

Abstract
Using pHluorin-tagged synaptic vesicle proteins we have examined the partitioning of these probes into recycling and non-recycling pools at hippocampal nerve terminals in cell culture. Our studies show that for 3 of the major synaptic vesicle components, vGlut-1, VAMP-2 and Synaptotagmin I, ∼50-60% of the tagged protein appears in a recycling pool that responds readily to sustained action potential stimulation by mobilizing and fusing with the plasma membrane, while the remainder is targeted to a non-recycling, acidic compartment. The fraction of recycling and non-recycling (or resting) pools varied significantly across boutons within an individual axon, from 100% resting (silent) to 100% recycling. Single bouton bleaching studies show that recycling and resting pools are dynamic and exchange between synaptic boutons. The quantitative parameters that can be extracted with the approaches outlined here should in the future help elucidate the potential functional role of the resting vesicle pool.
Keywords: synaptic vesicles, recycling, synapto-pHluorin, VAMP-2, VGlut
At nerve terminals, synaptic vesicles recycle in order to sustain neurotransmitter release during repetitive action potential (AP) firing. The size of the pool of vesicles that recycle and the kinetics of each of the recycling steps all play important roles in determining the efficacy of synaptic function during repetitive stimulation (Betz and Angleson 1998; Fernandez-Alfonso and Ryan 2006; von Gersdorff and Matthews 1999). Traditionally, vesicle pools at synaptic terminals have been defined based on kinetic criteria related to their depletion rates (Dobrunz and Stevens 1997; Elmqvist and Quastel 1965; von Gersdorff and Matthews 1997; Wu and Betz 1998). The readily-releasable pool is thought to correspond to vesicles in immediate apposition to the active zone (so-called docked vesicles) that undergo exocytosis during a small number of action potential stimuli. Beyond the active zone, presynaptic terminals have clusters of dozens to hundreds of synaptic vesicles that are recruited for exocytosis during prolonged stimulation and are referred to as the reserve pool (Neher 1998; Pieribone et al.1995; Stevens and Tsujimoto 1995). The entire recycling vesicle pool is comprised of both the readily-releasable and reserve pool of vesicles. FM dye labeling of synaptic vesicles in hippocampal nerve terminals show that the entire pool of recycling vesicles can be turned over with a few hundred stimuli, i.e. the amount of FM loaded saturates after a few hundred stimuli, and a similar number of stimuli causes >90% of the dye to be released (Ryan et al.1996; Ryan and Smith 1995). However, the number of vesicles loaded with such protocols appears significantly lower than those independently observed in electron micrographs (Murthy et al.1997; Murthy and Stevens 1999). Additionally, ultrastructural localization of FM dyes following photoconversion to an electron dense product, shows that only a fraction of the vesicles appear to have acquired dye even after loading under maximal stimulation (Harata et al.2001). Recently, using synapto-pHluorin to label synaptic vesicles, Li et al (Li et al.2005) recently suggested that some vesicles existed in a resting pool that would not fuse with the plasma membrane upon strong stimulation, however a detailed quantification of this pool or its dynamic across boutons and during long time scales was not provided. The simplest interpretation of these data is that a portion of the vesicle pool fails to recycle (Murthy et al.1997; Murthy and Stevens 1999; Sudhof 2000). Similar observations have been made in the Calyx of Held, a giant auditory brain stem synapse, where exhaustive FM labeling resulted in only a minor fraction of ultrastructurally-identifiable synaptic vesicle labeling (de Lange et al.2003). Here we have sought to examine the characteristics of both recycling and resting vesicles pools in a quantitative fashion in live cells. We made use of pHluorin-tagged synaptic vesicle proteins and made use of the pH-sensitivity of these probes to determine when and if they can be mobilized to fuse with the plasma membrane at synaptic terminals.
pHluorin is a modified form of GFP whose pH sensitivity has been shifted relative to that of EGFP by one log unit such that the pKa is ∼7.1 (Sankaranarayanan et al.2000). Upon binding protons the fluorescence is completely quenched, and therefore the fluorescence reflects the fraction of the time the fluorescent protein is in the proton-bound state. pHluorins can be targeted to the lumen of synaptic vesicles by fusion to one of the ∼10 identified synaptic vesicle proteins with membrane spanning domains (Takamori et al.2006). When expressed in neurons these proteins provide a useful readout of synaptic exocytosis and synaptic vesicle recycling. To date four of the most abundant synaptic vesicle proteins, VAMP-2 (Miesenbock et al.1998), vGlut-1 (Voglmaier et al.2006), Synaptotagmin I (Fernandez-Alfonso et al.2006) and Synaptophysin (Granseth et al.2006) have been successfully used for examining cycles of exo-endocytosis at nerve terminals with pHluorin-tagging.
