

Summary
In Salmonella enterica, an activated Rcs signaling system inhibits initiation of transcription of the flhD master operon. Under these conditions, where motility is shut down, microarray experiments showed an increased RNA signal for three flagellar genes - fliPQR - located upstream of rcsA. We show here that it is the anti-sense (AS) strand of these genes that is transcribed, originating at a weak promoter in the intergenic region between fliR and rcsA. RcsA is an auxiliary regulator for the Rcs system, whose transcription is dependent on the response regulator RcsB. Rcs-activated rightward transcription, but not translation, of rcsA is required for stimulation of leftward AS transcription. Our results implicate a combined action of RcsB and rcsA transcription in activating the AS promoter, likely by modulating DNA superhelicity in the intergenic region. We show that the AS transcript regulates many genes in the Rcs regulon, including SPI-1 and SPI-2 virulence and stress-response genes. In the wild-type strain the AS transcript is present in low amounts, independent of Rcs signaling. Here, AS transcription modulates complementary sense RNA levels and impacts swarming motility. It appears that the flagellar AS transcript has been co-opted by the Rcs system to regulate virulence.
Introduction
The Rcs phosphorelay signaling system was first identified by its role in the positive regulation of capsular polysaccharide or colanic acid synthesis in Escherichia coli, but is now known to control a vast genetic program (Majdalani and Gottesman, 2005; Wang et al., 2007). This system is present exclusively in the family Enterobacteriaceae (Huang et al., 2006). Pathways regulated by the Rcs system include maintenance of cell wall integrity, cell division, stationary phase sigma factor activity, motility, and virulence. The specific signals that activate the Rcs system are not known, although the general physiological signals that trigger the system are accepted to be osmotic or other membrane stresses acting through the outer membrane lipoprotein RcsF or the inner membrane protein IgaA, upstream of the hybrid kinase RcsCD (Cano et al., 2002; Castanie-Cornet et al., 2006; Dominguez-Bernal et al., 2004; Laubacher and Ades, 2008; Majdalani et al., 2005). The response regulator RcsB associates with an unstable protein RcsA to bind to an RcsAB box in the promoter region of several genes; not all genes require RcsA for regulation, suggesting that RcsB can bind independently of RcsA (Wang et al., 2007; Wehland and Bernhard, 2000). RcsB is required for rcsA transcription (Wehland and Bernhard, 2000). The Rcs system is expected to be most activated in a sessile state, where capsular polysaccharide helps cells survive drying (Ophir and Gutnick, 1994), and is apparently important for later stages of biofilm formation (Danese et al., 2000; Prigent-Combaret et al., 1999). A transition from a sessile to a motile state, which is proficient in the intestinal phase of host invasion, requires that the Rcs system to be deactivated, since motility and SPI-1 virulence are negatively regulated by this system. Once inside the host, however, the Rcs system likely interacts with other two-component signaling systems to selectively turn on the SPI-2 virulence pathway required for intracellular proliferation and for systemic infection (Wang et al., 2007).
Flagellar biogenesis in Salmonella enterica involves expression of more than 50 genes that are transcribed in operons of three temporal classes (Chilcott and Hughes, 2000). The class 1 genes are included in the master operon flhDC, whose proteins direct transcription of class 2 promoters which encode structural and assembly proteins required for the biosynthesis of the base of the flagellar structure. FliA (σ28) is made at this stage and regulates class 3 transcription which produces the functional flagellar filament. In a mutant Salmonella strain that is near-maximally activated for the Rcs system, motility is completely inhibited (Wang et al., 2007). Under these conditions, RcsB was observed to inhibit transcription of the master operon through an RcsB box downstream of the flhDC promoter; RcsA appeared to have no effect on flhDC transcription. Even though the flagellar regulon was turned off, microarray experiments showed a strong positive signal for three class 2 genes fliPQR, whose products are part of the Type III secretion apparatus that directs export of most extra-cytoplasmic flagellar proteins. These elevated RNA levels were dependent on RcsB, but not on RcsA (Wang et al., 2007).
We show in this study that the positive RNA signal observed for fliPQR in the Rcs-activated strain is due to anti-sense (AS) transcription of these genes, and that the AS transcript plays a regulatory role in both the Rcs and motility pathways, apparently by different mechanisms. We describe novel features involved in induction of the AS transcript, and propose a model to explain how RcsB might participate in this process. Throughout, ‘RcsB’ refers to its phosphorylated form, which functions as a transcriptional regulator.
Results
An activated Rcs system promotes anti-sense transcription of fliPQR
The flagellar fliPQR genes are located at the end of the fliL operon, upstream of rcsA (Fig. 1A). We had reported a large RNA signal for these genes in a strain carrying a point mutation in igaA (igaA*), where the Rcs system is highly activated (Wang et al., 2007). As determined by microarray analysis, RNA levels for fliR were elevated 10–50 fold, fliQ 3–15 fold and fliP 1–3 fold (log-stationary phase), the induction being dependent on RcsB. Since the methods employed in the Wang et al. study did not distinguish between sense (S) and anti-sense (AS) transcription, a 2-step RT-PCR reaction was carried out as described under Experimental Procedures.
Fig. 1. Anti-sense regulation of fliPQR by an activated Rcs signaling system.

A. Schematic showing the fliL operon and rcsA. Length of the regions corresponding to the individual genes within the operon are drawn approximately to scale with respect to each other. Arrows indicate transcription start. The square box in the intergenic region between fliR and rcsA is the RcsAB box, not drawn to scale.
B. RT-PCR results for RNA isolated from wild-type and QW119 (igaA*) strains. Primer pairs (listed in Suppl. Table 1) were designed to anneal to either the sense (S) or anti-sense (AS) strands of RNA from indicated genes. RNA for the wild-type strain was isolated at mid-log phase where flagellar gene expression is maximal, whereas that for QW119 was isolated at stationary phase where activity of the Rcs system is maximal. 40 ng of RNA were used for PCR reactions, with 26 cycles of amplification. M, size markers × 100 bp.
C. RT-PCR results for RNA isolated from QW698, which contains a constitutively active RscC kinase allele we refer to as rcsC*. Reaction conditions as in B.
D. RT-PCR results for RNA isolated after subjecting wild-type cells to osmotic stress by addition of 15% sucrose (Suc) as described under Experimental Procedures. The S transcript was monitored for wcaA. 200 ng of RNA were used for PCR reactions, with 29 cycles of amplification.
In a wild-type strain probed for fliQ and fliR along with the upstream gene fliM, an S transcript was readily detected for fliM (Fig. 1B, left panel). S-fliQ was present in lower amounts, and S-fliR was undetectable under the same RT-PCR conditions. A weak AS signal (marked with a dot) was present for both fliQ and fliR, but not for fliM.
In the igaA* strain QW119, where flagellar transcription is inhibited at the class 1 level, S transcripts were not detected for any of the three genes (Fig. 1B, right panel). However, induced levels of AS transcripts were seen for fliQ and fliR. As a control, we also monitored the downstream gene rcsA; only the S transcript was detected for this gene in QW119, showing that AS transcription in this strain is specific to the flagellar genes. These results show that the transcription data reported for these genes in the earlier study represents AS transcription (see Fig. 3 in Wang et al., 2007).
To ascertain that AS transcription observed in the igaA* mutant is through activation of the RcsC kinase, a mutant rcsC allele (rcsC*) reported to signal constitutively, was tested (Garcia-Calderon et al., 2005). AS-fliR induction was seen in this strain as well (Fig. 1C).
To determine whether the AS transcript is also induced under physiological conditions known to trigger the Rcs system, we tested the effect of two reported conditions in E. coli – osmotic shock (Sledjeski and Gottesman, 1996), and growth at low temperatures in the presence of glucose and zinc (Hagiwara et al., 2003). Only sucrose-induced osmotic shock resulted in induction of the Rcs system in S. enterica, as monitored by induction of transcription of wcaA, a colanic acid biosynthesis gene (Fig. 1C). AS-fliR transcription was induced under these conditions as well. Thus, production of the AS transcript is not an artifact of mutant backgrounds, and is seen under physiological conditions that activate the Rcs system as well.
In summary, an activated Rcs system promotes expression of an AS-fliQR transcript. In a wild-type strain, sense or mRNA levels of fliQR are much lower compared to those of the upstream gene fliM, with fliR being undetectable under similar PCR conditions.
