

Abstract
Learning is often prevented by events that occur after training, an outcome that is usually attributed to the disruption of consolidation – the transfer of learning to long-term memory. Here, we provide evidence from perceptual learning that improvements in performance can also be blocked by intervening events that occur during the acquisition phase of learning – the period of active practice. Listeners improved on each of two conditions of auditory temporal-interval discrimination (100 and 350 ms) when the two were practiced consecutively, even though that is a classic disruption-of-consolidation regimen. However, when practice on these two conditions was interleaved, there was no learning on either condition. The failure to improve in the interleaved case indicates that, at least in some circumstances, learning can be prevented during acquisition by events that do not disrupt consolidation itself. These results thus suggest that acquisition and consolidation are distinct phases in human learning.
Keywords: Perceptual learning, auditory learning, acquisition, consolidation, generalization, temporal interval discrimination
Introduction
Learning of new skills has often been conceptualized as consisting of two functional stages: An initial acquisition phase during which the experiences that have the potential to induce learning are obtained, usually through practice, and held in some form of short-term memory, and a subsequent consolidation phase in which learning stabilizes in long term memory and becomes immune to interference (e.g., Brashers-Krug et al., 1996; Shadmehr and Brashers-Krug, 1997; Roth et al., 2005; Seitz et al., 2005). The relationship between these two phases however remains unclear. To gain insight into this relationship, we asked whether learning on a perceptual discrimination task was susceptible to the same types of interference during the acquisition and consolidation phases. If not, this would suggest that the two stages are qualitatively distinct.
We are aware of only one previous investigation of the relationship between acquisition and consolidation in non-declarative learning. Zach et al. (2005) reported that the acquisition and consolidation of a target sensorimotor skill were differentially affected by the introduction of training on a non-target sensory motor task, depending on the characteristics of the non-target task and on the order of practice. They concluded from these results that acquisition and consolidation are, at least to some extent, independent in sensorimotor learning. Here we used a conceptually similar approach to investigate the relationship between acquisition and consolidation in perceptual learning. The presence of a prolonged consolidation phase has been shown using an interference paradigm for a visual hyperacuity task (Seitz et al., 2005), but there are no previous reports of the extent to which intervening training differentially affects acquisition and consolidation in perceptual learning.
We trained listeners on two conditions of auditory temporal-interval discrimination (standards: 100 ms, 1 kHz and 350 ms, 1 kHz) during each of 6 training sessions, using one of two training regimens, and assessed both learning on the trained conditions and generalization to an untrained stimulus frequency and to untrained temporal intervals. We chose to train temporal-interval discrimination because listeners reliably improve on this task when they practice a single condition at a time, so there was a good baseline for exploring the potential disruption of learning following training on two conditions. In addition, learning on this task generalizes in a distinct pattern, spreading to untrained stimulus frequencies but not to untrained intervals (Wright et al., 1997; Karmarkar and Buonomano, 2003). Thus, an assessment of generalization after two-condition training provided a further opportunity to evaluate how these training paradigms affected the overall learning pattern. The presence of generalization to any untrained stimulus would confirm that a given two-condition training regimen had induced learning. Generalization to an untrained frequency, but not to an untrained interval would indicate that this learning had similar characteristics to that induced by single-condition training.
The two training regimens were intended to reveal the vulnerability of learning on this task to intervening events that occurred during acquisition and consolidation. In one regimen, during each training session, the two conditions were presented consecutively, with 360 trials of the 100-ms condition followed by 360 trials of the 350-ms condition. Practicing 360 trials per session on the 100-ms condition alone yields learning on that condition (Wright and Sabin, 2007). Therefore, a failure of learning caused by the subsequent training of another condition would suggest a disruption of the consolidation of learning. In the other regimen, the two conditions were interleaved during each session, in sets of 120 trials, again for a total of 360 trials per condition. Thus, the per-session practice (acquisition) period on each condition was interrupted by training on the other condition. Therefore, a failure of learning with this regimen would suggest a disruption of learning during the acquisition phase. We reasoned that if the learning on temporal-interval discrimination was disrupted in one regimen, but not the other, it would suggest that events that affect learning during one stage (acquisition or consolidation) do not affect learning during the other, and therefore that these two stages are distinct.
Materials and Methods
Organization of the experiment
Three groups of listeners participated in this experiment, two trained groups—interleaved (n = 10) and consecutive (n = 8) -- and a control group (n = 11). During their first visit to the lab, all listeners participated in a screening session in which their hearing was assessed and their ability to perform auditory discrimination tasks confirmed. Subsequently all listeners completed pre- and post-test sessions, conducted 9–14 days apart, in which they were tested on seven conditions of auditory temporal-interval discrimination (described below). In between the pre- and post-test sessions, trained listeners practiced for 6 daily training sessions on one of two training regimens (described below). The controls received no such training.
Task, stimuli and procedure
Listeners performed an auditory temporal-interval discrimination task delivered in a 2-alternative forced choice (2AFC) paradigm. On each trial, two pairs of tone pips were presented. In one pair (the standard) the onsets of the two tones were separated by a fixed temporal interval of t ms. In the other pair (the comparison) the two onsets were separated by the fixed t plus a variable interval (Δt). Each tone pip had a total duration of 15 ms, including 5-ms raised-cosine onset/offset ramps. All tones in a given condition had the same frequency. On each trial, the standard and the comparison pairs were presented in random order and listeners had to determine which pair was the comparison (the ‘longer’) pair. The onsets of the standard and comparison pairs were separated by 900 ms (except for the 500-ms condition in which the onsets were separated by 1250 ms). Visual feedback was provided for both correct and incorrect responses throughout the entire experiment.