During synaptic vesicle recycling, the lumen of the vesicle is reacidified by virtue of the action of a V-type ATPase that develops a pH gradient and the proton-motive force providing the energy to drive neurotransmitter uptake. When an inhibitor of this proton pump is applied to synapses during repetitive stimulation, vesicles continue to recycle, however they fail to reacidify and remain alkaline after endocytosis (termed alkaline trapping, (Sankaranarayanan and Ryan 2001). Resting vesicles remain acidic and can be quantified by use of NH4Cl that can rapidly alkanalize internal organelles. Comparison of the steady-state level of fluorescence achieved with pHluorin-tagged vesicle proteins during prolonged AP stimulation and alkaline trapping, with that achieved by direct alkalization of the entire pool vesicle pool provides a real time estimate of the fraction of the total pool of these vesicle proteins that remain acidic and hence fails to recycle.
Using these approaches, we show that a variable fraction of the total pool of the synaptic vesicle proteins VAMP-2, vGlut-1 and Synaptotagmin I do not participate in recycling during repetitive stimulation, but are dynamically exchanged between synapses.
Materials and Methods
Cell Culture and Experimental Conditions
Hippocampal CA3-CA1 regions were dissected from 2-4 day old Sprague-Dawley rats, dissociated, and plated onto poly-ornithine coated glass as previously described (Ryan 1999). Cultures were maintained at 37°C in a 95% air, 5% CO2 humidified incubator, and culture media was replaced every week. In spH experiments, calcium phosphate mediated gene transfer was used to transfect 6-8 day old cultures as previously described (Sankaranarayanan et al.2000) with super-ecliptic spH, vGpH or Tagim-pH. Super-ecliptic pHluorin is a modified version of the original ecliptic pHluorin with enhanced optical properties but an identical pKa (∼7.1) for protonation (Sankaranarayanan et al.2000). Cells were used 12-17 days after plating (6-12 days after transfection), and the coverslips were mounted in a rapid switching, laminar-flow perfusion and stimulation chamber on the stage of a custom-built laser scanning confocal microscope. The total volume of the chamber was ∼75 μl and was perfused at a rate of 1-1.5 ml/min. Action potentials were evoked by passing 1 ms current pulses, yielding fields of ∼10 V/cm through the chamber via Platinum-Iridium electrodes. Except otherwise noted, cells were continuously perfused at room temperature (∼24°C) in a saline solution containing in mM: 119 NaCl, 2.5 KCl, 2 CaCl2, 2 MgCl2, 25 HEPES (buffered to pH 7.4), 30 glucose, 10 μM 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX; Research Biochemicals, Natick, MA) and 50 μM D,L-2-amino-5-phosphonovaleric acid (AP5; Research Biochemicals). When external CaCl2 was varied, the total divalent ion concentration was kept constant by simultaneously changing MgCl2. NaCl and CaCl2 were replaced with KCl (keeping the total ion concentration fixed) during application of Monensin (40 μM, CalBiochem) and Nigericin (10 μM, Molecular Probes), two carboxylic ionophores that together allow the exchange of cations for protons across cell membranes (both drugs were stored frozen as 10 mM stock solutions in MeOH). When NH4Cl (50 mM) was added to the saline, NaCl was reduced to 70 mM. Bafilomycin A1 (Baf; CalBiochem, San Diego, CA) was stored frozen as 0.2 mM aliquots (in DMSO) and diluted to a final concentration of 0.5 μM prior to spH experiments. In Figure1b, FM 4-64 (Molecular Probes, Eugene, OR) were used at a final concentration of 15 μM and vesicles were labeled by electrical field stimulation for 30s at 20 Hz in the presence of the dye in normal saline, followed by an additional 60 s of dye exposure to ensure labeling of all recycling membrane during endocytosis. Unless otherwise noted all chemicals were obtained from Sigma (St. Louis, MO).
