

Abstract
How does the brain determine what to learn and what not to learn? Previous studies showed that a feature or stimulus on which subjects performed a task was learned, while the features or stimuli that were irrelevant to the task were not learned. This led some researchers to conclude that attention to a stimulus was necessary for the stimulus to be learned. This thought was challenged by the discovery of a task-irrelevant perceptual learning, in which learning occurred by mere exposure to the unattended and subthreshold stimulus. However, this exposure-based learning does not necessarily indicate that all presented stimuli are learned. Rather, recent studies showed that the occurrence of this learning was very selective for the following new findings: unattended stimulus learning occurred only (1) when the unattended stimulus was associated temporally with the processing of an attended target, (2) when the unattended stimulus was synchronously presented with reinforcers, such as internal or external rewards, and (3) when the unattended stimulus had subliminal properties. These selectivities suggest some degrees of similarity between task-relevant and task-irrelevant perceptual learning, which has been the motivation for making a united model in which both task-relevant and task-irrelevant learning are formed with similar or same mechanisms.
Keywords: perceptual learning, task-irrelevant perceptual learning, exposure-based perceptual learning
What to learn and what not to learn? This is a very critical question for those interested in adaptation of the human brain to its environment. As the surrounding environments continuously change, the brain needs to selectively accept some critical changes and reject others in an attempt to maintain the basic architecture it needs to correctly perceive and recognize the environment. This ‘stability-plasticity dilemma’ (Grossberg, 1980; Spanis and Squire, 1987) is particularly interesting in the sensory areas of the adult brain. Learning has commonly been linked with the higher cognitive levels (i.e. decision making) rather than the lower sensory levels (i.e. various perception processes). This may have stemmed from our belief that perceptual abilities are not subject to change with training. Traditional thought of perceptual development has held the early sensory area to be adaptable only during early development (the “critical period”), after which it then becomes hard-wired. Some apparently non-significant differences between perceptual abilities of children and adults have served to confirm this belief.
However, recent studies have indicated that perceptual abilities of adults can improve with training. A study by Sowden, Davies and Roling (2000) has noted that trained radiologists are capable of distinguishing subtle patterns of tumors that naïve people could not possibly detect. Studies employing detection and/or discrimination tasks to observe more basic perceptual abilities also showed that abilities could be improved with training. The improvement was highly specific for trained features, including retinotopic location and orientation (Fiorentini and Berardi, 1980; see a review in Seitz and Watanabe, 2005). Such specificity suggested that learning occurred at the perceptual level (rather than the higher cognitive level); in other words, learning may be mediated in the early sensory area (Fahle, 2005).
The fact that perceptual learning can occur with adults as well gives rise to the importance of exploring the afore-mentioned question: how the brain selects for what to learn and what not to. This selection nature of learning had fueled numerous researchers towards focusing on the role of attention. Shiu and Pashler (1992) conducted a study evaluating subjects’ performances in discriminating line orientations. While subjects who were trained to do the orientation discrimination task showed improvement, those who were trained to discriminate the brightness of the line (instead of the orientation) showed no improvement. Another study has reported that learning did not transfer between two different tasks. Namely, practicing a task of discriminating the orientation of a global shape did not improve performance in a task of detecting a local element, even though both tasks used the same visual stimuli (Ahissar and Hochstein, 1993). These observations were consistent with results from several physiological studies. One study found the orientation tuning curves of macaque monkey V1 cells to be sharpened in relation to the location of the stimulus with which they performed an orientation discrimination task during the training session, but not in relation to the location of the stimulus that was merely exposed (Schoups et al., 2001). The above results thus led to the prevailing hypothesis that perceptual learning can only occur with focused attention to the feature relevant to the given task.