The comparison interval was adjusted adaptively in 60-trial blocks to estimate the discrimination threshold. Δt increased following an incorrect response and decreased following three consecutive correct responses. Δt values at which Δt changed from increasing to decreasing or vice versa were labeled reversals. The mean of the largest even number of reversal values, after discarding the first three reversals, yielded an estimate of the Δt value at the 79% correct point on the psychometric function (Levitt, 1971). This value is referred to here as the discrimination threshold. The Δt on the first trial of every block was 0 ms, forcing the listeners to guess. Δt changed in steps equal to 10 % of the standard temporal interval until the third reversal and in 1 % steps thereafter. For each listener, all threshold estimates obtained in a given session for a given condition (4 during the pre- and post-tests; 6 during training) were averaged to establish a daily threshold. Thresholds are expressed as fractions of the standard temporal interval (Δt/t).
Pre- and post-test conditions
In the pre- and post-test sessions, all listeners were tested on a set of seven conditions: two conditions used in training (100- and 350-ms standard intervals with sound frequencies of 1 kHz) and five additional conditions differing from the trained ones in either the sound frequency (100- and 350-ms standard intervals with sound frequencies of 4 kHz) or temporal interval (50-, 200- and 500-ms standard intervals with sound frequencies of 1 kHz). Four consecutive threshold estimates were obtained in each condition, therefore the pre- and post-tests consisted of 28 threshold estimates each. The order of conditions was counterbalanced across listeners and kept the same between the pre- and post- tests
Training regimens
All trained listeners participated in six daily training sessions. For the consecutive and interleaved groups, each daily session consisted of 720 practice trials that were equally distributed between the two trained conditions (360 trials each, in 6 blocks of 60 trials) in one of two regimens. In the interleaved regimen the two conditions alternated every 120 trials (2 blocks), starting with the 100-ms condition. In the consecutive regimen all 360 trials of the 100-ms condition (6 blocks) were presented before the 360 trials of the 350-ms condition. In each of these two regimens, the presentation order of the two trained conditions was the same across all listeners and during all training sessions.
Comparison to single-condition training
For comparison, we also report how two single-condition training regimens, examined during the course of other investigations, affected performance on the trained condition (100-ms standard interval with a sound frequency of 1 kHz). In one, listeners practiced for 360 trials per session, all blocked together (blocked, n = 6; data from Wright and Sabin, 2007), forming a comparison to learning on the 100-ms condition of the consecutive regimen. The other single-condition regimen served as a comparison for the interleaved case. In this regimen, each daily session consisted of 360 practice trials on the trained condition interleaved, every 120 trials, with periods of silence during which participants completed a written symbol-to-number matching task (spaced, n = 9; unpublished data). The duration of the silent periods was equivalent to the time ordinarily taken to complete two 60-trial blocks of auditory temporal interval-discrimination. The symbol-to-number matching task was administered only to impose some uniformity on the experiences of the controls during the breaks. Generalization data for these two groups will be reported elsewhere.
Listeners
A total of 29 listeners (aged 17 – 30 years) participated in the main experiment (consecutive, interleaved and control groups) and 15 listeners in the single-condition training regimens used for comparison (blocked and spaced). All participants were naïve to psychoacoustic tasks and reported no history of hearing or learning problems. Listeners were paid for their participation. The Northwestern Institutional Review Board approved all experimental procedures.
Data analysis
Prior to statistical analysis, the data were inspected for normality using a Kolmogorov-Smirnov (KS) test. The data in all pre- and post-test conditions in each of the groups was approximately normally distributed (KS Z < 0.93, p > 0.35).
We defined learning as a significantly larger pre- to post-test gain in trained listeners compared to controls as determined by 2 time (pre- vs. post- test) x 2 group (trained vs. control) ANOVAs, with time as a repeated measure, that were conducted separately for each regimen and condition. We concluded that there was training-induced learning if the interaction term was significant (p ≤ 0.025 to control for the use of the same control group in comparison to two trained groups) or if the effect size of the interaction was large (> 0.8). The effect size (ES) of the interaction term gives an indication of the magnitude of the difference in the amount of pre- to post- test change between the trained group and controls. It was calculated as follows. A pre- to post- test change score was first computed for each listener, and then the mean of this change score for the controls was subtracted from that for the trained listeners. Finally, this difference was divided by the standard deviation across all listeners entering the calculation. A positive ES indicates greater change in the trained group relative to controls, with ES > 0.8 considered large.
An inspection of the individual pre- and post-test data prior to statistical analysis revealed that in five instances the post-test value of a given listener on a given condition was at least twice that of the corresponding pre-test value. These values were all from two control listeners. Three values were from one control, (post test Δt/t = 0.65, 0.70 and 0.49 on the 100-ms 1-kHz, 100-ms 4-kHz and 200-ms 1-kHz conditions, respectively) and two from another (post test Δt/t = 0.53 and 1.57 on the 350-ms 4-kHz and the 50-ms 1-kHz conditions, respectively). Because such changes to the worse were uncharacteristic of either the current sample (5/77 data points) or of pre- to post-test changes of controls in previous investigations of learning on temporal-interval discrimination, these data (both pre- and post-test values) were removed from the statistical analyses reported in the Results section. Removal of these control outlying data points changed the statistical outcome in one case only (consecutive listeners vs. controls, 350-ms, 4-kHz condition: p = 0.081 with outliers, p= 0.034 without outliers). There were no such outliers in the data of the trained listeners.
