

Abstract
Cutaneous T-cell lymphomas (CTCL) are a group of skin neoplasms that originate from T lymphocytes and are difficult to treat in advanced stages. The present study is aimed at the identification of tumor-specific antigens from a human testis cDNA library using human sera known as the SEREX (serological identification of recombinantly expressed genes) approach. A cDNA library from normal testicle tissue was prepared and approximately 2 million recombinants were screened with sera from Sézary Syndrome and Mycosis fungoides patients. A total of 28 positive clones belonging to 15 different genes/ORFs were identified, including five hitherto unknown sequences. Whereas control sera did not react with most clones, 11–71% sera from CTCL patients were reactive against the identified clones. Expression analysis on 28 normal control and 17 CTCL tissues by reverse transcription–PCR (RT-PCR) and Northern blotting revealed seven ubiquitously distributed antigens, six differentially expressed antigens (several normal tissues were positive), and two tumor-specific antigens that were expressed only in testis and tumor tissues: (i) A SCP-1-like sequence, which has already been detected in various tumors, has been found in one CTCL tumor and four sera of CTCL patients reacted with various SCP-1-like clones and (ii) a new sequence named cTAGE-1 (CTCL-associated antigen 1) was detected in 35% of CTCL tumor tissues and sera of 6/18 patients reacted with this clone. The present study unravels CTCL-associated antigens independent of the T-cell receptor. The SCP-1-like gene and cTAGE-1 were shown to be immunogenic and immunologically tumor-specific and may therefore be candidates for immunotherapy targeting CTCL.
Keywords: mycosis fungoides, Sézary Syndrome, SEREX, cancer-testis-antigen, tumor immunology
Cutaneous T-cell lymphomas (CTCL) are a heterogeneous group of lymphoproliferative disorders with predominantly CD4 T-cells as the malignant cell type (1–3) with a mono- or, at least, oligo-clonal origin (4). Besides various other subtypes, Mycosis fungoides (MF) and the Sézary Syndrome (SS) are the most common forms of CTCL (1, 2). Both are monoclonal T helper memory lymphomas presented by cutaneous patches, plaques, tumors, or erythroderma, whereby SS is in addition characterized by a generalized lymphadenopathy and the presence of neoplastic T cells in the peripheral blood (1–3).
Therapeutic approaches include stage-dependent selection of PUVA (psoralen and ultra violet A), retinoids, interferon α-2a in combination with acitretin or PUVA (5–8), various immunomodulators (9), electron radiation (10), or extracorporeal photopheresis (11). These methods are successful in early, but not in the aggressive, later stages of the disease. New concepts for therapy in CTCL are being discussed (12) that include immunological therapies such as vaccination with peptides or peptide-loaded dendritic cells, as it was used for treatment of melanoma (13, 14).
The presence and activity of CD8+ T-cells in CTCL has been correlated with prognosis (15, 16). CD8+ reactive infiltrates were demonstrated to be CTCL-specific and lytic (17, 18). Although immunotherapies might be a promising concept for CTCL, prerequisite for such strategies is the identification of tumor-specific antigens. The T-cell receptor itself has been suggested as an antigen (19), which could function as a specific target like the idiotype immunoglobulins do for B-cell lymphoma-specific T-cells (20). In both cases treatment regimens face the disadvantage that the antigen/T-cell receptor would have to be identified for each patient individually.
In the last few years the SEREX (serological identification of recombinantly expressed genes) approach has increasingly been used to screen for new tumor antigens. This method is based on the detection of antigens within recombinantly expressed tumor cDNA phage libraries by autologous antibodies (21). One variant of the method (22), which has also been applied in the present study, aims directly at the identification of antigens that are expressed solely in tumors and germline cells such as MAGE genes (23, 24). Cancer germline genes are expressed only in spermatoids/spermatogones and in tumors and thus, in view of the fact that the testis is an immunoprivileged site, can be considered immunologically tumor-specific (25, 26). Because of their expression pattern, this group of genes has also been named cancer-testis-antigens (26). The fact that a variety of tumor antigens is also expressed within germ line cells is generally attributed to a genome-wide hypomethylation in both tissues (27, 28). The number of identified cancer germline genes has rapidly increased within the last years giving rise to groups of highly homologous genes like MAGE, BAGE, GAGE, SCP-1, SSX, and NY-ESO-1 (26, 29). Most of these have originally been identified in melanoma, but subsequently have been found to be expressed in various other tumors like neuroblastoma (30), gastric carcinoma (31), ovarian tumors (32), and hepatocellular tumors (33). The possible value of these antigens for tumor immunological treatment is currently being investigated (13, 34, 35).
No tumor-associated antigens are currently known for CTCL. For the identification of CTCL-specific antigens we have chosen the SEREX approach for several reasons: (i) Methods using synthetic peptide libraries have so far not been successful in identifying naturally expressed CTCL-specific antigens (36), whereas (ii) SEREX has been able to unravel new tumor antigens that have secondarily been shown to also possess epitopes for cellular immune responses (e.g., NY-ESO-1; ref. 37). (iii) The method itself proves that the identified antigens are of immunological relevance, because patients have generated antibodies against the detected proteins. The aim of the present study was to use the SEREX approach to discover tumor-associated antigens in CTCL and to define their specificity in terms of serological response as well as RNA-expression within different tissues.
Materials and Methods
Tissue and Sera.
Sera and tumor tissues were obtained during routine diagnostic procedures with a formal agreement signed by the patients and with the official permission of the local ethical review board.
Construction of Testis cDNA Library.
RNA was extracted from a testicular specimen by using an RNA isolation kit (RNeasy midi kit) and subsequently an mRNA isolation kit (oligotex mRNA kit; both from Qiagen, Chatsworth, CA) as recommended by the manufacturer. A total of 10.4 μg of mRNA was used for the construction of the λ-ZAP expression library (Uni-ZAP XR custom cDNA library; Stratagene). The library consisted of 106 primary recombinants with an insert size larger than 0.4 kb and was amplified to 1010 plaque forming units.
Immunoscreening.
The immunological screen was performed essentially as described (38, 39). A detailed protocol is published as supplemental data on the PNAS web site, www.pnas.org. Positive phagemids were further analyzed by using sera of MF (n = 15) and SS (n = 3) patients, as well as healthy controls (n = 10). Positive phagemids were submitted to in vivo excision of the pBluescript plasmid, which was then sequenced. Primers were chemically synthesized.
Tumor Tissues and Cell Lines.
