

Abstract
Background
Rab GTPases are regulators of intracellular membrane traffic. The Rab27 subfamily consists of Rab27a and Rab27b. Rab27a has been recently implicated in Griscelli Disease, a disease combining partial albinism with severe immunodeficiency. Rab27a plays a key role in the function of lysosomal-like organelles such as melanosomes in melanocytes and lytic granules in cytotoxic T lymphocytes. Little is known about Rab27b.
Results
The human RAB27B gene is organised in six exons, spanning about 69 kb in the chromosome 18q21.1 region. Exon 1 is non-coding and is separated from the others by 49 kb of DNA and exon 6 contains a long 3' untranslated sequence (6.4 kb). The mouse Rab27b cDNA shows 95% identity with the human cDNA at the protein level and maps to mouse chromosome 18. The mouse mRNA was detected in stomach, large intestine, spleen and eye by RT-PCR, and in heart, brain, spleen and kidney by Northern blot. Transient over-expression of EGF-Rab27b fusion protein in cultured melanocytes revealed that Rab27b is associated with melanosomes, as observed for EGF-Rab27a.
Conclusions
Our results indicate that the Rab27 subfamily of Ras-like GTPases is highly conserved in mammals. There is high degree of conservation in sequence and gene structure between RAB27A and RAB27B genes. Exogenous expression of Rab27b in melanocytes results in melanosomal association as observed for Rab27a, suggesting the two Rab27 proteins are functional homologues. As with RAB27A in Griscelli Disease, RAB27B may be also associated with human disease mapping to chromosome 18.
Background
Rab proteins constitute a large group within the Ras superfamily of low molecular weight GTPases. Rab GTPases regulate vesicular transport steps in endocytic and exocytic pathways, acting as molecular switches oscillating between active GTP-bound and inactive GDP-bound states [1,2]. Over 50 different Rab proteins have been identified to date in mammalian cells while the yeast S. cerevisae has 11 Rabs (termed Ypt and Sec4) [3,4]. Within this family of proteins, most members share about 35% identity, while there are some Rabs that show unusually high identity (more than 70%) and are considered isoforms, eg, Rab1a and Rab1b.
The Rab27 subfamily consists of Rab27a (previously designated Ram) [5] and Rab27b (previously designated c25KG) [6]. Rab27a was initially cloned from a megakaryocyte library and subsequent studies revealed that it is expressed in haemopoietic-derived cells, melanocytes in skin and eye, lung, pancreas and intestine [5,7,8]. Other tissues such as liver, kidney, brain and muscle show little if any expression of Rab27a. In a previous study, we reported a characterization of the human RAB27A gene [9]. The RAB27A gene comprises five coding exons and two non-coding exons, of which one is alternatively used, and spans approximately 65 kb of DNA. RAB27A was mapped to chromosome 15q21 by fluorescence in situ hybridization and radiation hybrid mapping.
More recently, RAB27A was identified as one gene responsible for Griscelli Disease [10]. Griscelli Disease is caused by two unrelated genes coincidentally mapping very close to each other at the 15q21 locus, RAB27A and MYO5A [11,12]. Griscelli Disease is a rare autosomal disorder characterized by partial albinism, variable cellular immunodeficiency and an acute phase of uncontrolled T lymphocyte and macrophage activation. In some cases, a neurological defect rather than immunological disease prevails. It appears that Griscelli Disease associated with immunological disease is caused by mutations in RAB27A, while Griscelli Disease associated with neurological disease is caused by mutations in MYO5A [10]. Mouse models for Griscelli Disease already exist and they are the result of mutations in Rab27a (ashen mouse, ash) or MyoVa (dilute mouse, d) [13,14].
Griscelli disease and related diseases such as Hermansky-Pudlak and Chediak-Higashi syndromes involve defects in intracellular sorting, maturation and/or secretion of specialized lysosome-like organelles. These have also been called "secretory" lysosomes and examples include lytic granules in T lymphocytes, dense platelet granules, and melanosomes in melanocytes [15]. Our recent studies suggest that Rab27a plays a critical role in the biology of secretory lysosomes. In melanocytes, Rab27a seems to be required with Myosin Va for the peripheral capture of melanosomes [16]. In cytotoxic T lymphocytes, Rab27a is required for the secretion of lytic granules and thus for target cell lysis [10,17].
Much less is known about Rab27b. Rab27b was originally purified from platelets and cloned from melanoma mRNA by RT-PCR [6,8]. As a first step towards the study of RAB27B and its possible involvement in human disease, we report here a characterization of the human and murine Rab27b genes.
Results
Gene Structure of the human RAB27B gene
To study the gene structure of RAB27B, three P1 clones containing inserts of 90 kb (designated gs15454), 86 kb (gs15455) and 97 kb (gs15502) respectively, were isolated. Hybridization of the RAB27B cDNA (U57093) probe to DNA digested with Eco RI, Hind III and Bam HI gave the same pattern of bands for genomic and P1 DNA indicating that RAB27B is a single copy gene and that the P1 clones contain this gene. The sizes of the restriction fragments hybridizing to each exon are given in Table 1. The exon/intron boundaries were determined by sequencing following subcloning of four Eco RI fragments containing the exons. The exon/intron boundaries are given in Table 2. In Table 3, we present the sequence of pairs of oligonucleotides that will amplify each of the coding exons of RAB27B from genomic DNA and therefore may be used for future mutation analysis.
Table 1.
Restriction fragment sizes hybridising to each exon
| RAB27B | Eco RI | Kpn I | Nde I | Nco I | Sac I | Sma I | Spe I |
| exon (no.) | |||||||
| 1 | 7 | 15 | 8.5 | 10 | 20 | 8.5 | 8 |
| 2 | 5.5 | 14 | 10 | 9 | 15 | 45 | 11 |
| 3 | 5.5 | 14 | 10 | 9 | 15 | 45 | 11 |
| 4 | 4.5 | 14 | 7.5 | 9a | 15 | 45 | 11 |
| 5 | 4.5 | 13 | 4 | 8 | 15 | 45 | <4 |
| 6 | 1.2 | 13 | 4 | 8 | 15 | 45 | <4 |
a There is a Nco I site within exon 4, and this Nco I fragment contains only the 5' end of exon 4 along with part of intron 1 and exons 2 and 3.
Table 2.