Results
Learning and generalization between the pre- and post-tests
Training resulted in two distinct outcomes for the two training regimens. The interleaved regimen resulted in no learning between the pre- and post-tests on any condition, whereas the consecutive regimen yielded both learning and generalization compared to untrained controls. No significant differences between the control group and either of the trained groups were observed in the pre-test (Figure 1, open diamonds; one-way ANOVA group effect, F(2,24) < 1.37, p ≥ 0.234, all pair-wise comparisons insignificant, p > 0.05), indicating that the initial level of performance was similar across the groups. The control group itself improved significantly on the 100-ms 4-kHz condition (p < 0.01), but not on any other condition (p ≥ 0.162), suggesting that exposure to the pre-test alone had little lasting influence on performance (Figure 1, leftmost set of data points in each panel).
Figure 1. Average group discrimination thresholds on the pre- (empty symbols) and post- (filled symbols) test sessions for the control, interleaved, and consecutive groups.
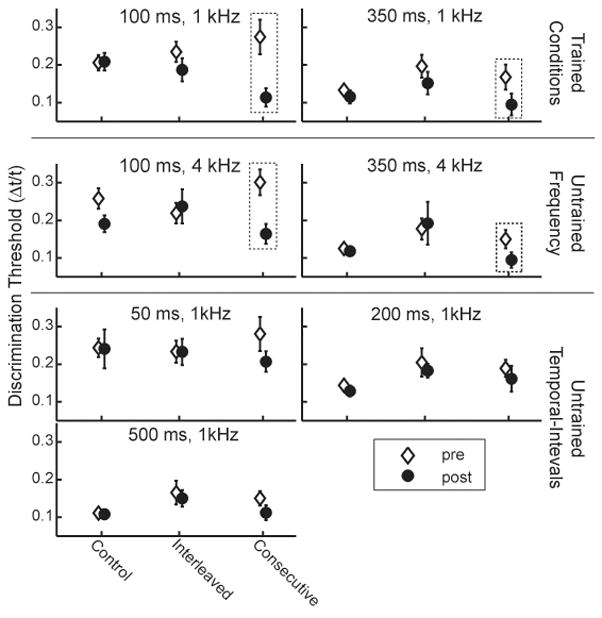
Results are shown for the two trained conditions (first row), and for untrained conditions that differed from the trained ones either only in sound frequency (second row) or temporal interval (third and fourth rows). Error bars denote ±1 standard error of the (group) mean. The dashed boxes denote that the trained group learned significantly more than the control group on that condition.
By our definition of learning (see Methods, Data Analysis), there was no significant pre- to post-test improvement on any of the trained or untrained conditions following training on the interleaved regimen. Listeners in the interleaved group did not learn significantly on either of the trained conditions (Figure 1, top row, middle set of data points in each panel), the untrained conditions sharing a temporal interval with the trained ones (middle row) or on any untrained temporal interval (bottom rows) (ANOVA, all F(1,18) < 2.62, p ≥ 0.124, all ES < 0.48). To determine whether individual trained listeners improved on one trained condition but not the other, thus obscuring a learning effect at the group level, we selected for each trained listener and control the trained condition on which improvement between the pre- and post-test threshold was greater, and compared pre- and post-test threshold on this “better” condition between the interleaved and control groups. These data are shown in Figure 2. For each listener the amount of improvement between the pre- and post-tests was plotted against the pre-test threshold. Irrespective of pretest threshold, the performance of the interleaved group was indistinguishable form controls even on the “better” condition (Figure 2 upper left panel; F = 1.55, p = 0.228, ES = 0.54). Naturally, there also was no learning on the condition on which the listeners improved the least (Figure 2, “poorer”, bottom left panel; F = 0.84, p = 0.373, ES = 0.43). It appears that this lack of learning with the interleaved regimen was not simply due to the passage of time between two successive encounters with each trained condition because the listeners who were trained with the spaced regimen did improve on their trained condition (from Δt/t = 0.19±0.05 at the pre-test to Δt/t = 0.11±0.06 at the post-test) relative to controls (F(1,17) = 5.31, p = 0.034, ES = 0.96).
Figure 2. Amount of improvement between the pre- and post-test sessions vs. initial performance for both the “better” (top) and “poorer” (bottom) of the two trained conditions. Results are shown for the interleaved (left column) consecutive (right column) and control (both columns) groups.
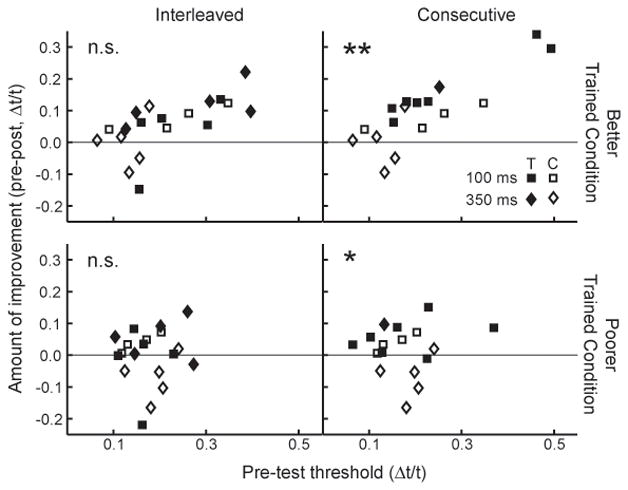
For both trained listeners (filled symbols) and controls (open symbols), the “better” condition was defined as the trained condition on which the difference between the pre- and post-test thresholds was the larger of the two. Listeners who improved more on the 100-ms condition are denoted with squares, those who improved more on the 350-ms condition are denoted with diamonds. Conditions in which trained listeners learned significantly more than controls are marked with * for p < 0.05 and ** for p < 0.01; n.s. indicates that there was no significant difference in improvement between the two groups.