Tissue specimens obtained from 17 CTCL patients served as a source for generating tumor cDNA: 13 MF (stage Ib to IVb, mainly IIb), 2 SS (stage III), 1 T-zone lymphoma (stage IVb), and 1 CD30+ CTCL (stage IIb). In addition, cDNA from the following four CTCL cell lines was generated: My-La (MF; ref. 40), SeAx (SS; ref. 41), HH (Lymphomatoide papulosis, ATCC number: CRL-2105), and HuT-78 (SS, ATCC Number: TIB-161). Furthermore, we generated cDNA from six leukemia cell lines (ARA-10, Jurkat, KG1, K562, Nalm-2, and SKW6.4) and 22 melanoma cell lines.
Extensive control cDNAs were used for analyzing tissue distribution within normal tissues, including panels of commercially available cDNAs and RNAs (CLONTECH) and cDNA of three activated, CD8+ T-cell lines (kindly provided by Y. Sun, German Cancer Research Center; ref. 42). All control tissues are given in Table 1.
Table 1.
RT-PCR analyses using primers against differentially expressed sequences and multiple tissue cDNA panels
| Primers against clone, homologous to | se2-1 (SCP-1) | se14-2 (cTAGE-1) | se20-10 (⊘) | se33-1 (NP220) | se57-1 (⊘) | se70-2 (⊘) |
|---|---|---|---|---|---|---|
| Bone marrow | − | − | + | + | + | − |
| Brain | − | − | + | + | − | + |
| Colon | − | − | − | − | + | − |
| Fetal brain | − | − | − | + | − | + |
| Fetal heart | − | − | − | + | − | + |
| Fetal kidney | − | − | + | + | − | + |
| Fetal liver | − | − | − | − | − | + |
| Fetal lung | − | − | − | + | − | + |
| Fetal skeletal muscle | − | − | − | + | − | + |
| Fetal spleen | − | − | − | + | − | + |
| Fetal thymus | − | − | − | − | − | + |
| Heart | − | − | + | − | − | − |
| Kidney | − | − | + | + | − | + |
| Liver | − | − | − | + | − | − |
| Lung | − | − | − | + | − | − |
| Ovary | − | − | − | + | − | − |
| Pancreas | − | − | − | + | − | − |
| Peripheral blood lymphocytes | − | − | − | − | − | − |
| Placenta | − | − | + | + | − | − |
| Prostate | − | − | + | + | − | − |
| Skeletal muscle | − | − | − | − | − | − |
| Skin | − | − | − | + | − | − |
| Small intestine | − | − | + | + | + | + |
| Spleen | − | − | + | + | + | + |
| Stomach | − | − | + | + | − | − |
| Testis | + | + | + | + | + | + |
| Thymus | − | − | − | + | − | + |
| Trachea | − | − | + | + | + | + |
Either commercially available cDNA panels or cDNA generated from RNA panels (both from CLONTECH) were used for RT-PCR. Each probe consists of several donor tissues of the indicated type. cDNA from skin tissue was generated by RNA isolation of one specimen. For details see Material and Methods. ⊘, No homologous sequences found.
Reverse Transcription–PCR (RT-PCR).
Because of a limited amount of RNA, RT-PCR was used for investigating the expression of identified sequences within different normal and tumor tissues. In selected cases this was complemented by Northern blotting. RT-PCR was performed in at least two independent experiments (annealing for 1 min at variable temperature by using 35 cycles). RNA isolation, primer sequences, conditions for RT-PCR, and Northern blotting, as well as sequencing are detailed in the supplemental data.
Results
Screening.
Approximately 1.9 × 106 recombinant clones of a cDNA library derived from normal testis tissue were screened by 17 individual sera from patients suffering from CTCL, including MF and SS, to identify new tumor-specific antigens. Twenty-eight positive clones representing 15 different ORFs or genes were detected and further analyzed for serological reactivity and molecular distribution (Table 2). Each reactive plaque was further examined by using additional sera of patients diagnosed with MF (n = 15) or SS (n = 3) and 10 control sera from healthy volunteers. Reactivity of the patients' sera was slightly elevated at clinical stage III (data not shown). However, this was not statistically significant (χ2 test and Mann–Whitney U test), probably because of the low number of sera at advanced tumor stages. Reactivity of patients' sera to clones identified during screening was in the range of 11–71% (Table 3).
Table 2.
Identified clones, their frequency, size, and accession numbers, as well as homologous sequences published in the GenBank or the SEREX databases
| Clone* | No.† | Size | Accession no. | Homology to published sequences in the GenBank
database
|
Homology to the SEREX database‡ (clone) | ||
|---|---|---|---|---|---|---|---|
| Gene | Accession no. | Description | |||||
| se1-1 | 2 | 2363 | AF273042 | KIAA0336 | AB002334 | brain cDNA clone (49) | ⊘ |
| se2-1 | 6 | 2564 | AF273043 | SCP-1 | D67035 | SCP-1 (43) | Hom-TSRCC1-7 (22) |
| se2-2 | 1 | 2317 | AF273044 | KIAA0373 | AB002371 | brain cDNA clone (49) | ⊘ |
| se2-5 | 2 | 2004 | AF177228 | ⊘ | ⊘ | ⊘ | |
| se14-2 cTAGE-1 | 1 | 1284 | AF177229 | ⊘ | ⊘ | ⊘ | |
| se14-3 | 1 | 2620 | AF273045 | Rack 7 | U48251 | protein kinase C-binding protein; Kuroda et al, not published | ⊘ |
| se20-4 | 5 | 2830 | AF273046 | HRIHFB 2216 | AB015345 | nuclear targeted protein (50) | ⊘ |
| se20-7 | 2 | 2100 | AF273047 | trans-golgi | U41740 | human trans-golgi p230 (52) | NGO-St-67 (and others)§ |
| se20-9 | 1 | 5629 | AF273048 | KIAA0204 | D86959 | brain cDNA clone (48) | ⊘ |
| se20-10 | 1 | 1991 | AF177227 | ⊘ | ⊘ | ⊘ | |
| se33-1 | 1 | 3962 | AF273049 | NP220 | D83032 | DNA-binding nuclear protein (51) | LONY-BR-13¶ |
| se37-2 | 1 | 2710 | AF273050 | UBE3A | U84404 | E6-assoc. protein E6-AP/ubiquitinprotein ligase (56) | NY-CO-30 (57) |
| se57-1 | 2 | 3997 | AF273051 | ⊘ | ⊘ | TL15-72‖ | |
| se70-2 | 1 | 1592 | AF273052 | ⊘ | ⊘ | ⊘ | |
| se89-1 | 1 | 3462 | AF273053 | RAP140 | AF180425 | retinoblastoma-associated protein | ⊘ |
⊘, no homologous sequences found. Similarity in the overlapping sequences was always above 98%.