Exon/intron organisation of human RAB27B gene
| RAB27B | Exon | Coded | Intron | Aminoacid(s) | |
| exon (no.) | size | aminoacid | 5' and 3' end sequence of exon | size | interrupted |
| (bp) | (no.) | (kb) | |||
| 1 | 71 | cgggagttccagGGCTTGGGAGG... | >30 | ||
| ...CTTCGCAGGCTGgtgagttgggaa | |||||
| 2 | 172 | 51 | tctcctatacagACCGACCAAGAC... | 1.6 | Val51/Val52 |
| ...GAAAAACGTGTGgtgagtttttaa | |||||
| 3 | 86 | 28 | tgtatctttcagGTTTATAATGCA... | 5.3 | Arg80 |
| ...GGGACAAGAGCGgtaatagtaaat | |||||
| 4 | 104 | 34 | tttccttggcagGTTCCGGAGTCT... | 3.5 | Ser115 |
| GAAACTGGATGAgtaagtgggact | |||||
| 5 | 124 | 40 | tttcttttcaagGCCAACTGCAAG... | 1.1 | Gly156 |
| ...TGACAAATATGGgtaagtcagtta | |||||
| 6 | 6,624 | 63 | tttctatgctagCATACCATATTT... |
Table 3.
Primers for amplification ofRAB27B coding regions
| RAB27B | PCR fragment | Sense primer | Antisense primer |
| exon (no.) | size (bp) | ||
| 2 | 252 | 5'-actttaatcaaaacatacttgttttcc | 5'-gggatgagtgacaaggttagaagtacg |
| 3 | 192 | 5'-tgagtgctgacaaaatccaaagcaacc | 5'-agtaagtttttctaagctatgtgtagc |
| 4 | 209 | 5'-tgtggactaactttcagaactaccc | 5'-aacaagcacctaaagcagttgagt |
| 5 | 237 | 5'-tttatcaataacttgctggttccat | 5'-atctaatgtctctgctccatttgt |
| 6 | 252 | 5'-tcaaaaatatttgccatgctttcta | 5'-gggttcttgatgttcagtttctatg |
Extensive Southern analysis with a variety of enzymes and probes specific for each exon allowed the exons to be mapped relative to each other (Fig. 1). Exons 2, 3, 4, 5 and 6 could all be accurately mapped in relation to each other and lie within about 19 kb and our studies suggested that exon 1 was located at least 30 kb from exon 2. While this work was in progress, sequence from two BACs containing the human RAB27B gene were deposited in the database of high throughput genomic sequence (NCBI). The genomic sequences have the accession numbers AC007673 and AP001910. The BAC AC007673 contains exon 1 (55,377-55,154), exon 2 (6,287-6,116) and exon 3 (4,485-4,400). The exons 4 and 6 are in the BAC AP001910 (96,303-96,200 and 116,297-115,786) but the partial sequences of this BAC are unordered. The genomic sequence for exon 5 was not found in the AP001910 sequence probably due to the sequence of this BAC being incomplete.
Figure 1.
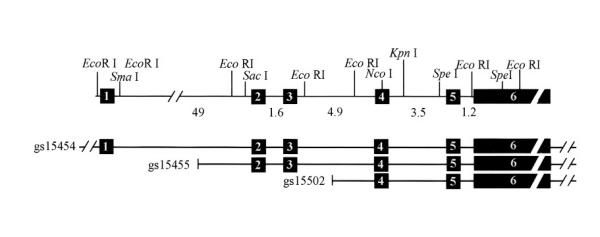
Restriction map of RAB27A and neighboring genomic sequences. The map at the top shows the RAB27B gene structure with some restriction sites. The numbers between exons represent the sizes of each intron and the black boxes represent the exons. The three lines below represent the P1 clones.
Northern blot analysis by Chen et al. [8] showed the most abundant transcript in most tissues to be of approximately 8 kb which is much larger than the 1.026 kb cDNA (U57093) reported by the same authors. The cDNA was cloned by RT-PCR of the coding region followed by 5' and 3' race and it is unlikely that the 3' race would have extended through a large 3' region of several kilobases. Database mining allowed us to identify the full 3' region of the mRNA. Using the U57093 sequence, we searched the human EST database and identified 9 ESTs; AA588753, AI432357, AA812916, AA846320, AW664866, AW370404, AW592937 (these 7 ESTs make up UniGene Hs.97189), BF088963 and BF243879. The most 3' of these, AW592937, did not bring up any more ESTs. However, searching the EST database with the corresponding region of the AP001910 BAC sequence identified a further 31 ESTs which are probably located in a single 6.6 kb 3' exon. Ten of these ESTs (BF438386, AI732288, AI798733, AW468475, AI473744, AW242160, AI277199, BF216951, R39044, BF002851) share the same 3' end and none extend beyond this point, making it highly likely that this is the most frequently used 3' end of the RAB27B mRNA. The other ESTs are; AA659732, AA621327, AA386051, AV749183, AA846320, AA580411, BF245563, BE878236, BF446484, BF246274, BF218405, AI697958, BF245714, BF216086, BF245940, AW340618, H25594, R69522, BF246185, R69389 and BE697849. Most of these ESTs fall within an anonymous 2.586 kb cDNA in Genbank called AF131784 [18]. The sequences from our gs15454, ESTs, AF131784, and AP001910 have been combined to compile a 7.3 kb cDNA, which may represent the complete cDNA for RAB27B. The compiled sequence is available at our laboratory web site (http://www.med.ic.ac.uk/divisions/4/hrab27b.txt or http://www.med.ic.ac.uk/divisions/4/seabra_research.asp)
Figure 1 shows the distribution of the exons determined by restriction mapping and sequence analysis, selected Eco RI, Sma I, Sac I, Nco I, Kpn I and Spe I sites present and the extent of each P1 clone are shown. P1 clone gs15454 contains all 6 exons and at least 3.5 kb of DNA 5' of exon 1. P1 clone gs15455 contains exons 2 to 6 and P1 clone gs15502 contains exons 4 to 6. The RAB27B gene spans about 69 kb with the untranslated exon 1 separated from exon 2 by 49 kb. Translation terminates approximately 200 bp into the very large exon 6, which is about 6.6 kb in size.