This hypothesis was challenged by recent findings of the task-irrelevant perceptual learning in which learning of a feature occurred in the absence of attention to the feature (Watanabe, Náñez and Sasaki, 2001). In their experiment, subjects were asked to select and identify letters from a rapid serial visual presentation (RSVP) display, a rapid succession of letters and numbers, which was presented against a background of dynamic random-dot (DRD) display consisting of moving dots (signal) and randomly moving dots (noise). Because this motion display was not related to the central task, the display was out of the focus of the subject’s attention. This task-irrelevant motion display, moreover, had a very weak motion direction signal. The display was on a 5% coherence level – five percent of the motion display consisted of coherently moving dots and 95 percent consisted of randomly moving dots. In this low coherent motion display, the direction of coherently moving dots could hardly be perceived, detected or recognized; in other words, the strength of the motion signals was below the thresholds, or subliminal. Despite its irrelevancy and indifferentiability, repeated exposure to the background motion display improved subjects’ sensitivity specifically to the trained direction (see Fig. 1). A follow-up study demonstrated that this task-irrelevant learning was highly specific to the local motion of the exposed stimuli, as opposed to the global motion, and that learning was retained for months after the training (Watanabe et al., 2002). These findings implied that focused attention to a feature was not necessary for perceptual learning of the feature.
Figure 1.

Illustration of an experiment conducted by Watanabe et al. (2001), which reported task-irrelevant perceptual learning. (a) Outline of the whole experiment. Identical tests were conducted before and after the exposure (or training) session to measure the learning effect, the difference between the pre- and post-test sessions. (b) Procedure of the task in test sessions. In each trial, dynamic random dot (DRD) displays were presented with each display containing either a 5% or 10% coherent motion moving in one of eight directions. 500 ms later, this display was replaced with eight arrows. Subjects selected the one arrow that best matched the coherent motion direction. (c) Procedure of the task in exposure sessions. Subjects were asked to report the two white target letters that appeared in a sequence of black distractor letters. In a peripheral annulus, a 5% coherent motion display was present in a consistent direction throughout the exposure sessions for each subject. (d) Mean performance of pre- and post-tests. Relative motion direction refers to the difference between tested direction and exposed direction. Thus, 0 indicates the exposed direction during exposure sessions. Significant improvement was found at and around the exposed direction for 10% coherent motion after the exposure.
HOW TO SELECT FOR WHAT TO LEARN AND WHAT NOT TO LEARN?
The dispensability of attention for perceptual learning complicates the question of selectivity. If perceptual learning of a feature can occur in the absence of attention to the feature, and as a result of exposure to the feature, how does the brain decide what to learn and what not to? Is this exposure-based learning purely passive? Recent studies have found three conditions in which this type of learning has occurred.
1. The processing of a task-target
The first condition is that task-irrelevant perceptual learning is related to the subject’s engagement in the task target during the training sessions. Seitz and Watanabe (2003) showed that a sensitivity enhancement occurred only when a task-irrelevant stimulus was paired with a task target, using a very similar experimental setup to the original one of the task-irrelevant perceptual learning (Watanabe et al., 2001). In their study, four subliminal motion displays moving in all different directions were presented an equal number of times in each trial during the training session. One motion with a specific direction was temporally paired with a target at all times, while the other three motion directions were consistently paired with distractors. Even though all directions of motion were exposed equally in quantity, learning occurred only for the direction paired with a task target.
The critical factor of exposure-based visual learning, however, turned out not to be temporal pairing with a task target, per se. Rather, it depended on whether a target was completely processed or not. When subjects failed to identify the target, learning of a feature paired with the target did not occur (Seitz et al., 2005). In order to interrupt the processing of a target, the attentional blink effect was employed. In this effect, an RSVP display consisting of two targets and several distractors is presented. The second target appears within a much shorter time frame after the first target appears, thereby lessening the subject’s ability to identify the second target. As in the study by Seitz and Watanabe (2003), a subliminal motion display was used as a task-irrelevant stimulus and paired temporally with a target. While the learning effect was found for the direction paired with a target that subjects could easily identify, no learning effect was found for the direction paired with a target that was hard to be detected because of this short interval from the previously presented target. These results indicated that the task-irrelevant features to be learned were determined in relation to the processing of a target in which subject are engaged.
2. The establishment of reinforcement
The importance of target processing for the exposure-based perceptual learning provided a new viewpoint in understanding how the selection process connected to learning. The fact that an irrelevant stimulus can be learned when paired with a task-relevant stimulus suggests that exposure-based learning has some similarities to classical conditioning. In classical conditioning, learning occurs through pairing of a conditioned stimulus (CS, which usually does not influence a subject’s response) with an unconditioned stimulus (US, which evokes a subject’s response directly).