First identified clone of each antigen.
Total number of homologous clones to this antigen found in the present study.
The SEREX database is maintained by the Ludwig Institute (Switzerland) and can be found on the internet (http://www.licr.org/SEREX.html).
Y. Obata, Aichi Cancer Center, Japan.
M. J. Scanlan, Ludwig Institute for Cancer Research, New York.
A. Güre, Ludwig Institute for Cancer Research, New York.
Table 3.
Serological analyses of identified clones
| Clone | CTCL
|
Controls
|
||
|---|---|---|---|---|
| Reactive | n | Reactive | n | |
| se1-1 | 50% | 10 | 0% | 5 |
| se2-1 (SCP-1) | 22% | 18 | 0% | 10 |
| se33-2 | 17% | 12 | 20% | 5 |
| (se2-1 homologous) | ||||
| se2-2 | 30% | 10 | 0% | 5 |
| se2-5 | 30% | 10 | 0% | 5 |
| se14-2 (cTAGE-1) | 33% | 18 | 0% | 10 |
| se14-3 | 11% | 9 | 0% | 5 |
| se20-4 | 30% | 10 | 0% | 5 |
| se20-6 | 20% | 10 | 20% | 5 |
| (se20-4 homologous) | ||||
| se20-7 | 30% | 10 | 0% | 5 |
| se20-9 | 22% | 9 | 0% | 5 |
| se20-10 | 11% | 18 | 0% | 10 |
| se33-1 | 29% | 17 | 0% | 10 |
| se37-2 | 29% | 17 | 0% | 10 |
| se57-1 | 33% | 9 | 0% | 5 |
| se70-2 | 13% | 8 | 0% | 5 |
| se89-1 | 71% | 14 | 0% | 5 |
Table gives the percentage of reacting and the total number of tested sera (n) during secondary screening. In the case of clones se2-1 and se20-4, one of the homologous clones is also given, as these differed in their reactivity pattern, which may be due to sequence differences. For details see text.
Table 2 summarizes the characteristics of the 15 genes identified during the study by giving the first antigenic clone of each gene, the number of homologous clones found, the size of the insert, and information derived from the GenBank database, as well as the SEREX database of the Ludwig Institute for Cancer Research (Switzerland; available via internet: http://www.licr.org/SEREX.html). Five antigens represent hitherto unknown sequences (se2-5, se14-2, se20-10, se57-1, and se70-2). The RNA expression pattern of the identified antigens as analyzed by RT-PCR varied between highly restricted to ubiquitously expressed in 28 normal and 17 CTCL tumor tissues and 33 tumor cell lines of diverse origin (Tables 1 and 4).
Table 4.
RT-PCR analyses using antigen-specific primers and cDNA from various tissues and cell lines
| cDNA, primers against clone | Controls
|
Tumor tissues (CTCL, n) | Cell lines
|
|||
|---|---|---|---|---|---|---|
| Multi-tissue panels* (n) | Activated CTLs† (n = 3) | CTCL (n = 4) | Leukemia (n = 6) | Melanoma (n) | ||
| se1-1 | 100% (16) | 100% | 91% (11) | 100% | 100% | 100% (6) |
| se2-1 | 4% (28)‡ | 0% | 6% (17) | 0% | 0% | 0% (11) |
| se2-2 | 100% (16) | 100% | 90% (10) | 100% | 83% | 80% (5) |
| se2-5 | 100% (16) | 0% | 55% (11) | 0% | 100% | 100% (11) |
| se14-2 (cTAGE-1) | 4% (28)‡ | 0% | 35% (17) | 100% | 0% | 0% (20) |
| se14-3 | 100% (16) | 100% | 100% (11) | 100% | 100% | 100% (11) |
| se20-4 | 100% (16) | 100% | 92% (12) | 100% | 100% | 100% (11) |
| se20-7 | 100% (16) | 100% | 100% (11) | 100% | 50% | 80% (5) |
| se20-9 | 100% (16) | 100% | 82% (11) | 100% | 83% | 45% (11) |
| se20-10 | 43% (28)‡ | 67% | 77% (13) | 100% | 100% | 55% (11) |
| se33-1 | 79% (28)‡ | 100% | 75% (16) | 100% | 83% | 100% (11) |
| se37-2 | 100% (16) | 100% | 93% (15) | 100% | 83% | 75% (5) |
| se57-1 | 21% (28)‡ | 0% | 6% (17) | 0% | 0% | 0% (23) |
| se70-2 | 54% (28)‡ | 33% | 31% (16) | 100% | 100% | 45% (22) |
| se89-1 | 79% (14) | 100% | 75% (16) | 100% | 100% | 73% (22) |
Table gives numbers as percentage of positive tissues or cell lines when analyzed by RT-PCR using clone-specific primers and the total number of tested specimen (n) in parentheses. Differentially expressed and cancer-germline clones are bold.
RT-PCRs using testis cDNA gave always positive results. In the case of clones se2-1 and se14-2 cDNA from testis tissue was the only positive probe. Content of panels see Table 1.
Activated cytotoxic T-cells (see material and methods).
For specific results on these clones see Table 1.
Cancer Germline Genes.
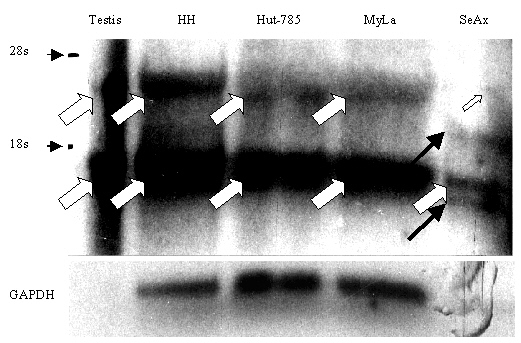
Clone se14-2 represented a new antigen, which was not found in either database with the exception of short expressed sequence tag (EST) sequences. se14-2 protein was detected by one third of the patients' sera (4 of 15 sera of MF and 2 of 3 sera of SS patients), whereas all 10 control sera were nonreactive (Table 3). Analyses of mRNA levels in 28 normal tissues by RT-PCR detected specific mRNA in testis only (Tables 1 and 4). An se14-2-specific message was found by RT-PCR in 35% of the investigated CTCL tissues and in all four CTCL cell lines (Table 4) (in detail, MF: 4/13, T-zone lymphoma, 1/1; SS: 1/2, CD30+ Lymphoma, 0/1; CTCL cell lines: 4/4). In contrast, none of the leukemia (0/6) and none of the melanoma cell lines (0/20) were found to express se14-2 specific mRNA. Therefore, the DNA insert of clone se14-2 was named cTAGE-1 (CTCL-associated antigen 1; accession no. AF177229). Sequence analyses revealed homology to an EST of 633 bp derived from a testis library (accession no. Aa868597). Interestingly, within the first 327 bp there is a 95% similarity, followed by a 397-bp gap in the EST sequence and another 307- bp overlap with 100% identity. cTAGE-1 is 1284 bp-long and shows no clear ORF. The longest possible polypeptide is 74 aa in length. Database searches revealed a 99% identity with chromosome 18p11.2 (accession no. AP001126) in a 1265 bp-overlap. Northern blot analysis by using RNA derived from testis and several CTCL cell lines revealed two to four bands (see Fig. 1 of the supplemental data): both testis tissue and cell lines showed bands at approximately 4 kb and 1.3 kb. The cell line SeAx showed additional bands at 2.3 kb and 0.5 kb.
