

Abstract
Over the past decade, important roles for the 84–88 kDa Group VIA Ca2+-independent phospholipase A2 (iPLA2β) in various organs have been described. We demonstrated that iPLA2β participates in insulin secretion, insulinoma cells and native pancreatic islets express full-length and truncated isoforms of iPLA2β, and certain stimuli promote perinuclear localization of iPLA2β. To gain a better understanding of its mobilization, iPLA2β was expressed in INS-1 cells as a fusion protein with EGFP, enabling detection of subcellular localization of iPLA2β by monitoring EGFP fluorescence. Cells stably-transfected with fusion protein expressed nearly 5-fold higher catalytic iPLA2β activity than control cells transfected with EGFP cDNA alone, indicating that co-expression of EGFP does not interfere with manifestation of iPLA2β activity. Dual fluorescence monitoring of EGFP and organelle Trackers combined with immunoblotting analyses revealed expression of truncated iPLA2β isoforms in separate subcellular organelles. Exposure to secretagogues and induction of ER stress are known to activate iPLA2β in β-cells and we find here that these stimuli promote differential localization of iPLA2β in subcellular organelles. Further, mass spectrometric analyses identified iPLA2β variants from which N-terminal residues were removed. Collectively, these findings provide evidence for endogenous proteolytic processing of iPLA2β and redistribution of iPLA2β variants in subcellular compartments. It might be proposed that in vivo processing of iPLA2β facilitates its participation in multiple biological processes.
Keywords: Golgi, ER, mitochondria, fusion protein, truncation, mass spectrometry
1. Introduction
Phospholipases A2 (PLA2)1 are a diverse group of enzymes that catalyze hydrolysis of the sn-2 substituent from glycerophospholipid substrates to yield a free fatty acid and a 2-lysophospholipid [1–3]. Among the PLA2s is a cytosolic PLA2 that does not require Ca2+ for catalysis and is designated Group VIA PLA2 or iPLA2β [4, 5]. At present two variants of iPLA2β are recognized and they are designated Group VIA-1 (84 kDa) and Group VIA-2 (88 kDa). The iPLA2β enzyme is expressed in various organs, stimulated by ATP, and inactivated by a bromoenol lactone (BEL) suicide substrate inhibitor of iPLA2β [6].
The iPLA2β variants contain 7–8 NH2-terminal ankyrin repeats, a caspase-3 cleavage site, an ATP-binding domain, a serine lipase consensus sequence (GXSXG), a bipartite nuclear localization sequence, a COOH-terminal calmodulin-binding domain(s) [6], and an ability to form a signaling complex with CamKIIβ [7]. Though regulation of iPLA2β during stimulation is not well-understood, the iPLA2β gene contains a sterol regulatory element (SRE) and binding of SRE binding proteins (SREBPs) to the SRE has been reported to promote iPLA2β transcription [8]. The Group VIA-2 iPLA2β, which is also expressed in human pancreatic islets, is the product of an mRNA species that arises from alternate splicing and contains a 54-amino acid sequence that interrupts the eighth ankyrin repeat [9].
The iPLA2β enzyme is proposed to be involved in phospholipid remodeling [10], signal transduction [11–16], sperm motility and fertility [17], bone formation [18], and fatty acid oxidation in skeletal muscle [19]. Recently, a role for iPLA2β in modulation of neurotransmission in the hippocampus [20], impairment in memory acquisition [21], nerve degeneration [22, 23], infantile neuroaxonal dystrophy [24, 25], schizophrenia [26, 27], muscle degeneration [28], skeletal muscle contractility [29], HIV-induced cardiomyopathy [30], exfoliation glaucoma [31], and onset of acute pleurisy [32] has also been recognized. Further, over 30 studies during the past five years have linked iPLA2β with apoptosis.
Studies in our laboratory reveal that pancreatic islet β-cells, but not non-βcells, express the Group VA-1 iPLA2β that participates in insulin secretion [33–36], cell proliferation [4], and apoptosis [37–39]. In support, inhibition or knockdown of iPLA2β suppresses [33, 34, 36, 37, 40, 41] and overexpression of iPLA2β amplifies these effects [37, 42, 43]. However, these protocols do not affect β-cell membrane phospholipid remodeling [4, 40, 42, 44], indicating a signaling role for iPLA2β in the β-cell.
Curiously, β-cells express different isoforms of iPLA2β protein that are not products of alternate splicing of the iPLA2β transcript and that are catalytic activity. These include a 70 kDa protein that results from truncation of C-terminal amino acids [34] and a 63 kDa protein that is generated by caspase-3-catalyzed cleavage of N-terminal amino acids [45]. Immunocytochemistry analyses suggested that in the presence of certain stimuli, the Group VA-1 iPLA2β localizes in the perinuclear region [45]. Subsequent expression of the Group VA-1 iPLA2β in INS-1 cells as a fusion protein (fp) with EGFP revealed a predominant localization of iPLA2β in the ER and Golgi [46].
In view of the presence of isoforms that result from truncation at either terminus of iPLA2β, herein, we compared the localization of the Group VA-1 iPLA2β expressed as a fusion protein. To facilitate identification of specific subcellular organelle(s) with which iPLA2β might associate during such stimulation, we engineered INS-1 cells that express iPLA2β as a fusion protein with EGFP fused to iPLA2β either at its C-terminus (NiPLA2β-EGFPC, designated fpN2) or at its N-terminus (NEGFP-iPLA2βC, designated fpC2). Expression of iPLA2β as a fusion protein permits tracking subcellular localization of iPLA2β by monitoring EGFP green fluorescence.
2. Materials and methods
2.1. Materials
INS-1 β-cells were generously provided by Dr. C. Newgard (Duke University Medical Center, Durham, NC). Other materials were obtained from the following sources: rainbow molecular mass standards and enhanced chemiluminescence (ECL) reagent (Amersham, Arlington Heights, IL); sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) supplies (BioRad, Hercules, CA); pEGFP-N2 and pEGFP-C2 vectors (Clontech, Palo Alto, CA); GenePORTER™ transfection reagent (Gene Therapy Systems Inc., San Diego, CA); pentex fraction V fatty acid-free bovine serum albumin (Miles Laboratories, Eckert, IN); Golgi Tracker BODIPY® TR ceramide, ER-Tracker™ Blue-White DPX (E-12353), Mito Tracker Deep Red 633, plasma membrane Tracker DiI, and Slow Fade® light antifade kit (Molecular Probes, Eugene, OR); (16:0/[14C]-18:2)-GPC (PLPC, 55 mCi/mmol), Coomassie reagent (Pierce, Rockford, IL); modified trypsin (Promega Corp., Madison, WI); peroxidase-conjugated goat anti-rabbit IgG antibody (Roche Diagnostic Corporation, Indianapolis, IN); primary antibodies against iPLA2β and GFP, GFP (B-2) mouse monoclonal IgG2α,, and goat anti-mouse IgG-HRP (Santa Cruz Biotechnology, Inc., Santa Cruz, CA); albumin, β-mercaptoethanol, protease inhibitor cocktail (PIC), and common reagents and salts (Sigma Chemical Co., St. Louis, MO); and antibiotic solutions and cell culture media (Tissue Culture Support Center, Wash. Univ., St. Louis, MO).
2.2. Preparation and expression of iPLA2β as a fusion protein with EGFP in INS-1 cells
Vectors pEGFP-N2 and pEGFP-C2 encode a red-shifted variant of wild-type GFP [47] that have been optimized for brighter fluorescence and higher expression in mammalian cells. INS-1 cells overexpressing a fused protein were generated as described [46]. Briefly, cDNA encoding the full-length Group VIA-1 iPLA2β (NP_058611) was amplified by PCR using the following primer set: sense, 5’-AGCTTCGAAT TCATGCAGTTCTTTGGACGC-3’, and antisense, 5’-TTCGATATCGGGAGATAGCAGCAGCTGG-3’. The amplified full-length iPLA2β from the pMSCV-neo- iPLA2β constructs were then subcloned into the pEGFP-N2 and pEGFP-C2 (Clontech, Palo Alto, CA). Control vectors (pEGFP-N2 and pEGFP-C2), and constructs encoding fusion protein (fp) were transfected into INS-1 cells with a Gene PORTER transfection system, according to the manufacture’s instructions (Gene Therapy Systems, San Diego, CA). Stably-transfected clones were selected using G418 (0.4 mg/ml) and fluorescence-activated cell sorting was utilized to optimize selection of stably-transfected cells with the highest iPLA2β expression. The cells were expanded in RPMI 1640 medium containing 11 mM glucose, 10% fetal calf serum, 10 mM HEPES buffer, 2 mM glutamine, 1 mM sodium pyruvate, 50 mM β-mercaptoethanol, and 0.1% (w/v) each of penicillin, fungizone, and streptomycin [4, 45]. Cells expressing EGFP-N2 and EGFP-C2 alone are designated N2 and C2, respectively, and were used as controls. Cells expressing iPLA2β as a fusion protein with EGFP were designated fpN2 (EGFP fused at the C-terminus) and fpC2 (EGFP fused at the N-terminus).