Yet a direct comparison of the two does not seem to be relevant. Whereas studies of conditioning focus on whether the behavioral response to a stimulus (that is, US) is evoked by another neutral stimulus (that is, CS), studies on learning are interested in the change of sensitivity for an irrelevant stimulus. In other words, the result of conditioning is a change of association between a stimulus and a response, and the result of perceptual learning is the strengthening of the sensitivity to a stimulus. A more critical difference between conditioning and learning is the role of an inducer. In conditioning, the inducer (US) functions as a type of a reinforcement, or a reward. On the other hand, it is not clear how the inducer of the task-irrelevant perceptual learning (a task target) can evoke sensitivity enhancement.
However, some researchers have tried to explore the properties of the exposure-based perceptual learning using concepts of conditioning. Seitz and Watanabe (2003) have suggested, for example, that successful performance of a given task could generate an internal reward that works like an external reward in reinforcement learning.
To test this hypothesis, attempts have been made to explore the connectivity of perceptual learning and reward. A recent study has shown this relation directly (Kim, Seitz and Watanabe, 2007; Seitz, Kim and Watanabe, in press). During the training session, subjects passively viewed (in other words, without being given any specific task) two differently oriented sinusoidal gratings. Whenever one grating with a specific orientation was presented, a small amount of water was delivered through a tube placed in their mouths, while no water was offered when the grating with the other orientation was presented. Because the subjects had been deprived of food and water for 5 hours before the experiment, it was assumed that the water would work as an external reward. The results showed the close relation between an external reward and perceptual learning. Namely, the sensitivity to the orientation paired with water drops was increased, relative to the sensitivity to the other orientation that was not paired with any external reward.
While this result does not directly verify the hypothesis that processing of a target works as a reward to form task-irrelevant learning, it suggests that a reward influences what stimulus is learned or not.
3. Subliminal visibility of task-irrelevant stimulus
The aforementioned two conditions about selectivity of task-irrelevant perceptual learning are linked to factors outside of the learned stimulus, rather than properties of the learned stimulus itself. Focus on the outside factors may reflect the characteristics of exposure-based learning. First, exposure-based learning is passive, and thus requires no effort by subjects on the learned stimulus. Second, learning occurs even when subjects cannot perceive the learned stimulus. Third, learning of an unattended stimulus depends on the feature the stimulus is paired with. Previous studies have shown sensitivity enhancement in the exposed stimulus, but no enhancement for the stimulus paired with a distractor.
A recent study, however, showed that properties of an irrelevant stimulus could determine whether learning could occur or not. Tsushima, Seitz and Watanabe (2008) explored the effect of visibility of the task-irrelevant stimulus on the task-irrelevant perceptual learning by varying the coherent ratio of the irrelevant motion that was employed in previous perceptual learning studies (e.g. Seitz and Watanabe, 2003; Watanabe, Náñez and Sasaki, 2001). It might be predicted, intuitively, that the motion stimulus with a higher coherence level would evoke a greater learning effect, since it would naturally be assumed that a stimulus with a stronger signal should induce a stronger learning signal. Surprisingly, the results contradicted such predictions. The learning took place for the relatively low coherent motion stimuli (5% and 15%), but did not take place for the relatively high coherent motion stimulus (50%), which was obviously suprathreshold.
Even though this result initially contradicted expectations, it was consistent with a previous study that observed higher activity in the visual cortex with the subthreshold motion as compared to the suprathreshold motion (Tsushima, Sasaki and Watanabe, 2006). These seemingly counterintuitive results may be because the human lateral prefrontal cortex (LPFC), which plays an important role in the inhibitory control of irrelevant stimulus, has a higher threshold for identifying the coherent motion direction in comparison to the human homologues of the middle temporal area (MT+) that is highly responsive to motion signals. The suprathreshold task-irrelevant motion display activates the LPFC so that the activation of MT+ is inhibited. However, when the task-irrelevant motion display has a sub- or parathreshold coherent level such as 5% or 10% coherency, the LPFC fails to notice this motion. Thus, MT+ can be activated without its inhibitory processing.
Therefore, the finding that task-irrelevant learning occurs only when the irrelevant feature is weak is also consistent with the intuition that stimuli with the stronger signal evoke the stronger learning signal. While the signal may not be strong in our perceptual experience, it is processed as a stronger signal in the brain.