Figure 1. Quantitative estimates of recycling and resting vesicle pool fractions in live cells.
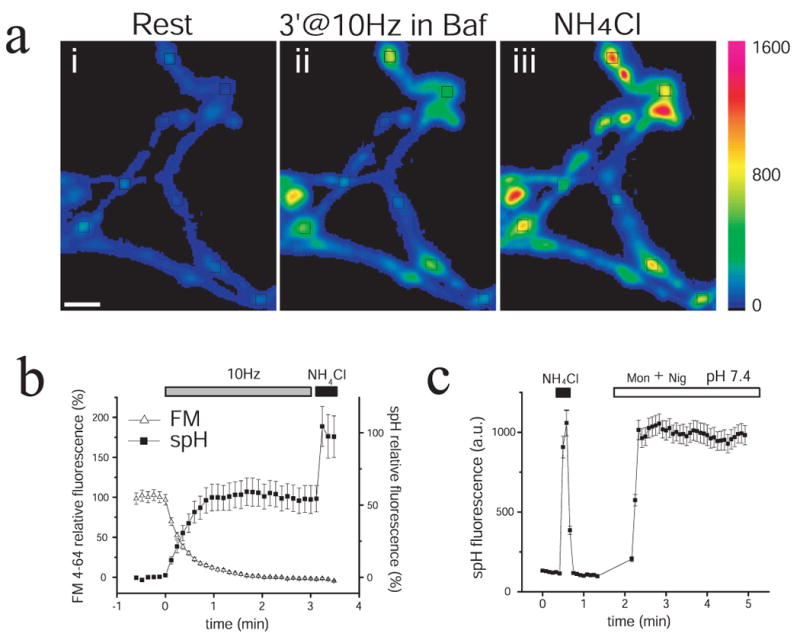
(a) Pseudo-color images of a representative axonal branch from a spH-transfected hippocampal neuron, during rest (i), after field stimulation with 1800 APs in the presence of 0.5μM Bafilomycin (Baf) (ii), and during a subsequent application of NH4Cl (50mM, pH 7.4) to equilibrate synaptic-vesicle lumen pH to 7.4 (iii). Black boxes represent examples of areas selected for analysis. Only a subset is shown for clarity. Scale bar, 3μm. Vertical pseudo-color scale shows spH fluorescence intensity values (arbitrary units, a.u.).
(b) Representative experiment showing the protocol used to visualize the fraction of the total pool of synaptic vesicles participating in recycling. (Solid squares) Average spH fluorescence response of one spH-transfected neuron (n = 49 boutons) to 1800 APs (10Hz, grey bar) normalized to the fluorescence level obtained during a subsequent NH4Cl exposure (black bar). Baf (0.5μM) was present throughout the experiment to prevent reacidification of vesicles after endocytosis (alkaline trapping); therefore any increase in spH fluorescence during stimulation reflects new vesicles undergoing exocytosis. The increase in fluorescence reached a steady-state level after ∼ 600 AP, indicating the time required to turnover the recycling pool of vesicles. NH4Cl application produced a further increase in fluorescence, reflecting the equilibration of resting vesicles to pH 7.4. FM 4-64 unloading (open triangles) was simultaneously recorded in the same set of boutons, which were preloaded with dye in a previous round of stimulation. The time to reach 50% recycling pool turnover was identical based on either spH or FM 4-64 responses (∼180 AP).
(c) NH4Cl effectively “unmasks” the total pool of synaptic vesicles present at axon terminals. The spH fluorescence level reached with NH4Cl (black bar) is the same as that reached when the pH of the vesicles was equilibrated to the extracellular pH (7.4) by applying 40μM Monensin and 10μM Nigericin (Mon + Nig, white bar), which allow neutral exchange of cations for protons across cell membranes.
