

Abstract
Although neuronal synchronization has been shown to exist in primary motor cortex (MI), very little is known about its possible contribution to coding of movement. By using cross-correlation techniques from multi-neuron recordings in MI, we observed that activity of neurons commonly synchronized around the time of movement initiation. For some cell pairs, synchrony varied with direction in a manner not readily predicted by the firing of either neuron. Information theoretic analysis demonstrated quantitatively that synchrony provides information about movement direction beyond that expected by simple rate changes. Thus, MI neurons are not simply independent encoders of movement parameters but rather engage in mutual interactions that could potentially provide an additional coding dimension in cortex.
It is well known that the discharge of single primary motor cortex (MI) neurons covaries with a number of arm movement-related parameters, including force, position, distance, speed, and direction (1–6). Particular emphasis has been given to the hypothesis that the intended direction of an arm movement is encoded by populations of MI neurons (7). In either single cell or population-coding hypotheses, however, it has been generally assumed that average firing rate mediates the representation of movement parameters. Synchronization of firing of ensembles of neurons has recently received attention as a novel coding dimension because of its prevalence in cortex and its possible functional relevance for perceptual binding (8, 9). Early work evaluated synchronous discharge of pairs of MI neurons largely in the context of identifying local connectivity patterns (10–12), rather than investigating its possible information content. More recent studies by using multi-electrode recordings in frontal cortex have suggested that the synchronous firing of two cells can provide general information regarding the decision to move (13) or expectation of predictable cues (14). However, none of these studies have addressed the relationship between synchrony and movement features. By using a multielectrode array to record from groups of neurons, we provide evidence that neuronal synchronization within MI carries information about movement direction beyond that available from the average firing rate of the same set of neurons.
METHODS
Behavioral Task.
Two macaque monkeys (Macaca fascicularis) were operantly conditioned to perform a reaching task with their left arm. Animals moved a two-joint manipulandum in the horizontal plane to direct a cursor from a central hold position to one of two (left or right) or eight possible (radially positioned) targets that were displayed on a computer monitor in front of the monkey. A trial was composed of three epochs: a “hold” period during which time the monkey had to maintain the cursor at the hold position for 0.5 s, a random 1–1.5 s “instructed delay” period during which one of the radial targets appeared but movement was withheld, and a movement period initiated by target blinking (time to movement onset ∼365 ms). The position of the manipulandum was monitored by using a digitizing tablet (Numonics, Montgomeryville, PA) sampled at 72 Hz.
Recording.
We used a silicon-based electrode array developed at the University of Utah (15, 16) to record neural discharge from multiple MI sites. The array was composed of 10 × 10 grid of 100 tapered electrodes, each 1.5 mm long with a 50-μm platinum-coated tip. Electrodes were separated by 400 μm. Typical impedance values were between 100 and 200 kΩ. Wires, which were attached to 22 of the possible 100 electrodes, led to connectors attached to the skull. The second monkey had connections from only 11 electrodes. The arrays were implanted into the arm area of MI on the right hemisphere. During a recording session, signals from up to 16 electrodes were amplified and recorded digitally onto disk at 20 kHz per channel (Datawave Technologies, Denver, CO). Only waveforms that crossed a threshold without surpassing an upper bound were stored (0.5 ms before and 1.0 ms after the threshold). There was no lock-out time between successive spike acquisitions so inter-spike intervals could be as small as 50 μs, the sampling interval of the analog to digital (A/D) board. By using up to 8 waveform features (i.e., spike height and width), manual and automatic feature clustering was used to isolate single units (Datawave Technologies). Autocorrelation functions were computed to verify single unit isolation. In addition, all synchronous spike waveforms from many pairs of neurons that were reported in this work were carefully examined to ensure that there were no noise artifacts and that there was no cross-talk between channels. As a final check, cross-correlations were computed by using 100-μs bins; no data presented here exhibited precise peaks in synchronous firing at that time scale which might suggest noise artifacts.