2.3. Preparation of INS-1 cell subcellular fractions for iPLA2β enzyme activity assays
INS-1 cell cytosol and membrane fractions were prepared and specific Ca2+-independent PLA2β activity in 25 µg protein aliquots was assayed in the presence of (16:0/[14C]-18:2)-GPC substrate and quantitated, as described [48]. To verify that the measured activity reflected that of iPLA2β, the abilities of ATP (10 mM) to stimulate and of BEL (10 µM) to inhibit activity were examined. To test the effects of BEL on activity, the sample protein was first pre-incubated (2 min, RT) with BEL before addition of substrate.
2.4. Secretagogue-stimulated translocation of iPLA2β
To examine if secretagogues induce translocation of iPLA2β, control and fusion protein expressing INS-1 cells were stimulated with glucose (G, 0 or 2 mM) in the absence and presence of forskolin (FSK, 2.5 µM). At 2 h, localization of iPLA2β, as reflected by EGFP, was visualized by fluorescence microscopy using a FITC filter.
2.5. Secretagogue-stimulated subcellular localization of iPLA2β
To identify organelles with which iPLA2β associates, fluorescences of EGFP and of specific organelle Trackers were monitored in live control and fusion protein-expressing cells after secretagogue stimulation using an inverted microscope (Nikon Eclipse TE300). The fluorescence images were captured using Soft Imaging System software (Lakewood, CO) and merged using Adobe Photoshop Software. Protocols with the different Trackers were as follows:
Golgi
Cells were incubated (4 °C, 30 min) with 5 µM ceramide-BSA (BODIPY® TR ceramide), washed, and incubated (37 °C, 1 h) in cu lture medium containing zero glucose and 0.1% BSA. The cells were then incubated (37 °C) in med ium supplemented with 2 mM glucose and 2.5 µM forskolin for up to 2 h. At various times during stimulation, EGFP and the Golgi Tracker (rhodamine) fluorescences were monitored.
ER, mitochondria, and plasma membrane
Cells were initially washed and incubated (37 °C, 1 h) in medium containing zero glucose and 0.1% BSA. The medium was then replaced with one supplemented with 2 mM glucose and 2.5 µM forskolin for up to 2 h. The ER (Blue-White DPX 600 nM), mitochondrial (Deep Red 633), or plasma membrane (Dil D-282) Tracker was added to the medium during the final 30 min of incubation. At 2 h, the medium was replaced with one not containing a Tracker and iPLA2β -associated EGFP fluorescence was monitored along with fluorescence associated with the ER (DAPI filter), mitochondria (rhodamine filter), or plasma membrane (rhodamine filter) Tracker.
2.6. ER stress-induced subcellular distribution of iPLA2β
Our earlier work indicated that inducing ER stress promotes activation of iPLA2β in the ER and mitochondria, leading to the release of cytochrome c and promoting apoptosis [38, 39, 49]. To determine if this process involves translocation of iPLA2β into the ER and/or mitochondria, a retroviral construct encoding a recombinant fusion protein consisting of EGFP attached to the N-terminus of the iPLA2β sequence (fpC2) was prepared and used to generate stably-transfected INS-1 cells as previously described [46]. Fusion proteins containing fluorescent tags attached to marker proteins for subcellular organelles were expressed for co-localization studies. The plasmid expressed to track ER encoded a fusion protein of the ER-targeting sequence of calreticulin attached to the N-terminus of MonoRed (Clontech, Mountain View, CA). The plasmid expressed to track mitochondria encoded a fusion protein of the mitochondrial-targeting sequence of human cytochrome c oxidase subunit VIII attached to Discosoma sp. red fluorescent protein (DsRed2). To compare the subcellular distribution of the tagged iPLA2β to that of the organelle markers, fpC2-expressing cells were transfected with either the pMonoRed-ER or the pDsRed2-Mito plasmid. To examine subcellular redistribution of the tagged iPLA2β upon induction of ER stress, cells were incubated with thapsigargin (1 µM, 37 °C) for various intervals. Cells were then fixed and visualized and analysed, as described above.
Preparation of mitochondria and ER fractions
To identify the iPLA2β isoforms that accumulate in the mitochondria and ER. Cells expressing iPLA2β with EGFP fused at the C-terminus (fpN2) were treated with vehicle (DMSO) alone or thapsigargin and harvested at 5 and 8 h. Mitochondria and ER fractions were then prepared and aliquots (30 µg protein) were processed for immunoblotting analyses, as described [38, 39].
2.7. Immunoblotting analyses
INS-1 cellular protein aliquots (30 µg) were analyzed by SDS-PAGE (7.5%), and processed for immunoblotting analyses as described [38, 39]. Immunoreactive protein bands were visualized by enhanced chemiluminescence (ECL). The primary antibodies were directed against GFP (IgG2α, 0.0002 µg/µl) or iPLA2β (0.0015 µg/µl). The secondary antibodies were goat anti-mouse IgG-HRP (1:2,000) for GFP and peroxidase-conjugated goat anti-rabbit IgG (1:40,000) for iPLA2β.
2.8. Mass spectrometric analyses of iPLA2β
To determine whether iPLA2β undergoes endogenous proteolytic processing, iPLA2β overexpressed in INS-1 cells was purified, concentrated, digested with trypsin, and analysed by MS, as described [34]. Briefly, iPLA2® was purified using ATP affinity chromatography. Fractions containing iPLA2® activity were pooled and concentrated. The concentrate was then resolved by SDS-PAGE. Protein bands were visualized by Coomassie stain and the band in the region of 85 kDa, corresponding to the molecular mass of iPLA2® was excised, reduced with 20 mM DTT and incubated with 50 mM iodoacetamide to alkylate the cysteine residue. Mass spectrometry grade trypsin (50:1) was then added and the gel pieces were incubated for 6 h at 37 °C. The digest was then centrifuged and the gel extract was injected into CapLC-Qtof (Waters) for data-dependent MS/MS analyses. Masslynx software was used to extract the peak list file from the MS/MS spectra. The peak list was then compared against a list of expected and unexpected peptides generated by a Signature Discovery Algorithm program recently developed in our laboratory [50]. These analyses facilitate identification of “signature” peptides derived from protein isoforms that reflect proteolytic processing events.
3. Results
3.1. Expression of iPLA2β enzymatic activity in INS-1 cells transfected with EGFP- iPLA2β cDNA
To confirm that the fpN2- and fpC2- transfected cells express a catalytically-active iPLA2β enzyme, cytosol and membrane fractions prepared from these cells were assayed for Ca2+-independent PLA2 activity. As shown in Fig. 1, both fpN2- and fpC2-expressing cells express a Ca2+-independent PLA2 activity that is four-to-five fold higher than that expressed in N2 and C2 cells. Such activity is stimulated by ATP and inhibited by BEL, which are recognized properties of iPLA2β [34, 36, 44, 45, 51]. These findings indicate preservation of iPLA2β activity when it is expressed as a fusion protein with EGFP.
Fig. 1. iPLA2β enzymatic activity in stably-transfected INS-1 cells.

Specific iPLA2β enzymatic activity in 25 µg protein aliquots of cytosol or membranes, prepared from transfected INS-1 cells, was assayed and quantitated as described in Methods. The data are presented as mean ± SEM (n = 8–10). A: Cytosolic activity. B: Membrane-associated activity.