Optical Measurements, Microscopy and Analysis
Laser-scanning fluorescence images were acquired using a custom-built laser-scanning microscope through a 40× 1.3 numerical aperture Zeiss Fluar objective (Oberkochen, Germany). Specimens were illuminated with ∼1.5 μwatts of the 488 nm line of an argon ion laser that was rapidly shuttered during all non-data acquiring periods using acousto-optic or electro-optic modulation. Time course of fluorescence response of spH and FM dyes was measured from time-lapse images taken every 3-10s. SpH fluorescence emission was collected using a 500 nm long pass filter (except for combined FM 4-64 and spH experiments, where a 498-538 nm band pass filter was used for spH); FM 4-64 emission was collected using a 650 nm long pass filter. Quantitative measurements of fluorescence intensity at individual boutons were obtained by averaging 4 × 4 area pixels (0.4 × 0.4 μm2) corresponding to the center of mass of 8×8-pixels positioned over individual puncta selected by hand from images (Figure 1a shows a subset of areas selected). Further criteria for including a fluorescent puncta as a synaptic bouton in our analysis, included requiring a minimum of a 1.5 fold increase in fluorescence upon alkalization with NH4Cl compared to that at rest ([FNH4Cl - Frest] / Frest). Fluorescence values for baseline, prolonged stimulation and NH4Cl were obtained by averaging 2 to 5 images at rest, after reaching a steady-state level during stimulation (10 or 20Hz), and during NH4Cl exposure respectively. The noise in our estimates of fraction recycling was calculated from fluorescence difference of the average of the first three images and the average of the following three images taken prior to stimulation (6 images were taken before stimulation), relative to the average change in fluorescence in NH4Cl (i.e., [F1-3 − F4-6] / ΔFNH4Cl). The standard deviation of the estimated noise was 0.11 (n = 148 boutons), therefore the fraction recycling was binned in 0.1 intervals. 4 out of 22 neurons, which had high basal rates of fluorescence increase on bafilomycin application were not included in the final analysis, since we could not distinguish with confidence whether the rise in fluorescence was due to spontaneous alkalization of vesicles or to spontaneous vesicle fusions (Sankaranarayanan and Ryan 2001). Data points that presented a fraction recycling of less than -0.1 or more than 1.1 probably represented slight shifts of boutons from focal plane and were not included in our analysis (55 out of 1085 boutons in Figure 3a). ΔF for each bouton was always normalized to ΔFNH4Cl before any further analysis. In Figure 5, bleaching was achieved by scanning the laser beam over a region of interest encompassing a single bouton (∼10 times more power and scanning rates than for regular imaging) in the presence of NH4Cl for ∼30 seconds. Errors are shown as standard error of mean except where stated.
Figure 3. The recycling vesicle fraction varies across different cells and synapses but represents on average 50% of the total pool.

(a) Measurements of average fraction of recycling vesicles for spH (n=10), synaptotagmin-pHluorin (tagmin-pH, n=4) and vGlut1-pHluorin (vGpH, n=5) are indistinguishable (p=0.05 ANOVA).
(b) Distribution of vesicle pool recycling fraction across different boutons in 18 cells from independent cultures. Each solid symbol represents a single measurement at a single bouton. The average recycling fraction and its standard deviation for each cell is shown as open triangles and grey bars respectively (n = 36-96 boutons in each cell).
(c) The average fraction recycling for each experiment is plotted as a function of the age in vitro of the individual culture. The fraction recycling did not correlate with age in culture for the range of ages analyzed (12 to 17 days in vitro).
(d) Boutons containing larger vesicles pools have a slight tendency to have higher recycling fractions. The increase in spH fluorescence in response to NH4Cl application (ΔF NH4Cl) gives an estimate of the size of the total vesicle pool. To compare across different neurons, this value was normalized to the average ΔF NH4Cl in each independent experiment. The grey line shows a linear fit to the data (R = 0.09; p<0.01).
Figure 5. Recycling and resting vesicle pools interchange across boutons.