Data from six recording sessions were analyzed for synchrony. Four recording sessions spaced over 1 year were analyzed from one monkey, and two recording sessions were analyzed from the second monkey spaced over 2 days. We were able to isolate 15 units on average during a recording session. Typically, only 1 unit/electrode was isolated although occasionally 2 units were isolated (19% of electrodes). To remove any possibility of noise artifact, we used only the best 6–7 units/recording session to perform our information analysis. The recordings in each session were treated as a unique data set. A preliminary recording stability analysis based on shape of waveforms, autocorrelation functions, and peri-event time histograms suggests that 50% of the electrodes record different units after a 1-mo period. Because five of the six recording sessions used in this work were collected more than 1 mo apart, it is likely that many of the units were unique.
Analysis.
Cross correlations. Cross-correlation histograms (CCHs) were computed by using either a 1 ms or 3 ms bin width and based on 17–52 trials per movement direction. One of the recording sessions, used to examine temporal modulation in synchrony for particular movement directions, required movements in only two directions (instead of eight directions) (see Fig. 2 A, C, and E) to obtain ∼150 trials per direction. For each CCH, one neuron in the pair was designated the reference cell and the other was referred to as the target cell. Cross-correlation values are reported either in number of counts per bin or in units of frequency. Frequency values should be read as the average number of action potentials in the target cell per action potential in the reference cell per unit time at a particular time lead/lag. Unless otherwise stated, a trial-based shift predictor is subtracted from the raw cross-correlation values shown in the figures. The shift predictor was obtained by recomputing the correlation after shifting the target cell’s spike train by one trial relative to that of the reference cell, by using the start of movement as the anchoring event. This procedure was repeated N times (with N approximately equal to the number of trials), each time shifting the target cell’s spike train by an additional trial. The resulting N correlations were averaged and designated as the trial-based shift predictor.
Figure 2.

Temporal modulation in synchrony over a trial. (A) A cross-correlogram between two single units. The x-axis represents time with respect to movement onset; the y-axis is lead/lag time in correlation; and color denotes the correlation strength (red is maximal and dark blue denotes nonsignificant values). Notice the transient increase in correlation around zero time lag (i.e., synchrony) at around movement onset. (B) Peaks in synchrony occur most often around the time of movement onset. This histogram (300 ms bin width) tabulates the times during the trial at which significant peaks in synchrony occurred with respect to movement onset (arrow). Significant peaks were defined as zero time lag correlation values (3 ms bin width) that crossed the upper bound of the 99% confidence limit. (C) Variations in synchrony are distinct from firing rate modulation. Sliding cross-correlation values (red line) at zero-time lag (i.e., synchrony) between two neurons are plotted along with their firing rates (black dotted lines) for movements to the left. The 99% confidence limit (dashed blue line) also is shown. Similar formats are used in D and E. (D) Different patterns of synchrony (red line) between one cell and two other simultaneously recorded neurons. Data from all 8 directions movement directions were pooled to compute cross correlations. (E) Pattern of synchrony across the trial varies with movement direction. Shown are synchrony values and firing rates between a given cell pair for leftward movements (Left side) and rightward movements (Right side). The red scale bar refers to synchrony magnitudes whereas the black bar refers to firing rates of the individual neurons. (C-E) Synchrony values are based on 3 ms bin widths and have been subtracted by the shift predictor. Vertical arrows indicate movement onset.
A 99% confidence limit was used to test for independence and assumed a Poisson counting process for the total number of coincident spikes within a bin presuming that both the reference and target cells fired independently (17). The mean rate of the process was estimated by multiplying the average firing rate of the target cell by the bin width of the CCH and multiplying this product by the number of spikes from the reference cell. This product is an estimate of the mean number of coincident spikes that would occur by chance if the two neurons fired independently. This product is very similar to the shift predictor estimate although they are calculated differently. A normal approximation to the Poisson distribution was used to set the confidence limit to 2.58 SDs above the mean.