3.2. Secretagogue-stimulated localization of iPLA2β
We previously observed using immunofluorescence analyses that exposure to secretagogues promotes translocation of iPLA2β to the perinuclear region of INS-1 cells [45]. Because potential antibody nonspecificity can lead to misinterpretation of immunofluorescence experiments, iPLA2β was expressed in INS-1 cells as a fusion protein (fp) with EGFP, and its subcellular redistribution after secretagogue stimulation was monitored by fluorescence microscopy. Our findings with the N2 construct, reported earlier [45], revealed that under non-stimulating conditions the green fluorescence in cells expressing N2 and fpN2 is dispersed throughout the cell. Following addition of glucose (2 mM) plus forskolin (2.5 µM), which was sufficient to increase insulin secretion 2-fold (data not shown), the fluorescence remains dispersed in the N2 expressing cells but in the fpN2 expressing cells, punctuated iPLA2β-associated green fluorescence is seen as a halo in the perinuclear region. Analogous analyses in the present study in cells expressing C2 and fpC2 (Fig. 2) reveal a similar dispersion of fluorescence under non-stimulatory conditions. Upon stimulation, in contrast to what was observed in the fpN2 cells, a punctate fluorescence that is dispersed in the cytosol is seen.
Fig. 2. Secretagogue-stimulated localization of fusion protein in INS-1 cells.

INS-1 cells expressing fpC2 (EGFP-iPLA2β) were incubated with glucose (G, 0 or 2 mM) in the absence and presence of forskolin (FSK, 2.5 µM). At 2 h, iPLA2β protein localization was monitored by fluorescence microscopy.
3.3. Secretagogue-stimulated subcellular redistribution of iPLA2β
To determine whether secretagogue stimulation of INS-1 cells promotes association of iPLA2β with specific subcellular compartments, dual fluorescence analyses using organelle Trackers were next performed. Targeted organelles were the Golgi, ER, mitochondria, and plasma membrane. The cells were treated with glucose and forskolin, and, at appropriate times, an individual organelle Tracker was added to the medium. The EGFP (FITC) and organelle Tracker fluorescences were then recorded in a field of cells. In Fig. 3, the separate iPLA2β-associated EGFP and organelle Tracker fluorescences are shown in the left and middle panels, respectively, and the overlays of the two fluorescences are shown in the right panels. Captured images were processed to reduce background fluorescence.
Fig. 3. Secretagogue-stimulated subcellular distribution of fusion protein.
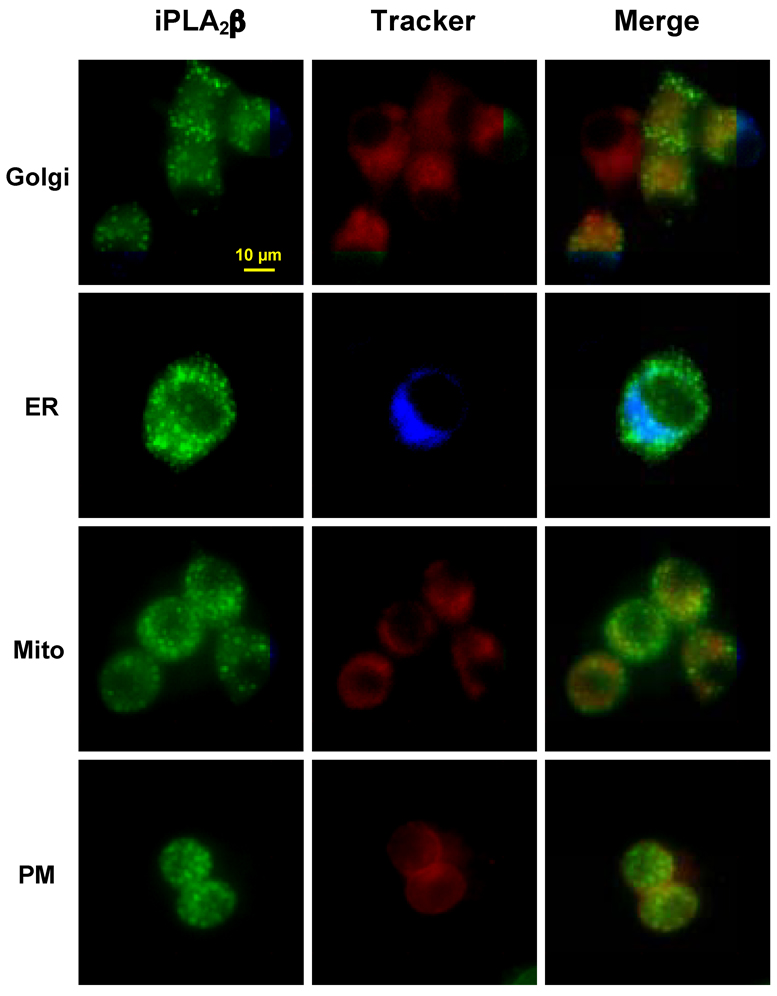
INS-1 cells expressing fpC2 (EGFP-iPLA2β) were incubated with glucose (G, 2 mM) and forskolin (FSK, 2.5 µM) along with organelle markers for Golgi (5 µM) or ER (600 nM), as described in Methods. At 2 h following stimulation, the EGFP (FITC) fluorescence was monitored separately from fluorescence associated with organelle Tracker; Golgi (rhodamine), ER (DAPI), mitochondria (rhodamine) and plasma membrane (PM, rhodamine). Left Panels, EGFP-associated iPLA2β fluorescence alone; Middle Panels, organelle Tracker fluorescence alone; and Right Panels, overlay of EGFP and organelle Tracker fluorescences.
In our earlier study, we found that the green fluorescence associated with iPLA2β in fpN2-expressing localized in the Golgi and ER, minimally associated with the plasma membrane, and not at all present in the mitochondria [45]. In contrast, as illustrated in Figure 3, the iPLA2β fluorescences in fpC2 expressing cells did not appear to overlap with any of the organelle trackers. In view of the findings with fpN2, the present observations with fpC2 raise the possibility that the presence of EGFP at one of the terminus of iPLA2β or possible proteolytic cleavage of the iPLA2β may contribute to distinct cellular redistribution of iPLA2β in secretagogue-stimulated β-cells.
3.4. Immunoblotting analyses provides evidence for endogenous processing of iPLA2β
To examine the possibility that iPLA2β undergoes endogenous proteolytic processing, cytosol prepared from fp-expressing cells was processed for immunoblotting analyses using antibodies against EGFP and iPLA2β. As illustrated in Fig. 4A, cells transfected with the EGFP-alone vector (N2 and C2) express only the EGFP-immunoreactive bands ca. 26 kDa (N2) and 29 kDa (C2), where the difference in mass is due to differences in the length of the insert encoded by the two vectors (left panel). The expected size of the fp, comprising EGFP and full-length iPLA2β (84 kDa) therefore is 110–113 kDa. However, immunoblotting analyses of cytosol from INS-1 cells expressing NiPLA2β-EGFPC (fpN2) and NEGFP-iPLA2βC (fpC2) revealed additional EGFP-immunoreactive bands that migrated with apparent molecular masses of ca. 90 kDa (fpN2) and 100 kDa (fpC2) (Fig. 4A, right panel). These bands appear to represent iPLA2β variants, as they are also recognized by polyclonal antibodies against iPLA2β (Fig. 4B). As seen in panel 4B, an additional iPLA2β-immunoreactive band is evident at 70 kDa.
Fig. 4. EGFP and iPLA2β immunoblotting analyses in INS-1 cells.

Following transfection of INS-1 cells with constructs encoding EGFP ± iPLA2β, cytosol protein fractions were prepared and analysed by SDS-PAGE and transferred onto Immobolin-P PVDF membrane. The electroblots were probed with antibodies directed against EGFP or iPLA2β and immunoreactive protein bands were visualized by ECL. A: EGFP immunoblotting. B: iPLA2β immunoblotting. C: Proposed scheme for generation and visualization of iPLA2β variants. (N2, EGFP-N2 vector only; C2, EGFP-C2 vector only; fpN2, iPLA2β-EGFP; fpC2, and EGFP- iPLA2β.)
