

The cerebral cortex is the primary seat of our sensory experiences, our plans, and the control of our actions. As such, it has been carefully examined since the inception of brain science and has been studied intensively at the level of individual neurons for over 40 years (1, 2). Nonetheless, it appears a fair assessment to say that the single most essential question about cortical physiology—“What is the basic cortical population signalling mechanism?”—has not yet been definitively answered and is still the subject of vigorous debate (3–9). Does the cortex function by pooling together essentially independent neural signals, as in an election, or does the signal come through the coordination of the elements, as in a symphony (3, 10, 11)? An article by Hatsopoulos et al., appearing in this issue of the Proceedings (12), provides interesting new examples of pairs of neurons in the primary motor cortex of monkeys conveying information by the relative timing of their action potentials during an eight-choice reaching task. Forming clear distinctions between independent and coordinated coding is challenging, because cortical neurons have overlapping functions and work together by distributing signals across many elements. These authors present a straightforward, measurable distinction between information transmission in independent and coordinated coding and use their data to evaluate these two hypotheses.
The Independent Coding Hypothesis.
Independent coding suggests that each cortical neuron represents a separate signal. Cortical neurons in both sensory and motor systems are broadly tuned for features of the world, with a given sensory stimulus or motor act engaging very large numbers of cortical neurons (4, 6, 10, 13). Whatever the relevant size of cortical populations, the essential feature captured by the independent coding hypothesis is that all of the information that can possibly be obtained from one neuron can be obtained from that one neuron alone, without reference to the activities of any others. Independent signals may be noisy, overlapping, or redundant and may need to be averaged or pooled to become clear, but they are carried independently.
One well known sensory example is the activity of neurons in cortical area MT during the performance of motion discrimination tasks by monkeys. Individual neurons in this area have been shown to have selectivities for visual motion that parallel the perceptual performance of the animal. These data show that there can be sufficient information available to complete a particular discrimination task in the independent signals of single neurons (8, 14). However, this does not indicate whether there is more information available in the coordination of populations of these neurons, perhaps information necessary for more complex tasks or tasks requiring a more rapid solution.
A well studied example of independent population coding in the primary motor cortex is the population vector method for computing predicted movement direction from the activities of a large number of independently recorded motor neurons (13, 15–17). Each neuron is presumed to “vote” independently for its own preferred direction of movement or for its full direction tuning curve in more modern implementations (18). The population vector method represents an independent population code, not a coordinated code, as defined below. As in the previous example, these methods do not tell us whether additional information is available in the coordinated activity of these populations.
The Coordinated Coding Hypothesis.
The coordinated coding hypothesis suggests that messages in the cortex are conveyed at least in part by the relative timing of the action potentials of a population of neurons. To understand the signal from each neuron, the cortex has to listen to other neurons at the same time. The defining characteristic of coordinated coding is that the signal must be derived from the relations between multiple neurons in a population, whether this relation is synchrony or any other pattern; it is not derived by tallying the independent votes of the individuals.
There are now a number of cases in which cortical neurons have been shown to convey information by their relative timing when information is absent in their independent firing rate signals (9, 11, 19–22). For example, data have demonstrated that temporally coordinated action potentials can systematically signal sensory object features, even in the absence of changes in firing rate (19). Many auditory cortical neurons in anesthetized monkeys do not change their mean firing rates during the ongoing phase of continuous stimuli, so independent firing rate coding can be largely excluded for the neurons being studied in this special circumstance. Auditory cortical neurons can change their rate of synchronous spikes in this condition, and changes in synchrony are stimulus-frequency specific, are mapped systematically across the cortical surface, and follow the timecourse of ongoing stimuli. These data from a sensory system address very similar questions to those raised by Hatsopoulos et al. in the motor system—both explore systematic representation of features by cortical spikes synchronized on a timescale of 5–15 ms in the absence of parallel increases in firing rate. Special case examples in which firing rates are unchanged are particularly interesting because they can cleanly demonstrate coordinated signaling in the absence of independent firing rate signaling. It is presumed that the same coordinated signaling mechanisms might be used in more general circumstances, when firing rates are changed.