The anti-sense transcript is present at low levels throughout the growth phase in a wild-type strain, independent of RcsB
Data in Fig. 1B showed trace amounts of an AS transcript for fliQ and fliR even in wild-type cells. To better detect this transcript and test if it arises from the low-level activity of RcsB reported in this strain (Wang et al., 2007), the amount of RNA used in RT-PCR reactions was increased 5-fold along with increased cycle numbers, and the S and AS transcripts for fliR were monitored throughout the growth cycle of wild-type and isogenic ΔrcsB strains (Fig. 2). Both S- and AS-fliR transcripts could be detected under these conditions. In the wild-type strain the AS transcript was observed throughout the growth cycle, in higher amounts than the S-fliR transcript (Fig. 2A). An essentially similar pattern of AS-fliR expression was seen in the ΔrcsB strain (Fig. 2B), suggesting that RcsB activity does not make a large contribution to the AS-fliR transcript levels during this phase. This result is consistent with earlier gene expression as well as motility data showing only a low activity of the Rcs system under these conditions (Wang et al., 2004; Wang et al., 2007). Thus, the AS-fliR transcript in the wild-type strain is largely RcsB-independent. This transcript was unaffected in a ΔrpoS strain, showing that the stationary phase sigma factor RpoS is not responsible for its transcription (data not shown). In contrast to the continued presence of the AS transcript, the S-fliR transcript began to decline after 4 hrs in both strains, consistent with the pattern of class 2 gene expression reported for the wild-type strain in microarray experiments (Wang et al., 2004). In these experiments, the non-class 2 RNA expression pattern reported for fliQ and fliR can now be explained as due to AS RNA.
Fig. 2. Presence of the anti-sense transcript through the time course of growth in a wild-type strain.

A. RT-PCR results for AS- and S-fliR transcripts monitored through 8 hrs of growth in LB in the wild-type strain. 200 ng of RNA were used for PCR reactions, with 29 cycles of amplification.
B. As in A except in an isogenic ΔrcsB mutant QW434.
The anti-sense transcript originates in the fliR-rcsA intergenic region
Northern blots were performed to detect the length of the AS transcript in the igaA* strain. The expected length of a fliPQR-sized transcript is ~1870 nt. When a 790 nt sense strand sequence of fliR was used as probe, five separate bands were detected, the most prominent of these being ~500 nt (Fig. 3A). The largest band of ~1500 nt was detected only upon a longer exposure of the blot. It is possible that the smaller bands are processed products of a larger precursor. A similar band pattern was seen in the wild-type, which required very long exposures for detection (data not shown).
Fig. 3. Length and origin of the anti-sense fliPQR transcript.

A. Northern blot of RNA prepared from QW119, hybridized to a 790 nt sense fliR RNA probe, prepared as described under Experimental Procedures (lane 2). Lane 1, size markers × 1000 nt. The segment of the blot around the 1500 nt marker was exposed for a longer time.
B. Primer extension results, where the primer a-fliR-PE1 (see C) was used for annealing to the anti-sense fliR transcript. T and A are DNA sequencing reactions using the same primer. Arrows indicate extension products.
C. DNA sequence of the intergenic region. Nucleotides are numbered starting after the TAA codon of fliR. Primers used in the experiments in B and C are indicated with half arrowheads below the sequence. fliR-F3 actually hybridizes to the 61–82 nt region upstream of fliR stop codon (not shown; see Suppl. Table 1). 3 dots, stop codon for fliR; box, RcsAB binding region identified in E. coli (Francez-Charlot et al., 2003); double arrows, transcription start sites identified in B; −10 and −35 regions of a putative antisense or AS-fliR promoter (see text) and of the rcsA promoter are underlined; single dot, rcsA transcription start site (Ebel and Trempy, 1999). The translation start site for rcsA would be at nucleotide 282 i.e.134 nucleotides downstream from the transcription start site (not shown).
D. RT-PCR results for RNA isolated from QW119 and probed with a fixed primer fliR-F3 paired with five different primers in the indicated fliR-rcsA intergenic region in C.
E. AS-fliR levels monitored by RT-PCR in strains carrying indicated mutations. * indicates igaA* mutant background. AS-fliR::T contains an insertion of the 426 bp rrnB terminator next to the S-fliR stop codon. PAS-fliR# is a promoter mutant where the −10 ATATAT has been changed to CGCGCG. PCR conditions in D and E as in Fig. 1B.
To map the start of the AS transcript, primer extension was performed using a complementary sense-strand primer (a-fliR-PE1; see Fig. 3C). Two bands positioned at 50 and 51 nucleotides from the end of fliR TAA were observed (Fig. 3B). This places the transcript start site to the right of the RcsAB box in the intergenic between fliR and rcsA (referred to henceforth as intergenic or RA region) (Fig. 3C). Consistent with these results, RT-PCR reactions using a common fliR primer (fliR-F3) and various primers in the RA region generated efficient products with RA-R2 and RA-R2a primers which include the RcsAB box, but not RA-R2b, RA-R2c or RA-R3 primers (Fig. 3D).
Possible candidates for a class 1 or σ70 promoter for AS-fliR could be −10 atatAT/−35 aaGtaA (marked as AS-fliR −10 and −35 in Fig. 3C; matches to the promoter consensus indicated by upper case letters), or −10 TAtccT/−35 aaaACA (this alternate −10 is 3 bp to the left of the marked AS-fliR −10 in Fig. 3C). A promoter search program (http://www-bimas.cit.nih.gov/molbio/proscan/) did not recognize either promoter, nor found any strong candidates for alternative promoters in the RA region. To ascertain the promoter location, we mutated all six base pairs in the marked −10 region (which would also affect the alternate −10), as well as inserted the strong rrnB T1T2 terminator just outside the S-fliR stop codon (see Table 1). These manipulations resulted in either loss or strong diminution of the AS-fliR transcript, confirming the general promoter location of the AS transcript (Fig. 3E). We designate this promoter PAS-fliR.
TABLE 1.
Strains and plasmids used
| Strain/plasmid | Genotypea | Source (Ref.) |
|---|---|---|
| Strains | ||
| 14028 | Wild type S. enterica | |
| QW119 | igaA* | Wang et al 2007 |
| QW422 | ΔfliR::kan, igaA* | This study |
| QW434 | ΔrcsB | Wang et al 2007 |
| QW441 | ΔrcsA, igaA* | Wang et al 2007 |
| QW442 | ΔrcsB, igaA* | Wang et al 2007 |
| QW471 | ΔfliQR, igaA* | This study |
| QW555 | ΔRA(49–88) | This study |
| QW556 | ΔRA(1–48), igaA* | This study |
| QW593 | anti-fliFLAG-R, igaA* | This study |
| QW594 | anti-fliR-FLAG, igaA* | This study |
| QW597 | pBAD33 | This study |
| QW610 | pfliPQRBAD | This study |
| QW605a | pLfliQRBAD | This study |
| QW663 | ΔtopA::kan | This study |
| QW667 | ΔPrcsA | This study |
| QW668 | ΔtopA | This study |
| QW669 | AS-fliR::T (TrrnB at bp 10 after fliR TAA) | This study |
| QW671 | PAS-fliR# (atatat @ -10 mutated to cgcgcg) | This study |
| QW672 | ΔPrcsA, igaA* | This study |
| QW673 | AS-fliR::T, igaA* | This study |
| QW674 | PAS-fliR#, igaA* | This study |
| QW675 | ΔtopA::kan, ΔPrcsA | This study |
| QW676 | ΔtopA::kan, ΔrcsB | This study |
| QW695 | ΔrpoS | This study |
| QW698 | SV4989 (rcsC P484L) | Garcia-Calderón et al 2005 |
| QW699 | AS-fliRSTOP2(186th n.t. C changed to A) | This study |
| QW700 | AS-fliRSTOP2, igaA* | This study |
| QW701 | AS-fliRSTOP1(13th n.t. A changed to T) | This study |
| QW702 | AS-fliRSTOP1, igaA* | This study |
| Plasmids | ||
| pBAD33 | vector | Guzman et al 1995 |
| pfliPQRBAD | fliPQR in pBAD33 | This study |
| pLfliQRBAD | fliQR in pBAD33 | This study |
refers to the point mutation described in the indicated study. Δ followed by :: refers to gene deletion concomitant with substitution of indicated antibiotic resistance cassette. Primers used for various manipulations are listed in Supp. Table 1, and extent of deletions created in Suppl. Table 2. RA refers to the intergenic region between fliR and rcsA, and deletion numbers refer to base pairs to the 3′ of fliR after the stop codon TAA. FLAG subscripts refer to epitope tags placed either at the 5′ or 3′ end of the indicated genes. L and BAD superscripts refer to fliL or PBAD promoters driving sense or anti-sense transcription of the indicated genes, respectively. T refers to the 426 bp rrnB T1T2, # to the 6 bp substitution at −10 of AS promoter. The stop codon numbering refers to the putative anti-FliR ORF that starts in Val; G of GTA is 17th nt after TAA of fliR. See Experimental Procedures for details.