In contrast to the lack of learning on the interleaved regimen, listeners trained on the consecutive regimen showed evidence of learning on both of the trained conditions as well as generalization to untrained conditions differing in frequency, but not temporal-interval from the trained conditions. Performance on both trained conditions improved with practice (Figure 1, top row, rightmost set of data points in each panel; 100-ms: F(1,16) = 11.98, p = 0.003, ES = 1.28; 350-ms: F(1,17) = 5.34, p = 0.034, ES = 0.92). As would be expected, the consecutive trained group also learned significantly on the “better” condition (Figure 2, top right panel; F(1,16) = 13.65, p = 0.003, ES = 1.27). However, demonstrating the large magnitude of the improvement, the consecutive-trained listeners learned significantly even on the condition on which they improved the least (Figure 2, “poorer”, bottom right panel; F(1,16) = 7.11, p = 0.017, ES = 1.08). These improvements were apparent across the entire range of pre-test thresholds. Learning on the 100-ms trained condition in this group was similar to that observed among listeners who were trained with the blocked regimen (see Wright and Sabin, 2007). Consecutive-trained listeners also showed evidence of generalization to the two untrained conditions sharing the same temporal intervals with, but differing in sound frequency from, the trained conditions (Figure 1 middle column, 100-ms, 4-kHz, F(1,16) = 3.91, p = 0.066, ES = 0.86; 350-ms, 4-kHz, F(1,15) = 5.68, p = 0.031, ES = 1.03), but not to any of the untrained temporal intervals (Figure 1, bottom 2 rows, p ≥ .103)1. This generalization pattern was similar to that obtained with single-condition regimens in which more intensive training was provided on the 100-ms, 1-kHz trained condition (Wright et al., 1997; Karmarkar and Buonomano, 2003).
Learning curves
The conclusion that consecutive-trained but not interleaved-trained listeners benefited from training received further support from analyses of the learning curves of the two groups. We used two indices of performance – average performance (defined for each listener as the average of all threshold estimates for a particular condition completed during a single session), and within-listener consistency (defined for each listener as the standard deviation of all threshold estimates completed for a particular condition during a single session). The mean group learning curves are shown in Figure 3. For each listener and condition, we fitted a regression line through the daily average or consistency values from each session, either including or excluding the pre- and post-tests sessions, against the log of the session number. We analyzed the slopes of these regression lines to determine whether improvement within a group was significantly different from 0. The consecutive-trained group improved significantly on both trained conditions using either measure of performance, regardless of the inclusion or exclusion of the pre- and post-tests, with one exception (average performance on the 350-ms condition across training sessions only; See Table 1). The interleaved-trained group on the other hand learned significantly only on one condition with one performance index (average performance in the 350-ms condition, both including and excluding the pre- and post-tests; see Table 1). This significant improvement in the interleaved group was not sufficient however to distinguish their pre- to post-test change from that of controls (Figure 1). Thus, two partially distinct learning patterns were observed during the training phase. Consecutive training resulted in significant improvement of both average performance and performance consistency on both trained intervals. Interleaved training on the other hand, resulted in no improvement in performance consistency on either trained interval with average performance improvement on the 350-ms interval only.
Figure 3. Group learning curves on average performance (top) and within-listener consistency (bottom).
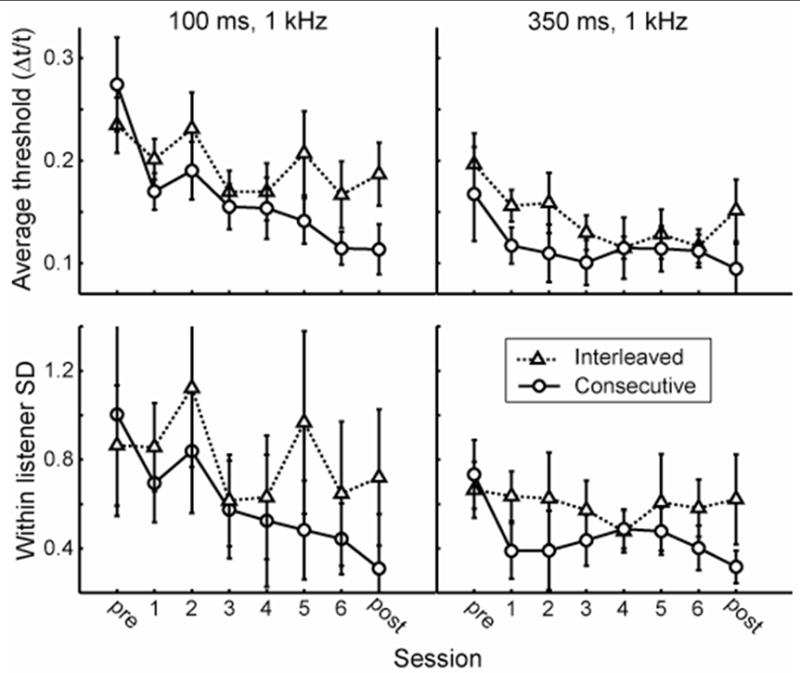
Results are shown for the interleaved (triangles) and consecutive (circles) groups for the 100-ms (left column) and the 350-ms (right column) conditions. For each listener, the two performance indices are defined as the mean (average performance) or standard deviation (within-listener consistency) of all threshold estimates completed for a particular condition during a single session. See Table 1 for the results of statistical analyses of these data.
Table 1.