A potential rationale for the observed immunoreactive bands is presented in Fig. 4C. The 90 kDa EGP-immunoreactive band evident only in fpN2-expressing cells most likely represents a variant of iPLA2β from which amino acid residues have been removed from the N-terminal region. As the EGFP is fused to the C-terminus of fpN2, this truncated isoform retains EGFP, is recognized by the EGFP antibody, and would exhibit fluorescence. In contrast, similar cleavage of fpC2 would result in the elimination of the EGFP that is fused to the iPLA2β N-terminus. The product of this cleavage that retained the majority of the iPLA2β sequence would thus fail to be recognized by EGFP antibody and to exhibit fluorescence, although it would react with iPLA2β antibody. The 100 kDa EGFP-immunoreactive band that is evident only in the fpC2-expressing cells probably represents a variant of iPLA2β from which 10–12 kDa of amino acid sequence has been removed from the C-terminal. As the EGFP in fpC2 is fused to the N-terminus of iPLA2β, this truncated isoform is recognized by the EGFP antibody. In contrast, similar cleavage of fpN2 would result in the elimination of EGFP fused to the C-terminus of iPLA2β, and this variant would not be recognized by the EGFP antibody.
Because the control cells (N2 and C2) are transfected with only the EGFP vectors and hence do not express a fusion protein, the iPLA2β-immunoreactive bands seen in these cells (Fig. 4C) most likely represent endogenous full-length iPLA2β produced from the transcripts of the native gene and a 70 kDa iPLA2β variant produced by removal of about 14 kDa of C-terminal sequence [52].
Consistent with the fluorescence analyses, the plasma membrane fraction contains an immunoreactive band (ca. 110–113 kDa) that is recognized by both EGFP and iPLA2β antibodies. This suggests that only the full-length iPLA2β protein associates with plasma membranes (Fig. 5).
Fig. 5. Immunoblotting analyses of plasma membrane-associated iPLA2β.

Plasma membrane protein fractions were prepared and analysed by SDS-PAGE and transferred onto Immobolin-P PVDF membrane. The electroblots were probed with antibodies directed against EGFP or iPLA2β and the immunoreactive protein bands were visualized by ECL. A: EGFP immunoblotting. B: iPLA2β immunoblotting.
3.5. ER stress-induced redistribution of iPLA2β between the ER and mitochondria
The findings that ER- and mitochondrial-associated iPLA2β activities are increased following induction of ER stress with thapsigargin in INS-1 cells suggest that an interaction between iPLA2β and the ER and mitochondria occurs during the evolution of apoptosis. To examine this further, INS-1 cells that express fpC2 and ER (calreticulin) or mitochondria (cytochrome c oxidase subunit VII) markers tagged to fluorophores were generated and visualized by fluorescence microscopy following vehicle or thapsigargin treatment. Individual fluorescence associated with iPLA2β-tagged with EGFP (fpC2, left panels) and organelle markers (middle panels), and the merged fluorescences (right panels) are presented in Fig. 6 (ER) and Fig. 7 (mitochondria).
Fig. 6. iPLA2β association with the ER following induction of ER stress in INS-1 cells.

INS-1 cells expressing fpC2 (EGFP-iPLA2β) alone or fpC2 along with a fusion protein of ER-targeting sequence of calreticulin tagged to the N-terminus of MonoRed were treated with DMSO vehicle (Con) or with thapsigargin (0.5 µM) to induce ER stress. At 8 and 24 h the cells were harvested, fixed, and assessed by fluorescence microscopy. Images of individual fluorescence associated with iPLA2β (left panels) and ER marker (middle panels) and the merged fluorescence (right panels) are presented.
Fig. 7. ER stress-induced accumulation of iPLA2β in INS-1 cell mitochondria.

INS-1 cells expressing fpC2 (EGFP-iPLA2β) alone or fpC2 along with a fusion protein of mitochondrial-targeting sequence of human cytochrome c oxidase subunit VIII tagged to Discosoma sp. red fluorescent protein were treated with DMSO vehicle (Con) or with thapsigargin (0.5 µM) to induce ER stress. At 8 and 24 h the cells were harvested, fixed, and assessed by fluorescence microscopy. Images of individual fluorescence associated with iiPLA2β (left panels) and mitochondria marker (middle panels) and the merged fluorescence (right panels) are presented.
The choice of using the N-terminally tagged enzyme was based on the following: (1) We previously demonstrated that thapsigargin induces accumulation of full-length iPLA2β in the ER [38] and then in the mitochondria [39], therefore we focused on potential translocation of this iPLA2β isoform. (2) As illustrated in Fig. 4 above and reported earlier [34], iPLA2β undergoes proteolytic processing at the C-terminal region in vivo to generate a 70 kDa isoform that is catalytically active. Tagging the C-terminal of iPLA2β could potentially result in the loss of the EGFP label and hence, preclude the ability to visualize any potential translocation of the full-length protein.
As seen in Fig. 6, the merged image in the vehicle-treated group (top) exhibits a homogeneous yellow-orange color consistent with co-localization of fluorescences associated with iPLA2β and ER marker calreticulin, suggesting that iPLA2β is associated with ER under basal conditions. Following induction of ER stress, there is a time-dependent segregation of green fluorescence, suggesting that iPLA2β undergoes subcellular redistribution away from the ER. In contrast, though control iPLA2β fluorescence is prominent in Fig. 7, the merge image reveals that iPLA2β fluorescence is not strongly associated with the mitochondrial tracker suggesting that interaction between iPLA2β and mitochondria under basal conditions is minimal. However, following induction of ER stress the merged images reveal a time-dependent appearance of yellow-orange structures consistent with co-localization of fluorescences associated with iPLA2β and mitochondrial marker cytochrome c oxidase subunit VII, suggesting that iPLA2β undergoes progressive association with mitochondria.
3.6. ER stress-induced accumulation of iPLA2β in the ER and mitochondria
We next sought to identify the iPLA2β isoforms that accumulate in the mitochondria and ER. Because our earlier studies suggested that iPLA2β in INS-1 cells undergoes proteolytic cleavage at the N-terminus during ER stress [37], we performed these studies using cells expressing iPLA2β with EGFP fused at the C-terminus (fpN2). This allows detection of subcellular localization of iPLA2β isoforms that might be cleaved at the N-terminus because they would remain fused to EGFP. The cells were treated with vehicle (DMSO) alone or thapsigargin and harvested at 5 and 8 h for immunoblotting analyses. Probing for either iPLA2β (data not shown) or EGFP (Fig. 8A) reveals a similar immunoreactive pattern where only one band migrating with an apparent molecular mass of ca. 110 kDa is evident. Because this mass is reflective of a full-length iPLA2β fused to EGFP, these findings suggest that it is the full-length iPLA2β that localizes in the mitochondria and ER during ER stress. And, consistent with the findings with the organelle Trackers, ER stress caused a progressive decrease in ER iPLA2β and a concomitant increase in mitochondria-associated iPLA2β (Fig. 8B).
Fig. 8. ER stress-induced accumulation of iPLA2β isoforms in the ER and mitochondria.

INS-1 cells overexpressing fpN2 (iPLA2β-EGFP) were treated with vehicle (DMSO) alone or thapsigargin (1 µM, 37 °C) for 5 or 8 h. The cells were harvested and mitochondria and ER fractions prepared and aliquots (30 µg protein) of the fractions were analysed by SDS-PAGE (7.5 %) and processed for EGFP immunblotting analyses. A: EGFP Immnoublot B: Denistometry comparison. (Mito, mitochondria; and ER, endoplasmic reticulum; IDV, Integrated Density Value).
3.7. Mass spectrometry analyses of iPLA2β
To obtain further evidence of endogenous proteolytic processing, iPLA2® was purified from INS-1 cells and analysed by mass spectrometry. Using LC-MS/MS analyses, we detected not only peptides expected after digestion of iPLA2® with trypsin, but also a peptide arising from non-trypsin proteolytic processing. The doubly-charged ion at m/z 955.01 identified at retention time of 49 min (Fig. 9A) matches the theoretical molecular mass (1908.94 Da) of iPLA2® tryptic peptide 7L-R23, and MS/MS analysis (Fig. 9B) confirmed its amino acid sequence to be 7LVNTLSSVTNLFSNPFR23.
Fig. 9. Analyses of fragment ion with m/z of 955.01 arising from tryptic digestion of iPLA2β by mass spectrometry.