In summary, the AS-fliPQR transcript observed in an Rcs-activated strain is transcribed from a sub-optimal promoter PAs-fliR located in the RA region. The transcript is long and appears to be processed. The weak nature of PAS-fliR is consistent with low AS transcript levels observed in the wild-type strain (Figs. 1 and 2).
Induction of anti-sense transcription is dependent on rcsA transcription but not RcsA
The RcsAB box in the RA region is located 100 bp upstream of PrcsA (Fig. 4A; see also Fig. 3C). This box is reported to bind both RcsB and RcsA in E. coli (Wehland and Bernhard, 2000). RcsB is required for rcsA transcription, while RcsA serves as an autoregulator. Wang et al., 2007 reported that deletion of rcsB or of the RcsAB box (ΔRA1–48) in the igaA* strain strongly diminished both rcsA and fliPQR (AS) transcripts. This is shown again for AS-fliR in Fig. 4B. Note that rcsA transcription is eliminated in the ΔrcsB strain, but is still detectable in the RcsAB box mutant (ΔRA 1–48). An internal deletion that removed most of rcsA, resulted in an increase in remnant rcsA RNA levels, but no significant difference in AS-fliR (Fig. 4B; the rcsA probe in these experiments is to an upstream untranslated region of the transcript). Thus, RcsA itself is not required for AS transcription. To test if transcription of rcsA is required, a 25 bp deletion was generated that removed the rcsA promoter PrcsA (see Fig. 4A). The deletion abolished thercsA transcript, with a concomitant loss of the AS-fliR transcript (Fig. 4B). The PrcsA deletion did not alter PAS-fliR, as judged by similar basal levels of AS-fliR in the ΔPrcsA mutation in the wild-type strain (Fig. 4C). Mutation of the -10 region (PAS-fliR#) abolished the AS transcript in the wild-type strain, as it did in the igaA* strain (see Fig. 3E), showing a similar origin of the transcript in both strains. While loss of rcsA transcription abolished the AS transcript, the reverse was not the case i.e. rcsA transcription (which isdependent on RcsB binding to the RcsAB box) was unaffected in the PAS-fliR# mutant (data not shown). In summary, induction of AS transcription requires the RcsAB box, RcsB, and rcsA transcription, but not RcsA.
Fig. 4. Dependence of induction of anti-sense transcription on rcsA transcription.
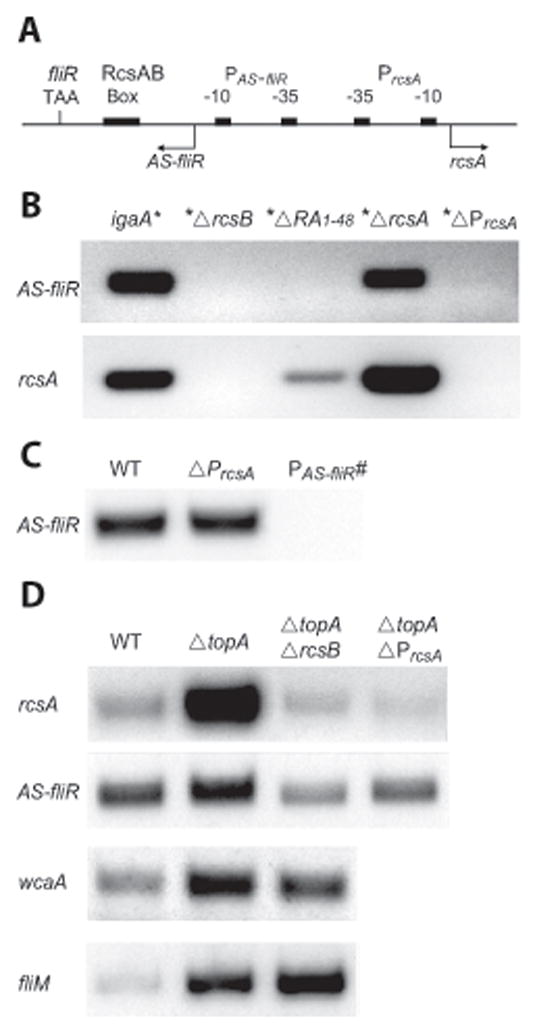
A. Schematic diagram showing regulatory elements in the fliR – rcsA intergenic region. These include the stop codon TAA of fliR, the RcsAB binding box, and the divergent promoters PAS-fliR and PrcsA. Other symbols as in Fig. 1A.
B. RT-PCR results for AS-fliR and rcsA RNA isolated from igaA* and its indicated derivative strains. An upstream untranslated region of rcsA (see Suppl. Table 1), which is still present in the ΔrcsA strain, was used for monitoring rcsA RNA levels in these strains. RNA was isolated at stationary phase. 40 ng of RNA were used for PCR reactions, with 26 cycles of amplification.
C, D. RT-PCR results for AS-fliR, rcsA, wcaA and fliM in wild-type, topA::kan (ΔtopA), and their indicated derivative strains. RNA was isolated from these strains at 6 hr. 200 ng of RNA were used for PCR reactions, with 29 cycles of amplification for all except fliM reactions, which used 100 ng RNA and 28 cycles.
How might transcription of rcsA promote anti-sense transcription? Several studies have demonstrated that the process of transcription can enhance negative DNA supercoiling (Liu and Wang, 1987; Wang and Lynch, 1993). An advancing polymerase generates positive supercoils in the DNA template ahead of it and negative supercoils behind it. Negative supercoiling generally stimulates transcription (Gralla, 1996), a well-known example being activation of an inactive leu-500 promoter in S. enterica and E. coli by negative supercoiling of DNA templates (Chen et al., 1992; Lilley and Higgins, 1991; Tan et al., 1994; Wu and Fang, 2003). To test if the activation of PAS-fliR is due to increased negative supercoiling generated in the wake of transcription from PrcsA, a topA null mutation was generated in the wild-type strain, and transcription from both PAS-fliR and PrcsA was monitored (Fig. 4D). The topA mutation is viable in Salmonella and produces increased negative DNA supercoiling (Richardson et al., 1984). Since changes in supercoiling can have positive effects on transcription generally, we also constructed a topA-rcsB double mutant, to distinguish topology-related changes from RcsB-related changes. As controls, we monitored the behavior of two other genes - wcaA and fliM – which are regulated positively and negatively, respectively, by RcsB. RNA was isolated at stationary phase, where the Rcs system is expected to be slightly active, although not as active as in the igaA* strain (see Figs. 1 and 2). This is evident in the wcaA control, where there is only a small difference between transcript levels in the topA versus topA rcsB strain. The increase in wcaA transcript levels in the topA strains compared to wild-type can therefore be attributed to a positive effect of increased negative supercoiling on this promoter. fliM transcription (from the fliL promoter; see Fig. 1A) is also stimulated in the topA strains, and the stimulation is enhanced in the topA rcsB strain, consistent with the negative regulation of the master flagellar operon by RcsB. Thus, both wcaA and fliM transcripts, respond positively to changes in DNA topology in the topA strain, independent of RcsB.
By contrast, rcsA transcript levels increase significantly in the topA mutant alone (Fig. 4D, top panel). Increased negative supercoiling itself is not sufficient for this stimulation, as seen by its absence in the topA rcsB strain. This suggests that supercoiling and RcsB have a synergistic effect on rcsA transcription. The stimulation is not seen in the topAΔPrcsA strain, confirming that the induced rcsA transcript originates from PrcsA in the topA strain.
Like rcsA, AS-fliR levels also increase in the topA mutant (Fig. 4D, second panel); the increase depends on RcsB and on transcription from PrcsA (compare topA with topA rcsB and topAΔPrcsA). However, while rcsA levels are induced ~5–6 fold in the topA strain, the increase in AS-fliR is modest (~1.5 fold). We note that the decrease in AS-fliR levels in the topA rcsB mutant compared to wild-type is consistent with the small contribution of RcsB to AS-fliR levels in wild-type cells (compare AS-fliR in Fig. 2A and 2B at 6 hr and 8 hr).