Learning curve slopes and 95% confidence intervals
| Training sessions only | With pre- and post-tests | |||
|---|---|---|---|---|
| 100-ms | 350-ms | 100-ms | 350-ms | |
| Consecutive | ||||
| Average | −0.030 (−0.05, −0.011) | 0.002 (−0.021, 0.024) | −0.069 (−0.102, −0.036) | −0.020 (−0.039, −0.001) |
| Consistency | −0.018 (−0.035, −0.0004) | 0.006 (−0.01, 0.023) | −0.030 (−0.043, −0.018) | −0.009 (−0.018, −0.001) |
| Interleaved | ||||
| Average | −0.019 (−0.055, 0.016) | −0.025 (−0.035, −0.015) | −0.026 (−0.060, 0.007) | −0.031 (−0.053, −0.010) |
| Consistency | −0.012 (−0.028, 0.004) | −0.004 (−0.012, 0.004) | −0.009 (−0.024, 0.005) | −0.004 (−0.011, 0.003) |
Top value in each box is the mean learning curve slope; values in parentheses denote the 95% confidence interval (lower bound, upper bound). If the confidence interval is all in the negative range (slopes denoted in bold), the slope is significantly less than 0, indicating significant (p < 0.05) training phase learning for that condition and index of performance.
Discussion
The major goal of the present investigation was to determine whether the acquisition and consolidation of perceptual learning are differentially sensitive to interference induced by intervening events. To investigate this question, we trained listeners on auditory temporal-interval discrimination in one of two training regimens differing in the distribution of two trained conditions during each of six training sessions. In one regimen, the acquisition phase of each condition was interleaved with periods of practice on the other condition. This regimen yielded no learning between the pre- and post-test sessions on either condition. In contrast, practicing the conditions consecutively during each session, such that practice on the second condition started during the consolidation period of the first, resulted in learning on both. Further, this consecutive regimen yielded the same generalization pattern as was previously observed following single-condition training; that is, the learning generalized to two untrained conditions sharing the temporal-intervals of the trained conditions, but differing in tone frequency, but did not generalize to untrained temporal intervals. These outcomes suggest that an intervening event (training on a second condition) can have different effects depending on the learning phase during which it occurs. This difference implies that, at least for auditory temporal-interval discrimination, acquisition and consolidation are qualitatively different aspects of learning, potentially governed by distinct neural mechanisms.
Interleaved training: Disruption of acquisition
Intervening events are known to interfere with both perceptual (Roth et al., 2005; Seitz et al., 2005; Zhang et al., 2008) and motor (Brashers-Krug et al., 1996; Shadmehr and Brashers-Krug, 1997; Balas et al., 2007) learning. Those interruptions are typically attributed to a disruption of consolidation. Arguing against the idea that a failure in consolidation accounts for the lack of learning in the interleaved regimen here, there was significant learning on both trained conditions in the consecutive training regimen even though it had the characteristics of a typical disruption-of-consolidation experiment. In this regimen, listeners practiced one temporal-interval discrimination condition for 360 trials per day, an amount known to be sufficient for learning on this task when only a single condition is practiced (Wright and Sabin, 2007). However, here, the introduction of the second condition immediately after the first, which could have interrupted the consolidation of this learning, did not do so. Given the lack of interference of consolidation in the consecutive regimen it seems unlikely that disruption of consolidation can account for the failure of learning in the interleaved regimen. If each of the sets of 120 practice trials in the interleaved regimen were sufficient to initiate the consolidation of each trained condition, then there should have been no interruption of this consolidation on either condition by the introduction of the other one, just as was the case in the consecutive regimen in which we know consolidation was successful. This, however, was not the case. Instead, learning in the interleaved regimen was interrupted on both trained conditions.
It therefore appears that the lack of learning in the interleaved regimen stems from a failure in the acquisition phase of learning. This failure appears to have arisen from a true disruption of learning induced by alternating between the two trained conditions, rather than from the simple decay of the influence of the active training on each condition during practice on the other as indicated by the successful learning of the group that was trained on a single condition with the spaced regimen. It is interesting to note that during single-condition training on temporal-interval discrimination there are no obvious performance gains within the training sessions (during the acquisition periods), instead the improvements occur between sessions (Wright and Sabin, 2007). Thus, the present results indicate that the acquisition period on temporal-interval discrimination is susceptible to interference even when improvements within that period are not observed in the absence of interference.
The disruption of learning observed in the interleaved regimen in the current study adds to the known list of requirements that must be fulfilled during acquisition in order for successful learning to occur. Two such requirements observed in single-condition training are of particular interest here. First, for learning to occur on many tasks, the participant must actively practice that task (e.g., Shiu and Pashler, 1992; Ahissar and Hochstein, 1993; Polley et al., 2006). Second, the participant must receive sufficient training within each session (Hauptmann et al., 2005; Wright and Sabin, 2007). Indeed, practicing auditory temporal-interval discrimination for 50 trials per day (for 20 days) resulted in no learning (Rammsayer, 1994), even though daily practice for more trials per day (>360) for 5–10 days did (Wright et al., 1997; Wright and Sabin, 2007; Lapid et al., 2009). The current results suggest that, at least for some condition combinations, successful acquisition may have to fulfill a third requirement: the proper distribution of the training trials on each condition within each training session.
That interleaving two temporal intervals prevented acquisition in the present case suggests that the acquisition process did not proceed separately on each trained stimulus. Task performance adequate for the selection of the neural substrate to be modified and a sufficient amount of practice per session were provided even in the interleaved regimen, as indicated by the learning observed in the group who practiced sets of 120 trials interleaved with silent periods. Nevertheless acquisition was disrupted, possibly because the alternation between the two intervals prevented the buildup over time of the processes relevant for the acquisition of each interval. This may have happened either because these processes required shared resources that were insufficient or not efficiently distributed in the interleaved condition, or because they mutually inhibited each other.