Six clones, representing together five different recombinants, were homologous to SCP-1, a meiosis-associated protein: clone se2-1 and se3-2 were identical, whereas clones se5-1, se20-8, se33-2, and se37-1 showed differences in length. An SCP-1 homologous clone has also been identified by the SEREX approach by Türeci et al. by using a testis library and a renal cell carcinoma patient's serum, Hom-Tsrcc1-7 (22). This gene has already been described as a cancer-testis-antigen, which could not be found in any control tissues besides testis, but could be detected in a variety of different cancer tissues. Sequence alignment showed that all identified SCP-1-like clones belong to one gene and had only four base pair differences in comparison to Hom-Tsrcc1-7 resulting in three amino acid exchanges, but several differences in comparison to SCP-1 as published by Kondoh et al. (ref. 43; see Figs. 2 and 3 of the supplemental data). The latter leads, e.g., to a region of 43 amino acid mismatches (see Fig. 3, amino acids 483 to 533, of the supplemental data). All SCP-1-like clones were truncated at the 3′ end, which also resulted in shortened ORF and thus may reflect truncated cDNA clones belonging to the same gene as Hom-Tsrcc1-7. Additionally, one clone (se33-2) exhibited a gap of 9 base pairs resulting in a loss of 3 amino acids, and a shortening at the 5′ end so that the ORF theoretically started at base pair 14. Interestingly, the degree of serological reactivity of various sera varied for specific SCP-1 clones: Clone se2-1 was detected by sera from 2/15 MF and 2/3 SS patients (Table 3). Another clone with high homology to SCP-1 (se37-1) was detected by 3/9 MF sera, but only one of 3 SS sera. This might reflect multiple epitopes of SCP-1, as the clones varied in length. Moreover, clone se33-2 also reacted with 1/5 control sera (Table 3), which might be explained by the sequence differences (see above). PCR analysis was done by using the same primers as published by Türeci et al. (22); these primers were also suited perfectly for our SCP-1-like clones. Among all of the tested normal tissues, as well as tumor specimen and cell lines, we found only testis and one MF (patient H.S.) specimen expressing SCP-1 mRNA (Tables 1 and 4). The positive RT-PCR result of H.S. cDNA could be confirmed by Northern blotting resulting in an ≈4.3-kb-sized band (data not shown). In addition, the serum of patient H.S. also reacted with se2-1 and one of the other SCP-1 homologous clones.
In the following two paragraphs 13 antigens with differential or ubiquitous expression patterns as detected by RT-PCR (Tables 1 and 4) are described. All sequences were submitted to GenBank; the corresponding accession numbers and, where available, published homologous genes are given in Table 2. Details on possible ORFs, as well as alignments/differences to homologous sequences, will be published elsewhere.
Antigens with Restricted Expression Patterns.
Four new antigens (se2-5, se20-10, se57-1, and se70-2) and two antigens with homologies to known genes (se33-1: NP220; se89-1: retinoblastoma-associated protein RAP140) were found to be differentially expressed on the molecular level. The serological reactivity against these clones as defined by the percentage of reactive sera during secondary screening was 31% on the average (Table 3). A low rate of reactivity was noticed against clones se20-10 and se70-2 (2/18 and 1/8 reactive sera, respectively), whereas 71% of CTCL patients' sera (n = 14) responded against clone se89-1. All six clones were not reactive with up to 10 normal control sera (Table 3).
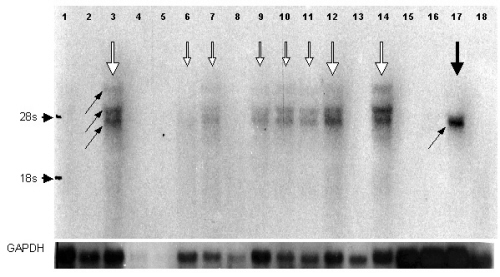
By RT-PCR analysis, the clone se2-5 specific message was almost ubiquitously expressed within normal tissues, but was absent in activated T-cells and expressed in only 55% of the CTCL tissues (Table 4). Northern blot analysis, in contrast, revealed a restricted expression pattern even within normal tissues (see Fig. 4 of the supplemental data). Strong signals were detectable in kidney, trachea, and testis, and weaker signals in colon, small intestine, thymus, bone marrow, and stomach. Whereas in all positive normal tissues three bands could be found (5.2, 4.2, and 3.9 kb), the only positive CTCL cell line SeAx exhibited one signal at 3.9 kb.
Specific mRNA for clones se20-10 and se57-1 were found in 43% and 21% of the investigated control tissues by RT-PCR (Tables 1 and 4), respectively. Interestingly, expression of se57-1-specific message was dramatically down-regulated in all tumor tissues and cell lines (Table 4). In contrast, clone se70-2 mRNA expression was up-regulated within CTCL and leukemia cell lines (100%) in comparison to normal tissues (54%), whereas CTCL tissue and melanoma cell lines showed a medium expression level (33% and 45%, respectively). Remarkably, among the control tissues all fetal tissues were positive in RT-PCR.
Two antigens with homologies to known sequences were found to be differentially expressed: se33-1 cDNA is homologous to NP220, a DNA-binding protein, and se89-1 cDNA is homologous to RAP140, a retinoblastoma-associated clone (Table 2). Clone se33-1 showed 99% similarity in a 3830-bp overlapping stretch at the 3′ end of NP220, but is truncated at the 5′ end, which resulted in a shortened ORF. RT-PCR using the se33-1-specific primer revealed mRNA in 6/8 fetal and in 16/20 normal tissues (Table 1), as well as within CTCL tissues (12/16), activated cytotoxic T-cells, and in most cell lines (Table 4).