We draw several conclusions from these results. First, PrcsA is stimulated by increased negative DNA supercoiling, but only in the presence of RcsB. Second, increased negative DNA supercoiling itself is not sufficient to activate transcription from PAS-fliR, which is still dependent on transcription from PrcsA and on RcsB. Third, relative AS-fliR levels (compared to rcsA) are not as high in the topA strain as that seen in the igaA* strain. Thus, maximal stimulation of PAS-fliR requires higher activity of RcsB. A model rationalizing these results is presented under ‘Discussion’, where we propose that RcsB binding to the RcsAB box traps negative supercoils generated by transcription from PrcsA, stimulating both PrcsA and PAS-fliR.
A regulatory role for the anti-sense transcript in the Rcs pathway
To test if the AS transcript participates in regulation of genes in the Rcs regulon, a microarray analysis was performed, comparing RNA from the igaA* strain with that from three different igaA* derivatives carrying disruptions that eliminated the anti-sense transcript as determined by RT-PCR experiments. These were QW422 (ΔfliR::kan, igaA*), QW471 (ΔfliQR, igaA*) and QW556 (ΔRA1–48, igaA*) (see Fig. 3E and Fig. 4B). The microarray data showed a similar transcription profile for all three strains. Differentially regulated genes belonging to the Rcs regulon and common to all three pairwise combination of strains analyzed are shown in Table 2. Similar results were also seen for QW674 (PAS-fliR#, igaA*) (data not shown). None of the affected genes are reported to be regulated by RcsA (Wang et al., 2007). The largest functionally related group of genes to be negatively regulated by the AS transcript i.e. genes whose expression increased in the AS minus strains compared to the parent AS plus strain, were virulence genes belonging to SPI-I (hil, inv, prg, sic, sop) and SPI-2 (ssa, sse, ssr) (SPI is short for ‘Salmonella pathogenicity island’) (Galan, 1999; Waterman and Holden, 2003). Other regulated genes are listed in their alphabetical or numerical order. Among genes with a known function, cadB was strongly negatively regulated (>30 fold). The cadBA operon encodes a lysine decarboxylase (CadA) and a lysine-cadaverine antiporter (CadB); cadB is an essential component of the acid tolerance response in Salmonella (Park et al., 1996). Known genes that were strongly positively regulated (10–150 fold) include aceAB, manXYZ, and ompF. The ace operon encodes enzymes for the glyoxylate shunt pathway, required for growth of E. coli on acetate or fatty acids as the sole carbon source (Maloy and Nunn, 1982); aceA is required for persistence of Salmonella in mice (Fang et al., 2005). The manXYZ operon encodes a sugar transporter of the phosphoenol pyruvate-dependent phosphotransferase system (Plumbridge, 1998), and plays a role in organic solvent tolerance in E. coli (Okochi et al., 2007). ompF is a major outer membrane porin that is regulated in response to various stresses (Begic and Worobec, 2006; Pratt et al., 1996; Batchelor et al., 2005). ompF translation is known to be regulated by the micF anti-sense RNA (Guillier et al., 2006). A seven-member ego operon of unknown function was also strongly induced.
TABLE 2.
Regulation of the Rcs regulon by anti-sense fliPQR RNAa
| Name b | Ratio c | SD | Ratio | SD | |
|---|---|---|---|---|---|
| Virulence genes negatively regulated | Others positively regulated (cont’nd) | ||||
| avrA | 2.6 | 0.5 | melA | 3.0 | 0.9 |
| hilC | 122 | 95 | narV (narZ) | 3.1 | 0.7 |
| hilD | 2.3 | 0.3 | nirB (nirB) | 2.2 | 0.6 |
| invC,F,G,J,spaO,S (invF) | 1.4–2.8 | 0.1–0.7 | ompF | 24 | 11.8 |
| prgH,I,K,orgA,STM2868 (prgH) | 2.0–4.9 | 0.3–1.1 | osmE | 2.5 | 0.4 |
| sicA,P,sipA~D,iacP,sptP (sicA) | 1.7–2.3 | 0.1–1.0 | ptsG | 3.1 | 1.1 |
| sopA | 2.4 | 0.2 | rmf | 4.3 | 1.6 |
| sopB (sopB) | 4.8 | 1.4 | rpsP,rimM,trmD,rplS (rpsP) | 2.2–3.0 | 0.3–0.5 |
| ssaC (ssaB) | 1.6 | 0.1 | sodC | 2.0 | 0.7 |
| ssaJ,L (ssaG) | 2.5–4.3 | 0.2–0.9 | spvA | 4.5 | 1.7 |
| sseA,B,sscA (sseA) | 1.7–2.1 | 0.1 | spvB | 2.6 | 0.8 |
| sseF,G (sscB) | 1.9–2.1 | 0.3–0.4 | spy | 2.2 | 0.4 |
| sseJ | 9.8 | 2.1 | STM0950 (STM0951) | 2.7 | 0.9 |
| ssrA,B (ssrA) | 1.6–2.6 | 0.2–0.6 | STM1273 | 3.4 | 0.2 |
| Others negatively regulated | STM1561 | 2.4 | 0.5 | ||
| cadB | 36.5 | 7.0 | STM1633 (STM1633) | 2.3 | 0.4 |
| cdd | 8.4 | 1.5 | STM1638 | 7.3 | 2.0 |
| fabB | 2.5 | 0.1 | STM4071 | 5.3 | 0.8 |
| ppk (ppk) | 2.4 | 0.3 | STM4239,yjbJ (dinF) | 3.2–5.3 | 0.7–1.6 |
| sfbA (sfbA) | 2.3 | 0.5 | STM4242 | 3.6 | 0.6 |
| STM1328 | 6.6 | 0.4 | STM4510 | 5.6 | 1.5 |
| STM2287 | 4.9 | 0.4 | STM4519 | 5.4 | 2.4 |
| tdcE (tdcD) | 3.2 | 0.7 | yahoo | 5.6 | 1.7 |
| Others positively regulated | yaiA (aroL) | 2.8 | 1.0 | ||
| aceA~B (aceB) | 103–143 | 43–46 | yciE,STM1731 (yciE) | 3.5–3.9 | 1.6–1.9 |
| bax | 3.0 | 0.5 | yciF | 2.6 | 1.2 |
| chaB | 2.5 | 0.8 | ydeV | 5.6 | 0.8 |
| dniR | 3.6 | 2.2 | ydeW | 7.0 | 2.0 |
| ecnB | 2.5 | 0.6 | yeaG | 2.6 | 0.9 |
| ego, ydeY,Z, yneA~C, STM4080 (ego) | 2.5–20 | 0.1–4.6 | yebF | 2.2 | 0.5 |
| fbaB | 4.1 | 1.4 | yehZ (yehZ) | 2.1 | 0.7 |
| galP | 2.2 | 0.6 | ygiM | 2.3 | 0.8 |
| ggt | 5.6 | 5.0 | yhbO | 2.0 | 0.9 |
| katE | 2.0 | 0.6 | yibD | 2.8 | 0.9 |
| malE | 2.3 | 0.5 | yieF (yieE) | 2.5 | 0.8 |
| manX~Z (manX) | 7.3–13 | 1.5–4.9 | yncB | 2.7 | 1.1 |
The microarray data are for RNA harvested from cells at stationary phase.
Genes and open reading frames from the same operon are listed together. The first gene of the operon is indicated in parenthesis. ~ indicates intervening genes in the operon.
Transcription ratios are expressed such that the numbers are >1 for both positively and negatively regulated genes, because it is easier to compare fold regulation using whole numbers rather than fractions. Positive ratios were derived by dividing QW119 (igaA*) values by either QW422 ( fliR::kan, igaA*), QW471 ( fliQR, igaA*) or QW556 (ΔRA 1–48, igaA*); negative ratios were reverse of these. The differentially regulated Rcs regulon genes common to all three pairs of strains are shown. The range indicates low and high values for individual genes within the operon. The ratios represent an average of 4 independent repeats. SD, standard deviation.
The microarray data were validated by RT-PCR using hilC and aceA as representative genes that show negative and positive regulation, respectively (Fig. 5). The expression of these genes in the igaA* strain compared with its isogenic PAS-fliR# mutant derivative is consistent with repression of hilC and activation of aceA by AS-fliR.
Fig. 5. Differential effects of AS-RNA on regulation of genes in the Rcs pathway and motility.

A. Putative ORFs in the anti-sense strand of fliPQR. Open boxes show sense strand proteins, with their amino acid (aa) lengths indicated on top. Dark boxes indicate antisense or AS-ORFs starting with Met, and the light box the AS-ORF starting in Val. The aa lengths of these putative ORFs are shown below the boxes. c′ and n′ FLAG refer to FLAG epitopes engineered into the 210 aa AS-fliR ORF at the indicated locations. Numbers on the bottom refer to nucleotides, with 1 marking the start of the AS transcript as determined in Fig. 3B. Position of stop codons introduced into AS-fliR are marked.
