doi: 10.1158/0008-5472.CAN-07-0312. [DOI] [PubMed] [Google Scholar]
- 144.Cowley SM, et al. The mSin3A chromatin-modifying complex is essential for embryogenesis and T-cell development. Mol Cell Biol. 2005;25:6990–7004. doi: 10.1128/MCB.25.16.6990-7004.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Williams CJ, et al. The chromatin remodeler Mi-2beta is required for CD4 expression and T cell development. Immunity. 2004;20:719–733. doi: 10.1016/j.immuni.2004.05.005. [DOI] [PubMed] [Google Scholar]
- 146.Kaji K, et al. The NuRD component Mbd3 is required for pluripotency of embryonic stem cells. Nat Cell Biol. 2006;8:285–292. doi: 10.1038/ncb1372. [DOI] [PubMed] [Google Scholar]
- 147.Marino S, Nusse R. Mutants in the mouse NuRD/Mi2 component P66alpha are embryonic lethal. PLoS ONE. 2007;2:e519. doi: 10.1371/journal.pone.0000519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Wang J, et al. Opposing LSD1 complexes function in developmental gene activation and repression programmes. Nature. 2007;446:882–887. doi: 10.1038/nature05671. [DOI] [PubMed] [Google Scholar]
- 149.Hermanson O, Jepsen K, Rosenfeld MG. N-CoR controls differentiation of neural stem cells into astrocytes. Nature. 2002;419:934–939. doi: 10.1038/nature01156. [DOI] [PubMed] [Google Scholar]
- 150.Vega RB, et al. Histone deacetylase 4 controls chondrocyte hypertrophy during skeletogenesis. Cell. 2004;119:555–566. doi: 10.1016/j.cell.2004.10.024. [DOI] [PubMed] [Google Scholar]
- 151.Chang S, et al. Histone deacetylases 5 and 9 govern responsiveness of the heart to a subset of stress signals and play redundant roles in heart development. Mol Cell Biol. 2004;24:8467–8476. doi: 10.1128/MCB.24.19.8467-8476.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Chang S, et al. Histone deacetylase 7 maintains vascular integrity by repressing matrix metalloproteinase 10. Cell. 2006;126:321–334. doi: 10.1016/j.cell.2006.05.040. Refs. 150 & 152 report the unexpected phenotypes of HDAC4 and HDAC7 null mice, thereby revealing the essential roles of these deacetylases in bone development and maintenance of vascular integrity, respectively. [DOI] [PubMed] [Google Scholar]
- 153.Zhang CL, et al. Class II histone deacetylases act as signal-responsive repressors of cardiac hypertrophy. Cell. 2002;110:479–488. doi: 10.1016/s0092-8674(02)00861-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.