Clone se89-1 cDNA showed a 98% similarity to RAP140 in a 3444-bp overlap and a gap of 60 bp, which is located within the ORF resulting in a 20-aa gap. RT-PCR was performed by using various primers: first a combination of primers (RAP140for and RAP140rev; see Materials and Methods) detected both RAP140 and se89-1 and revealed three bands in testis cDNA and two bands in various other cDNAs. To specify the expression of se89-1 we generated a new reverse primer spanning the gap within se89-1. By using this primer together with the forward primer against RAP140, which also detects se89-1, only one band was amplified. The frequency of positive cDNAs to the se89-1-specific primers did not differ significantly between control tissues (79%), CTCL tissues (75%), and cell lines (CTCL and leukemia lines, 100%; melanoma lines, 73%, Table 4). By using Northern blotting, the presence of se89-1-specific RNA was confirmed within RNA derived from brain, kidney, colon, and testis and the CTCL line SeAx (data not shown).
Ubiquitously Expressed Antigens.
Six antigens (se1-1, se2-2, se14-3, se20-4, se20-9, and se37-2) homologous to known genes (Table 2) were found to be ubiquitously expressed. In all control tissue (n = 28), mRNA specific for these clones was detectable by RT-PCR (Table 4). In two clones, se14-3 and se20-4, all tumor tissues and cell lines were also RT-PCR-positive. In contrast, clones se2-2, se20-7, se20-9, and se37-2 were expressed only in a subset of melanoma and leukemia cell lines, whereas CTCL tissues and CTCL cell lines showed a high percentage of RT-PCR-positive cDNAs (Table 4).
The reactivity of patients' sera with these clones averaged about 29% (Table 3) including two extremes: reactivity against clone se14-3 was found in 11% (1/9) and against clone se1-1 in 50% (5/10) CTCL sera. Two control sera (n = 10) reacted to clone se20-6, which is homologous to clone se20-4 (Table 3). se20-6 was shown to code for a different, 72-aa peptide in the first reading frame, which was not present in either se20-4 or its homologous gene HRIHFB2216 (Table 2).
Sequence analyses of these clones and comparison to their homologous counterparts revealed insertions, deletions, or elongations in some cases. A detailed analysis on these matters and on the consequences for the corresponding ORFs will be published separately.
Discussion
Up to now only one attempt, that of Linnemann et al. (36), by using a synthetic peptide library, was undertaken to find new tumor antigens that are expressed in various CTCL tumor tissues. Whereas Linnemann and colleagues could prove that peptide-specific cytotoxic T-cells can be generated that are able to kill the CTCL line MyLa in vitro when the tumor cells were loaded with the identified epitope, they failed to identify a natural homologue to the synthetic epitope. In a recent study (44), however, the same group was able to generate, by using another synthetic peptide, cytotoxic T-cells that were able to kill the HLA-matched, allogenic cell line MyLa, indicating that they are specific for a naturally expressed peptide that is still unknown.
The present study identifies 15 CTCL antigens that are independent of the T-cell receptor. The specificity of these antigens was examined on two levels: (i) Taking the RNA expression pattern of the investigated CTCL antigens as analyzed by RT-PCR, only two antigens, namely an SCP-1 homologous gene (represented by six clones) and cTAGE-1, were cancer/testis-specific, as they were not expressed in any control tissue besides testis. Six antigens (8 clones) could be found in at least 21% of control tissues, whereas another seven antigens (13 clones) were expressed in almost every tissue examined. (ii) This contrasts to the serological specificity: 26 of the 28 identified clones were detected only by sera of CTCL patients and not of healthy control sera. Future experiments with an increased number of tested sera have to unravel the relevance of these findings. Antigens inducing autoantibodies, which should be expected in the SEREX approach (21, 45), were represented by two clones: one of the six clones that were homologous to SCP-1 (se33-2) and one of the five clones that were homologous to HRIHFB2216 (se20-6). In both cases an unusual ORF at the 5′ end of the sequence might code for a peptide responsible for the reactivity with autoantibodies in normal control sera.
Two antigens, namely an SCP-1-like sequence and cTAGE-1, were tumor-specific in the humoral response of the tested sera and could be addressed as cancer germline cells because of their restricted RNA expression as assessed by RT-PCR using 28 control cDNAs and a large number of cancer tissues and cell lines. SCP-1 is part of the synaptonemal complex that is responsible for the accurate pairing of the homologous chromosomes during meiosis (43). A homologous clone to SCP-1 (Hom-Tsrcc1-7) was also found by the SEREX approach using serum of a renal cell carcinoma patient analyzing a testis library (22). We found six clones with a higher similarity to Hom-Tsrcc1-7 than to SCP-1, although they differed in length. Four of 18 sera of CTCL patients were reactive against clone se2-1, which contained the longest ORF, although it is still truncated at the C-terminal in comparison to Hom-Tsrcc1-7 and SCP-1. Although the mRNA expression within the tested CTCL tissues was low (6%), we could examine tumor tissue-derived RNA of one serologically positive patient, resulting in a positive signal both in RT-PCR and Northern blotting. Unfortunately, there was no tumor RNA available from the other serum-positive patients.
cTAGE-1 (clone se14-2) is a new cancer germline gene established in the present study. Its sequence is quite complex, as no long ORF could be found. A 74-aa sequence could be detected within the second reading frame starting at bp 14, which might be the possible immunogenic target. cTAGE-1 is located at chromosome 18p11.2 downstream of—but not overlapping with—a region with high homology to the MEA family (46)/MGEA-family (47). The relationship between cTAGE-1 and MEA is not solved and whether cTAGE-1 is a cancer-testis-specific splicing variant of an unknown MEA-family member is currently under investigation. In contrast to SCP-1, the cTAGE-1 sequence has been found in a substantial number of CTCL tissues (35%) and in all four investigated CTCL cell lines by RT-PCR. Although cTAGE-1 could not been detected within melanoma cell lines, preliminary results suggest that cTAGE-1 is expressed in head–neck carcinomas and carcinoma of ovarial tissues to a similar extent (23% and 30%, respectively; manuscript in preparation). This makes cTAGE-1 attractive for use as antigen in tumor therapy. The sequence of cTAGE-1 as well as the two to four bands in the Northern blot analysis of four CTCL cell lines imply that cTAGE-1 is a part of, or a splicing product of, a longer sequence. Studies are currently being carried out to identify the other putative members of the cTAGE-family and their possible immunogenic epitopes.
Besides the cancer germline sequence cTAGE-1, we were able to identify another four sequences that could be found in databases only as ESTs and in some cases as chromosomal sequences: clones se2-5, se20-10, se57-1, and se70-2. All of these clones code for mRNAs that could be shown to be differentially expressed within normal and tumor tissues as well as tumor cell lines. A detailed analysis of these sequences with respect to ORF, containing motifs, splicing variants, and chromosomal location, will be published separately.