B. RT-PCR validation of the microarray data in Table 2 for hilC and aceA using as control the AS-fliR defective mutant PAS-fliR#. The AS-fliR stop codon mutants show a similar loss of regulation of hilC and aceA even though their AS-fliR RNA levels are unaffected. 200 ng RNA/29 cycles of amplification were used for hilC and aceA, and 40 ng RNA/26 cycles for AS-fliR in RT-PCR reactions.
C. Swimming and swarming motility of wild-type and indicated promoter (ΔRA49–88 and PAS-fliR#), terminator (AS-fliR::T), and stop codon (AS-fliRstop1) mutant strains. Plates were incubated at 37°C for 4 hr.
D. S- and AS-fliR RNA in wild-type and indicated AS-RNA mutant derivative strains. RNA was isolated at 6 hr. RT-PCR conditions as in Fig. 2.
To test if the AS transcript regulates gene expression in the absence of an activated Rcs system, we cloned fliPQR on a plasmid, with a PBAD promoter positioned to the right of fliR driving the AS transcript (pfliPQRBAD). The expression profile of a wild-type strain carrying pfliPQRBAD (QW610) showed a consistent repression (1.5 – 5.5 fold) of virulence genes belonging to SPI-1 (hil, inv, prg, sic, sop) and SPI-4 (STM4257 operon). The rtsAB genes, which encode regulators of the SPI-1 genes (Ellermeier and Slauch, 2003), were also repressed (data not shown). These data were validated for hilC using RT-PCR (data not shown).
In summary, the AS transcript regulates a large number of genes in the Rcs regulon in both a positive and negative manner. The largest functional blocks of genes showing negative regulation belong to the SPI-1 and SPI-2 virulence pathways. Genes showing >10-fold regulation are involved in various stress responses. SPI-1 genes were regulated by the AS transcript even when expressed from a plasmid in the absence of Rcs signaling. Genes for colanic acid synthesis, which are the major target of the Rcs system (Majdalani and Gottesman, 2005), were not regulated by the AS transcript.
Gene regulation in the Rcs pathway is eliminated in AS-fliR stop codon mutants
A fairly long potential ORF (either 210 or 255 aa) is present in AS-fliR, while short ORFs are found in AS-fliQ and AS-fliP (40 and 75 aa, respectively) (Fig. 5A). The 255 aa AS-fliR ORF starts in Val, while the others all start in Met. A BLAST search revealed that in the closely related organisms E. coli and Shigella, there is a stop codon in the middle of their AS-fliR ORF. However, we found a 253 aa ORF in the AS-fliR strand from Burkholderia pseudomallei; this ORF has 37% amino acid identity to the S. enterica AS-fliR. To test if AS-fliR might be translated, we created FLAG epitopes at either the N- or C-terminal regions of the putative anti-FliR ORF. However, no protein bands were detected in Western blots probed with anti-epitope antibody, suggesting that the RNA is either not translated, or the translated product is unstable. To distinguish between these possibilities, we created two stop codons in AS-fliR ORF at two different locations (Fig. 5A) and monitored their effect on gene regulation in the igaA* strain (QW700 and QW702 in Table 1) by RT-PCR of the representative genes hilC and aceA genes. Like the PAS-fliR# mutant, the stop codon mutants also showed relief of repression of hilC and suppression of activation of aceA, even though unlike the PAS-fliR# mutant, AS-fliR expression was not abolished (Fig. 5B). Microarray data for these mutants were consistent with these results (data not shown). These data implicate a translated product of AS-fliR in regulating gene expression, although we were unable to detect such a product.
The anti-sense transcript plays a regulatory role in motility
To test if the AS transcript observed in wild-type cells (Fig. 2) plays a role in motility, we tested the effect of absence of this transcript on both swimming and swarming. Three strains not making the AS transcript – carrying the mutant promoter PAS-fliR#, the terminator insertion (AS-fliR::T), or a 40 bp deletion that selectively removed the PAS-fliR promoter as well as the start site of the AS transcript (ΔRA 49–88) - were examined. All three mutant strains showed a moderate inhibition of swimming (~10%), but a more severe effect on swarming motility (Fig. 5C). The more pronounced effect on swarming could due to the development dynamics of the swarmer colony, in which motility initiates only after a substantial lag. The growth rates of all these strains in LB broth were similar to wild-type (data not shown). The stop codon mutants, which do not affect AS-fliR transcription did not affect motility, in contrast to their effect on gene regulation in the igaA* strain (Fig. 5B). These results implicate a role for the AS-RNA, and not its translation product, in regulation of motility.
To test if the decrease in motility associated with the loss of the AS transcript might be explained by a decrease in levels of the S transcript, S- and AS-fliR transcripts were analyzed in the AS transcript mutants. All three mutants showed ~1.5–2.5 fold increase in S-fliR levels (Fig. 5D). This surprising result i.e. increased S-RNA but decreased motility, implicates the AS transcript in either decreasing the stability of the S transcript, or AS transcription in interfering with S transcription. The negative impact on motility of increased S-RNA levels may suggest that the specific stoichiometry of FliPQR proteins, which are part of a multi-component type III secretion apparatus, is critical for flagella biogenesis. Indeed, providing extra copies of fliQR from a plasmid (pLfliQR) had a negative effect on both swimming and swarming motility of the wild-type strain (data not shown).
In summary, AS transcription from PAS-fliR decreases complementary S transcript levels and plays a positive role in motility. This regulation is not via a translated product of AS-RNA.
Discussion
In this work we report the discovery of an anti-sense (AS) flagellar fliPQR transcript induced by the Rcs signaling system. We show that the transcript originates in the intergenic region between fliR and rcsA, at a weak promoter adjacent to PrcsA. Strong induction of leftward transcription from the PAS-fliR promoter is due to RcsB-controlled rightward transcription of rcsA. The Rcs system, which shuts down motility, appears to have exploited a translated product of the anti-sense transcript to regulate virulence and other stress-related genes. In a wild-type strain, AS transcription contributes to improving motility by a mechanism that controls levels of the complementary sense-strand RNA. A diagram summarizing our main findings is presented in Fig. 6, where we propose a model for RcsB function as discussed below.
Fig. 6.

Summary of regulatory interactions in the fliR-rcsA intergenic region, and model for how RcsB functions as a transcription activator. Steps 1–3 are described in text. Wavy lines, transcripts; solid arrows, positive regulation; dotted arrow, proposed intermediate;Θ, increased negative DNA supercoiling; ⊥, negative regulation.
Anti-sense regulation
Anti-sense RNAs were first discovered in controlling processes such as phage lysogeny, plasmid DNA replication, and DNA transposition in E. coli (Brantl, 2007). A common feature of these systems is that the anti-sense RNAs are cis-encoded, i.e they are transcribed from the same loci, but in an anti-sense direction from the target RNA. Their perfect complementarity to large segments of the target RNA leads to the formation of stable double-stranded RNA structures. These RNAs are generally small (~50 to 300 nt), are not translated, display highly structured stem-loops, and bind their target or sense RNAs to regulate gene expression in many different ways including transcription attenuation, translation inhibition, mRNA degradation and mRNA stabilization. Recent studies have identified many sRNAs or small regulatory RNAs in E. coli, many of which act in trans by base-pairing to activate or repress translation or to destabilize mRNAs (Storz et al., 2004). Base-pairing between these regulatory RNAs and their target mRNAs is not always perfect, and requires the Hfq protein which likely functions as an RNA chaperone to increase RNA unfolding or local target RNA concentration in order to stabilize the partial duplexes formed with their target RNA(s) (Brennan and Link, 2007). Non-coding RNAs are abundant in eukaryotes, varying in size from 200 bp to >10,000 bp (Wilusz et al., 2009). While the regulatory mechanisms involving many of the eukaryotic small RNAs are known (e.g. micro and siRNAs), those of the long RNAs are still enigmatic.
The AS transcript described in this study is long and is processed (Fig. 3). We surmise that the regulatory function of AS-fliPQR is encoded mainly in AS-fliQR, since AS-fliP is not present in high levels in the igaA* strain (Wang et al., 2007), and is not detectable in the wild-type strain (our unpublished data). We will therefore refer to this regulatory transcript as AS-fliQR. AS-fliR has a 255 aa ORF that spans nearly the entire fliR gene. Interruption of this ORF with two separate stop codon mutants abolished the AS RNA-dependent Rcs regulation in the igaA* strain (Table 2 and Fig. 5B), but did not affect motility in the wild-type strain (Fig. 5C), suggesting that the transcript appears to exert its regulatory role in distinctly different ways in the Rcs and molitity pathways.