RECONCILING OPPOSITE FINDINGS RELATED TO TASK-IRRELEVANT LEARNING
These results indicate that even in cases of task-irrelevant learning the brain has its own conditions for determining which stimuli trigger learning. At the same time, these conditions can reconcile controversies over whether perceptual learning without attention is possible or not.
As mentioned earlier, many studies suggested that focused attention to a feature is necessary for the feature to be learned, by showing that unattended features during training were not learned. Even though a series of experiments had repeatedly confirmed task-irrelevant perceptual learning, it was still necessary to explain why many previous studies had failed to find perceptual learning without attention. The aforementioned conditions thus provide insight into a plausible explanation and, when applied, can determine whether task-irrelevant learning would be possible under a given situation.
For example, in the study by Ahissar and Hochstein (1993) where learning of a task-irrelevant stimulus was not observed, two different orientations (vertical or horizontal) of the global texture array (task-irrelevant stimulus) were paired with a target present display and a target absent display with equal probability. If the target worked as an internal reward, this probability would be problematic in producing learning. It has been suggested that conditioning is dependent on the relative probability of the appearance of an unconditioned stimulus in the presence of a conditioned stimulus. When this probability is high, excitatory conditioning occurs; when this probability is low, inhibitory conditioning occurs. The equal probability situation of the Ahissar and Hochstein study, therefore, created a situation where no learning could occur. In addition, the task-irrelevant stimulus which was employed in their studies prevented learning from occurring because the stimulus strength was over the threshold and an attentional mechanism might have inhibited the stimulus. Since task-irrelevant perceptual learning did not occur with suprathreshold stimuli, the results of their experiment were not inconsistent with the observations of studies which have shown the successful task-irrelevant perceptual learning.
A UNIFIED MODEL FOR PERCEPTUAL LEARNING
Thus far, we have explored certain conditions under which task-irrelevant perceptual learning can occur. This type of perceptual learning initially appeared to be incompatible with the results of previous studies that task-irrelevant features were not leaned just as a result of exposure. The aforementioned new findings (e.g. Kim, Seitz and Watanabe, 2007; Seitz, Kim and Watanabe, in press; Seitz et al., 2005; Seitz and Watanabe, 2003; Tsushima et al., 2008), however, have suggested the model that bridges the gap between both lines of studies. This may lead one to ask if such reconciliation can suggest that both types of learning occur by a single underlying mechanism.
Few attempts were made to explain both types of learning (task-relevant and task-irrelevant) with a single model. The role of attention (the most critical factor) is different between the two learning types: in task-relevant learning, attention to a feature is a key element in generating perceptual learning of the feature, while in task-irrelevant learning attention to a feature is unnecessary for the feature to be leaned. This has made it difficult to compromise both types into a single model. Another problem faced when attempting to build a single model is that task-irrelevant perceptual learning has not been observed in all studies. The inconsistency has therefore led to controversies over the existence of task-irrelevant perceptual learning. Due to these difficulties in combining the two types of learning into one model, task-irrelevant learning has, more often than not, been classified as an exceptional case.
The results of recent studies have suggested, however, that the differences between the two learning types might not be as large as expected, especially with regards to the involvement of attention. Although the occurrence of task-irrelevant learning indicated that the learning of a stimulus occurs without focused attention to the stimulus, it did not rule out the possibility that attention is not involved. The necessity of synchronization between presentation of an attended target and a learned stimulus indicates that attention to some stimulus or location in the visual field may be necessary for a stimulus to be learned.
As aforementioned in the previous section, recent studies provided possible reasons as to why learning of an unattended feature did not occur in every study. Learning does not occur for all unattended stimuli, but rather occurs only when some conditions are fulfilled. Thus, the incongruity of findings related to the task-irrelevant learning cannot suffice as evidence against the existence of the task-irrelevant learning.
These findings have led to the development of a model that may explain both task-relevant and task-irrelevant perceptual learning. Seitz and Watanabe (2005) proposed a unified model based on concepts taken from reinforcement learning and conditioning. In this model, perceptual learning occurs through the coincidence of diffusive signals driven by a task activity (reinforcement signals) and signals induced by the presentation of a stimulus (stimulus-driven signals).