An EST found in GenBank (BF243879) contains an extra 152 bp of sequence in the 5' UTR, which is not present in the cDNA reported by Chen [8]. This extra sequence is present in a BAC (accession no. AC007673) and in our P1 clone (gs15454), is probably part of exon 1 and was therefore included in the compiled cDNA sequence. Two kb upstream of the start of transcription (accession no. AC007673) was analysed using MatInspector/TRANSFAC (http://www.hgsc.hcm.tmc.edu). Numerous potential transcription-factor binding sites can be found in the possible promoter region. This region contains 41% of C+G and the promoter prediction by neural network (http://www.hgsc.hcm.tmc.edu) shows three possible promoter sequences (at 13,104; 13,357 and 14,780) and two for the reverse sequence (at 14,546 and 13,542) in the BAC (accession no. AC007673).
cDNA cloning and expression of the murine Rab27b gene
In order to obtain a cDNA encoding mouse Rab27b, we attempted RT-PCR from different mouse tissues using primers derived from the known human sequences around the start and stop codons. We successfully amplified a fragment from intestinal RNA which was then subcloned and sequenced. The fragment showed 89% and 94% identity at the DNA and amino acid levels respectively with human RAB27B, and only 67% and 71% identity at the DNA and amino acid levels respectively with RAB27A. BLAST searching the mouse EST database identified one IMAGE clone (Genbank AI317047) that is identical to our sequence. Further sequencing of this clone showed that it spans the RAB27B cDNA coding region. We concluded that the sequence represents the mouse ortholog of RAB27B and was deposited in Genbank under accession number AF328893.
Searching the mouse EST database with the corresponding 3' end region of the AI317047 sequence identified a further 33 ESTs (AI643928, BB559600, BB316219, BB378261, AV376027, BB549470, BB254935, BB317086, BB183652, BB147117, BB110158, BB319088, BB112138, AV086385, BB268115, BB267030, AV330192, BB128597, BB005939, AV227565BB106617, AV371120, BB524432, BB263040, AV360152, BB144635, BB149815, AV229102, BB008967, BB273472, BB311744, AV226744 and BB535273) sharing the same 3' end and none extend beyond this point, making it highly likely that this is the most frequently used 3' end of the mouse Rab27b mRNA.
RT-PCR also allowed us to also isolate a fragment spanning the coding region of the mouse Rab27a cDNA. The sequence shows 88% and 96% identity at the DNA and amino acid levels respectively with human RAB27A, and only 67% and 72% identity at the DNA and amino acid levels respectively with RAB27B. We concluded that this sequence represents the mouse ortholog of RAB27A. While this work was in progress, a mouse cDNA for Rab27a was deposited in GenBank under accession number AF304376. Searching the mouse EST database with the mouse Rab27a identified 5 ESTs (BE369888, AI036507, AA204414, BE573054 and BE226035). A comparison between human and mouse Rab27a and Rab27b amino acid sequences reveals extensive sequence conservation across mammals (Fig. 2).
Figure 2.
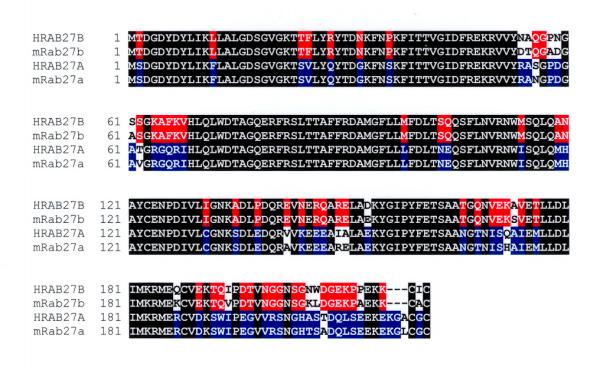
Protein sequence alignment between human RAB27A, mouse Rab27a, human RAB27B and mouse Rab27b.
We then compared the expression pattern of Rab27a and Rab27b in the mouse. We first conducted extensive RT-PCR studies and we observed that Rab27a mRNA appears more abundant than Rab27b in every one of over 10 different tissues studied (Fig. 3). Rab27b mRNA could only be amplified from stomach, large intestine, spleen and eye. In cultured mouse melanocyte cell lines, melan-a, melan-b and melan-c, we observe only Rab27a expression (Fig. 3). Then we performed Northern analysis. mRNA from various mouse tissues revealed a 2.5 kb Rab27b message which may correspond to our 2.47 kb cDNA and also an 8 kb message present in lower levels that appears similar in size to the prevailing message in Homo sapiens. Rab27b mRNA is detectable in heart, brain, spleen and kidney, and at low levels in lung and liver (Fig. 4). A 3.5 kb Rab27a message was detected in heart, spleen, lung and kidney, and faintly in brain, liver and testis (Fig. 4).
Figure 3.

Expression of Rab27a and Rab27b in mouse tissues and melanocyte cell lines. Rab27a and Rab27b specific primers were used to amplify fragments of 445 bp and 550 bp respectively from the indicated mouse tissues (A) and melanocyte cell lines (B), as described in Materials and Methods. The PCR products were analysed on an agarose gel stained with ethidium bromide. The arrows on the left-hand side of the figure indicate positions of DNA marker sizes. Hprt was used as an internal control. Reaction performed without reverse transcriptase did not produce any detectable product.
Figure 4.
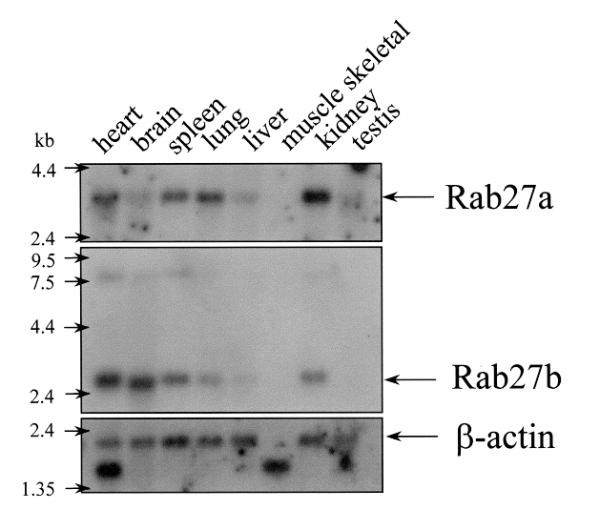
Northern blot analysis of expression of Rab27a and Rab27b in mouse tissues. Rab27a and Rab27b-specific probes were used as described in Materials and Methods. The arrows on the left-hand side of the figure indicate positions of the RNA marker sizes. β-actin was used as a loading control.
Mapping the human and mouse Rab27b genes
Radiation hybrid mapping with the GeneBridge 4 Radiation Hybrid Panel indicated that RAB27B is located on chromosome 18. More precisely, RAB27B was found to map 9.5 cR below marker WI-4115 and 3.9 cR above marker AFM3577TD5. These and surrounding markers have been localised to 18q12.3-21.1. For example, marker WI-8740 has been placed between D18S474 and D18S64 (71.3-83.0 cM) on the GB4 RH map. To confirm the localization obtained by radiation hybrid mapping, we performed FISH analysis using P1 clone gs15454 as a probe on cells from a patient containing a deletion in chromosome 18 (Fig. 5) [19]. We observed in all cases only one signal with the RAB27B probe while a chromosome 18 centromere probe revealed two signals in almost all metaphases. Therefore RAB27B appears to be located on chromosome 18 and lies within the region q21.1-qter, which is deleted in this patient. The combined mapping results strongly suggest that RAB27B is located on 18q21.1.