ATP affinity chromatography was used to purify iPLA2β from INS-1 cells overexpressing iPLA2β. Fractions containing iPLA2β catalytic activity were pooled, concentrated, and the concentrate was resolved by SDS-PAGE. Protein band migrating with an apparent molecular mass of 84 kDa corresponding to iPLA2β was identified by Coomassie staining, excised, and processed for digestion with trypsin. The tryptic digest was then analyzed by mass spectrometry. A: MS spectrum of doubly charged ion with m/z 955.01. B: Tandem MS spectrum of precursor ion with m/z 955.01.
As illustrated in Fig. 10A, at retention time 44 min, an abundant doubly-charged ion at m/z 791.90 was detected. However, this ion is not expected to arise from trypsin digestion of iPLA2®. Processing its MS/MS spectrum (Fig. 10B) using a Signature Discovery Algorithm program [50] revealed that it represents TLSSVTNLFSNPFR (1581.82 Da), which is part of the tryptic peptide 7LVNTLSSV10TNLFSNPFR23. The detection of 10T-R23 therefore reflects a “signature peptide” that derives from an iPLA2β variant produced by in vivo proteolytic processing.
Fig. 10. Analyses of fragment ion with m/z of 791.90 arising from tryptic digestion of iPLA2β by mass spectrometry.

Tryptic digest of iPLA2β was prepared as described for Fig. 9 and analysed by mass spectrometry. A: MS spectrum of doubly charged ion with m/z 791.90. B: Tandem MS spectrum of precursor ion with m/z 791.90.
4. Discussion
The Group VIA phospholipase A2 (iPLA2β) is proposed to play a role in several biological processes [8]. In the β-cell, we reported participation of iPLA2β in insulin secretion and apoptosis. Consistent with its involvement in these processes, inhibition or knock-out of iPLA2β activity suppresses [33, 35–41, 52, 53] and overexpression of iPLA2β amplifies [37–39, 42, 43, 45] secretagogue-stimulated insulin secretion and apoptosis.
In subsequent studies, insulin secretagogues were found to cause subcellular redistribution of iPLA2β from the cytoplasm to the perinuclear region. These observations were derived from immunofluorescence experiments that in principle could be confounded by non-specificity of iPLA2β antibodies and/or by cellular fixation artifacts [45]. To circumvent those issues iPLA2β was express as a fusion protein (fp) with EGFP fused to either the N-terminus (EGFP-iPLA2β, designated fpC2) or the C-terminus (iPLA2β-EGFP, designated fpN2) of iPLA2β, and subcellular localization within the β-cells was re-examined. INS-1 cells stably-transfected with the iPLA2β/EGFP constructs expressed several-fold higher Ca2+-independent PLA2 enzymatic activity than cells stably-transfected with the EGFP vector alone. Such activity is inhibited by BEL and stimulated by ATP, and both properties are characteristic of iPLA2β. This suggests that the fusion of EGFP to iPLA2β does not interfere with manifestation of iPLA2β catalytic activity.
The fusion of EGFP to iPLA2β allowed us to track iPLA2β movements within the cell by monitoring the green fluorescence of EGFP. The present findings reveal that stimulation of INS-1 cells that express iPLA2β/EGFP with secretagogues causes the appearance of punctate areas of fluorescence that reflect a subcellular redistribution of iPLA2β. Curiously, the punctate distribution of fluorescence differs between cells that express EGFP fused to the N-terminus compared to the C-terminus of iPLA2β. In the fpN2-expressing cells, the punctate fluorescence accumulates in the perinuclear region, resembling the pattern observed with immunofluorescence analyses [45]. In contrast, in the fpC2-expressing cells, the punctate area of fluorescence appears throughout the cytoplasm. Association of iPLA2β with subcellular compartments was next examined in combination with fluorescent Trackers targeted at specific organelles and in contrast to the findings of fpN2 localization in the ER and Golgi, fpC2 did not appear to associate with the ER, Golgi, mitochondria, or plasma membrane.
The differential subcellular iPLA2β localization in the fpN2- and fpC2-expressing cells is probably a reflection of endogenous processing of iPLA2β and the presence or absence of amino acid sequences that might target iPLA2β to specific subcellular organelles. The expected sizes of the fp are 110–113 kDa, of which EGFP accounts for 26–29 kDa and such bands are observed. Two additional EGFP-immunoreactive bands that migrate with apparent molecular masses of ca. 90 kDa and 100 kDa are also visualized. The 90 kDa band is evident only in fpC2-expressing cells, and the 100 kDa band only in fpN2-expressing cells. We reported that β-cells express at least two catalytically-active truncated isoforms of iPLA2β; a 63 kDa isoform that is a product of caspase-3-mediated cleavage [37] and a 70 kDa isoform that arises from proteolytic removal of C-terminus sequence [34]. Interestingly, the 70 kDa iPLA2β variant is the predominant isoform in parental INS-1 cells and native pancreatic islets.
Our observations here and elsewhere [34, 37] are consistent with the possibility that the 90 kDa band represents the iPLA2β variant that is produced by caspase-3 cleavage. As the EGFP is fused to the C-terminus of fpN2, the 90 kDa iPLA2β caspase-3-cleavage product is still recognized by the EGFP antibody. Similar cleavage of the fpC2 would result in the loss of EGFP fused to the N-terminus of iPLA2β, and this would prevent recognition by the EGFP antibody. Analogously, the 100 kDa iPLA2β-EGFP band probably represents the fusion product of EGFP and that region of iPLA2β contained in the native 70 kDa iPLA2β isoform. Cleavage of C-terminal residues from fpN2 would result in the loss of EGFP fused to the C-terminus of iPLA2β, and this would prevent recognition by the EGFP antibody. Collectively, these findings suggest that iPLA2β undergoes post-translational proteolytic processing and that an isoform from which N-terminal sequence has been removed accumulates in the ER and Golgi following cell stimulation. The iPLA2β isoform that forms a punctate distribution in the cytosol upon cell stimulation retains the N-terminus. The iPLA2β isoform that accumulates in the ER and Golgi upon cell stimulation retains the C-terminus.
Stimulus-induced translocation of the Group V sPLA2 and Group IV cPLA2 to the perinuclear region has also been demonstrated [54, 55], and cPLA2 has been reported to translocate to the Golgi and/or ER depending on the stimulus [56]. In this regard, we recently found evidence of time-dependent accumulation of iPLA2β protein in the ER, mitochondria, and nucleus of INS-1 cells following long-term (4–24 h) exposure to the SERCA inhibitor thapsigargin [37–39]. Consistent with these observations, organelle tracking analyses (Fig. 6 and Fig. 7) performed in the present study reveal that induction of ER stress in INS-1 cells promotes redistribution of iPLA2β from the ER and its subsequent accumulation in the mitochondria. This is supported by immunoblotting analyses (Fig. 8) of isolated ER and mitochondrial fractions
However, it should be noted that the basal levels of iPLA2β in the two fractions appear to be different depending on the analyses used. Results with the organelle Trackers suggest that basal ER accumulation of iPLA2β is greater than in the mitochondria but the immunonoblotting findings, similar to our earlier observations [38, 39], suggest the opposite. To reconcile this discrepancy, it is necessary to consider the fusion protein used in the two analyses. Organelle Trackers were used in cells expressing iPLA2β fused to EGFP at the N-terminus and, as described above, proteolytic cleavage at the C-terminal end of this fusion protein would generate a truncated iPLA2β protein. It might be speculated that this leads to deletion of a mitochondrial targeting sequence precluding accumulation of the truncated iPLA2β in the mitochondria. It could be further speculated that induction of ER stress activates enzymes and/or generates factors that inhibit proteolytic cleavage of iPLA2β allowing the full-length iPLA2β to translocate to the mitochondria. Because the immunoblotting analyses were performed using cells expressing iPLA2β fused to EGFP at the C-terminus, it is plausible that this fusion protein is protected from proteolytic cleavage at the C-terminal and therefore is able to retain the mitochondrial targeting sequence. This would allow its localization in the mitochondria under basal conditions and its accumulation following induction of ER stress.
Interestingly, there appear to be differences in iPLA2β localization following secretagogue stimulation and induction of ER stress. For instance, translocation events following exposure to secretagogue occur within 30 min and last up to 2 h. In contrast the onset of iPLA2β mobilization after exposure to thapsigargin is later and lasts up to 20–24 h. Further, while the localization after secretagogue treatment concurrently appears predominantly in the ER and the Golgi, it proceeds from the ER to the mitochondria following ER stress induction. Additionally, we previously reported that ER stress induces nuclear association of iPLA2β [45] and recently observed that ER stress induces iPLA2β expression (Lei et al., in press). To date, we have no evidence of nuclear accumulation or induction of iPLA2β expression in INS-1 cells following secretagogue exposure. These findings suggest that the type and duration of stimulus can influence subcellular distribution of iPLA2β.