Several identified antigens showed a high homology to already known genes, some of which have not been associated with cancer, although five of these sequences and one additional antigen that is not published in gene banks have also been found in other SEREX studies (Table 2). Four clones were homologous to three different KIAA genes (see Table 2), which were identified by sequencing a cDNA library derived from brain tissue and which code for large proteins (48, 49). All of these are ubiquitously distributed, which is in line with the original publication. Up to now, nothing is known about the function of theses proteins.
Five clones are homologous to HRIHFB2216, which codes for a nuclear targeted protein with known DNA binding motifs (50). The sequence of the longest clone (se20-4) is longer than HRIHFB2216 in both the 5′ and 3′ directions and the deduced amino acid sequence is elongated at the N-terminal by 207 aa. Another DNA binding, nuclear protein named NP220 (51) is homologous to clone se33-1, although the ORF of se33-1 translates only to 903 aa of the C-terminal of NP220. These sequences were ubiquitously expressed (se20-4) or detected in most tissues (se33-1). About 30% of patients' sera reacted to each clone. Two clones (se20-7 and se33-3) were homologous to a gene located at chromosome 6p12-22 and coded for the human trans-golgi p230 protein, which is assumed to play a key role during transport of proteins by the Golgi apparatus (52). Interestingly, a homologous clone to this gene has also been found in another SEREX study by using a stomach cDNA library (see Table 2).
Clone se37-2 cDNA is homologous to a gene named UBE3A, which codes for the E6-AP ubiquitin-protein-ligase, a protein interacting between human papilloma virus E6 oncoproteins and the tumor suppressor p53 (53). This gene gives rise to a variety of splicing products (54, 55). Clone se37-2 fits best in its nucleic acid and its deduced amino acid sequence to an alternatively spliced variant (accession no.: U84404) that is associated with Angelmann syndrome (56). Interestingly, another homologous clone named NY-CO-30 has been identified by the SEREX approach by using a cDNA library from a lung metastasis of a colon carcinoma and autologous serum for screening (57). In contrast to the present study, these authors found 63% of the patients' sera and 38% of control sera reacting with clone NY-CO-30. Amino acid sequence analyses revealed that NY-CO-30 protein fits best to a slightly different sequence that has been submitted as “homo sapiens papillomavirus E6 oncogenic protein-associated protein” (accession no.: L07557; ref. 53). Differences in reactivity of the control sera may in part be due to these sequence differences.
Clone se89-1 shows homology to the retinoblastoma-associated antigen rap140 (accession no.: AF180425; Quan et al., unpublished data), but is truncated at the 5′ end by 1300 bp and possesses a 60-bp gap within the coding sequence. Consequently, the deduced amino acid sequence is shortened by 178 aa at the N-terminal and by a 20-aa gap. In addition, there are only three mismatches in the protein sequence. Both sequences may be splicing variants of the same gene.
The list of tumor antigens generating both humoral and cellular immune responses in tumor patients is still growing (21, 38, 45, 58). Jäger et al. (59) showed in a recent publication that the presence of NY-Eso-1-specific, cytotoxic T-cells correlates with the presence of NY-Eso-1-specific antibodies in the same patient. Tumor antigens themselves might also be used for the generation of bispecific antibodies (60, 61), provided they are specifically expressed.
Our study proves that tumor antigens of CTCL tumors can be identified by antibodies derived from tumor patients for screening a recombinant library. Moreover, two groups of tumor antigens could be discovered with different possible clinical applications: (i) Two antigens, namely cTAGE-1 and an SCP-1-like gene, are cancer germline genes because of their RNA expression and thus may be applicable for immunological therapies, although ongoing research in respect to cTAGE-1 has to establish the essential evidence. (ii) All antigens described in this paper are specific in that only tumor patients, no healthy volunteers, generated antibodies against them, although 13 of these tumor antigens are expressed in at least 21% or even within every tested control tissue. Yet, the question needs to be answered, whether the expression of the derived proteins is also widespread or possibly tumor-specific. In any case, the described antigens may be of further interest for use as tumor markers for diagnostic or prognostic purposes or for monitoring. After further investigations, the new tumor antigens, especially cTAGE-1, may be added to the growing list of potential targets for immunotherapies—not only for CTCL but also for various other tumor entities. Finally, because of the knowledge of specific antigens, CTCL may become amenable to “cancer immunotherapy based on the genes that encode cancer antigens” (14).
Supplementary Material
Acknowledgments
We thank Dr. Ö. Türeci and Prof. M. Pfreundschuh for giving us an introduction to the SEREX approach; Mrs. Anita Jochim for excellent technical assistance; Dr. H. Delius (German Cancer Research Center) for sequencing; and Bennedikt Brors (German Cancer Research Center) for help in bioinformatics. This work was supported in part by grants from the Deutsche Forschungsgemeinschaft (DFG Ei 433-1) and the “Forschungsfond der Fakultät für Klinische Medizin Mannheim” (to S.E.).
Abbreviations
- CTCL
cutaneous T-cell lymphoma
- MF
Mycosis fungoides
- RT-PCR
reverse transcription–PCR
- SS
Sézary Syndrome
- SEREX
serological identification of recombinantly expressed genes
- cTAGE-1
CTCL-associated antigen 1
- EST
expressed sequence tag
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
Data deposition: The sequences reported in this paper have been deposited in the GenBank database (accession nos. AF177227–AF177229 and AF273042–AF273053).