Figure 5.
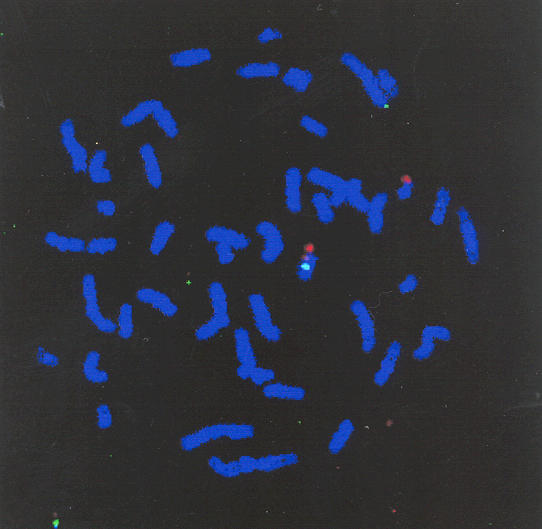
Chromosomal localization of human RAB27B locus. A P1 clone containing RAB27B was used to probe by FISH as described under Materials and Methods human lymphocyte metaphase spreads from a patient containing a deletion in chromosome 18 [24].
Syntenic conservation between human chromosome 18q21.1 and the murine genome suggested that Rab27b is located on either mouse chromosome 1 or 18 (http://www.ncbi.nlm.nih.gov/Homology/). We used the cDNA sequence to design oligonucleotides for radiation hybrid mapping in the mouse genome. Using the Mouse/Hamster Radiation Hybrid panel, we mapped Rab27b to chromosome 18 between markers D18Mit33 and D18Mit141, more precisely 5.6 cR from D18Mit33 (lod>3.0).
A BAC (AP001910) that contains the 3' end of RAB27B, also contains an additional gene called CTCL tumor antigen se57-1 [20]. This corresponds to the cDNA AF273051 in Genbank and 10 sequences in the EST database; AI800280 (this is UniGene cluster Hs.208095), H02496, AI823801, AW148635, N51464 (these 4 are UniGene cluster Hs.30315), N53604, AI767468, AI350671, and H02495, AW600293. It consists of at least 11 exons and is largely included in the BAC except its 5' most 70 bp. It is probably the next gene 3' of RAB27B but its distance and relative orientation is not clear as the BAC sequence is not complete.
Subcellular distribution of Rab27b
We recently studied Rab27a subcellular distribution in melanocytes [16]. We showed that Rab27a is associated with melanosomes, the specialized pigment-containing organelle in melanocytes. The similarity of Rab27a and b isoforms led us to hypothesize that Rab27a and b isoforms may fulfill similar functions. As a first step to test this possibility, we compared the subcellular location of over-expressed GFP-fusion proteins (EGFP-Rab27a and EGFP-Rab27b) in cultured mouse melanocyte cell line melan-c (Fig. 6) [21]. We found that both EGFP-Rab27a (Fig. 6A) and EGFP-Rab27b (Fig. 6D) exhibit a similar distribution pattern. In particular, both fusion proteins are found to be associated with 0.5 μm cytoplasmic structures distributed at the periphery of melanocytes, often in the dendritic processes. Staining of transfected melan-c cells using antibodies reactive to the melanosome resident protein TRP-1 (Fig. 6B,E,H), revealed a high degree of co-localisation between the proteins, suggesting that the structures containing over-expressed EGFP-Rab27a (Fig. 6C) and EGFP-Rab27b (Fig. 6F) are melanosomes. Consistent with melanosomal association of Rab27b with melanosomes in melan-c cells, we also observed association of EGFP-Rab27b with pigment granules in the pigmented melanocyte cell line melan-b (data not shown). The specificity of the association of Rab27 isoforms with melanosomes was shown by transient over-expression of EGFP-Rab1a (Fig. 6G-I), EGFP-Rab5, EGFP-Rab6 and EGFP-Rab8 (data not shown) proteins in these cells. None of these Rabs co-localised with melanosome markers.
Figure 6.
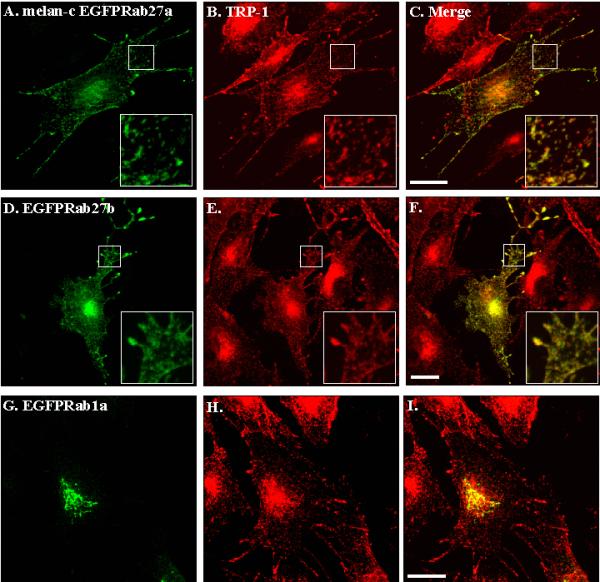
Transiently over-expressed EGFP-Rab27a and EGFP-Rab27b, but not EGFP-Rab1a, associate with melanosomes in melanocytes. Melan-c cells were transfected with pEGFP-Rab27a (A-C), EGFP-Rab27b (D-F) or pEGFP-Rab1a (G-I) and fixed 48 hours later, as described in Materials and Methods. Panels A, D and G show the distribution of the indicated over-expressed EGFP-Rab fusion protein in melan-c cells. B, E and H show the distribution of the melanosomes resident protein TRP-1. C, F and I show the extent of colocalisation of each overexpressed fusion protein with melanosomes. Bars = 20 μm
Discussion
We report here an initial characterization of the Rab27b gene in mammals. The human RAB27B gene is found to consist of 6 exons spread over about 69 kb. The first, non-coding, exon is separated from the others by 49 kb and the end of translation is located within exon 6. This structure is very similar to the human RAB27A gene [9], the exon/intron junctions are present at the equivalent positions and there is high sequence similarity (67% identity at the DNA level and 72% in the protein). It is likely that a duplication event has occurred to generate the two genes encoding the two isoforms of Rab27. Unlike RAB27A however, exon 6 in RAB27B has a very long (6.4 kb) 3' untranslated region but the significance of this finding is unclear at present.