In the Rcs pathway, a translation product of AS-fliR appears to regulate a diverse set of genes. However, FLAG epitopes engineered at either the N- or C-terminal regions of the anti-FliR ORF failed to detect any protein. This suggests that the product is either processed or unstable. We note that RcsA, the auxiliary regulator in the Rcs pathway, is also unstable (Majdalani and Gottesman, 2005). The anti-FliR regulator can apparently function independently of RcsB, since overexpression of the AS-RNA from a plasmid in the absence of an activated Rcs system, was seen to regulate a subset of virulence genes common to those regulated in the igaA* strain.
In the motility pathway, the sense or mRNA of the promoter-distal genes fliQR in the fliL operon shows lower abundance compared to mRNA of the promoter-proximal genes (see Fig. 1B). This could be due to the lack of an obvious Rho-independent terminator after fliR (Malakooti et al., 1994), increasing the susceptibility of the 3′ end of the fliL-R transcript to degradation. However, absence of the AS-fliQR transcript increases S-fliR levels (Fig. 5D), suggesting that 3′ end of the fliL-R transcript may be targeted for degradation by the AS-RNA. Failure of the stop codon mutants in AS-fliR ORF to affect motility (Fig. 5C), suggests that the observed AS-RNA regulation is not dependent on translation of the AS transcript. An RNA-dependent mechanism for degradation could include direct base-pairing between the S and AS transcripts. Alternatively, AS transcription could create a DNA structure in the intergenic region that releases the S transcript immediately from the DNA template, shortening its life. Or, S transcript levels might be controlled by interference of S transcription from RNA polymerase traveling in the AS direction. The decrease in motility associated with increase in S-fliR levels upon loss of the AS transcript (Fig. 5C,D), suggests that the stoichiometry of these proteins is critical to flagellar assembly. FliO, FliP, FliQ and FliR are integral membrane proteins located within the MS ring; they function in concert with two other membrane proteins FlhA and FlhB to form the Type III secretion apparatus for extracytoplasmic flagellar components (Chevance and Hughes, 2008). The numbers of these components and their mechanism of assembly are currently unknown.
Transcription, DNA topology and RcsB
Transcription and DNA topology are mutually linked (Wang and Lynch, 1993). Liu and Wang formulated the “twin-supercoiled-domain” model to explain the enhanced local negative DNA supercoiling associated with transcription (Liu and Wang, 1987). They hypothesized that as the size of the growing RNA transcript increases, it becomes progressively more difficult for the transcribing complex to rotate around the DNA double helix, reaching a point where it is energetically more feasible for DNA to rotate around its own helical axis to generate a transient positively supercoiled domain in front of the RNA polymerase and a transient negatively supercoiled domain behind it. DNA gyrase rapidly converts the positive supercoils into negative supercoils (Wang, 1996), which are thought to stabilize the formation of open transcription complexes by changing DNA twist (Gartenberg and Wang, 1992; Travers and Muskhelishvili, 2007). A striking stimulation of transcription-coupled DNA supercoiling has been demonstrated in vitro in the presence of site-specific DNA binding proteins such as the λ O protein, or the Gal or Lac repressors (Leng and McMacken, 2002). Such nucleoprotein complexes have been proposed to act as barriers by either wrapping or bending DNA in a manner that hinders diffusion of DNA supercoils, helping to localize transcription-generated supercoils in the vicinity of their recognition sequences.
The location of RcsB binding sites on DNA differs in promoters that do and do not require RcsA (Majdalani and Gottesman, 2005). RcsB sites are located immediately adjacent to the −35 region of promoters that do not require RcsA, and there is evidence for co-operative binding of RcsB and RNA polymerase at such promoters. In promoters that are modulated by RcsA, RcsB binding sites are located 50–100 bp upstream of the promoters, and the mechanism of activation from these distant sites is not known. From the results reported in this study, where (a) increased negative supercoiling and RcsBwere observed to act synergistically to activate transcription from PrcsA (Fig. 4D), (b) maximal activation of PAS-fliR was dependent on transcription from PrcsA, a highly activated RcsB, as well as the RcsAB box (compare data in Fig. 4B and 4D), and (c) low-level transcription of rcsA was observed even the absence of the RcsAB box (Fig. 4B; see also Fig. 3C in Wang et al., 2007), we propose the following model for transcription activation by RcsB in the fliR-rcsA intergenic region (Fig. 6): 1) RcsB contributes to a low-level stimulation of transcription from PrcsA, independent of the RcsAB box. 2) RcsB binding to the RcsAB box enhances transcription from PrcsA by slowing the diffusion of rcsA transcription-generated supercoils. 3) Strong binding of RcsB to the RcsAB box acts as a barrier against dissipation of the negative supercoils generated by rcsA transcription, producing a feed-forward loop that increases negative supercoiling even further, activating PAS-fliR. 4) The affinity of RcsA for the RcsAB box may be modulated in response to the altered DNA topology to autoregulate this process.
The curious placement of rcsA
Finally, we find it curious that rcsA is the lone gene in the Rcs signaling pathway that is positioned downstream of the fliL operon, while three other rcs genes are located together elsewhere on the genome. We speculate that the original role of AS transcription was to regulate fliQR mRNA levels from its cis locus. We suggest that this transcript was later co-opted by the Rcs system for trans regulatory functions. We note that genes for the biosynthesis of colanic acid, which are the major target of the Rcs system and thought to be required for survival outside the host, are not regulated by the AS transcript. Instead, AS-regulated genes include those required for host virulence or other stress responses. These data might also argue for a later appearance of the trans AS regulation in the evolutionary history of this organism.
Experimental Procedures
Bacterial strains, growth conditions and motility assays
Strains are listed in Table 1. Bacteria were grown either in L-broth (LB) base (20g/liter), or in M63 minimal medium for experiments involving either pBAD33 plasmid derivatives or for inducing osmotic shock (see below). Strains carrying pBAD33 constructs were first grown in M63-Glycerol overnight and subsequently diluted 1:100 in M63-Arabinose for induction. M63 was prepared according to the recipe from Molecular Cloning (Sambrook et al., 1989). Final concentration of carbon sources (glycerol, glucose or arabinose) in M63 was 0.2%. For preparation of RNA samples, cultures grown in LB were harvested at OD600 of 0.6 (~2.5 hr) for mid-log phase, and OD600 of ~2.3 (6 hr) for stationary phase; cultures grown in M63 minimal medium reached OD600 of 0.6 at ~4 hrs. Strains carrying the topA mutation grew more slowly than wild-type. The initial inoculum was therefore increased ~3-fold for topA strains, in order for both strains to reach a similar OD600 at 6hr. Swim plates were solidified with 0.3% agar, swarm plates with 0.6% Eiken agar (Wang et al., 2007).
To mimic physiological conditions that activate the Rcs system, two previously reported protocols were followed. In one, wild-type cells were grown in EB (1.8% Eiken broth, 0.4% glucose, 50 mM MOPS (pH 7.0), 1 mM ZnCl2) at 20°C (Hagiwara et al., 2003). Control cells were incubated in LB at 37°C. Cultures were harvested at OD600 of 0.6 for RNA extraction. In the second protocol, wild-type cells were incubated in M63-glucose at 30°C for 4 hrs, and sucrose was added to a final concentration of 15% (Sledjeski and Gottesman, 1996). Control cultures received an equal volume of M63 salts. After one hour, cells were harvested for RNA extraction. Eiken agar and broth were purchased from Eiken Chemical Co., Japan.
Deletion or insertion of genes and regulatory regions was achieved by the onestep mutagenesis procedure (Datsenko and Wanner, 2000) as described (Wang et al., 2005). The extent of coding or regulatory sequences deleted is provided in Suppl. Table 2. Point mutations and insertions were introduced on the chromosome by first amplifying DNA fragments from genomic template DNA using appropriate primers containing the mutations (see Suppl. Table 1), followed by replacement of a tetRA cassette previously inserted into the relevant region, by selecting recombinants for Tet sensitivity as described (Wang et al., 2007). Design of the anti-sense stop codon 2 mutation ensured that the sense strand amino acid code was unaltered. The 426 bp rrnB terminator fragment (Orosz et al., 1991) was amplified from pBAD33 (Guzman et al., 1995), and inserted 10 bp after the stop codon of fliR. FLAG epitopes at the N- and C terminus of the putative 210 aa anti-fliR ORF were constructed by a similar tetRA replacement method. All constructs were confirmed by DNA sequencing. Mutant combinations were constructed by P22 transduction where appropriate. FLAG antibodies were purchased from Sigma.