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.021386498.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.021386498
References
- 1.Burg G, Kempf W, Haeffner A, Nestle F, Hess-Schmid M, Doebbeling U, Mueller B, Dummer R. Curr Probl Dermatol. 1997;9:137–204. [Google Scholar]
- 2.Willemze R, Kerl H, Sterry W, Berti E, Cerroni L, Chimenti S, Diaz Perez J L, Geerts M L, Goos M, Knobler R, et al. Blood. 1997;90:354–371. [PubMed] [Google Scholar]
- 3.Jahn S, Asadullah K, Walden P, Sterry W. Immunol Today. 1998;19:100–103. doi: 10.1016/s0167-5699(97)01212-7. [DOI] [PubMed] [Google Scholar]
- 4.Weiss L M, Hu E, Wood G S, Moulds C, Cleary M L, Warnke R, Sklar J. N Engl J Med. 1985;313:539–544. doi: 10.1056/NEJM198508293130903. [DOI] [PubMed] [Google Scholar]
- 5.Dreno B, Claudy A, Meynadier J, Verret J, Souteyrand P, Ortonne J, Kalis B, Godefroy W, Beerblock K, Thill L. Br J Dermatol. 1991;125:456–459. doi: 10.1111/j.1365-2133.1991.tb14772.x. [DOI] [PubMed] [Google Scholar]
- 6.Bunn P A, Jr, Hoffman S J, Norris D, Golitz L E, Aeling J L. Ann Intern Med. 1994;121:592–602. doi: 10.7326/0003-4819-121-8-199410150-00007. [DOI] [PubMed] [Google Scholar]
- 7.Dummer R, Häffner A, Hess M, Burg G. Onkologie. 1996;19:226–230. [Google Scholar]
- 8.Stadler R, Otte H, Luger T, Henz B, Kühl P, Zwingers T, Sterry W. Blood. 1998;92:3578–3581. [PubMed] [Google Scholar]
- 9.Rogalski C, Dummer R, Burg G. J Eur Acad Dermatol Venereol. 1999;113:83–90. [PubMed] [Google Scholar]
- 10.Jones G, Rosenthal D, Wilson L. Cancer. 1999;85:1985–1995. doi: 10.1002/(sici)1097-0142(19990501)85:9<1985::aid-cncr16>3.0.co;2-o. [DOI] [PubMed] [Google Scholar]
- 11.Russel-Jones R. Br J Dermatol. 2000;142:16–21. doi: 10.1046/j.1365-2133.2000.03286.x. [DOI] [PubMed] [Google Scholar]
- 12.Duvic M, Cather J. Dermatologic Clinics. 2000;18:147–156. doi: 10.1016/s0733-8635(05)70155-8. [DOI] [PubMed] [Google Scholar]
- 13.Nestle F O, Alijagic S, Gilliet M, Sun Y, Grabbe S, Dummer R, Burg G, Schadendorf D. Nat Med. 1998;4:328–332. doi: 10.1038/nm0398-328. [DOI] [PubMed] [Google Scholar]
- 14.Rosenberg S. Immunity. 1999;10:281–287. doi: 10.1016/s1074-7613(00)80028-x. [DOI] [PubMed] [Google Scholar]
- 15.Hoppe R T, Medeiros L J, Warnke R A, Wood G S. J Am Acad Dermatol. 1995;32:448–453. doi: 10.1016/0190-9622(95)90067-5. [DOI] [PubMed] [Google Scholar]
- 16.Asadullah K, Friedrich M, Docke W D, Jahn S, Volk H D, Sterry W. J Invest Dermatol. 1997;108:743–747. doi: 10.1111/1523-1747.ep12292129. [DOI] [PubMed] [Google Scholar]
- 17.Berger C L, Wang N, Christensen I, Longley J, Heald P, Edelson R L. J Invest Dermatol. 1996;107:392–397. doi: 10.1111/1523-1747.ep12363378. [DOI] [PubMed] [Google Scholar]
- 18.Bagot M, Echchakir H, Mami Chouaib F, Delfau Larue M H, Charue D, Bernheim A, Chouaib S, Boumsell L, Bensussan A. Blood. 1998;91:4331–4341. [PubMed] [Google Scholar]
- 19.Berger C L, Longley B J, Imaeda S, Christensen I, Heald P, Edelson R L. Int J Cancer. 1998;76:304–311. doi: 10.1002/(sici)1097-0215(19980504)76:3<304::aid-ijc3>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- 20.Osterroth F, Garbe A, Fisch P, Veelken H. Blood. 2000;95:1342–1349. [PubMed] [Google Scholar]
- 21.Türeci Ö, Sahin U, Pfreundschuh M. Mol Med Today. 1997;3:342–349. doi: 10.1016/s1357-4310(97)01081-2. [DOI] [PubMed] [Google Scholar]
- 22.Türeci Ö, Sahin U, Zwick C, Koslowski M, Seitz G, Pfreundschuh M. Proc Natl Acad Sci USA. 1998;95:5211–5216. doi: 10.1073/pnas.95.9.5211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Boon T, Cerottini J C, Van den Eynde B, van der Bruggen P, Van Pel A. Annu Rev Immunol. 1994;12:337–365. doi: 10.1146/annurev.iy.12.040194.002005. [DOI] [PubMed] [Google Scholar]
- 24.Castelli C, Rivoltini L, Andreola G, Carrabba M, Renkvist N, Parmiani G. J Cell Physiol. 2000;182:323–331. doi: 10.1002/(SICI)1097-4652(200003)182:3<323::AID-JCP2>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- 25.Takahashi K, Shichijo S, Noguchi M, Hirohata M, Itoh K. Cancer. 1995;55:3478–3482. [PubMed] [Google Scholar]
- 26.Chen Y T, Old L J. Cancer J Sci Am. 1999;5:16–17. [PubMed] [Google Scholar]
- 27.Sahin U, Türeci Ö, Pfreundschuh M. Curr Opin Immunol. 1997;9:709–716. doi: 10.1016/s0952-7915(97)80053-2. [DOI] [PubMed] [Google Scholar]
- 28.De Smet C, Lurquin C, Lethe B, Martelange V, Boon T. Mol Cell Biol. 1999;19:7327–7335. doi: 10.1128/mcb.19.11.7327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Old L, Chen Y. J Exp Med. 1998;187:1163–1167. doi: 10.1084/jem.187.8.1163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Corrias M V, Scaruffi P, Occhino M, De Bernardi B, Tonini G P, Pistoia V. Int J Cancer. 1996;69:403–407. doi: 10.1002/(SICI)1097-0215(19961021)69:5<403::AID-IJC9>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 31.Li J, Yang Y, Fujie T, Baba K, Ueo H, Mori M, Akiyoshi T. Clin Cancer Res. 1996;2:1619–1625. [PubMed] [Google Scholar]
- 32.Gillespie A M, Rodgers S, Wilson A P, Tidy J, Rees R C, Coleman R E, Murray A K. Br J Cancer. 1998;78:816–821. doi: 10.1038/bjc.1998.585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Tahara K, Mori M, Sadanaga N, Sakamoto Y, Kitano S, Makuuchi M. Cancer. 1999;85:1234–1240. [PubMed] [Google Scholar]