Related examples also have been found in the frontal and motor cortex (23–26). Vaadia et al. found that neurons in the frontal cortex of monkeys performing a motor task could show changes in their number of synchronous spikes in relation to behavior, even in the absence of changes in mean firing rate (27). There also have been examples shown of frontal cortical neurons emitting patterns of synchronous spikes in relation to behavior (11, 27).
Independent and coordinated signaling typically are found intermixed, and it is often not clear how to separate the two. For example, at stimulus or movement onsets, many neurons fire a burst of action potentials and thereby signal these events independently. However, the spikes are nearly coincident, so the relevant signal also could be the synchronous burst across the population. Neurons integrate their inputs temporally as well as spatially, so it is clear that, at the level of synapses, the number and timing of input spikes both contribute to postsynaptic response. Both codes are decoded by using the same set of synaptic mechanisms. Recent data have shown that, for cortical neurons, synchronization of input signals may play a particularly important role (28). Given the close relationship between population rate and timing, how can we ask most clearly whether cortical signaling primarily involves independent voting by individuals or coordinated signaling? A formalism based on information theory can address this issue.
An Introduction to Information Theory.
“Information theory” is a somewhat misleading name; it is really less of a theory than a simple type of statistics used for measuring arbitrary quantities, analogous to correlation coefficients and P values. Taking a familiar statistic as an example, the correlation coefficient expresses, roughly, the extent to which two parameters are related to one another. If a neural response is highly correlated with some feature of the world or behavior, then there will be a high correlation coefficient between the neural response and that feature: if one knows about one of them, one can make a good prediction about the other. “Mutual Information” is a central information theoretic statistic that is somewhat similar to a traditional correlation. In our example, if a neural response is highly correlated with some feature, then there also will be high mutual information between the response and that feature. If one knows the neuron’s response, then one has some information about what the feature likely was, and if one knows the feature, then one has some information about what the neuron’s response likely was. Mutual information also has some benefits beyond correlation: it can quantify noncontinuous relationships between a feature and a response, and it takes into account the full distributions of features and responses in question.
A full description of information theoretic statistics is, of course, far beyond this commentary and can be found elsewhere (29, 30). However, it is important to be clear that information theory is not a theory of coding in the nervous system that one can support or refute any more than a correlation coefficient is such a theory; it is just a set of useful statistical methods. In particular, showing high mutual information between a neural signal and a stimulus or movement merely shows that there is a strong and consistent relationship between the two. It does not show that the neural signal is important for coding, that it is the most relevant signal to measure, or that it is used by the nervous system at all, any more than a high correlation coefficient would show these things. Inferences about the importance of a signal depend on a theory of neural function.
Fig. 1 illustrates some cartoon examples of using information theoretic statistics. The black and white blocks in Fig. 1A indicate the presence or absence of some feature of the world (stimulus, movement, transistor in a computer, no matter). The lines below indicate the firing pattern of a particular neuron (or any other signaling element), with action potentials indicated by vertical lines. The stimulus has two possibilities, and these constitute a single either/or choice of white or black. The information needed to indicate the answer to a single either/or choice is defined to be one bit. In the upper case, the neuron’s activity is perfectly correlated with the feature, so each spike indicates the full answer to this choice and thereby conveys one bit of information. There is only one bit of information to be had about the feature (one either/or decision), so the neuron cannot possibly code any more than one bit per event. The neuron in the lower case is less well correlated, so its spike gives some information about the stimulus, but not enough to remove all uncertainty, and so conveys less than one bit.
Figure 1.

Information theoretic analysis of spike trains (see text).