Plasmids
pfliPQRBAD contains DNA from ATG of fliP to TAA of fliR, cloned into pBAD33 such that the anti-sense strand was driven by the PBAD promoter, with no specific promoter driving the sense strand (Guzman et al., 1995). Primers for PCR amplification of this region included SphI and XbaI restriction sites, respectively, for ligating into similar sites on pBAD33. In pLfliQRBAD, the fliL promoter was introduced to drive sense strand expression of fliQR.
Northern blots and Primer extension
QW119 was grown in LB medium for 6 hrs. RNA was extracted using MasterPure™ RNA Purification Kit (Epicentre, MD). DNA contamination was removed by using DNA-free™ (Ambion, TX). The Northern blot preparation followed the instruction manual of NorthernMax®-Gly Kit (Ambion, TX). Briefly, 15 μg total RNA was loaded to a 1% agarose gel, which was run at 65 volts for 2 hrs and transferred to a positively charged nylon membrane. An RNA probe covering nearly the entire fliR coding sequence was synthesized with Strip-EZ®RNA T7 Kit (Ambion, TX) with addition of αP32-dUTP, using primers fliR-T7 and RA-R1 (see Suppl. Table 1) and fliR DNA template.
Primer extension for mapping the start of the transcript was performed using Primer Extension System from Promega according to the instruction manual, using the primer a-fliR-PE1. DNA sequence reactions were run alongside using the same primer, and the fmol® DNA cycle sequencing system from Promega.
Microarray experiments and RT-PCR
RNA preparation and microarray analyses were done as described (Wang et al., 2004). RT-PCR was performed as described (Wang et al., 2007) using a one-step RT-PCR kit (Qiagen). However, in order to distinguish the sense and anti-sense transcript, a strategy of “two-step RT-PCR” was used: the reverse transcription step was done first with one primer (the forward primer for detecting the anti-sense strand or the reverse primer for the sense strand) with cycling condition of 50°C for 35 min and 95°C for 15 min. After the initial reverse transcription, the other primer was added to the reaction tube to perform regular PCR amplification using the following steps: 94°C for 15 s, 55 °C for 25 s and 72 °C for 45 s. RNA amounts and cycling numbers varied depending on the RNA abundance of target genes. All samples were normalized using 30S ribosomal RNA. Primer pairs were designed to amplify ~150–350 bp of DNA. DNA bands were quantitated using Bio-Rad Gel-doc QuantityOne.
Supplementary Material
Suppl. Table 1. Primers used in indicated experiments.
Suppl. Table 2. Extent of deletion in genes or regulatory regions.
Acknowledgments
This work was supported in part by National Institutes of Health grant GM57400 and a University Research Grant. We thank Dr Ramos-Morales, Universidad de Sevilla, Spain, for the rcsC* strain, and Makkuni Jayaram and Shelley Payne for helpful comments on the manuscript.
References
- Batchelor E, Walthers D, Kenney LJ, Goulian M. The Escherichia coli CpxA-CpxR envelope stress response system regulates expression of the porins ompF and ompC. J Bacteriol. 2005;187:5723–5731. doi: 10.1128/JB.187.16.5723-5731.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Begic S, Worobec EA. Regulation of Serratia marcescens ompF and ompC porin genes in response to osmotic stress, salicylate, temperature and pH. Microbiology. 2006;152:485–491. doi: 10.1099/mic.0.28428-0. [DOI] [PubMed] [Google Scholar]
- Brantl S. Regulatory mechanisms employed by cis-encoded antisense RNAs. Curr Opin Microbiol. 2007;10:102–109. doi: 10.1016/j.mib.2007.03.012. [DOI] [PubMed] [Google Scholar]
- Brennan RG, Link TM. Hfq structure, function and ligand binding. Curr Opin Microbiol. 2007;10:125–133. doi: 10.1016/j.mib.2007.03.015. [DOI] [PubMed] [Google Scholar]
- Cano DA, Dominguez-Bernal G, Tierrez A, Garcia-Del Portillo F, Casadesus J. Regulation of capsule synthesis and cell motility in Salmonella enterica by the essential gene igaA. Genetics. 2002;162:1513–1523. doi: 10.1093/genetics/162.4.1513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castanie-Cornet MP, Cam K, Jacq A. RcsF is an outer membrane lipoprotein involved in the RcsCDB phosphorelay signaling pathway in Escherichia coli. J Bacteriol. 2006;188:4264–4270. doi: 10.1128/JB.00004-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen D, Bowater R, Dorman CJ, Lilley DM. Activity of a plasmid-borne leu-500 promoter depends on the transcription and translation of an adjacent gene. Proc Natl Acad Sci U S A. 1992;89:8784–8788. doi: 10.1073/pnas.89.18.8784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chevance FF, Hughes KT. Coordinating assembly of a bacterial macromolecular machine. Nat Rev Microbiol. 2008;6:455–465. doi: 10.1038/nrmicro1887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chilcott GS, Hughes KT. Coupling of flagellar gene expression to flagellar assembly in Salmonella enterica serovar Typhimurium and Escherichia coli. Microbiol Mol Biol Rev. 2000;64:694–708. doi: 10.1128/mmbr.64.4.694-708.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Danese PN, Pratt LA, Kolter R. Exopolysaccharide production is required for development of Escherichia coli K-12 biofilm architecture. J Bacteriol. 2000;182:3593–3596. doi: 10.1128/jb.182.12.3593-3596.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Datsenko KA, Wanner BL. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci U S A. 2000;97:6640–6645. doi: 10.1073/pnas.120163297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dominguez-Bernal G, Pucciarelli MG, Ramos-Morales F, Garcia-Quintanilla M, Cano DA, Casadesus J, Garcia-del Portillo F. Repression of the RcsC-YojN-RcsB phosphorelay by the IgaA protein is a requisite for Salmonella virulence. Mol Microbiol. 2004;53:1437–1449. doi: 10.1111/j.1365-2958.2004.04213.x. [DOI] [PubMed] [Google Scholar]
- Ebel W, Trempy JE. Escherichia coli RcsA, a positive activator of colanic acid capsular polysaccharide synthesis, functions to activate its own expression. J Bacteriol. 1999;181:577–584. doi: 10.1128/jb.181.2.577-584.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellermeier CD, Slauch JM. RtsA and RtsB coordinately regulate expression of the invasion and flagellar genes in Salmonella enterica serovar Typhimurium. J Bacteriol. 2003;185:5096–5108. doi: 10.1128/JB.185.17.5096-5108.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fang FC, Libby SJ, Castor ME, Fung AM. Isocitrate lyase (AceA) is required for Salmonella persistence but not for acute lethal infection in mice. Infect Immun. 2005;73:2547–2549. doi: 10.1128/IAI.73.4.2547-2549.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francez-Charlot A, Laugel B, Van Gemert A, Dubarry N, Wiorowski F, Castanie-Cornet MP, Gutierrez C, Cam K. RcsCDB His-Asp phosphorelay system negatively regulates the flhDC operon in Escherichia coli. Mol Microbiol. 2003;49:823–832. doi: 10.1046/j.1365-2958.2003.03601.x. [DOI] [PubMed] [Google Scholar]
- Galan JE. Interaction of Salmonella with host cells through the centisome 63 type III secretion system. Curr Opin Microbiol. 1999;2:46–50. doi: 10.1016/s1369-5274(99)80008-3. [DOI] [PubMed] [Google Scholar]
- Garcia-Calderon CB, Garcia-Quintanilla M, Casadesus J, Ramos-Morales F. Virulence attenuation in Salmonella enterica rcsC mutants with constitutive activation of the Rcs system. Microbiology. 2005;151:579–588. doi: 10.1099/mic.0.27520-0. [DOI] [PubMed] [Google Scholar]
- Gartenberg MR, Wang JC. Positive supercoiling of DNA greatly diminishes mRNA synthesis in yeast. Proc Natl Acad Sci U S A. 1992;89:11461–11465. doi: 10.1073/pnas.89.23.11461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gralla JD. Activation and repression of E. coli promoters. Curr Opin Genet Dev. 1996;6:526–530. doi: 10.1016/s0959-437x(96)80079-7. [DOI] [PubMed] [Google Scholar]