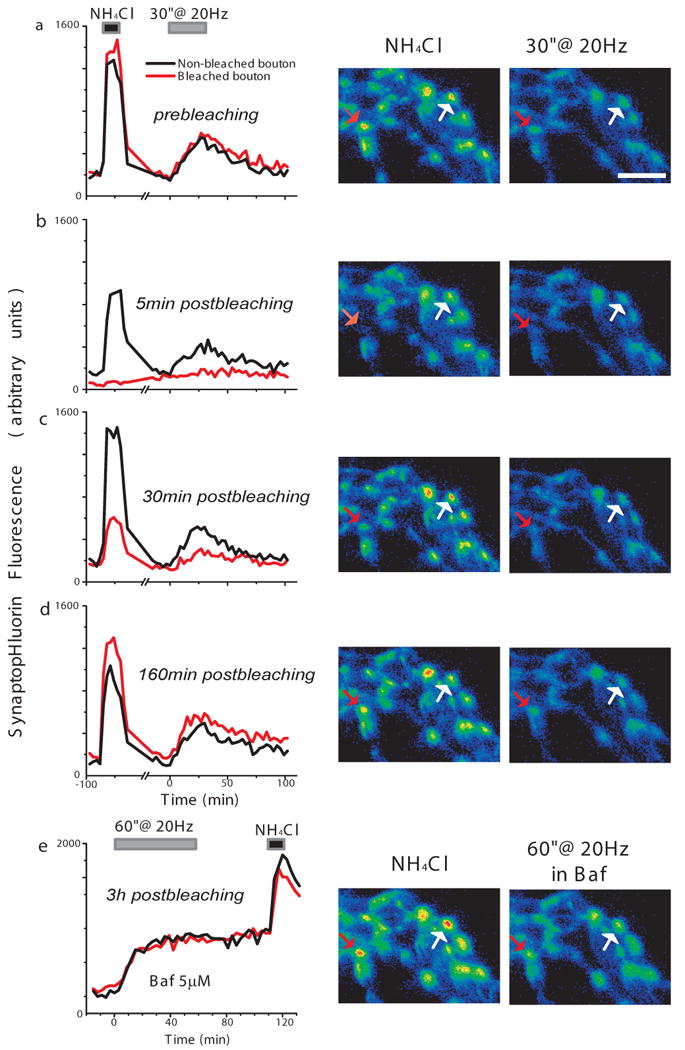
After assessing the size of the total vesicle pool with NH4Cl and estimating its recycling pool with a train of stimulation (a), all vesicles in a single bouton where bleached (red arrow in right panel pictures, red trace in left plots). (b-d) The same protocol applied 5 (b), 30 (c), and 160 (d) minutes after bleaching indicates that non-bleached vesicles reached the originally bleached bouton and distribute in both recycling and resting pools. (e) After 3 hours, non-bleached vesicles account for 91% of originally bleached ones (NH4Cl, compare with a); of these, 51% are capable of recycling (stimuli in Baf). The response to the same series of trials is also shown for a neighboring, non-bleached, bouton (a-e, white arrow in right panel pictures, black traces in left plots). Scale bar 5μm.
Results
Measuring the fraction of the total pool of vesicles that participates in recycling
The main goal of this work was to quantify the fraction of the synaptic vesicle (SV) pool located at presynaptic terminals that participates in recycling during prolonged action potential (AP) firing. We developed a protocol using pHluorin as an optical tracer to visualize and quantify both the total SV pool and the fraction of it that can be engaged to recycle. We have explicitly assumed that synaptic vesicles are the only spH-containing organelles in synaptic terminals (see Discussion). Figure 1a shows confocal images of a rat hippocampal neuron in culture transfected with spH, depicting GFP fluorescence intensity in a pseudo-color scale. The fluorescence of this pH-sensitive GFP fused to the lumenal aspect of VAMP-2 is largely quenched at the resting lumenal pH (5.5), and undergoes an ∼ 20 fold increase in its fluorescence when the pH shifts to 7.4 upon exocytosis (Miesenbock et al.1998; Sankaranarayanan et al.2000). The fluorescence observed at rest (panel i in Figure 1a) largely reflects VAMP-2 on the plasma membrane surface. Superfusion with NH4Cl (50mM) equilibrates the SV lumen to a pH of 7.4 (Miesenbock et al.1998), thus unmasking clusters of vesicles located in axon terminals and allowing the visualization of individual boutons as puncta of fluorescence (panel iii). These puncta were chosen as regions of analysis in subsequent studies (square boxes in Figure 1a, see Experimental Procedures). Synaptic vesicles are normally re-acidified after endocytosis by a V-type ATPase whose action can be blocked by the reversible cell-permeant inhibitor bafilomycin (baf). Measurements of spH fluorescence during AP stimulation in baf provide a direct measure of exocytosis, independent of any endocytic event, since vesicles become trapped in the alkaline state during recycling (alkaline trapping). Bafilomycin does not significantly perturb exocytosis or endocytosis in this system (Cousin and Nicholls 1997; Li et al.2005; Sankaranarayanan and Ryan 2001; Zhou et al.2000). In order to visualize the pool of vesicles that can be engaged to recycle, neurons were field-stimulated with 1800 APs at 10Hz (which far exceeds the stimulus required to turnover the entire recycling pool of SVs, see below) in the presence of baf (panel ii). The size of the vesicle pool that participates in recycling can be estimated for each bouton by subtracting the fluorescence during rest from that obtained after the stimulus (ΔF1800AP, i.e., panel ii – panel i). A similar subtraction can be done to the fluorescence in NH4Cl to get the size of the total vesicle pool (ΔFNH4Cl, i.e., panel iii – panel i). Finally, the fraction of the total pool of vesicles recycling can be calculated for each bouton as ΔF1800AP / ΔFNH4Cl. Figure 1b shows the spH fluorescence increase during stimulation in the presence of baf (alkaline trapping) and a subsequent application of NH4Cl, plotted as a function of time. A steady-state level is reached after ∼ 600 APs, reflecting the stimulation required to turnover the entire recycling pool (Fig. 1b, solid squares). This complete turnover was confirmed by simultaneously measuring FM 4-64 destaining from the same set of boutons (Fig. 1b, open triangles), which were preloaded with the dye in a previous round of stimulation. FM 4-64 destaining kinetics agreed quantitatively very well with the kinetics of alkaline-trapped spH fluorescence increase (time to half FM signal decay = 18s, time to half spH signal increase = 20s), further supporting the idea that this methodology provides a robust measure of AP-driven synaptic vesicle