“Sliding” CCHs were computed by using 100-ms intervals of data incremented in 20-ms steps. As before, synchrony was measured by taking the cross-correlation value at zero time lag and subtracting the corresponding value of the trial-based shift predictor. Movement onset was used as the anchoring event. The start of movement was defined as the time at which the position-feedback cursor left the hold position.
To estimate how synchrony varied with movement direction, we computed CCHs by using data within a 400-ms window centered on the start of movement for each of eight movement directions taken separately. The preferred direction (PD) of each neuron was defined as the peak of a cosine function fitted to the observed firing rate versus movement direction (5) during a 400-ms interval centered about movement onset.
Mutual information estimation.
We estimated the mutual information between the number of coincident spikes and movement direction in 50-ms nonoverlapping intervals so that we could assume stationarity in the firing rates of the neurons within each interval. Spike times were rounded to the nearest millisecond by binning. A coincidence was defined as at least one spike occurring in one neuron within ±0 (1 ms resolution), ±2 (5 ms resolution), or ±7 bins (15 ms resolution) of a spike in a second neuron. We assumed that the number of coincident spikes fit a Poisson distribution; this assumption was checked visually by comparing the observed frequency distributions to analytical distributions. This assumption allowed us to estimate the conditional distributions on the number of synchronous spikes given each movement direction by simply estimating the mean number of coincidences for each direction within the interval. We estimated the number of bits of mutual information, I, synchrony provided about direction by using the following equation:
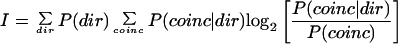 |
1 |
where dir is the movement direction and coinc is the number of coincident spikes in the interval for a given pair of neurons; P(dir) is the a priori probability distribution of movement directions which was set by the task presentation program to be uniform (i.e., all movement directions are equally likely); P(coinc|dir) is the probability of coinc coincident spikes given a particular movement direction; and P(coinc) is the a priori probability of coinc coincident spikes knowing nothing about the movement direction. Eq. 1 describes the average reduction in uncertainty (in bits) about the number of coincident spikes knowing in which direction the monkey moved. This is equal to the average reduction in uncertainty about the movement direction knowing how many coincident spikes occurred.
To test whether the observed mutual information between direction and number of coincidences was not merely a result of firing rate modulations, we computed the mutual information by randomly shuffling the trials of one neuron relative to the other. By computing multiple trial-shuffled estimates (30 shuffles), a distribution of values under the null hypothesis (i.e., that the neurons fire independently) was obtained. Mutual information estimates that were larger than 29 of 30 trial-shuffled estimates (i.e., P < 0.03) were considered significant. Those cell pairs that showed significant mutual information in at least one of ten 50-ms intervals around movement onset (300 ms before and 200 ms after movement onset) were tallied to come up with the percentages reported in Table 1. Given that ten 50-ms intervals were examined and a P value of 0.03 was used, we would expect that 26% (i.e., the probability of a cell pair having at least one interval of significant information is: 1–0.9710) of cell pairs would show significance by chance within that period. The assumption of independence among the intervals used to estimate this percentage is justified given that a different random shuffle was used for each interval. This percentage was used as the mean under the null hypothesis for a binomial test. A more accurate estimate of the distribution of values under the null hypothesis by using 100 trial-shuffled estimates instead of 30 for six cell pairs gave essentially the same results.
Table 1.
Percentage of cell pairs providing directional information from synchrony
| Monkey | 1 ms | 5 ms | 15 ms |
|---|---|---|---|
| 1 | 47% (30/64), P = 3 × 10−4 | 63% (40/64), P = 9 × 10−10 | 53% (34/64), P = 4 × 10−6 |
| 2 | 43% (18/42), P = 0.01 | 40% (17/42), P = 0.03 | 43% (18/42), P = 0.01 |
The Poisson model of coincidences also was tested by computing a non-parametric estimate of the mutual information for one data set. This was done by estimating the underlying distribution of coincidences by using ∼150 trials of data for each of two directions. The number of cell pairs with significant information did not substantially differ by using parametric or non-parametric methods.