The sequence of the human RAB27B cDNAs and genomic DNA that we have analysed is found to differ slightly from that previously described by Chen et al. [8]. One important difference affects the putative translation of the gene as it occurs in the coding region. We found that codon 206 is TTG (leucine) rather than the reported TGG (tryptophan). We sequenced several independent cDNA clones and genomic clones and our sequence matches five ESTs (Genbank AI432357, AW664866, AA812916, AW370404 and BF088963) in the database that overlap in the region where there is disparity with the Chen et al. [8] sequence.
The mouse rab27b cDNA shows 89% at the DNA and 95% at the protein level with the human RAB27B cDNA. On Northern analysis, the mouse mRNA is only 2.5 kb in size which is considerably smaller than the 8 kb human mRNA. This smaller mRNA size corresponds with the 3' ends determined by a large number of mouse ESTs and is likely due to a much shorter 3' UTR in the mouse compared to the human gene.
Our expression analysis by RT-PCR suggests that Rab27b is expressed at lower levels than Rab27a. However, the PCR was not done under quantitative conditions and this could be the result of different rates of amplification of the different PCR fragments. On the other hand, both mRNAs were detected at approximately the same levels on Northern blots. Our Northern blots of mouse tissues indicated that mouse Rab27a is expressed in heart, spleen, lung and kidney while Rab27b is expressed in heart, brain, spleen and kidney. This distribution is somewhat different from our RT-PCR results for mouse tissues but this could be due to the presence of contaminating blood in the tissues used for analysis as Rab27 proteins are expressed in haemopoietic cell types. It remains to be established whether Rab27b is expressed in melanocytes. The initial cloning of Rab27b by RT-PCR from human melanoma cells [22] would suggest that it is but our RT-PCR analysis on several melanocyte cell lines proved negative. Future studies with specific antibodies should clarify this issue.
The sequences of the human and mouse Rab27a and Rab27b genes confirm the extensive similarity between the two proteins. This suggests that these two Rab27 isoforms may be functional homologues which are active in different tissues. One would then expect the two proteins to be targeted to the same intracellular location in the same cells. We decided to study the subcellular localization of Rab27a and Rab27b in melanocytes as we have recently shown a very specific association of Rab27a with melanosomes, the specialised pigment-containing structures in these cells [16]. Upon expression of GFP-Rab27 fusion proteins in melanocytes, we found that GFP-Rab27b, like GFP-Rab27a, co-localises to melanosomes. This suggests that, although Rab27b does not seem to be expressed in melan-c cells, it contains targeting determinants as present in Rab27a. However, further experiments are required to test whether there is functional redundancy between the two proteins.
Mutations in Rab27a in humans (Griscelli Disease) and mice (ashen) result in melanosome transport and secretion defects [10,13,16]. The lack of phenotype in the other tissues where Rab27a is expressed could be due to functional redundancy between the Rab27a and Rab27b proteins such that there is enough Rab27b in most tissues to complement the lack of Rab27a except in melanocytes where Rab27b was not detectable by RT-PCR.
We had previously implicated Rab27a in the pathogenesis of Choroideremia (CHM), an X-linked slow-onset retinal degenerative disease resulting from loss-of-function mutations in Rab Escort Protein-1 [7]. As the CHM phenotype is very different from Griscelli Disease, it remains unclear whether Rab27a has any role on eye disease. It is unclear at present whether Griscelli patients develop retinal disease as the disease has a poor prognosis and patients don't survive past childhood [12]. Also, the equivalent mutation in mice, ash arose in the C3H strain homozygous for the rd mutation and all C3H mice are blind a few weeks after birth [23]. A likely possibility is that both Rab27 isoforms are partially affected in CHM and therefore contribute, possibly with other Rabs to the initiation and propagation of the degenerative process in CHM.
We show that the human RAB27B gene maps to chromosome 18q21.1 by radiation hybrid mapping and FISH. This region of human 18q21.1 is syntenic with two mouse chromosomal regions in chromosomes 1 and 18. As the leaden (ln) mouse mutant locus maps to mouse chromosome 1 next to Bcl2 (which maps to 18q21.3 in humans), we investigated whether Rab27b could be involved in ln. The ln mutation attracted our attention because ln is phenotypically very similar to ash, which is caused by mutations in Rab27a and d, which is caused by MyoVa mutations [13,14]. We therefore hypothesized that Rab27b was a candidate gene for leaden. However, the mapping of mouse Rab27b to chromosome 18 suggests that Rab27b is not responsible for leaden as leaden maps to chromosome 1. If Rab27a and Rab27b are indeed functional homologues, we would expect Rab27b mutations to cause deficiency in lysosomal transport in tissue(s) other than melanocytes where Rab27b, but not Rab27a, is expressed. The studies presented here will aid the future analysis of a possible association of RAB27B with human disease.
Materials and methods
Isolation of human RAB27B P1 clones
Two DNA oligonucleotides, 5'-TTCGAAGGCTGACCGACCAAGACCATCACT (primer JA158, sense) and 5'-TATAAACCACACGTTTTTCCCGAAAGTCTA (primer JA169, antisense), both corresponding to the 5' end of the gene, amplified a specific 190 bp fragment from human genomic DNA by PCR. The PCR product was verified to correspond to the RAB27B gene by DNA sequencing. These oligonucleotides were used to isolate two P1 clones from a human genomic library through PCR screening in a pyramidal fashion. Colonies containing PCR-positive P1 clones were isolated by filter hybridisation and two P1 clones were obtained and designated gs15454 and gs15455 (Genome Systems). Another set of oligonucleotides, 5'-GTGCAGCAACTGGACAGAATGTGGAGAAAG (primer JA168, sense) and 5'-CTCTGTATATGTCTATATACTCATTTTCTC (primer JA180, antisense) both corresponding to the 3' end of the gene, amplified a specific 460 bp fragment from human genomic DNA by PCR. These oligonucleotides were used as above to isolate one P1 clone designated gs15502.
PCR conditions
PCR amplifications were carried out in a Hybaid-Touch Down thermal cycler (Hybaid). To estimate the sizes of the introns of the RAB27B gene, oligonucleotides were designed and used in PCR reactions to amplify each intron. PCR for introns up to 6 kb in size was carried out in a final volume of 25 μl, containing 10 ng of P1 or 1-2 ng of pBS-Rab27b cDNA clone (Genbank: U57093; a generous gift from Dr. W. Gahl, NIH), 500 nM oligonucleotides and 0.125 units of BioTaq DNA polymerase (Bioline) in a buffer containing 16 mM (NH4)2SO4, 67 mM Tris-HCl (pH 8.8), 0.1% Tween-20, 2.5 mM MgCl2 and each dNTP at 250 μM. PCR reactions were run for 32 cycles of 94ºC denaturing for 1 min, 58ºC annealing for 1 min and 72ºC extension for 5 min, followed by a final elongation step of 72ºC for 10 min. PCR products were analysed on 1% agarose gels after staining with ethidium bromide. Each intron was amplified at least twice from different P1 clones. Intron 1 could not be amplified using this protocol. Its size was estimated by restriction digestion and hybridisation with specific radiolabelled probes as described below.