pool turnover. In general, this steady-state level in spH fluorescence was no different if neurons were stimulated at a frequency of 20Hz and it was reached within 600 to 1200 APs, in agreement with previous reports (Ryan and Smith, 1995). In order to confirm that NH4Cl (50mM, pH 7.4) effectively equilibrates the lumen of synaptic vesicles with the extracellular pH, the peak fluorescence level reached in NH4Cl was compared with that reached when a combination of ionophores that allow free exchange of cations for protons across cell membranes was applied. Figure 1c shows such an experiment. Monensin (40μM) and nigericin (10μM) were perfused in a Ca2+ and Na+-free (replaced with K+) saline to prevent vesicle fusion and allow pH equilibration across all membranous compartments (Lee and Epel 1983; Lemasters et al.1995). spH reached the same fluorescence level in both conditions (peak mon+nig / peak NH4Cl = 0.96 ± 0.1, Mean ± SD of 6 independent experiments),, indicating that pH inside SV-lumen is effectively equilibrated with the extracellular pH during NH4Cl exposure. Taken together, these experiments indicate that spH fluorescence values reached during prolonged stimulation in baf, as compared to that reached under NH4Cl, provides a good estimate of the fraction of the total pool of synaptic vesicles that can be engaged in recycling as well as the fraction that remains acidic and hence refractory to recycling.
The fraction of recycling vesicles varies from synapse to synapse and neuron to neuron
The distribution of recycling vesicle fractions in a population of 47 boutons arising from an individual neuron is presented in Figure 2. For each bouton, the spH ΔF during both alkaline trapping and NH4Cl application is depicted in Figure 2a. Displaying the fraction recycling as a function of the entire vesicle pool shows that that terminals with larger vesicle pools do not necessarily have larger recycling fractions (Fig. 2b, but see also below and Fig. 3d). Figure 2c shows the frequency distribution of the fraction recycling for the 47 boutons analyzed. The percentage of vesicles recycling for this neuron varied from 0% to 100% for different boutons, with a mean of 54% with a standard deviation of 27 %. There were no obvious differences in the kinetics of vesicle pool mobilization and fusion among boutons that made use of a small, a medium or a large fraction of its total pool to release neurotransmitter (0.1-0.4, 0.4 0.7 and 0.7-1 fraction groups respectively), suggesting that the fraction recycling is independent of the efficiency of the fusion machinery (Fig. 2c). We found the average value to be 50% ± 3% across 18 cells (Fig. 3a,b). Using vGlut-pHluorin (vGpH) or synaptotatmin I-pHluorin (tagmin-pH) as reporters in identical experiments we found the average recycling pool size was very similar (Fig 3a, 60.1 ±9.9%, vGpH, n=5; 51.0 ± 10 % tagmin-pH, n=4 cells). Analysis of recycling fractions recycling over many cells shows that in addition to varying across boutons the mean recycling fraction also varies from neuron to neuron (Fig. 3b). The average fraction recycling was 22% and 67% for the neurons presenting the smallest and the highest values respectively (cells number 14 and 11 respectively, see Fig. 3b). Similar ranges were obtained for vGpH and tagmin-pH. We also assessed whether the fraction recycling varied with age of cultures. It was recently reported that the amount of FM 1-43 uptake is two-fold higher in neuronal cultures of 15 days versus 12 days in vitro (Renger et al., 2001) suggesting that the number of vesicles recycling increases during synapse maturation. Contrary to what we expected, we found no increase in the fraction recycling in more mature cultures (Fig. 3c). However, it should be noted that if the total pool of vesicles increased proportionally to the recycling pool, then this fraction would not be affected (see above). We also tested the hypothesis that boutons containing larger vesicle pools are able to engage a larger fraction of the pool in recycling. There is a very small (R = 0.09) but significant (p < 0.01) tendency of larger pools to recycle a larger fraction of its vesicles (Fig. 3d). Taken together, these experiments indicate that the fraction of the total pool of vesicles that recycles varies across synapses (0% to 100%) and across neurons (∼ 20 to 70%), that it is not correlated with the age of the culture and that terminals with more vesicles are likely to recycle a larger fraction of its vesicles.