RESULTS
The majority of cells recorded during reaching modulated their discharge in association with movement, usually beginning before movement onset, as commonly encountered in MI (Fig. 1A). Of 63 cells tested, 81% exhibited directional tuning which could be fit to a cosine function (r2 > 0.5; P < 0.05) (4, 5, 7, 18). CCHs revealed significant synchronous discharge in 35% of 114 neuronal pairs and 19% of 84 pairs tested for each of the two monkeys, respectively. The precision of synchrony was typically within the range of 10–15 ms where precision was defined as the width of the cross-correlation peak at half height (64% of 55 pairs) (Fig. 1B, left side), although examples of CCHs with only a sharper 1–3 ms peak at zero time lag were sometimes observed (36% of 55) (Fig. 1B, right side). Fig. 1C demonstrates that that each of the synchronous waveforms, which were characteristic of the waveforms recorded on each of the two channels, were similar, indicating that this precise form of synchrony results from the concerted firing of two isolated single units. As others have shown in motor cortex (11, 19), we found that the strength of temporal correlations generally decreased with inter-electrode distance (Fig. 1D). However, the incidence of synchrony at near distances were quite variable and synchrony could be detected at distances up to 3 mm apart in the MI arm area. Such long range interactions are consistent with the extensive horizontal connections and distributed nature of the MI arm area (20, 21).
Figure 1.

(A) Firing rate modulation of two simultaneously recorded MI neurons showing a movement related increase (Left side) and decrease (Right side). Peri-event time histograms are aligned on movement onset (dashed line) and are based on movements in a single direction. (B) Range of temporal precision of synchrony for pairs of recorded neurons. The relative positions of the recording sites on the array are shown as gray boxes on a grid. Below are shown CCHs between two pairs of neurons. A trial-based shift predictor representing the expected number of chance coincidences has been subtracted from the histograms. Broadly correlated discharge peaked at zero time lag (1 ms bin widths) (Left side), computed from data in the interval from trial start to the end of movement for all movement directions. Sharply correlated discharge at zero time lag (Right side). In this example, the data is restricted to a 700-ms interval after the go cue for a single movement direction. (C) All synchronous extracellular waveforms (±500 μs) recorded from two electrode sites that were used to compute the histogram on the Right side of B. These waveforms were recorded during a single recording session. Synchronous and nonsynchronous waveforms for both neurons were not different in initial peak amplitude (two-tailed t test, P < .01). Autocorrelation functions are shown below. (D) Synchronous interactions between neurons is evident at sites up to 3 mm apart. The strength of synchrony (normalized) shows a negative log-linear relationship with inter-electrode distance. Normalized synchrony values were computed by taking the ratios of the cross-correlation value at zero time lag (1 ms bin width) to the trial-based shift predictor value. Data were taken from a 700-ms period about movement onset. Error bars represent one SD.
The incidence of synchronous discharge varied across a behavioral trial. Importantly, the majority of significant peaks appeared in conjunction with movement initiation (Fig. 2A), although additional peaks were distributed throughout the trial (Fig. 2B). The dynamics of synchronous discharge could not be simply explained by firing rate modulations of the two cells. As illustrated in Fig. 2C peaks in synchrony were encountered during periods of low firing. Further, a given cell could participate in different forms of synchronous interactions with other cells. For example, one cell that exhibited a single phasic peak in synchrony with one cell (Fig. 2D Upper) revealed recurring episodes of synchrony with a different, simultaneously recorded cell (Fig. 2D Lower).