- 34.Thurner B, Haendle I, Roder C, Dieckmann D, Keikavoussi P, Jonuleit H, Bender A, Maczek C, Schreiner D, von den Driesch P, et al. J Exp Med. 1999;190:1669–1678. doi: 10.1084/jem.190.11.1669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mackensen A, Herbst B, Chen J L, Kohler G, Noppen C, Herr W, Spagnoli G C, Cerundolo V, Lindemann A. Int J Cancer. 2000;86:385–392. doi: 10.1002/(sici)1097-0215(20000501)86:3<385::aid-ijc13>3.0.co;2-t. [DOI] [PubMed] [Google Scholar]
- 36.Linnemann T, Brock C, Sparbier K, Muche M, Mielke A, Lukowsky A, Sterry W, Kaltoft K, Wiesmuller K H, Walden P. Adv Exp Med Biol. 1998;451:231–235. doi: 10.1007/978-1-4615-5357-1_36. [DOI] [PubMed] [Google Scholar]
- 37.Jäger E, Jäger D, Karbach J, Chen Y T, Ritter G, Nagata Y, Gnjatic S, Stockert E, Arand M, Old L J, et al. J Exp Med. 2000;191:625–630. doi: 10.1084/jem.191.4.625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sahin U, Türeci Ö, Schmitt H, Cochlovius B, Johannes T, Schmits R, Stenner F, Luo G, Schobert I, Pfreundschuh M. Proc Natl Acad Sci USA. 1995;92:11810–11813. doi: 10.1073/pnas.92.25.11810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Türeci O, Sahin U, Schobert I, Koslowski M, Schmitt H, Schild H J, Stenner F, Seitz G, Rammensee H G, Pfreundschuh M. Cancer Res. 1996;56:4766–4772. [PubMed] [Google Scholar]
- 40.Kaltoft K, Bisballe S, Dyrberg T, Boel E, Rasmussen P B, Thestrup Pedersen K. In Vitro Cell Dev Biol Anim. 1992;28a:161–167. doi: 10.1007/BF02631086. [DOI] [PubMed] [Google Scholar]
- 41.Kaltoft K, Bisballe S, Rasmussen H F, Thestrup Pedersen K, Boehncke W H, Volker H, Sterry W. Arch Dermatol Res. 1988;280:264–267. doi: 10.1007/BF00440598. [DOI] [PubMed] [Google Scholar]
- 42.Möller P, Sun Y S, Dorbic T, Alijagic S, Makki A, Jurgovsky K, Schroff M, Henz B M, Wittig B, Schadendorf D. Br J Cancer. 1998;77:1907–1916. doi: 10.1038/bjc.1998.317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kondoh N, Nishina Y, Tsuchida J, Koga M, Tanaka H, Uchida K, Inazawa J, Taketo M, Nozaki M, Nojima H, et al. Cytogenet Cell Genet. 1997;78:103–104. doi: 10.1159/000134637. [DOI] [PubMed] [Google Scholar]
- 44.Linnemann T, Wiesmuller K H, Gellrich S, Kaltoft K, Sterry W, Walden P. Ann Oncol. 2000;11,Suppl.1:95–99. [PubMed] [Google Scholar]
- 45.Chen Y T, Güre A O, Tsang S, Stockert E, Jäger E, Knuth A, Old L J. Proc Natl Acad Sci USA. 1998;95:6919–6923. doi: 10.1073/pnas.95.12.6919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Heckel D, Brass N, Fischer U, Blin N, Steudel I, Türeci O, Fackler O, Zang K D, Meese E. Hum Mol Genet. 1997;6:2031–2041. doi: 10.1093/hmg/6.12.2031. [DOI] [PubMed] [Google Scholar]
- 47.Comtesse N, Heckel D, Racz A, Brass N, Glass B, Meese E. Clin Cancer Res. 1999;5:3560–3568. [PubMed] [Google Scholar]
- 48.Nagase T, Seki N, Ishikawa K, Ohira M, Kawarabayasi Y, Ohara O, Tanaka A, Kotani H, Miyajima N, Nomura N. DNA Res. 1996;3:321–329. doi: 10.1093/dnares/3.5.321. [DOI] [PubMed] [Google Scholar]
- 49.Nagase T, Ishikawa K, Nakajima D, Ohira M, Seki N, Miyajima N, Tanaka A, Kotani H, Nomura N, Ohara O. DNA Res. 1997;4:141–150. doi: 10.1093/dnares/4.2.141. [DOI] [PubMed] [Google Scholar]
- 50.Ueki N, Oda T, Kondo M, Yano K, Noguchi T, Muramatsu M. Nat Biotechnol. 1998;16:1338–1342. doi: 10.1038/4315. [DOI] [PubMed] [Google Scholar]
- 51.Inagaki H, Matsushima Y, Nakamura K, Ohshima M, Kadowaki T, Kitagawa Y. J Biol Chem. 1996;271:12525–12531. doi: 10.1074/jbc.271.21.12525. [DOI] [PubMed] [Google Scholar]
- 52.Erlich R, Gleeson P A, Campbell P, Dietzsch E, Toh B H. J Biol Chem. 1996;271:8328–8337. doi: 10.1074/jbc.271.14.8328. [DOI] [PubMed] [Google Scholar]
- 53.Huibregtse J, Scheffner M, Howley P. Mol Cell Biol. 1993;13:775–784. doi: 10.1128/mcb.13.2.775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kishino T, Wagstaff J. Genomics. 1998;47:101–107. doi: 10.1006/geno.1997.5093. [DOI] [PubMed] [Google Scholar]
- 55.Yamamoto Y, Huibregtse J M, Howley P M. Genomics. 1997;41:263–266. doi: 10.1006/geno.1997.4617. [DOI] [PubMed] [Google Scholar]
- 56.Kishino T, Lalande M, Wagstaff J. Nat Genet. 1997;15:70–73. doi: 10.1038/ng0197-70. [DOI] [PubMed] [Google Scholar]
- 57.Scanlan M J, Chen Y-T, Williamson B, Güre A O, Stockert E, Gordan J D, Türeci Ö, Sahin U, Pfreundschuh M, Old L J. Int J Cancer. 1998;76:652–658. doi: 10.1002/(sici)1097-0215(19980529)76:5<652::aid-ijc7>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- 58.Jäger E, Chen Y T, Drijfhout J W, Karbach J, Ringhoffer M, Jäger D, Arand M, Wada H, Noguchi Y, Stockert E, et al. J Exp Med. 1998;187:265–270. doi: 10.1084/jem.187.2.265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Jäger E, Nagata Y, Gnjatic S, Wada H, Stockert E, Karbach J, Dunbar P R, Lee S Y, Jungbluth A, Jäger D, et al. Proc Natl Acad Sci USA. 2000;97:4760–4765. doi: 10.1073/pnas.97.9.4760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Segal D M, Weiner G J, Weiner L M. Curr Opin Immunol. 1999;11:558–562. doi: 10.1016/s0952-7915(99)00015-1. [DOI] [PubMed] [Google Scholar]
- 61.Hudson P J. Curr Opin Immunol. 1999;11:548–557. doi: 10.1016/s0952-7915(99)00013-8. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}