To generate DNA probes for each exon of RAB27B, oligonucleotides were designed by comparison to RAB27A gene structure and used in PCR reactions to amplify each exon. PCR for exons was carried out in a final volume of 25 μl, containing 10 ng of RAB27B P1 or 1-2 ng pBS-RAB27B clones or pBS-RAB27B cDNA. Exons were amplified in conditions as described above, with an extension time of 1 min. The PCR products were analysed on 2.5% agarose gels stained with ethidium bromide and gel purified using Quick Extract (Qiagen) as described by the manufacturer.
Determination of exon/intron junctions
P1 DNA was isolated using Large-Construct kit (Qiagen). Forty μg of DNA was digested with EcoRI as recommended by the manufacturer (Life Technologies). After digestion, 10 μg of DNA was subjected to standard 0.8% gel electrophoresis. The DNA was transferred onto Hybond N+ nylon membrane (Amersham) by alkaline blotting. Blots were prehybridised in 10 ml of 0.2% bovine serum albumin, 7% SDS, 100 μg/ml sheared denatured salmon sperm DNA, 1 mM EDTA, 0.3 M sodium phosphate buffer (pH 7.4) for 4 hours and then hybridised with RAB27B probes specific for each exon. Hybridisation was carried out at 65ºC overnight in the same buffer. Membranes were washed at 65ºC in 0.25 % SDS and 1 × SSC for 20 min, repeated 4 times and then exposed to Hyperfilm MP (Amersham) in the presence of an intensifying screen. Membranes were stripped at 45ºC twice for 30 min in 0.4 M NaOH, and then incubated for 15 min in 0.1 × SSC, 0.1% SDS, 0.2 M Tris-HCl (pH 7.5). After identification of the bands correspondent to the exons, 30 μg of digested DNA were subjected to standard 0.8% gel electrophoresis. The fragments containing RAB27B exons were isolated, subcloned into pBS (Stratagene) and used to determined the exon/intron junctions by sequencing. Sequencing was performed with an automated ABI 377 DNA sequencer using 1 μg of pBS DNA for each reaction.
Restriction mapping of RAB27B
One microgram of DNA from all P1 clones was singly or doubly digested under conditions recommended by the manufacturer (Life Technologies or New England Biolabs). After digestion, DNA was subjected to standard gel electrophoresis or to PFGE in 0.5x TBE with 5-10 s switching for 16 h. The DNA was transferred onto Hybond N+ nylon membrane (Amersham Pharmacia) by alkaline blotting. Blots were prehybridised and hybridised as described above with theRAB27B cDNA or individual exon probes. After hybridisation to each exon, membranes were stripped and neutralised as described above.
RT-PCR analysis
Total RNA was isolated from eyes using TRIzol® Reagent (Life Technologies). RT-PCR was carried out using 5 μg of total RNA in a reaction volume of 20 μl mixed with 500 ng of oligo(dT)12-18, denatured at 70ºC for 10 min and chilled on ice. The annealed samples were then incubated with 0.5 mM dNTPs, 20 mM Tris-HCl (pH 8.4), 50 mM KCl, 2.5 mM MgCl2, 10 mM DTT and 200 U of SuperScript II reverse transcriptase (Life Technologies) for 1 h at 42ºC, stopped by heating for 15 min at 70ºC and chilling on ice. In order to get a better PCR amplification, the samples were treated with 0.5 U of RNase H for 45 min at 37ºC after reverse transcriptase inactivation. Hprt gene expression was used as an internal control. The specific oligonucleotides for Hprt were 5'-CCTGCTGGATTACATTAAAGCACTG (sense) and 5'-GTCAAGGGCATATCCAACAACAAAC (antisense) [24]. Rab27b expression was detected using the oligonucleotides: 5'-GATGGAGACGATTATCTG (primer JR92, sense) and 5'-TCCATCGACAGTGTCCGGAACC (primer JR94, antisense). Rab27a expression was detected using the oligonucleotides: 5'-GCATTGATTTCAGGGAAAAGAGAG (primer JR62, sense) and 5'-TTCTCCACACACCGCTCCATCCGC (primer JR63, antisense). Amplification products using these oligonucleotides were 350 bp, 445 bp and 550 bp for Hprt, Rab27a and Rab27b, respectively. DNA from two microliters of RT-PCR reaction were amplified using 500 nM oligonucleotides and 0.125 units of BioTaq DNA polymerase (Bioline) in a buffer containing 16 mM (NH4)2SO4, 67 mM Tris-HCl (pH 8.8), 0.1% Tween-20, 2.5 mM MgCl2 and each dNTP at 200 μM. PCR reactions were run for 32 cycles of 94ºC denaturing for 1 min, 58ºC annealing for 1 min and 72ºC extension for 1 min, followed by final incubations at 72ºC for 10 min. PCR products were analysed on 2.5% agarose gels stained with ethidium bromide.
Radiation hybrid mapping
Oligonucleotide primers were designed to the 3' end of the RAB27B cDNA (Genbank: U57093) [8] to amplify a 249 bp product (primer JR135, 5'-GACAGAATGTGGAGAAAGCTGTAGAAACCC corresponding to nucleotide positions 592-621 of the cDNA, and primer JR136, 5'-TGCGTGGTTTACTCAACAACAATTGTGTGG complementary to nucleotide positions 813-842 of the cDNA). PCR was performed as above using DNA from the UK-HGMP subset of the Genebridge 4 Radiation Hybrid DNA Panel [25]. The result (0211202100220000010001001000110001010 11010021000011000110200110000100201101001020001011001000) was submitted to the online radiation hybrid server at The Whitehead Institute for Biomedical Research/MIT Center for Genome Research (http://genome.wi.mit.edu).