Not only did synchrony vary during the course of a trial, but it also varied by movement direction for a subset of recorded neuronal pairs. Note in the example shown in Fig. 2E, significant synchrony was observed around movement onset for rightward but not for leftward movements. Directional variations in synchrony were observed in 7%, 11%, and 19% of 144 neuron pairs reexamined in both animals by using the eight-direction task at 1-, 5-, and 15-ms temporal resolutions, respectively. A cell pair was classified as directional tuned if synchrony was significant in at least one direction. Cell pairs that were not directionally selective lacked significant synchrony in any of the eight directions. The directional selectivity of temporal correlations were not clearly related to the directional tuning obtained from the firing rate of either cell of the pair. The pair of cells shown in Fig. 3A engaged in synchronous (and mildly oscillatory) firing for movements to the right but not the left, even though both cells were not firing maximally during rightward movements. An example of particularly sharp synchrony distinct from firing rate tuning is shown in Fig. 3B. In this case, synchrony was only evident for movements in the 225 degree direction, whereas the individual neurons had preferred directions (i.e., peak direction in the cosine function) that were greater than 50 degrees from the peak synchrony. Fig. 3C summarizes the lack of a systematic relationship between the peak synchrony direction and the direction of maximal firing of the constituent neurons. In particular, there was no prevalence for the two directions to be the same. Maximal synchrony occurred in a variety of movement directions depending on the cell pair examined. Unlike the cosine-tuning of the individual cells based on firing rate, synchrony could possess additional submaximal peaks for different directions.
Figure 3.

Two examples of directional tuning in synchrony. (A) CCHs computed from two cells recorded during movements to the left and right during a 1-sec period around movement onset. Side bands (see arrows) in the CCH for movements to the right indicate oscillations. Below each CCH, the peri-movement time histograms are plotted for each of the two neurons. (B) CCHs between one pair of neurons computed over ±200-ms period with respect to movement onset for each movement direction. The 99% confidence limit (gray line) assumes independence of firing of the two cells. This pair of neurons exhibits strong synchrony for movements in the 225 degree direction. In contrast, the constituent cells have preferred directions on either side (170 and 293 degrees, gray and dark arrows, respectively). The shift predictor has not been subtracted in these histograms. (C) The number of directionally tuned cell pairs as a function of the difference between the direction of peak synchrony and the direction of the cell’s peak firing rate which is closest to the peak synchrony direction.
We quantified the amount of directional information provided by synchrony across the trial by estimating the mutual information (22) between the number of coincident spikes and movement direction. Mutual information is a particularly useful quantity because it provides an objective measure of the directional information content of synchrony that can be compared across time and across cell pairs having different physiological properties. This quantity also can be directly compared with the maximum possible information for the task (i.e., 3 bits for eight directions and 1 bit for two directions). Consistent with our cross-correlation results, we found that a large percentage of pairs examined revealed significantly more information regarding movement direction (P < 0.03) in their synchronous interactions than would be expected if the two cells fired independently of one another. Examples of the dynamics of this information is shown in Fig. 4. Significant information is seen to emerge around movement onset for one cell pair (Fig. 4A), whereas another cell pair exhibits significant information that persists for several hundred milliseconds after movement onset (Fig. 4B). The number of 50-ms intervals that showed significant information was significantly higher than would be expected given a P value of 0.03 by using a binomial test (P < 0.05): 7%, 8%, and 5% for 1, 5, and 15 ms bin widths computed from one data set. Moreover, for all three bin widths used in the analysis (1, 5, and 15 ms) in all data sets for two monkeys, a significant number of cell pairs exhibited directional information in their synchronous discharge around movement onset. Table 1 shows the percentage of these cell pairs for both monkeys as well as the significance levels by using a binomial test.
Figure 4.