Fig. 1B shows a slightly more complex example analogous to that explored by Hatsopoulos et al. Here, there are eight possible choices of direction, indicated by the arrows pointing to eight points around a circle. A single choice among eight can be divided into three choices among two (23 = 8), and a representation of the same choice as patterns of three independent black/white choices also is shown. In this case, a neuron can signal up to three bits per feature, and the neuron shown signals three bits per spike but .54 bits averaged across the eight features.
Mutual information corresponds to the change in uncertainty that one has about an element in a first signal, given a second signal. If the number of possibilities for the first signal is large, and many of the possibilities are quite likely, then there is a great deal of uncertainty. If the likely possibilities are few, then there is little uncertainty. If uncertainty changes from eight possibilities to one possibility on learning the second signal, as in Fig. 1B, then the uncertainty changes by log2(8) − log2(1) = 3 bits. Uncertainty is measured by the sum of the probabilities of each event times the log of the probability of that event, and this is called an entropy. From this, one can derive the definition of mutual information used by Hatsopoulos et al.,
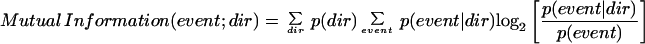 |
where p(dir) is the probability of a given movement direction, p(event dir) is the probability of observing a given number of events (e.g., spikes) in a bin during a particular movement direction, and p(event) is the probability of observing that number of events in a bin during any movement direction.
Separating Independent from Coordinated Coding.
The challenge is to clearly define the difference between information being carried independently by neurons of a population and information being carried by population coordination (31, 32) and then to explore whether the independent or coordinated model better characterizes cortical neurons during behavior. The paper by Hatsopoulos et al. is focused directly on this central issue. These authors succeed in recording from populations of neurons in the primary motor cortex of monkeys during a reaching task requiring the animals to move a manipulandum to a selected target. They also succeed in defining independent and synchrony coding and developing an estimate of the information content in each by using their data. This paper thereby challenges us to decide whether we see an answer to the long-term question of cortical population coding beginning to take shape in their findings.
The authors develop a method to compare the information in synchronous spikes with the information in shifted synchronous spikes. Shifted synchronous spikes are defined as spikes that are evoked from one neuron during one trial and a second neuron during a different trial, a long-standard method (33, 34). The question then becomes whether there is more mutual information between the synchronous spikes and the movement directions than there is in the independent signals of the two neurons, as measured by using the shifted synchronous spikes. Using the distinction between independent and coordinated coding suggested above, one could compare information content between independent coding and any type of coordinated coding, not just synchronous spikes.
The authors show example pairs that do exhibit more mutual information in their synchronous spikes than in the independent signals. A cartoon example of this is shown in Fig. 1C. The two individual neurons shown here have no mutual information with the depicted stimulus or movement feature. Each neuron is equally likely to fire during each feature. However, the synchronous spikes between the two neurons convey information about the stimulus—they are more likely to be found during black than during white features.
Information theoretic measures allow one to set a minimum to the amount of information potentially available in a signal, but not a maximum, because it is always possible that another decoding of the signal could convey even more information. Therefore, comparing estimates of the minimum information that can be found in two codes does not necessarily indicate which one actually carries more signal in the cortex.
Information theoretic analyses are also prone to significant sampling problems, in that it is common to overestimate the amount of information in a signal when insufficient data are present to make a correct estimation. This makes it crucial to verify information theoretic estimates for reliability by methods such as subsampling the data. Using different portions of the data set, one can verify that the same conclusion is reached and that using subsamples of increasing size produces convergence to the estimated value.