In order to map murine Rab27b, oligonucleotide primers were designed to the 3' end of the mouse Rab27b cDNA to amplify a 235 bp product (primer JR100, 5'-CATACCATACTTCGAAACAAGCGCAGC corresponding to nucleotide positions 737-763 of the cDNA; primer JR112, 5'-TCATGGTTTGACATTGTCTCAGGACGTCGG corresponding to nucleotide positions 965-994 of the cDNA). PCR was performed as above using DNA from the Mouse/Hamster Radiation Hybrid Panel (Research Genetics) [26]. The result (000011100000001000100000100110010100220000010 010110000000001100000001000110010101001000110100) was submitted to the online radiation hybrid server at The Whitehead Institute for Biomedical Research/MIT Center for Genome Research (http://genome.wi.mit.edu)
Northern blot analysis
Mouse Multiple Tissue Northern (MTN) Blot containing 2 μg of poly(A)+ RNA per lane was purchased from Clonetech. The blot was hybridized to DNA radiolabelled probes in ExpressHyb buffer (Clonetech) and washed under high stringency conditions (0.2X SSC/0.1% SDS, 50°C). To generate radiolabelled probes for Rab27a and Rab27b transcripts, oligonucleotides were designed and used in PCR reactions to amplify Rab27a and Rab27b fragments using cDNAs as templates. The oligonucleotides used for Rab27a were 5'-ATGTCGGATGGAGATTACGATTACGATTAC (primer JR95, sense) and 5'-TCACTTAGCTGATCCGCAGAGGTA (primer JR96, antisense) and for Rab27b were 5'-GATGGAGACTATGATTATCTG (primer JR92, sense) and 5'-TCCATCGACAGTGTCCGGAACC (primer JR94, antisense). The PCR fragments were gel purified and radiolabelled with [α-32P]dCTP by random oligopriming (Amersham) and used as probes.
Fluorescence in situ hybridization
Cells from a patient with a chromosome 18q deletion [Patient 1 case no. 180569, [46,XY,del(18)(q211-qter)] in [19] were analysed by FISH using a centromere probe for chromosome 18 (Oncor), detected with rhodamine, and P1 clone gs15454 containing the RAB27B gene detected with FITC. Preparation of metaphase spreads, labelling and detection of probes were carried out using standard techniques described previously [27].
Melanocyte culture, transfection and immunofluorescence
The melanocyte cell line melan-c from an albino mouse was kindly provided by Prof. Dorothy Bennett, St George's Hospital Medical School, London, UK. Melan-c cells were cultured in RPMI 1640 supplemented with 10% fetal calf serum, 2 mM glutamine, 0.1 mM 2-mercaptoethanol, 200 nM phorbol 12-myristate 13-acetate (Calbiochem), 100 U/ml penicillin G and 100 U/ml streptomycin at 37°C with 10% CO2. Cells for immunofluorescence were grown on coverslips for 24 h, transfected using the liposomal transfection reagent Fugene 6 (Roche) and then processed as follows. Transfected cells were washed in PBS, permeabilized for 5 min in 80 mM K PIPES, pH 6.8, 5 mM EGTA, 1 mM MgCl2, 0.05% saponin (in order to remove non-membrane associated fusion protein) and then fixed in 3% paraformaldehyde in PBS for 15 min. Excess fixative was removed by extensive washing in PBS and quenched by incubation in 50 mM NH4Cl for 10 min. Fixed cells were then incubated with anti-tyrosinase related protein-1 (TRP-1) monoclonal antibody (ID Labs) diluted 1:50 in buffer (PBS, 0.5% BSA, 0.05% saponin) for 30 min, washed extensively in buffer and incubated for 30 min with Alexa568 conjugated goat anti-mouse secondary antibodies (Molecular Probes) diluted 1:200 in buffer. Coverslips containing fixed cells were washed as before in buffer, mounted in Immuno Fluor medium (ICN) and observed using a Leica DM-IRBE confocal microscope. The plasmid pEGFP-Rab27b was produced by subcloning of the Rab27b cDNA into the pEGFP-C3 vector (Clontech) downstream of the EGFP ORF. The resulting plasmid encodes a fusion protein containing the Rab27b protein, attached to the carboxyl-terminus of EGFP. The production of other plasmids transfected in this study have been described elsewhere [16].
Sequences
We have deposited the following sequences in Genbank; human RAB27B cDNA (accession number AF329499) and mouse Rab27b cDNA (accession number AF328893). All searches described in this work were current as of 10 December 2000.
Acknowledgments
Acknowledgments
We thank Karen Brøndrum-Nielsen, Bill Gahl and Dot Bennett for reagents and Jim Bellingham for initial mapping of RAB27B. This work was supported by the Medical Research Council, the Wellcome Trust, the Foundation Fighting Blindness and the British Retinitis Pigmentosa Society. J. S. R. was supported by a PhD studentship from the Portuguese Government through Fundação Ciência e Tecnologia.
Contributor Information
José S Ramalho, Email: jose.ramalho@ic.ac.uk.
Tanya Tolmachova, Email: t.tolmachova@ic.ac.uk.
Alistair N Hume, Email: a.hume@ic.ac.uk.
Amanda McGuigan, Email: amanda.mcguigan@kcl.ac.uk.
Cheryl Y Gregory-Evans, Email: c.gregory-evans@ic.ac.uk.
Clare Huxley, Email: c.huxley@ic.ac.uk.
Miguel C Seabra, Email: m.seabra@ic.ac.uk.