Figure 2. Individual bouton analysis of the recycling vesicle pool fraction.

The study of 47 boutons corresponding to one neuron (same as that presented in Figure 1a) is shown.
(a) The increase in spH fluorescence (with respect to the baseline at rest, ΔF spH) after 1800 action potentials in baf (solid squares) and after NH4Cl perfusion (open circles) is shown for each bouton. The length of the line linking the symbols represents the size of the resting pool of vesicles. ΔF1800AP / ΔFNH4Cl was calculated to get the fraction recycling in (b) and (c).
(b) The fraction of vesicles recycling is plotted as a function of the total vesicle pool size as reflected in the fluorescence increase in the presence of NH4Cl.
(c) Cumulative frequency histogram of the fraction of the total pool of vesicles that participates in recycling during prolonged stimulation (fraction recycling). The fraction recycling varied from 0 to 1 with an average value of 0.54 ± 0.04.
(d) The fraction recycling does not appear to be related to the efficiency of the fusion machinery. Boutons were pooled in 3 groups according to the fraction of vesicles recycling: 0.1 to 0.4 (solid squares, n=11); 0.4 to 0.7 (solid circles, n=17); and 0.7 to 1 (solid triangles, n=14). The average normalized fluorescence for each group is plotted as a function of time (dotted line delimits the 3 groups). The kinetics of exocytosis during stimulation (3min. at 10Hz in baf, gray bar) is similar in the 3 groups (τ0.1-0.4 = 32 s, τ0.4-0.7 =37 s, τ0.7-1 =32 s).
The fraction of vesicles recycling varies little in the time scale of hours
The heterogeneity in the fraction of vesicles recycling across boutons from a single cell (Fig. 3) may indicate that, at the single bouton level, the fraction of vesicles recycling could be modulated over time. To test this hypothesis, we measured the change in the fraction recycling at single boutons over a period of 3 hours. Figure 4a shows that the average fraction of all boutons remains stable over 3 hours. Interestingly, some boutons showed large variations in the fraction recycling (Fig. 4b, black bars). However these represented only 19% of all boutons, and included increasing and decreasing fraction of recycling vesicles (Fig. 4b). Thus, although a few boutons present large variation in the fraction recycling over time, the overall fraction across all boutons appeared to remain stable over a period of hours.
Figure 4. The fraction of vesicles recycling is stable over a 3 hour time period.

(a) The average fraction of vesicles recycling measured in two consecutive trials with an interval of 3 hours indicates that recycling and resting vesicles proportions are stable over time. Recycling fractions are 53% ± 2% and 47% ± 2% in the first and second trial respectively (n=16 boutons).
(b) The majority of boutons showed little variation (<10%) in the recycling fraction (13 boutons, dark grey bars). A few presented a larger variation (between 15% and 30%, n=3 boutons, light grey bars). The error bars are the standard deviation of the steady state fluorescence after all recycling vesicles were trapped in the alkaline state during the stimulus (2%-6%). The inset depicts the example of individual boutons presenting a 4% increase (I) and 29% decrease (II) in their recycling fraction.