Synchronous discharge carries directional information beyond that expected by firing rate alone. (A) The temporal evolution of mutual information between synchrony defined at 1 ms precision and movement direction (blue crosses and line) and trial-shuffled estimates of mutual information (red dots) for a single pair of neurons. The mutual information is significantly (P < 0.03) larger than chance at around movement onset (green dots). (B) In another cell pair, a sustained level of significant mutual information is observed after movement onset by using a 5 ms temporal precision. The means and SDs of the trial-shuffled estimates are shown in red. (Inset) The difference between the mutual information and the mean of the shuffled estimates. (C and D) The temporal evolution of mutual information for the same cell pair used in A by using a 5 ms bin width excluding coincident spikes at 1 ms precision and by using a 15 ms bin width excluding coincident spikes at 5 ms precision, respectively. (E) Z-scores of mutual information derived from the pair of neurons used in A, C, and D for all three levels of temporal precision: 1 ms (red), 5 ms exclusive of 1 ms synchrony (blue), and 15 ms exclusive of 5 ms synchrony (green), respectively. Scale bar represents a z-score of 4. Z-scores were computed by subtracting the mean from the estimates of mutual information and normalizing for variance. This measure reveals the additional information about direction that occurs at movement onset that exceeds chance. (F) For the same cell pair used in A, C, and D, all three levels of temporal precision together were used to estimate the mutual information between synchronous spikes and direction. Scale bar for all graphs except for E represents 0.05 bits/50 ms interval. All figures except for B are based on a two-direction task (i.e., 1-bit task). Figure B is based on an eight-direction task (i.e., 3-bit task).
Because our results at multiple time scales depend on each other to a certain degree (because broad synchrony includes fine synchrony), we examined information from synchrony at broad time scales while excluding coincident events at finer time scales. Mutual information was estimated from the same cell pair shown in Fig. 4A but by using a 5 ms bin width excluding synchronous spikes at 1 ms precision (Fig. 4C) and by using a 15 ms bin width excluding synchronous spikes at 5 ms (Fig. 4D). The information added by temporal synchrony is more clearly illustrated by plotting z-scores for these information estimates, which subtract the expected information provided by firing rate and normalize for variance differences (Fig. 4E). If all three temporal resolutions are included together in the estimation of mutual information, a strong degree of significant information is observed immediately after movement onset (Fig. 4F).
We compared the residual information provided by synchrony with the information provided by the firing rates of the constituent cells. Estimates of the information provided by rate alone were calculated by using data obtained from randomly selected, nonsimultaneously recorded trials (shuffling method) (see Methods). Residual information was considered to be the difference between the unshuffled (simultaneously recorded) estimate and the mean of the shuffled estimates. By using one data set in the two-direction task, we found that synchrony provided a maximum of 0.21 bits of residual information per coincidence event in a 50-ms interval as compared with a maximum of 0.33 bits per spike provided by a single cell. This result suggests that the additional directional information that can be obtained from synchrony when measured on a per event basis can be of the same magnitude as that available from single cell spike occurrence alone.
DISCUSSION
Our results provide the first demonstration that information about movement direction is available from the synchronous discharge of motor cortex neurons. Four features suggest that synchrony may constitute a distinct coding variable for directional information. First and foremost, the amount of information obtained from synchrony exceeded that predicted from the expected distribution assuming the neurons fired independently. Second, we observed that increased information could occur in more than one successive interval over the trial (as in Figs. 4 A, B, and F), which also suggests that it is not simply a chance occurrence. Third, significant synchrony clustered around a specific behavioral event, the onset of movement. Fourth, the dynamic modulation in synchrony did not necessarily parallel the firing rate modulations of the constituent cells. Taken together these findings argue that synchrony is not a reflection of changes in average discharge rate and that the directional information obtained from temporally correlated discharge provides is separate from that provided by firing rate modulations alone. Thus, synchrony may form an information channel distinct from firing rate.