Practicing two different conditions within each session prevented learning in the current investigation, but does not always do so. For example, there was learning on visual contrast (Yu et al., 2004) and motion-direction (Kuai et al., 2005) discrimination when each of four conditions were interleaved in sequences of fewer than 50 trials, on auditory frequency discrimination when each of three conditions were systematically interleaved on a trial by trial basis (Demany and Semal, 2002) and when one of five conditions was presented at random on each and every trial (Amitay et al., 2005), and even on auditory temporal interval-discrimination when one of two conditions was presented at random on each and every trial (Karmarkar and Buonomano, 2003). Multiple factors could contribute to the difference between the present lack of learning on the interleaved regimen and the presence of learning in these other investigations. We consider three possibilities here. First, learning on different tasks may be governed by distinct underlying mechanisms that have task-specific requirements for the distribution of training trials (see Wright and Zhang, 2009 for a review of the distinct learning patterns across different auditory tasks). Second, different learning mechanisms may be engaged when multiple trained conditions alternate rapidly (Demany and Semal, 2002; Karmarkar and Buonomano, 2003; Amitay et al., 2005) as opposed to slowly (current study). Some support for this idea comes from the finding that learning on visual contrast (Yu et al., 2004) and motion-direction (Kuai et al., 2005) discrimination occurred when four conditions alternated relatively slowly during training, but not when those conditions were randomized trial by trial. Third, just as some conditions but not others interfere with consolidation, there may be specific combinations of conditions that prevent acquisition when they are interleaved (e.g., current conditions) or randomly presented (e.g., the conditions used by Kuai et al., 2005) and other combinations that do not (e.g., the conditions used by Demany and Semal 2002, Amitay et al., 2005 and Karmarkar and Buonomano, 2003).
Consecutive training: No disruption of consolidation
The presence of learning on temporal-interval discrimination in the consecutive regimen indicates that both acquisition and consolidation were successful; therefore, the same event--training on a second temporal interval--that prevented learning on that task when encountered during the acquisition period (interleaved regimen) did not interfere with learning when encountered during the consolidation phase (consecutive regimen), even though this paradigm disrupts consolidation on other tasks (Seitz et al., 2005; Balas et al., 2007; Korman et al., 2007). The learning observed in the consecutive regimen does not appear to have resulted from the generalization of learning on one temporal interval to the other. Had this been the case, we would have expected learning to generalize to untrained temporal intervals as well, but it did not. The pattern of learning generalization following consecutive training was the same as that resulting from single-condition training on this task (Wright et al., 1997; Karmarkar and Buonomano, 2003), with generalization across auditory frequencies for the trained temporal intervals and no generalization to untrained intervals. Particularly noteworthy, with the consecutive training, learning did not even generalize to an untrained interval (200 ms) that was in between the two successfully learned ones (100 ms and 350 ms). Thus consecutive training did not broaden the pattern of generalization relative to single-condition training. Consequently we are led to the conclusion that in the consecutive regimen the learning on the two intervals occurred separately. The lack of interference of consolidation between the two temporal intervals in the current study raises an interesting possibility, namely that if learning does not generalize between two stimuli in single-condition training, consecutive practice on those two stimuli will not disrupt consolidation. This idea is also consistent with the finding of Seitz et al. (2005) that consecutive training on two visual hyperacuity tasks was successful unless the two stimuli were presented at the same orientation and the same retinotopic location – conditions under which single-condition visual training does generalize.
Implications for the neural mechanisms underlying temporal-interval discrimination learning
Taken together, the current results suggest that learning on auditory temporal-interval discrimination proceeds along two independent phases – acquisition and consolidation, each of which is differentially influenced by intervening events. Further, that consecutive but not interleaved training resulted in improvement suggests that whereas acquisition did not proceed separately on each trained interval, once acquisition was successful and consolidation initiated, the two temporal intervals were treated separately. One potential account for the apparent independence of the acquisition and consolidation of temporal-interval discrimination is that each phase engaged a different neural substrate. Indeed there is evidence that following more intensive training, primary sensory areas can become less engaged in the performance of the trained task than after less intensive training (Nieuwenhuis et al., 2008), whereas non sensory areas can become more engaged (Takashima et al., 2007). An alternative account is that acquisition and consolidation occurred within the same neural substrate such that acquisition resulted in changes that allowed this substrate to now treat each interval independently (see Dudai, 2004 for review). Evidence for this view comes from observations that the same sensory and motor pathways that are involved in naive performance can reorganize and become more robustly activated with training (Penhune and Doyon, 2005; Sigman et al., 2005).
To the extent that the present learning modified temporal processing rather than cognitive processes involved with other aspects of the discrimination task, the current results are somewhat more consistent with models that attribute temporal processing to a distributed network rather than to those based on a single centralized mechanism (see Ivry and Schlerf, 2008 for a recent review). Central models of timing propose that time is processed by a dedicated mechanism, typically described as either a single internal clock (e.g., the pacemaker-accumulator model) or as multiple clocks each dedicated to a specific interval. The idea of a single clock is inconsistent with the lack of generalization to untrained intervals seen here and elsewhere (Wright et al., 1997; Karmarkar and Buonomano, 2003). Multiple clocks could account for this temporal specificity. However, they are inconsistent with the finding that two temporal intervals are treated separately during consolidation but not during acquisition, unless it is assumed that the processes relevant for the acquisition of each interval are shared amongst the different clocks and that consolidation occurs locally. Distributed models of timing instead postulate that time is processed in distributed networks that are either dedicated to temporal processing (Harrington et al., 1998) or that yield timing information as an emergent property (Karmarkar and Buonomano, 2007). These models can explain the specificity of learning to the trained temporal interval. They also offer several possible scenarios to account for the distinction between acquisition and consolidation of interval learning reported here. For example, the successful consolidation of learning may have resulted in the modification of populations of duration tuned neurons, such as those reported to be found in the inferior colliculus of many species (Casseday et al., 1994; Brand et al., 2000; Edwards et al., 2002), whereas acquisition may have affected a different time-encoding substrate that was not specifically tuned to any particular interval (hence the interference between the intervals during acquisition). Another possibility is that both acquisition and consolidation occurred in a cortical network that could be dynamically tuned to particular intervals (Mauk and Buonomano, 2004; Karmarkar and Buonomano, 2007). If so, consecutive training created a network with stable states corresponding to each of the intervals, whereas the interleaved regimen failed to do so.