References
- Chavrier P, Goud B. The role of ARF and Rab GTPases in membrane transport. Curr Opin Cell Biol. 1999;11:466–475. doi: 10.1016/S0955-0674(99)80067-2. [DOI] [PubMed] [Google Scholar]
- Brennwald P. Reversal of fortune. Do Rab GTPases act on the target membrane? J Cell Biol. 2000;149:1–4. doi: 10.1083/jcb.149.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pereira-Leal JB, Seabra MC. The mammalian Rab family of small GTPases: Definition of family and subfamily sequence motifs suggests a mechanism for functional specificity in the Ras super family. J Mol Biol. 2000;301:1077–1087. doi: 10.1006/jmbi.2000.4010. [DOI] [PubMed] [Google Scholar]
- Lazar T, Gotte M, Gallwitz D. Vesicular transport: how many Ypt/Rab-GTPases make a eukaryotic cell? Trends Biochem Sci. 1997;22:468–72. doi: 10.1016/S0968-0004(97)01150-X. [DOI] [PubMed] [Google Scholar]
- Nagata K-I, Satoh T, Itoh H, Kozasa T, Okano Y, Doi T, Kaziro Y, Nozawa Y. The ram : a novel low molecular weight GTP-binding protein cDNA from a rat megakaryocyte library. FEBS Lett. 1990;275:29–32. doi: 10.1016/0014-5793(90)81431-M. [DOI] [PubMed] [Google Scholar]
- Nagata K-I, Itoh H, Katada T, Takenaka K, Ui M, Kaziro Y, Nozawa Y. Purification, identification and characterization of two GTP-binding proteins with molecular weights of 25,000 and 21,000 in human platelet cytosol. J Biol Chem. 1989;264:17000–17005. [PubMed] [Google Scholar]
- Seabra MC, Ho YK, Anant JS. Deficient geranylgeranylation of Ram/Rab27 in choroideremia. J Biol Chem. 1995;270:24420–24427. doi: 10.1074/jbc.270.41.24420. [DOI] [PubMed] [Google Scholar]
- Chen D, Guo J, Miki T, Tachibana M, Gahl WA. Molecular cloning and characterization of Rab27a and Rab27b, novel human Rab proteins shared by melanocytes and platelets. Biochem Mol Med. 1997;60:27–37. doi: 10.1006/bmme.1996.2559. [DOI] [PubMed] [Google Scholar]
- Tolmachova T, Ramalho JS, Anant JS, Schultz RA, Huxley CM, Seabra MC. Cloning, mapping and characterization of the human RAB27A gene. Gene. 1999;239:109–116. doi: 10.1016/S0378-1119(99)00371-6. [DOI] [PubMed] [Google Scholar]
- Ménasché G, Pastural E, Feldmann J, Certain S, Ersoy F, Dupuis S, Wulffraat N, Bianchi D, Fischer A, Le Deist F, de Saint Basile G. Mutations in RAB27A cause Griscelli syndrome associated with haemophagocytic syndrome. Nat Genet. 2000;25:173–176. doi: 10.1038/76024. [DOI] [PubMed] [Google Scholar]
- Pastural E, Barrat FJ, Dufourcq-Lagelouse R, Certain S, Sanal O, Jabado N, Seger R, Griscelli C, Fischer A, de Saint Basile G. Griscelli disease maps to chromosome 15q21 and is associated with mutations in the myosin-Va gene. Nat Genet. 1997;16:289–292. doi: 10.1038/ng0797-289. [DOI] [PubMed] [Google Scholar]
- Pastural E, Ersoy F, Yalman N, Wulffraat N, Grillo E, Ozkinay F, Tezcan I, Gediköglu G, Philippe N, Fischer A, de Saint Basile G. Two genes are responsible for Griscelli syndrome at the same 15q21 locus. Genomics. 2000;63:299–306. doi: 10.1006/geno.1999.6081. [DOI] [PubMed] [Google Scholar]
- Wilson SM, Yip R, Swing DA, O'Sullivan N, Zhang Y, Novak EK, Swank RT, Russell LB, Copeland NG, Jenkins NA. A mutation in Rab27a causes the vesicle transport defects observed in ashen mice. Proc Natl Acad Sci USA. 2000;97:7933–7938. doi: 10.1073/pnas.140212797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mercer JA, Seperack PK, Strobel MC, Copeland NG, Jenkins NA. Novel myosin heavy chain encoded by murine dilute coat colour locus. Nature. 1991;349:709–713. doi: 10.1038/349709a0. [DOI] [PubMed] [Google Scholar]
- Stinchcombe JC, Griffiths GM. Regulated secretion from hemopoietic cells. J Cell Biol. 1999;147:1–6. doi: 10.1083/jcb.147.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hume AN, Collinson LM, Rapak A, Gomes AQ, Hopkins CR, Seabra MC. Rab27a regulates the peripheral distribution of melanosomes in melanocytes. J Cell Biol. 2001. [DOI] [PMC free article] [PubMed]
- Stinchcombe JC, Barral DC, Mules EH, Booth S, Hume AN, Machesky LM, Seabra MC, Griffiths GM. Rab 27a is required for regulated secretion in cytotoxic T lymphocytes. J Cell Biol. 2001. [DOI] [PMC free article] [PubMed]
- Yu W, Andersson B, Worley KC, Muzny DM, Ding Y, Liu W, Ricafrente JY, Wentland MA, Lennon G, Gibbs RA. Large-scale concatenation cDNA sequencing. Genome Res. 1997;7:353–358. doi: 10.1101/gr.7.4.353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warburg M, Sjo O, Tranebjaerg L, Fledelius HC. Deletion mapping of a retinal cone-rod dystrophy: assignment to 18q211. Am J Med Genet. 1991;39:288–293. doi: 10.1002/ajmg.1320390309. [DOI] [PubMed] [Google Scholar]
- Eichmuller S, Usener D, Dummer R, Stein A, Thiel D, Schadenforf D. Serological detection of cutaneous T-cell lymphoma-associated antigens. Proc Natl Acad Sci USA. 2001 doi: 10.1073/pnas.021386498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett DC, Cooper PJ, Dexter TJ, Devlin LM, Heasman J, Nester B. Cloned mouse melanocyte lines carrying the germline mutations albino and brown: complementation in culture. Development. 1989;105:379–385. doi: 10.1242/dev.105.2.379. [DOI] [PubMed] [Google Scholar]
- Chen D, Guo J, Gahl WA. RAB GTPases expressed in human melanoma cells. Biochim Biophys Acta. 1997;1355:1–6. doi: 10.1016/S0167-4889(96)00169-3. [DOI] [PubMed] [Google Scholar]
- Pittler SJ, Baehr W. Identification of a nonsense mutation in the rod photoreceptor cGMP phosphodiesterase -subunit gene of the rd mouse. Proc Natl Acad Sci USA. 1991;88:8322–8326. doi: 10.1073/pnas.88.19.8322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koopman P. Analysis of gene expression by reverse transcriptase by polymerase chain reaction. In Essential Developmental Biology A Practical Approach Edited by Stem CD, Holand PWH Oxford: IRC Press, 1993. pp. 233–242.
- Gyapay G, Schmitt K, Fizames C, Jones H, Vega-Czarny N, Spillett D, Muselet D, Prud'Homme JF, Dib C, Auffray C, Morissette J, Weissenbach J, Goodfellow PN. A radiation hybrid map of the human genome. Hum Mol Genet. 1996;5:339–46. doi: 10.1093/hmg/5.3.339. [DOI] [PubMed] [Google Scholar]
- McCarthy LC, Terrett J, Davis ME, Knights CJ, Smith AL, Critcher R, Schmitt K, Hudson J, Spurr NK, Goodfellow PN. A first-generation whole genome-radiation hybrid map spanning the mouse genome. Genome Res. 1997;7:1153–1161. doi: 10.1101/gr.7.12.1153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGuigan A, Huxley C. Replication of yeast DNA and novel chromosome formation in mouse cells. Nucleic Acids Res. 1996;24:2271–2280. doi: 10.1093/nar/24.12.2271. [DOI] [PMC free article] [PubMed] [Google Scholar]