Discharge synchrony has been previously noted throughout cerebral cortex, including motor cortex (23). Recent studies have indicated that dynamic synchronization between neurons can emerge in conjunction with more general aspects of motor behavior, such as the expectation of sensory cues signaling the initiation of movement (14) or the decision to move (13). By contrast we have been able to extract specific information about movement direction from synchrony. If synchrony were not directionally tuned, one might explain its occurrence as a reflection of some global modulation process. The dynamics of synchrony, i.e., its directional tuning and linkage to behavioral events, argues that the information carried is related to specific aspects of motor planning and action.
The physiological mechanisms underlying synchronous discharge are unknown. It seems unlikely that common input from a single cell could generate such precise firing given the weak and unreliable nature of synaptic interactions between cortical neurons (24–26). Common input from a synchronized group of cells from within MI or from other areas might underlie synchrony, but it is unclear how this hypothetical group of neurons would become synchronous. Nevertheless, it would be of interest to know whether input areas to MI, such as the premotor cortex or ventrolateral thalamus, contribute to MI synchrony. Recent work suggests that transient synchronous input to a cell may explain the irregular firing patterns observed in cortical neurons (27). Other mechanisms that rely upon network interactions among groups of cortical neurons have been proposed but remain to be experimentally verified (28, 29). Whatever the mechanism, our data suggest that directional information can be obtained from synchrony that is comparable on a per event basis to that obtained from the firing rates of individual neurons.
Acknowledgments
We would like to thank Daniel M. Stein for his technical assistance with spike sorting and Beth Travers for her assistance in running the electrophysiological experiments. We also would like to thank David Mumford and Stuart Geman for their help in developing the information-theoretic analysis and Jerome N. Sanes for his helpful comments on earlier drafts of this manuscript. Supported by National Institutes of Health grants awarded to N.G.H. and J.P.D.
ABBREVIATIONS
- MI
primary motor cortex
- CCH
cross-correlation histogram
Footnotes
A Commentary on this article begins on page 15166.
References
- 1.Fu Q-G, Flament D, Coltz J D, Ebner T J. J Neurophysiol. 1995;73:836–854. doi: 10.1152/jn.1995.73.2.836. [DOI] [PubMed] [Google Scholar]
- 2.Fu Q-G, Suarez J I, Ebner T J. J Neurophysiol. 1993;70:2097–2116. doi: 10.1152/jn.1993.70.5.2097. [DOI] [PubMed] [Google Scholar]
- 3.Thach W T. J Neurophysiol. 1978;41:654–676. doi: 10.1152/jn.1978.41.3.654. [DOI] [PubMed] [Google Scholar]
- 4.Evarts E V. In: Handbook of Physiology. Brooks V B, editor. Vol. 2. Washington, DC: Am. Physiol. Soc.; 1981. pp. 1083–1120. [Google Scholar]
- 5.Georgopoulos A P, Kalaska J F, Caminiti R, Massey J T. J Neurosci. 1982;2:1527–1537. doi: 10.1523/JNEUROSCI.02-11-01527.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ashe J, Georgopoulos A P. Cereb Cortex. 1994;6:590–600. doi: 10.1093/cercor/4.6.590. [DOI] [PubMed] [Google Scholar]