Acquisition and consolidation as distinct phases of learning
The outcomes of the current investigation demonstrate distinct and at least partially independent acquisition and consolidation phases of auditory perceptual learning. A similar distinction has been identified in a single investigation of sensorimotor learning, where the acquisition and consolidation of a target condition were differentially affected by additional training on one of two non-target conditions (Zach et al., 2005). Furthermore, the distinction between acquisition and consolidation appears to characterize training regimens in which learning is rapid and occurs within a single training session (Zach et al., 2005) as well as regimens in which learning is more gradual and is not observed during any single training session (current study). Therefore, distinct learning phases may be a feature of improvements that occur over different time scales and a characteristic of both perceptual and motor learning.
Acknowledgments
We thank Julia Huyck, Nicole Marrone, Andrew Sabin, Dan Sanes and Yuxuan Zhang for their assistance at various stages of this project. We also thank Christophe Micheyl and an anonymous reviewer for their insightful comments on a previous version of this manuscript.
This work was supported by NIH/NIDCD, the Hugh Knowles Center for Clinical and Basic Science in Hearing and its Disorders (at Northwestern University) and by the Israel Science Foundation (LHSI 1842/07).
Footnotes
Note that the same conclusions were reached when the data from the present trained listeners were compared to those of a larger control group comprised of the current controls and an additional 9–14 controls from whom data were available on a subset of the conditions examined here. These additional controls completed different subsets of the current conditions, among other conditions not tested here, as part of other investigations (unpublished data). The different condition combinations did not appear to have any systematic effect on the performance of controls on the current conditions. When all of the available control data were included in the analyses, the interleaved group still showed no evidence of learning or generalization (all p ≥ 0.16, all ES < 0.45). However, the consecutive trained listeners learned significantly on both trained conditions (100-ms, 1-kHz p = 0.003, ES = 1.14; 350-ms, 1-kHz p = 0.013, ES = 0.92) and significantly generalized their learning to the untrained sound frequencies (100-ms, 4-kHz, p = 0.004, ES = 1.15; 350-ms, 4-kHz, p = 0.012, ES = 1.06), but not to the untrained temporal intervals (p ≥ 0.18).
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References Cited
- Ahissar M, Hochstein S. Attentional control of early perceptual learning. Proceedings of the National Academy of Sciences of the United States of America. 1993;90:5718–5722. doi: 10.1073/pnas.90.12.5718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amitay S, Hawkey DJ, Moore DR. Auditory frequency discrimination learning is affected by stimulus variability. Percept Psychophys. 2005;67:691–698. doi: 10.3758/bf03193525. [DOI] [PubMed] [Google Scholar]
- Balas M, Roitenberg N, Giladi N, Karni A. When practice does not make perfect: well-practiced handwriting interferes with the consolidation phase gains in learning a movement sequence. Exp Brain Res. 2007;178:499–508. doi: 10.1007/s00221-006-0757-3. [DOI] [PubMed] [Google Scholar]
- Brand A, Urban R, Grothe B. Duration tuning in the mouse auditory midbrain. J Neurophysiol. 2000;84:1790–1799. doi: 10.1152/jn.2000.84.4.1790. [DOI] [PubMed] [Google Scholar]
- Brashers-Krug T, Shadmehr R, Bizzi E. Consolidation in human motor memory. Nature. 1996;382:252–255. doi: 10.1038/382252a0. [DOI] [PubMed] [Google Scholar]
- Casseday JH, Ehrlich D, Covey E. Neural tuning for sound duration: role of inhibitory mechanisms in the inferior colliculus. Science. 1994;264:847–850. doi: 10.1126/science.8171341. [DOI] [PubMed] [Google Scholar]
- Demany L, Semal C. Learning to perceive pitch differences. J Acoust Soc Am. 2002;111:1377–1388. doi: 10.1121/1.1445791. [DOI] [PubMed] [Google Scholar]
- Dudai Y. The neurobiology of consolidations, or, how stable is the engram? Annu Rev Psychol. 2004;55:51–86. doi: 10.1146/annurev.psych.55.090902.142050. [DOI] [PubMed] [Google Scholar]
- Edwards CJ, Alder TB, Rose GJ. Auditory midbrain neurons that count. Nat Neurosci. 2002;5:934–936. doi: 10.1038/nn916. [DOI] [PubMed] [Google Scholar]
- Harrington DL, Haaland KY, Knight RT. Cortical networks underlying mechanisms of time perception. J Neurosci. 1998;18:1085–1095. doi: 10.1523/JNEUROSCI.18-03-01085.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauptmann B, Reinhart E, Brandt SA, Karni A. The predictive value of the leveling off of within session performance for procedural memory consolidation. Brain Res Cogn Brain Res. 2005;24:181–189. doi: 10.1016/j.cogbrainres.2005.01.012. [DOI] [PubMed] [Google Scholar]