The key idea is that the task activities evoked reinforcement signals, which caused learning. It has been suggested that the task activities, such as detecting and identifying a target, might function as an internal reward. This idea was supported by previous studies showing internal motivation to be regarded as an important reinforcer and that successful performance of a task worked as an internal reward (Herzog and Fahle, 1999; Karni and Bertini, 1997).
When subjects identify a target successfully, reinforcement signals spatially diffuse to trigger perceptual learning of the stimulus exposed at that moment. According to this schema, learning always occurs for a task-relevant stimulus when a target is detected, since the target evokes both the reinforcement and stimulus-driven signals. On the other hand, learning occurs for a task-irrelevant stimulus only when the stimulus is presented with the diffusion of reinforcement signals.
This model includes subsystems of attention that provides an explanation for how attention is involved in perceptual learning. Recent studies have suggested that there are three independent subsystems of attention, each having different effects on stimulus processing. Posner and Petersen (1990), for example, suggested three subsystems – the alerting, orienting, and executive function. The alerting system controls a non-specific arousal state; the orienting system directs resources to a specific spatial cue or feature; the executive function system helps solve task-involving conflicts. In the model by Seitz and Watanabe (2005), the processing of a target is facilitated by the orienting system, which directs attentional resources to the location of the target when subjects are performing a task. The alerting system is activated by processing of a target and releases diffusive signals. Thus, in this model, the alerting system and reward system work in the same way to trigger diffusive reinforcement signals (see Figure 2).
Figure 2.
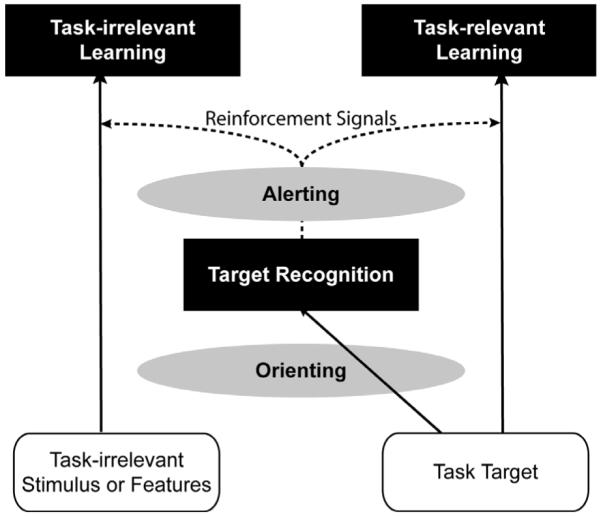
Depiction of a unified model of perceptual learning proposed by Seitz and Watanabe (2005). Successful task performance due to focused attention (by the orienting system) on task targets leads to diffusive reinforcement signals, which trigger perceptual learning to occur for the stimulus exposed at that moment, regardless of whether the stimulus is task relevant or not.
Therefore, according to the principles of this model, task-irrelevant perceptual learning is not necessarily regarded as perceptual learning which occurs without attention. Despite the lack of oriented (focused) attention to task-irrelevant stimulus, learning still occurs via the alerting attention system. Thus, it can be said that attention plays a role in both the task-irrelevant and task-relevant perceptual learning.
CONCLUSION
This article focuses on the selection process of the brain when undergoing task-irrelevant learning. Unattended stimuli can be learned when synchronized with processes of a target, reinforced by an internal or external reward and when the stimuli are weak or subthreshold. This perspective not only gives rise to better understanding of task-irrelevant learning, but also resolves controversies based on observations related to the learning of unattended stimuli.
ACKNOWLEDGEMENT
This work was supported by NIH R01EY015980-04A2 and R21EY018925.