The likelihood that iPLA2β is processed endogenously was first evident from studies demonstrating that the 70 kDa protein was an iPLA2β isoform that was truncated at the C-terminus and was not a product resulting from alternate splicing of the iPLA2βtranscript [34]. Subsequently, we obtained evidence for other iPLA2β variants that were truncated at various amino acid residues at the N-terminus [50, 57]. In the present study, further evidence of iPLA2β variants generated by endogenous proteolytic processing of iPLA2β was obtained using a Signature Discovery Algorithm program developed in our laboratory to analyse peptide fragments identified by mass spectrometry protocols [50]. Analyses of tryptic digest of purified iPLA2β by this method identified several peptide fragments arising from tryptic digestion of iPLA2β. In addition to the expected fragments, however, a doubly-charged ion with m/z 791.90 is evident and this was identified by tandem mass spectrometry as having the amino acid sequence of 10T-R23. Because this is an unexpected fragment, 10T-R23 reflects a “signature peptide” that derives from an iPLA2β variant produced by in vivo proteolytic processing.
The size of these proteolytic processed proteins are most likely 1–2 kDa smaller than the full length protein and this is deduced based on the following observations: (1) The identified N-terminal fragments arose from digestion of an iPLA2β-immunoreactive band which migrated with an apparent molecular mass of ca. 80–84 kDa. (2) The protein was His-tagged at the C-terminus and was purified using a cobalt-affinity column and visualization of an iPLA2β-immunoreactive band would only be possible if the His-tag was still present on the protein, allowing it to be eluted from the column. Thus, the fragments analysed by MS arose from a protein that spanned the N-terminal amino acids (minus the 1–2 kDa truncated residues) to the end of the C-terminal residues.
Though direct evidence is not yet available, the current observations raise the possibility that proteolytic processing of iPLA2β may be a factor in its redistribution. Consistent with this possibility are the findings that iPLA2β accumulates in different subcellular organelles under different conditions and differential localization is evident fpN2 and fpC2 following secretagogue stimulation. This raises the likelihood that the two termini contain potential targeting sequences that may be lost from the cleaved proteins. Future studies that will include expression of processed iPLA2β isoforms to more directly track their mobilization following different stimuli should facilitate clarification of the affect of proteolytic processing on iPLA2β translocation.
In summary, we find that iPLA2β undergoes endogenous proteolytic processing and exhibits variable subcellular localization. Studies are currently underway to express the different iPLA2β isoforms in INS-1 cells to determine the identities of the variants that associate with specific subcellular organelles under different stimulus conditions. These studies should result in identification of the regions of iPLA2β sequence that mediate interaction with an organelle and potentially, to the design of targeted compounds that could enhance or suppress interaction of iPLA2β with a specific organelle.
Acknowledgements
This research was supported by grants from National Institutes of Health (DK69455, DK34388, P41-RR00954, P60-DK20579, P30-DK56341) and by an Award (to SR) from the American Diabetes Association.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
ABBREVIATIONS
Abbreviations used in this manuscript are: BEL, bromoenol lactone suicide substrate inhibitor of iPLA2β; BSA, bovine serum albumin; C2 and N2, EGFP-C2 and EGFP-N2 vectors, respectively; ECL, enhanced chemiluminescence; EGFP, enhanced green fluorescence protein; ER, endoplasmic reticulum; fp, iPLA2β/EGFP fusion protein; fpC2, NEGFP-iPLA2βC fusion protein; fpC2, NiPLA2β-EGFPC fusion protein; iPLA2β, Group VIA Ca2+-independent phospholipase A2; kDa, kilodaltons; MS, mass spectrometry; OE, iPLA2β overexpressing cells; O/N, overnight; PAGE, polyacrylamide gel electrophoresis; PLA2, phospholipase A2; PM, plasma membrane; Q-TOF, Quadrupole-time of flight; SDS, sodium dodecyl sulfate; and RT, room temperature
References
- 1.Dennis EA. The growing phospholipase A2 superfamily of signal transduction enzymes. Trends Biochem. Sci. 1997;22:1–2. doi: 10.1016/s0968-0004(96)20031-3. [DOI] [PubMed] [Google Scholar]
- 2.Gijon MA, Leslie CC. Phospholipases A2. Semin. Cell Dev. Biol. 1997;8:297–303. doi: 10.1006/scdb.1997.0151. [DOI] [PubMed] [Google Scholar]
- 3.Gijon MA, Spencer DM, Kaiser AL, Leslie CC. Role of phosphorylation sites and the C2 domain in regulation of cytosolic phospholipase A2. J. Cell Biol. 1999;145:1219–1232. doi: 10.1083/jcb.145.6.1219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ma Z, Ramanadham S, Wohltmann M, Bohrer A, Hsu FF, Turk J. Studies of insulin secretory responses and of arachidonic acid incorporation into phospholipids of stably transfected insulinoma cells that overexpress group VIA phospholipase A2 (iPLA2β) indicate a signaling rather than a housekeeping role for iPLA2β. J. Biol. Chem. 2001;276:13198–13208. doi: 10.1074/jbc.M010423200. [DOI] [PubMed] [Google Scholar]
- 5.Mancuso DJ, Jenkins CM, Gross RW. The genomic organization, complete mRNA sequence, cloning, and expression of a novel human intracellular membrane-associated calcium-independent phospholipase A2. J. Biol. Chem. 2000;275:9937–9945. doi: 10.1074/jbc.275.14.9937. [DOI] [PubMed] [Google Scholar]
- 6.Ma Z, Turk J. The molecular biology of the group VIA Ca2+-independent phospholipase A2. Prog Nucleic Acid Res Mol. Biol. 2001;67:1–33. doi: 10.1016/s0079-6603(01)67023-5. [DOI] [PubMed] [Google Scholar]
- 7.Wang Z, Ramanadham S, Ma ZA, Bao S, Mancuso DJ, Gross RW, Turk J. Group VIA Phospholipase A2 forms a signaling complex with the calcium/calmodulin-dependent protein kinase IIβ expressed in pancreatic islet β-cells. J. Biol. Chem. 2005;280:6840–6849. doi: 10.1074/jbc.M405287200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Seashols SJ, del Castillo Olivares A, Gil G, Barbour SE. Regulation of group VIA phospholipase A2 expression by sterol availability. Biochim. Biophys. Acta (BBA) – Mol. Cell Biol. Lipids. 2004;1684:29–37. doi: 10.1016/j.bbalip.2004.05.003. [DOI] [PubMed] [Google Scholar]
- 9.Ma Z, Wang X, Nowatzke W, Ramanadham S, Turk J. Human pancreatic islets express mRNA species encoding two distinct catalytically active isoforms of group VI phospholipase A2 (iPLA2) that arise from an exon-skipping mechanism of alternative splicing of the transcript from the iPLA2 gene on chromosome 22q13.1. J. Biol. Chem. 1999;274:9607–9616. doi: 10.1074/jbc.274.14.9607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Balsinde J, Bianco ID, Ackermann EJ, Conde-Frieboes K, Dennis EA. Inhibition of calciumin-dependent phospholipase A2 prevents arachidonic acid incorporation and phospholipid remodeling in P388D1 macrophages. Proc. Natl. Acad. Sci. 1995;92:8527–8531. doi: 10.1073/pnas.92.18.8527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Boilard E, Surette ME. Anti-CD3 and concanavalin A-induced human t cell proliferation is associated with an increased rate of arachidonate-phospholipid remodeling. Lack of involvement of group iv and group VI phospholipase A2 in remodeling and increased susceptibility of proliferating T cells to CoA-independent transacylase inhibitor-induced apoptosis. J. Biol. Chem. 2001;276:17568–17575. doi: 10.1074/jbc.M006152200. [DOI] [PubMed] [Google Scholar]