- Guillier M, Gottesman S, Storz G. Modulating the outer membrane with small RNAs. Genes Dev. 2006;20:2338–2348. doi: 10.1101/gad.1457506. [DOI] [PubMed] [Google Scholar]
- Guzman LM, Belin D, Carson MJ, Beckwith J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J Bacteriol. 1995;177:4121–4130. doi: 10.1128/jb.177.14.4121-4130.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagiwara D, Sugiura M, Oshima T, Mori H, Aiba H, Yamashino T, Mizuno T. Genome-wide analyses revealing a signaling network of the RcsC-YojN-RcsB phosphorelay system in Escherichia coli. J Bacteriol. 2003;185:5735–5746. doi: 10.1128/JB.185.19.5735-5746.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YH, Ferrieres L, Clarke DJ. The role of the Rcs phosphorelay in Enterobacteriaceae. Res Microbiol. 2006;157:206–212. doi: 10.1016/j.resmic.2005.11.005. [DOI] [PubMed] [Google Scholar]
- Laubacher ME, Ades SE. The Rcs phosphorelay is a cell envelope stress response activated by peptidoglycan stress and contributes to intrinsic antibiotic resistance. J Bacteriol. 2008;190:2065–2074. doi: 10.1128/JB.01740-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leng F, McMacken R. Potent stimulation of transcription-coupled DNA supercoiling by sequence-specific DNA-binding proteins. Proc Natl Acad Sci U S A. 2002;99:9139–9144. doi: 10.1073/pnas.142002099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lilley DM, Higgins CF. Local DNA topology and gene expression: the case of the leu-500 promoter. Mol Microbiol. 1991;5:779–783. doi: 10.1111/j.1365-2958.1991.tb00749.x. [DOI] [PubMed] [Google Scholar]
- Liu LF, Wang JC. Supercoiling of the DNA template during transcription. Proc Natl Acad Sci U S A. 1987;84:7024–7027. doi: 10.1073/pnas.84.20.7024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Majdalani N, Gottesman S. The Rcs phosphorelay: a complex signal transduction system. Annu Rev Microbiol. 2005;59:379–405. doi: 10.1146/annurev.micro.59.050405.101230. [DOI] [PubMed] [Google Scholar]
- Majdalani N, Heck M, Stout V, Gottesman S. Role of RcsF in signaling to the Rcs phosphorelay pathway in Escherichia coli. J Bacteriol. 2005;187:6770–6778. doi: 10.1128/JB.187.19.6770-6778.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malakooti J, Ely B, Matsumura P. Molecular characterization, nucleotide sequence, and expression of the fliO, fliP, fliQ, and fliR genes of Escherichia coli. J Bacteriol. 1994;176:189–197. doi: 10.1128/jb.176.1.189-197.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maloy SR, Nunn WD. Genetic regulation of the glyoxylate shunt in Escherichia coli K-12. J Bacteriol. 1982;149:173–180. doi: 10.1128/jb.149.1.173-180.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okochi M, Kurimoto M, Shimizu K, Honda H. Increase of organic solvent tolerance by overexpression of manXYZ in Escherichia coli. Appl Microbiol Biotechnol. 2007;73:1394–1399. doi: 10.1007/s00253-006-0624-y. [DOI] [PubMed] [Google Scholar]
- Ophir T, Gutnick DL. A role for exopolysaccharides in the protection of microorganisms from desiccation. Appl Environ Microbiol. 1994;60:740–745. doi: 10.1128/aem.60.2.740-745.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orosz A, Boros I, Venetianer P. Analysis of the complex transcription termination region of the Escherichia coli rrnB gene. Eur J Biochem. 1991;201:653–659. doi: 10.1111/j.1432-1033.1991.tb16326.x. [DOI] [PubMed] [Google Scholar]
- Park YK, Bearson B, Bang SH, Bang IS, Foster JW. Internal pH crisis, lysine decarboxylase and the acid tolerance response of Salmonella typhimurium. Mol Microbiol. 1996;20:605–611. doi: 10.1046/j.1365-2958.1996.5441070.x. [DOI] [PubMed] [Google Scholar]
- Plumbridge J. Control of the expression of the manXYZ operon in Escherichia coli: Mlc is a negative regulator of the mannose PTS. Mol Microbiol. 1998;27:369–380. doi: 10.1046/j.1365-2958.1998.00685.x. [DOI] [PubMed] [Google Scholar]
- Pratt LA, Hsing W, Gibson KE, Silhavy TJ. From acids to osmZ: multiple factors influence synthesis of the OmpF and OmpC porins in Escherichia coli. Mol Microbiol. 1996;20:911–917. doi: 10.1111/j.1365-2958.1996.tb02532.x. [DOI] [PubMed] [Google Scholar]
- Prigent-Combaret C, Vidal O, Dorel C, Lejeune P. Abiotic surface sensing and biofilm-dependent regulation of gene expression in Escherichia coli. J Bacteriol. 1999;181:5993–6002. doi: 10.1128/jb.181.19.5993-6002.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson SM, Higgins CF, Lilley DM. The genetic control of DNA supercoiling in Salmonella typhimurium. EMBO J. 1984;3:1745–1752. doi: 10.1002/j.1460-2075.1984.tb02041.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning. Cold Spring Harbor: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- Sledjeski DD, Gottesman S. Osmotic shock induction of capsule synthesis in Escherichia coli K-12. J Bacteriol. 1996;178:1204–1206. doi: 10.1128/jb.178.4.1204-1206.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Storz G, Opdyke JA, Zhang A. Controlling mRNA stability and translation with small, noncoding RNAs. Curr Opin Microbiol. 2004;7:140–144. doi: 10.1016/j.mib.2004.02.015. [DOI] [PubMed] [Google Scholar]
- Tan J, Shu L, Wu HY. Activation of the leu-500 promoter by adjacent transcription. J Bacteriol. 1994;176:1077–1086. doi: 10.1128/jb.176.4.1077-1086.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Travers A, Muskhelishvili G. A common topology for bacterial and eukaryotic transcription initiation? EMBO Rep. 2007;8:147–151. doi: 10.1038/sj.embor.7400898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang JC, Lynch AS. Transcription and DNA supercoiling. Curr Opin Genet Dev. 1993;3:764–768. doi: 10.1016/s0959-437x(05)80096-6. [DOI] [PubMed] [Google Scholar]
- Wang JC. DNA topoisomerases. Annu Rev Biochem. 1996;65:635–692. doi: 10.1146/annurev.bi.65.070196.003223. [DOI] [PubMed] [Google Scholar]
- Wang Q, Frye JG, McClelland M, Harshey RM. Gene expression patterns during swarming in Salmonella typhimurium: genes specific to surface growth and putative new motility and pathogenicity genes. Mol Microbiol. 2004;52:169–187. doi: 10.1111/j.1365-2958.2003.03977.x. [DOI] [PubMed] [Google Scholar]
- Wang Q, Suzuki A, Mariconda S, Porwollik S, Harshey RM. Sensing wetness: a new role for the bacterial flagellum. EMBO J. 2005;24:2034–2042. doi: 10.1038/sj.emboj.7600668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Q, Zhao Y, McClelland M, Harshey RM. The RcsCDB signaling system and swarming motility in Salmonella enterica serovar Typhimurium: dual regulation of flagellar and SPI-2 virulence genes. J Bacteriol. 2007;189:8447–8457. doi: 10.1128/JB.01198-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waterman SR, Holden DW. Functions and effectors of the Salmonella pathogenicity island 2 type III secretion system. Cell Microbiol. 2003;5:501–511. doi: 10.1046/j.1462-5822.2003.00294.x. [DOI] [PubMed] [Google Scholar]
- Wehland M, Bernhard F. The RcsAB box. Characterization of a new operator essential for the regulation of exopolysaccharide biosynthesis in enteric bacteria. J Biol Chem. 2000;275:7013–7020. doi: 10.1074/jbc.275.10.7013. [DOI] [PubMed] [Google Scholar]
- Wilusz JE, Sunwoo H, Spector DL. Long noncoding RNAs: functional surprises from the RNA world. Genes Dev. 2009;23:1494–1504. doi: 10.1101/gad.1800909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu HY, Fang M. DNA supercoiling and transcription control: a model from the study of suppression of the leu-500 mutation in Salmonella typhimurium topA- strains. Prog Nucleic Acid Res Mol Biol. 2003;73:43–68. doi: 10.1016/s0079-6603(03)01002-x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Suppl. Table 1. Primers used in indicated experiments.
Suppl. Table 2. Extent of deletion in genes or regulatory regions.