REFERENCES
- Ahissar M, Hochstein S. Attentional control of early perceptual learning. The Proceedings of the National Academy of Science USA. 1993;90:5718–5722. doi: 10.1073/pnas.90.12.5718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fahle M. Perceptual learning: specificity versus generalization. Current Opinion in Neurobiology. 2005;15:154–160. doi: 10.1016/j.conb.2005.03.010. [DOI] [PubMed] [Google Scholar]
- Fiorentini A, Berardi N. Perceptual learning specific for orientation and spatial frequency. Nature. 1980;287:43–44. doi: 10.1038/287043a0. [DOI] [PubMed] [Google Scholar]
- Grossberg S. How does a brain build a cognitive code? Psychological Review. 1980;87:1–51. doi: 10.1007/978-94-009-7758-7_1. [DOI] [PubMed] [Google Scholar]
- Herzog MH, Fahle M. Effects of biased feedback on learning and deciding in a vernier discrimination task. Vision Research. 1999;39:4232–4243. doi: 10.1016/s0042-6989(99)00138-8. [DOI] [PubMed] [Google Scholar]
- Karni A, Bertini G. Learning perceptual skills: Behavioral probes into adult cortical plasticity. Current Opinion in Neurobiology. 1997;7:530–535. doi: 10.1016/s0959-4388(97)80033-5. [DOI] [PubMed] [Google Scholar]
- Kim D, Seitz A, Watanabe T. Effect of reward on perceptual learning [Abstract] Journal of Vision. 2007;7(9):85, 85a. http://journalofvision.org/7/9/85/, doi:10.1167/7.9.85. [Google Scholar]
- Posner MI, Petersen SE. The attention system of the human brain. Annual Review of Neuroscience. 1990;14:340–347. doi: 10.1146/annurev.ne.13.030190.000325. [DOI] [PubMed] [Google Scholar]
- Schoups A, Vogels R, Qian N, Orban G. Practising orientation identification improves orientation coding in V1 neurons. Nature. 2001;412:549–553. doi: 10.1038/35087601. [DOI] [PubMed] [Google Scholar]
- Seitz A, Lefebvre C, Watanabe T, Jolicoeur P. Requirement for high-level processing in subliminal learning. Current Biology. 2005;15:R753–R755. doi: 10.1016/j.cub.2005.09.009. [DOI] [PubMed] [Google Scholar]
- Seitz A, Watanabe T. Psychophysics: Is subliminal learning really passive? Nature. 2003;422:36. doi: 10.1038/422036a. [DOI] [PubMed] [Google Scholar]
- Seitz A, Watanabe T. A unified model for perceptual learning. Trends in Cognitive Sciences. 2005;9(7):329–334. doi: 10.1016/j.tics.2005.05.010. [DOI] [PubMed] [Google Scholar]
- Seitz A, Kim D, Watanabe T. Rewards evoke learning of unconsciously processed visual stimuli in adult humans. Neuron. doi: 10.1016/j.neuron.2009.01.016. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shiu LP, Pashler H. Improvement in line orientation discrimination is retinally local but dependent on cognitive set. Perception and Psychophysics. 1992;52:582–588. doi: 10.3758/bf03206720. [DOI] [PubMed] [Google Scholar]
- Sowden PT, Davies IR, Roling P. Perceptual learning of the detection of features in X-ray images: A functional role for improvements in adults’ visual sensitivity? Journal of Experimental Psychology: Human Perception and Performance. 2000;26(1):379–390. doi: 10.1037//0096-1523.26.1.379. [DOI] [PubMed] [Google Scholar]
- Spanis CW, Squire LR. Stability of long temporal gradients of retrograde amnesia in mice. Behavioral and Neural Biology. 1987;48:237–245. doi: 10.1016/s0163-1047(87)90794-1. [DOI] [PubMed] [Google Scholar]
- Tsushima Y, Sasaki Y, Watanabe T. Greater disruption due to failure of inhibitory control on an ambiguous distractor. Science. 2006;314:1786–1788. doi: 10.1126/science.1133197. [DOI] [PubMed] [Google Scholar]
- Tsushima Y, Seitz A, Watanabe T. Task-irrelevant learning occurs only when the irrelevant feature is weak. Current Biology. 2008;18(12):R516–R517. doi: 10.1016/j.cub.2008.04.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe T, Náñez JE, Koyama S, Mukai I, Liederman J, Sasaki Y. Greater plasticity in lower-level than higher-level visual motion processing in a passive perceptual learning task. Nature Neuroscience. 2002;5:1003–1009. doi: 10.1038/nn915. [DOI] [PubMed] [Google Scholar]
- Watanabe T, Náñez JE, Sasaki Y. Perceptual learning without perception. Nature. 2001;413:844–848. doi: 10.1038/35101601. [DOI] [PubMed] [Google Scholar]