- 12.Isenovic E, LaPointe MC. Role of Ca2+-Independent phospholipase A2 in the regulation of inducible nitric oxide synthase in cardiac myocytes. Hypertension. 2000;35:249–254. doi: 10.1161/01.hyp.35.1.249. [DOI] [PubMed] [Google Scholar]
- 13.Maggi LB, Jr, Moran JM, Scarim AL, Ford DA, Yoon J-W, McHowat J, Buller RML, Corbett JA. Novel role for calcium-independent phospholipase A2 in the macrophage antiviral response of inducible nitric-oxide synthase expression. J. Biol. Chem. 2002;277:38449–38455. doi: 10.1074/jbc.M206247200. [DOI] [PubMed] [Google Scholar]
- 14.Moran JM, Buller RML, McHowat J, Turk J, Wohltmann M, Gross RW, Corbett JA. Genetic and pharmacologic evidence that calcium-independent phospholipase A2β regulates virus-induced inducible nitric-oxide synthase expression by macrophages. J. Biol. Chem. 2005;280:28162–28168. doi: 10.1074/jbc.M500013200. [DOI] [PubMed] [Google Scholar]
- 15.Tithof PK, Olivero J, Ruehle K, Ganey PE. Activation of neutrophil calcium-dependent and - independent phospholipases A2 by organochlorine compounds. Toxicol. Sci. 2000;53:40–47. doi: 10.1093/toxsci/53.1.40. [DOI] [PubMed] [Google Scholar]
- 16.Williams SD, Ford DA. Calcium-independent phospholipase A2 mediates CREB phosphorylation and c-fos expression during ischemia. Am. J. Physiol. Heart Circ. Physiol. 2001;281:H168–H176. doi: 10.1152/ajpheart.2001.281.1.H168. [DOI] [PubMed] [Google Scholar]
- 17.Bao S, Miller DJ, Ma Z, Wohltmann M, Eng G, Ramanadham S, Moley K, Turk J. Male mice that do not express group VIA phospholipase A2 produce spermatozoa with impaired motility and have greatly reduced fertility. J. Biol. Chem. 2004;279:38194–38200. doi: 10.1074/jbc.M406489200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ramanadham S, Yarasheski KE, Silva MJ, Wohltmann M, Novack DV, Christiansen B, Tu X, Zhang S, Lei X, Turk J. Age-related changes in bone morphology are accelerated in group VIA phospholipase A2 (iPLA2β)-null mice. Am. J. Pathol. 2008;172:868–881. doi: 10.2353/ajpath.2008.070756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Carper MJ, Zhang S, Turk J, Ramanadham S. Skeletal muscle group VIA phospholipase A2 (iPLA2β): Expression and role in fatty acid oxidation. Biochemistry. 2008;47:12241. doi: 10.1021/bi800923s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.St-Gelais F, Menard C, Congar P, Trudeau LE, Massicotte G. Postsynaptic injection of calcium-independent phospholipase A2 inhibitors selectively increases AMPA receptor-mediated synaptic transmission. Hippocampus. 2004;14:319–325. doi: 10.1002/hipo.10176. [DOI] [PubMed] [Google Scholar]
- 21.Schaeffer EL, Gattaz WF. Inhibition of calcium-independent phospholipase A2 activity in rat hippocampus impairs acquisition of short- and long-term memory. Psychopharmacology (Berl) 2005;181:392–400. doi: 10.1007/s00213-005-2256-9. [DOI] [PubMed] [Google Scholar]
- 22.Mendes CT, Gattaz WF, Schaeffer EL, Forlenza OV. Modulation of phospholipase A2 activity in primary cultures of rat cortical neurons. J. Neural. Transm. 2005;112:1297–1308. doi: 10.1007/s00702-004-0271-3. [DOI] [PubMed] [Google Scholar]
- 23.Berti-Mattera LN, Harwalkar S, Hughes B, Wilkins PL, Almhanna K. Proliferative and morphological effects of endothelins in Schwann cells: roles of p38 mitogen-activated protein kinase and Ca2+-independent phospholipase A2. J. Neurochem. 2001;79:1136–1148. doi: 10.1046/j.1471-4159.2001.00642.x. [DOI] [PubMed] [Google Scholar]
- 24.Biancheri R, Rossi A, Alpigiani G, Filocamo M, Gandolfo C, Lorini R, Minetti C. Cerebellar atrophy without cerebellar cortex hyperintensity in infantile neuroaxonal dystrophy (INAD) due to PLA2G6 mutation. Eur. J. Paediatr. Neurol. 2007;11:175–177. doi: 10.1016/j.ejpn.2006.11.013. [DOI] [PubMed] [Google Scholar]
- 25.Westaway SK, Gregory A, Hayflick SJ. Mutations in PLA2G6 and the riddle of Schindler disease. J. Med. Genet. 2007;44:e64. doi: 10.1136/jmg.2006.044966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Junqueira R, Cordeiro Q, Meira-Lima I, Gattaz WF, Vallada H. Allelic association analysis of phospholipase A2 genes with schizophrenia. Psychiatr. Genet. 2004;14:157–160. doi: 10.1097/00041444-200409000-00007. [DOI] [PubMed] [Google Scholar]
- 27.Smesny S, Kinder D, Willhardt I, Rosburg T, Lasch J, Berger G, Sauer H. Increased calcium-independent phospholipase A2 activity in first but not in multiepisode chronic schizophrenia. Biol. Psychiatry. 2005;57:399–405. doi: 10.1016/j.biopsych.2004.11.018. [DOI] [PubMed] [Google Scholar]
- 28.Boittin FX, Petermann O, Hirn C, Mittaud P, Dorchies OM, Roulet E, Ruegg UT. Ca2+-independent phospholipase A2 enhances store-operated Ca2+ entry in dystrophic skeletal muscle fibers. J. Cell Sci. 2006;119:3733–3742. doi: 10.1242/jcs.03184. [DOI] [PubMed] [Google Scholar]
- 29.Gong MC, Arbogast S, Guo Z, Mathenia J, Su W, Reid MB. Calcium-independent phospholipase A2 modulates cytosolic oxidant activity and contractile function in murine skeletal muscle cells. J. Appl. Physiol. 2006;100:399–405. doi: 10.1152/japplphysiol.00873.2005. [DOI] [PubMed] [Google Scholar]
- 30.Kan H, Xie Z, Finkel MS. iPLA2 inhibitor blocks negative inotropic effect of HIV gp120 on cardiac myocytes. J. Mol. Cell. Cardiol. 2006;40:131–137. doi: 10.1016/j.yjmcc.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 31.Ronkko S, Rekonen P, Kaarniranta K, Puustjarvi T, Terasvirta M, Uusitalo H. Phospholipase A2 in chamber angle of normal eyes and patients with primary open angle glaucoma and exfoliation glaucoma. Mol. Vis. 2007;13:408–417. [PMC free article] [PubMed] [Google Scholar]
- 32.Gilroy DW, Newson J, Sawmynaden P, Willoughby DA, Croxtall JD. A novel role for phospholipase A2 isoforms in the checkpoint control of acute inflammation. FASEB J. 2004;18:489–498. doi: 10.1096/fj.03-0837com. [DOI] [PubMed] [Google Scholar]
- 33.Ramanadham S, Gross RW, Han X, Turk J. Inhibition of arachidonate release by secretagogue-stimulated pancreatic islets suppresses both insulin secretion and the rise in beta-cell cytosolic calcium ion concentration. Biochemistry. 1993;32:337–346. doi: 10.1021/bi00052a042. [DOI] [PubMed] [Google Scholar]
- 34.Ramanadham S, Song H, Hsu FF, Zhang S, Crankshaw M, Grant GA, Newgard CB, Bao S, Ma Z, Turk J. Pancreatic islets and insulinoma cells express a novel isoform of group VIA phospholipase A2 (iPLA2β) that participates in glucose-stimulated insulin secretion and is not produced by alternate splicing of the iPLA2β transcript. Biochemistry. 2003;42:13929–13940. doi: 10.1021/bi034843p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ramanadham S, Wolf MJ, Jett PA, Gross RW, Turk J. Characterization of an ATP-stimulatable Ca2+-independent phospholipase A2 from clonal insulin-secreting HIT cells and rat pancreatic islets: a possible molecular component of the beta-cell fuel sensor. Biochemistry. 1994;33:7442–7452. doi: 10.1021/bi00189a052. [DOI] [PubMed] [Google Scholar]