The conceptual approach taken by Hatsopoulos et al. leads in the important new direction of distinguishing the information content of independent and coordinated population codes in awake animals. Their data, however, raise many additional questions. It will be critical to address quantitatively the extent to which the illustrated examples of neurons exhibiting synchrony coding represent a statistically robust and general phenomenon in cortical coding. Future data sets containing more spikes, larger numbers of trials and larger numbers of neurons should allow more statistically powerful measurement of the phenomena they suggest and should provide an assessment of the magnitude and selectivity of added information carried by many pairs of neurons. For example, one wishes to know not just whether synchrony becomes statistically significant during a particular event, but whether it is significantly changed in different conditions, by how much it is changed for many pairs of neurons (19), and how much information this change carries on average. Ultimately, we will need new analytical methods to evaluate the coordinated information carried across whole populations of increasing size, rather than by pairs of neurons taken in isolation (11, 25, 35, 36). One hopes that the question of whether the fundamental cortical code acts by pooling independent signals or by coordinated populations ultimately will be resolved empirically in this way.
Footnotes
A commentary on this article begins on page 15706.
References
- 1.Mountcastle V B, Davies P W, Berman A L. J Neurophysiol. 1957;20:374–407. doi: 10.1152/jn.1957.20.4.374. [DOI] [PubMed] [Google Scholar]
- 2.Hubel D H, Wiesel T N. J Physiol. 1962;160:106–154. doi: 10.1113/jphysiol.1962.sp006837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Abeles M. Corticonics: Neural Circuits of the Cerebral Cortex. Cambridge, U.K.: Cambridge Univ. Press; 1991. [Google Scholar]
- 4.Andersen R A, Snyder L H, Bradley D C, Xing J. Annu Rev Neurosci. 1997;20:303–330. doi: 10.1146/annurev.neuro.20.1.303. [DOI] [PubMed] [Google Scholar]
- 5.Bair W, Koch C. Neural Comput. 1996;8:1185–1202. doi: 10.1162/neco.1996.8.6.1185. [DOI] [PubMed] [Google Scholar]
- 6.Deadwyler S A, Hampson R E. Annu Rev Neurosci. 1997;20:217–244. doi: 10.1146/annurev.neuro.20.1.217. [DOI] [PubMed] [Google Scholar]
- 7.Middlebrooks J C, Clock A E, Xu L, Green D M. Science. 1994;264:842–844. doi: 10.1126/science.8171339. [DOI] [PubMed] [Google Scholar]
- 8.Parker A J, Newsome W T. Annu Rev Neurosci. 1998;21:227–277. doi: 10.1146/annurev.neuro.21.1.227. [DOI] [PubMed] [Google Scholar]
- 9.Singer W, Gray C M. Annu Rev Neurosci. 1995;18:555–586. doi: 10.1146/annurev.ne.18.030195.003011. [DOI] [PubMed] [Google Scholar]
- 10.Arieli A, Sterkin A, Grinvald A, Aertsen A. Science. 1996;273:1868–1871. doi: 10.1126/science.273.5283.1868. [DOI] [PubMed] [Google Scholar]
- 11.Riehle A, Grun S, Diesmann M, Aertsen A. Science. 1997;278:1950–1953. doi: 10.1126/science.278.5345.1950. [DOI] [PubMed] [Google Scholar]
- 12.Hatsopoulos N G, Ojakangas C L, Paninski L, Donoghue J P. Proc Natl Acad Sci USA. 1998;95:15706–15711. doi: 10.1073/pnas.95.26.15706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Georgopoulos A P. Cold Spring Harbor Symp Quant Biol. 1990;55:849–859. doi: 10.1101/sqb.1990.055.01.080. [DOI] [PubMed] [Google Scholar]