- Ivry RB, Schlerf JE. Dedicated and intrinsic models of time perception. Trends Cogn Sci. 2008;12:273–280. doi: 10.1016/j.tics.2008.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karmarkar UR, Buonomano DV. Temporal specificity of perceptual learning in an auditory discrimination task. Learn Mem. 2003;10:141–147. doi: 10.1101/lm.55503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karmarkar UR, Buonomano DV. Timing in the absence of clocks: encoding time in neural network states. Neuron. 2007;53:427–438. doi: 10.1016/j.neuron.2007.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korman M, Doyon J, Doljansky J, Carrier J, Dagan Y, Karni A. Daytime sleep condenses the time course of motor memory consolidation. Nat Neurosci. 2007;10:1206–1213. doi: 10.1038/nn1959. [DOI] [PubMed] [Google Scholar]
- Kuai SG, Zhang JY, Klein SA, Levi DM, Yu C. The essential role of stimulus temporal patterning in enabling perceptual learning. Nat Neurosci. 2005;8:1497–1499. doi: 10.1038/nn1546. [DOI] [PubMed] [Google Scholar]
- Lapid E, Ulrich R, Rammsayer T. Perceptual learning in auditory temporal discrimination: no evidence for a cross-modal transfer to the visual modality. Psychon Bull Rev. 2009;16:382–389. doi: 10.3758/PBR.16.2.382. [DOI] [PubMed] [Google Scholar]
- Levitt H. Transformed up-down methods in psychoacoustics. J Acoust Soc Am. 1971;49(Suppl 2):467. [PubMed] [Google Scholar]
- Mauk MD, Buonomano DV. The neural basis of temporal processing. Annu Rev Neurosci. 2004;27:307–340. doi: 10.1146/annurev.neuro.27.070203.144247. [DOI] [PubMed] [Google Scholar]
- Nieuwenhuis IL, Takashima A, Oostenveld R, Fernandez G, Jensen O. Visual areas become less engaged in associative recall following memory stabilization. Neuroimage. 2008;40:1319–1327. doi: 10.1016/j.neuroimage.2007.12.052. [DOI] [PubMed] [Google Scholar]
- Penhune VB, Doyon J. Cerebellum and M1 interaction during early learning of timed motor sequences. Neuroimage. 2005;26:801–812. doi: 10.1016/j.neuroimage.2005.02.041. [DOI] [PubMed] [Google Scholar]
- Polley DB, Steinberg EE, Merzenich MM. Perceptual learning directs auditory cortical map reorganization through top-down influences. J Neurosci. 2006;26:4970–4982. doi: 10.1523/JNEUROSCI.3771-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rammsayer TH. Effects of practice and signal energy on duration discrimination of brief auditory intervals. Percept Psychophys. 1994;55:454–464. doi: 10.3758/bf03205302. [DOI] [PubMed] [Google Scholar]
- Roth DA, Kishon-Rabin L, Hildesheimer M, Karni A. A latent consolidation phase in auditory identification learning: time in the awake state is sufficient. Learn Mem. 2005;12:159–164. doi: 10.1101/87505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seitz AR, Yamagishi N, Werner B, Goda N, Kawato M, Watanabe T. Task-specific disruption of perceptual learning. Proc Natl Acad Sci U S A. 2005;102:14895–14900. doi: 10.1073/pnas.0505765102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shadmehr R, Brashers-Krug T. Functional stages in the formation of human long-term motor memory. J Neurosci. 1997;17:409–419. doi: 10.1523/JNEUROSCI.17-01-00409.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shiu LP, Pashler H. Improvement in line orientation discrimination is retinally local but dependent on cognitive set. Perception & psychophysics. 1992;52:582–588. doi: 10.3758/bf03206720. [DOI] [PubMed] [Google Scholar]
- Sigman M, Pan H, Yang Y, Stern E, Silbersweig D, Gilbert CD. Top-down reorganization of activity in the visual pathway after learning a shape identification task. Neuron. 2005;46:823–835. doi: 10.1016/j.neuron.2005.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takashima A, Nieuwenhuis IL, Rijpkema M, Petersson KM, Jensen O, Fernandez G. Memory trace stabilization leads to large-scale changes in the retrieval network: a functional MRI study on associative memory. Learn Mem. 2007;14:472–479. doi: 10.1101/lm.605607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright BA, Sabin AT. Perceptual learning: how much daily training is enough? Exp Brain Res. 2007;180:727–736. doi: 10.1007/s00221-007-0898-z. [DOI] [PubMed] [Google Scholar]
- Wright BA, Zhang Y. Insights into human auditory processing gained from perceptual learning. In: Gazzaniga MS, editor. The cognitive neurosciences. 4. Cambridge, Ma: MIT Press; 2009. pp. 353–366. [Google Scholar]
- Wright BA, Buonomano DV, Mahncke HW, Merzenich MM. Learning and generalization of auditory temporal-interval discrimination in humans. J Neurosci. 1997;17:3956–3963. doi: 10.1523/JNEUROSCI.17-10-03956.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu C, Klein SA, Levi DM. Perceptual learning in contrast discrimination and the (minimal) role of context. J Vis. 2004;4:169–182. doi: 10.1167/4.3.4. [DOI] [PubMed] [Google Scholar]
- Zach N, Kanarek N, Inbar D, Grinvald Y, Milestein T, Vaadia E. Segregation between acquisition and long-term memory in sensorimotor learning. Eur J Neurosci. 2005;22:2357–2362. doi: 10.1111/j.1460-9568.2005.04415.x. [DOI] [PubMed] [Google Scholar]
- Zhang JY, Kuai SG, Xiao LQ, Klein SA, Levi DM, Yu C. Stimulus coding rules for perceptual learning. PLoS Biol. 2008;6:e197. doi: 10.1371/journal.pbio.0060197. [DOI] [PMC free article] [PubMed] [Google Scholar]