- 36.Ramanadham S, Wolf MJ, Li B, Bohrer A, Turk J. Glucose-responsitivity and expression of an ATP-stimulatable, Ca2+-independent phospholipase A2 enzyme in clonal insulinoma cell lines. Biochim. Biophys. Acta. 1997;1344:153–164. doi: 10.1016/s0005-2760(96)00139-7. [DOI] [PubMed] [Google Scholar]
- 37.Ramanadham S, Hsu FF, Zhang S, Jin C, Bohrer A, Song H, Bao S, Ma Z, Turk J. Apoptosis of insulin-secreting cells induced by endoplasmic reticulum stress is amplified by overexpression of group VIA calcium-independent phospholipase A2 (iPLA2β) and suppressed by inhibition of iPLA2β. Biochemistry. 2004;43:918–930. doi: 10.1021/bi035536m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lei X, Zhang S, Bohrer A, Bao S, Song H, Ramanadham S. The Group VIA calcium-independent phospholipase A2 participates in ER stress-induced INS-1 insulinoma cell apoptosis by promoting ceramide generation via hydrolysis of sphingomyelins by neutral sphingomyelinase. Biochemistry. 2007;46:10170–10185. doi: 10.1021/bi700017z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lei X, Zhang S, Bohrer A, Ramanadham S. Calcium-independent phospholipase A2 (iPLA2β)-mediated ceramide generation plays a key role in the cross-talk between the endoplasmic reticulum (ER) and mitochondria during er stress-induced insulin-secreting cell apoptosis. J. Biol. Chem. 2008;283:34819–34832. doi: 10.1074/jbc.M807409200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bao S, Song H, Wohltmann M, Ramanadham S, Jin W, Bohrer A, Turk J. Insulin secretory responses and phospholipid composition of pancreatic islets from mice that do not express group VIA phospholipase A2 and effects of metabolic stress on glucose homeostasis. J. Biol. Chem. 2006;281:20958–20973. doi: 10.1074/jbc.M600075200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bao S, Bohrer A, Ramanadham S, Jin W, Zhang S, Turk J. Effects of stable suppression of group VIA phospholipase A2 expression on phospholipid content and composition, insulin secretion, and proliferation of INS-1 insulinoma cells. J. Biol. Chem. 2006;281:187–198. doi: 10.1074/jbc.M509105200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bao S, Jacobson DA, Wohltmann M, Bohrer A, Jin W, Philipson LH, Turk J. Glucose homeostasis, insulin secretion, and islet phospholipids in mice that overexpress iPLA2β in pancreatic β-cells and in iPLA2β-null mice. Am. J. Physiol. Endocrinol. Metab. 2008;294:E217–E229. doi: 10.1152/ajpendo.00474.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ma Z, Bohrer A, Wohltmann M, Ramanadham S, Hsu F-F, Turk J. Studies of phospholipid metabolism, proliferation, and secretion of stably transfected insulinoma cells that overexpress group VIA phospholipase A2. Lipids. 2001;36:689–700. doi: 10.1007/s11745-001-0774-9. [DOI] [PubMed] [Google Scholar]
- 44.Ramanadham S, Hsu F-F, Bohrer A, Ma Z, Turk J. Studies of the role of group vi phospholipase A2 in fatty acid incorporation, phospholipid remodeling, lysophosphatidylcholine generation, and secretagogue-induced arachidonic acid release in pancreatic islets and insulinoma cells. J. Biol. Chem. 1999;274:13915–13927. doi: 10.1074/jbc.274.20.13915. [DOI] [PubMed] [Google Scholar]
- 45.Ma Z, Zhang S, Turk J, Ramanadham S. Stimulation of insulin secretion and associated nuclear accumulation of iPLA2β in INS-1 insulinoma cells. Am. J. Physiol. Endocrinol. Metab. 2002;282:E820–E833. doi: 10.1152/ajpendo.00165.2001. [DOI] [PubMed] [Google Scholar]
- 46.Bao S, Jin C, Zhang S, Turk J, Ma Z, Ramanadham S. Beta-cell calcium-independent group VIA phospholipase A2 (iPLA2β): Tracking iPLA2β movements in response to stimulation with insulin secretagogues in INS-1 cells. Diabetes. 2004;53:S186–S189. doi: 10.2337/diabetes.53.2007.s186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Cormack BP, Valdivia RH, Falkow S. FACS-optimized mutants of the green fluorescent protein (GFP) Gene. 1996;173:33–38. doi: 10.1016/0378-1119(95)00685-0. [DOI] [PubMed] [Google Scholar]
- 48.Gross RW, Ramanadham S, Kruszka KK, Han X, Turk J. Rat and human pancreatic islet cells contain a calcium ion independent phospholipase A2 activity selective for hydrolysis of arachidonate which is stimulated by adenosine triphosphate and is specifically localized to islet beta-cells. Biochemistry. 1993;32:327–336. doi: 10.1021/bi00052a041. [DOI] [PubMed] [Google Scholar]
- 49.Bao S, Li Y, Lei X, Wohltmann M, Jin W, Bohrer A, Semenkovich CF, Ramanadham S, Tabas I, Turk J. Attenuated free cholesterol loading-induced apoptosis but preserved phospholipid composition of peritoneal macrophages from mice that do not express group VIA phospholipase A2. J. Biol. Chem. 2007;282:27100–27114. doi: 10.1074/jbc.M701316200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Song H, Hecimovic S, Goate A, Hsu FF, Bao S, Vidavsky I, Ramanadham S, Turk J. Characterization of N-terminal processing of group VIA phospholipase A2 and of potential cleavage sites of amyloid precursor protein constructs by automated identification of signature peptides in LC/MS/MS analyses of proteolytic digests. J. Am. Soc. Mass Spectrom. 2004;15:1780–1793. doi: 10.1016/j.jasms.2004.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ramanadham S, Hsu F, Zhang S, Bohrer A, Ma Z, Turk J. Electrospray ionization mass spectrometric analyses of phospholipids from INS-1 insulinoma cells: comparison to pancreatic islets and effects of fatty acid supplementation on phospholipid composition and insulin secretion. Biochim. Biophys. Acta. 2000;1484:251–266. doi: 10.1016/s1388-1981(00)00022-6. [DOI] [PubMed] [Google Scholar]
- 52.Ma Z, Ramanadham S, Kempe K, Chi XS, Ladenson J, Turk J. Pancreatic islets express a Ca2+-independent phospholipase A2 enzyme that contains a repeated structural motif homologous to the integral membrane protein binding domain of ankyrin. J. Biol. Chem. 1997;272:11118–11127. [PubMed] [Google Scholar]
- 53.Ma Z, Ramanadham S, Hu Z, Turk J. Cloning and expression of a group IV cytosolic Ca2+-dependent phospholipase A2 from rat pancreatic islets. Comparison of the expressed activity with that of an islet group VI cytosolic Ca2+-independent phospholipase A2. Biochim. Biophys. Acta. 1998;1391:384–400. doi: 10.1016/s0005-2760(98)00027-7. [DOI] [PubMed] [Google Scholar]
- 54.Fatima S, Yaghini FA, Ahmed A, Khandekar Z, Malik KU. CaM kinase IIα mediates norepinephrine-induced translocation of cytosolic phospholipase A2 to the nuclear envelope. J. Cell Sci. 2003;116:353–365. doi: 10.1242/jcs.00242. [DOI] [PubMed] [Google Scholar]
- 55.Kim YJ, Kim KP, Rhee HJ, Das S, Rafter JD, Oh YS, Cho W. Internalized group V secretory phospholipase A2 acts on the perinuclear membranes. J. Biol. Chem. 2002;277:9358–9365. doi: 10.1074/jbc.M110987200. [DOI] [PubMed] [Google Scholar]
- 56.Evans JH, Spencer DM, Zweifach A, Leslie CC. Intracellular calcium signals regulating cytosolic phospholipase A2 translocation to internal membranes. J. Biol. Chem. 2001;276:30150–30160. doi: 10.1074/jbc.M100943200. [DOI] [PubMed] [Google Scholar]
- 57.Ramanadham S, Song H, Bao S, Hsu F-F, Zhang S, Ma Z, Jin C, Turk J. Islet complex lipids: Involvement in the actions of group VIA calcium-independent phospholipase A2 in β-Cells. Diabetes. 2004;53:S179–S185. doi: 10.2337/diabetes.53.2007.s179. [DOI] [PMC free article] [PubMed] [Google Scholar]