- 14.Britten K H, Shadlen M N, Newsome W T, Movshon J A. J Neurosci. 1992;12:4745–4765. doi: 10.1523/JNEUROSCI.12-12-04745.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Georgopoulos A P. Annu Rev Neurosci. 1991;14:361–377. doi: 10.1146/annurev.ne.14.030191.002045. [DOI] [PubMed] [Google Scholar]
- 16.Georgopoulos A P, Schwartz A B, Kettner R E. Science. 1986;233:1416–1419. doi: 10.1126/science.3749885. [DOI] [PubMed] [Google Scholar]
- 17.Georgopoulos A P, Lurito J T, Petrides M, Schwartz A B, Massey J T. Science. 1989;243:234–236. doi: 10.1126/science.2911737. [DOI] [PubMed] [Google Scholar]
- 18.Sanger T D. J Neurophysiol. 1996;76:2790–2793. doi: 10.1152/jn.1996.76.4.2790. [DOI] [PubMed] [Google Scholar]
- 19.deCharms R C, Merzenich M M. Nature (London) 1996;381:610–613. doi: 10.1038/381610a0. [DOI] [PubMed] [Google Scholar]
- 20.Gray C M, Singer W. Proc Natl Acad Sci USA. 1989;86:1698–1702. doi: 10.1073/pnas.86.5.1698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gray C M, Konig P, Engel A K, Singer W. Nature (London) 1989;338:334–337. doi: 10.1038/338334a0. [DOI] [PubMed] [Google Scholar]
- 22.von der Malsburg C. Internal Report. Göttingen, Germany: Max Planck Institute for Biophysical Chemistry; 1981. [Google Scholar]
- 23.Abeles M, Bergman H, Margalit E, Vaadia E. J Neurophysiol. 1993;70:1629–1638. doi: 10.1152/jn.1993.70.4.1629. [DOI] [PubMed] [Google Scholar]
- 24.Prut Y, Vaadia E, Bergman H, Haalman I, Slovin H, Abeles M. J Neurophysiol. 1998;79:2857–2874. doi: 10.1152/jn.1998.79.6.2857. [DOI] [PubMed] [Google Scholar]
- 25.Abeles M, Bergman H, Gat I, Meilijson I, Seidemann E, Tishby N, Vaadia E. Proc Natl Acad Sci USA. 1995;92:8616–8620. doi: 10.1073/pnas.92.19.8616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Murthy V N, Fetz E E. Proc Natl Acad Sci USA. 1992;89:5670–5674. doi: 10.1073/pnas.89.12.5670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Vaadia E, Haalman I, Abeles M, Bergman H, Prut Y, Slovin H, Aertsen A. Nature (London) 1995;373:515–518. doi: 10.1038/373515a0. [DOI] [PubMed] [Google Scholar]
- 28.Stevens C F, Zador A M. Nature Neuroscience. 1998;1:210–217. doi: 10.1038/659. [DOI] [PubMed] [Google Scholar]
- 29.Shannon C E. The Mathematical Theory of Communication. Urbana, IL: Univ. of Illinois Press; 1949. [Google Scholar]
- 30.Rieke F, Warland D, de Ruyter van Steveninck R R, Bialek W. Spikes: Exploring the Neural Code. Cambridge, MA: MIT Press; 1997. [Google Scholar]
- 31.Dan Y, Alonso J M, Usrey W M, Reid R C. Nature Neuroscience. 1998;1:501–507. doi: 10.1038/2217. [DOI] [PubMed] [Google Scholar]
- 32.Warland D K, Reinagel P, Meister M. J Neurophysiol. 1997;78:2336–2350. doi: 10.1152/jn.1997.78.5.2336. [DOI] [PubMed] [Google Scholar]
- 33.Aertsen A M, Gerstein G L, Habib M K, Palm G. J Neurophysiol. 1989;61:900–917. doi: 10.1152/jn.1989.61.5.900. [DOI] [PubMed] [Google Scholar]
- 34.Perkel D H, Gerstein G L, Moore G P. Biophys J. 1967;7:419–440. doi: 10.1016/S0006-3495(67)86597-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.deSa V R, deCharms R C, Merzenich M M. Neural Information Processing Systems. 1998;10:131–137. [Google Scholar]
- 36.Seidemann E, Meilijson I, Abeles M, Bergman H, Vaadia E. J Neurosci. 1996;16:752–768. doi: 10.1523/JNEUROSCI.16-02-00752.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]