- 7.Georgopoulos A P, Schwartz A B, Kettner R E. Science. 1986;233:1416–1419. doi: 10.1126/science.3749885. [DOI] [PubMed] [Google Scholar]
- 8.Singer W, Gray C M. Annu Rev Neurosci. 1995;18:555–586. doi: 10.1146/annurev.ne.18.030195.003011. [DOI] [PubMed] [Google Scholar]
- 9.Roelfsema P R, Engel A K, Konig P, Singer W. J Cognit Neurosci. 1996;8:603–625. doi: 10.1162/jocn.1996.8.6.603. [DOI] [PubMed] [Google Scholar]
- 10.Allum J H J, Hepp-Reymond M C, Gysin R. Brain Res. 1982;231:325–334. doi: 10.1016/0006-8993(82)90369-9. [DOI] [PubMed] [Google Scholar]
- 11.Murphy J T, Kwan H C, Wong Y C. Can J Neurol Sci. 1985;12:11–23. doi: 10.1017/s0317167100046527. [DOI] [PubMed] [Google Scholar]
- 12.Murphy J T, Kwan H C, Wong Y C. Can J Neurol Sci. 1985;12:24–30. doi: 10.1017/s0317167100046539. [DOI] [PubMed] [Google Scholar]
- 13.Vaadia E, Haalman I, Abeles M, Bergman H, Prut Y, Slovin H, Aertsen A. Nature (London) 1995;373:515–518. doi: 10.1038/373515a0. [DOI] [PubMed] [Google Scholar]
- 14.Riehle A, Grun S, Diesmann M, Aertsen A. Science. 1997;278:1950–1953. doi: 10.1126/science.278.5345.1950. [DOI] [PubMed] [Google Scholar]
- 15.Campbell P K, Jones K E, Huber R J, Horch K W, Normann R A. IEEE Trans Biomed Eng. 1991;38:758–768. doi: 10.1109/10.83588. [DOI] [PubMed] [Google Scholar]
- 16.Nordhausen C T, Rousche P J, Normann R A. Brain Res. 1994;637:27–36. doi: 10.1016/0006-8993(94)91213-0. [DOI] [PubMed] [Google Scholar]
- 17.Abeles M. J Neurosci Methods. 1982;5:317–325. doi: 10.1016/0165-0270(82)90002-4. [DOI] [PubMed] [Google Scholar]
- 18.Donoghue J P, Sanes J N, Hatsopoulos N G, Gaal G. J Neurophysiol. 1998;77:159–173. doi: 10.1152/jn.1998.79.1.159. [DOI] [PubMed] [Google Scholar]
- 19.Kwan H C, Murphy J T, Wong Y C. Brain Res. 1987;400:259–269. doi: 10.1016/0006-8993(87)90625-1. [DOI] [PubMed] [Google Scholar]
- 20.Donoghue J P, Hess G, Sanes J N. In: Acquisition of Motor Behavior. Bloedel J, Ebner T, Wise S P, editors. Cambridge: MIT Press; 1996. pp. 363–386. [Google Scholar]
- 21.Huntley G W, Jones E G. J Neurophysiol. 1991;66:390–413. doi: 10.1152/jn.1991.66.2.390. [DOI] [PubMed] [Google Scholar]
- 22.Cover T, Thomas J. Elements of Information Theory. New York: Wiley; 1991. [Google Scholar]
- 23.Fetz E E, Toyama K, Smith W. In: Cerebral Cortex. Peters A, Jones E G, editors. New York: Plenum; 1991. pp. 1–80. [Google Scholar]
- 24.Bekkers J M, Stevens C F. Nature (London) 1990;346:724–729. doi: 10.1038/346724a0. [DOI] [PubMed] [Google Scholar]
- 25.Bekkers J M, Stevens C F. Nature (London) 1989;341:230–233. doi: 10.1038/341230a0. [DOI] [PubMed] [Google Scholar]
- 26.Freund T F, Martin K A C, Soltesz P, Somogyi P, Whitteridge D. J Comp Neurol. 1989;289:315–336. doi: 10.1002/cne.902890211. [DOI] [PubMed] [Google Scholar]
- 27.Stevens C, Zador A M. Nat Neurosci. 1998;1:210–217. doi: 10.1038/659. [DOI] [PubMed] [Google Scholar]
- 28.Hansel D, Sompolinsky H. J Comput Neurosci. 1996;3:7–34. doi: 10.1007/BF00158335. [DOI] [PubMed] [Google Scholar]
- 29.Juergens E, Eckhorn R. Biol Cybern. 1997;76:217–227. doi: 10.1007/s004220050334. [DOI] [PubMed] [Google Scholar]