Resting and recycling vesicles are interchanged across boutons
The stability in the recycling and resting vesicle pools (Fig. 4) may indicate that vesicles remain stably bound at their terminals. Alternatively this may reflect a steady state behavior, with a constant flux of vesicles into and out of these pools, interchanging across boutons or even between recycling and resting pools within boutons while maintaining a constant relative abundance. To test these possibilities, we developed an approach to photobleach the fluorescence of the entire pool of vesicles (i.e., recycling and resting) in individual boutons and examined recovery of fluorescence over time of each of the pools (Fig. 5). We reasoned that photobleaching the spH fluorescence in the presence of NH4Cl at an individual bouton would irreversibly mark vesicles in both recycling and resting pools. A typical bleaching event is shown in Fig 5. Prior to photobleaching the boutons show a robust NH4Cl response reflecting the alkalinization of both the recycling and resting vesicles. Following NH4Cl washout and a return to baseline fluorescence, the recycling pool can be mobilized to fuse with the plasma membrane with a 30 s 20 Hz stimulus. Individual boutons were then photobleached during a second NH4Cl challenge. 5 min after this bleaching episode the NH4Cl response at the bleached bouton was eliminated indicating both recycling and resting pools were bleached. A second round of AP stimulation gave rise to only a minimal signal, likely corresponding to diffusional spillover from adjacent non-bleached boutons (Fernandez-Alfonso et al.2006; Li and Murthy 2001; Sankaranarayanan and Ryan 2000; Wienisch and Klingauf 2006). Nearby boutons in the field retained both their NH4Cl signal as well as the stimulus driven recycling pool signal. Repeated querying of fluorescence with NH4Cl and AP firing at different times after bleaching revealed that the fluorescence of both pools gradually returned to original values reaching near-completion within ∼3 hours (87% ± 7% of total pool, and 81% ± 11% recycling fluorescence recovered, n=3 boutons). A final stimulation in the presence of bafilomycin followed by NH4Cl confirmed that both recycling and resting vesicles are likely under dynamic exchange with neighboring boutons (Fig. 5e).
Discussion
We used a novel strategy to determine what fraction of vesicle pools in living synaptic terminals participates in vesicle recycling during prolonged stimulation. We relied upon measurements of genetically encoded tracers of synaptic vesicles, pHluorin-tagged synaptic vesicle proteins, to determine what portion of labeled vesicle pools can engage in recycling. In resting terminals, spH fluorescence is quenched at the low vesicular pH (∼5.5). During recycling in the presence of bafilomycin, a blocker of the vesicular proton pump responsible for vesicle reacidification, vesicles become trapped in the alkaline (pH 7.4) and thus fluorescent state. If all labeled vesicles in the pool recycle, then the fluorescence should reach a value that would be equivalent to direct alkalization of vesicles to pH 7.4 (via either NH4Cl or protonophores). Any pHluorin-tagged protein that fails to recycle will remain acidic and maintain a low fluorescence value.
The robustness of our estimates of synaptic vesicle pool recycling fractions with pHluorin-tagged proteins relies on two main assumptions. First that the proteins we examined, VAMP-2, vGlut-1 and Synaptotagmin I, and their pHluorin-tagged variants, reside exclusively in synaptic vesicles at synaptic terminals. An alternate possibility is that some of the reporters may be targeted to endosomes as has been suggested for VAMP-2 (West et al.1997). We feel this is unlikely since presynaptic spH appears to maintain a resting pH similar to that of synaptic vesicles, not endosomes (Miesenbock et al.1998; Mukherjee et al.1997; Sankaranarayanan et al.2000), although the definition of an endosome could potentially be expanded to include the resting synaptic vesicle proteins we see here. The second assumption is that pHluorin-tagged proteins are randomly distributed between recycling and resting vesicle pools. At present, it is very difficult to explicitly test this hypothesis. However, even if the expression of pHluorin-tagged sv proteins is biased towards one pool or another, the basic observation that a large fraction of the vesicles seem to be held in a state of “deep” reserve or are simply not competent for recycling will not change. We recently demonstrated that the expression of both spH (Fernandez-Alfonso et al.2006) and vGpH (Balaji and Ryan 2007) results in only 1-2 copies of the pHluorin-tagged protein per vesicle, much lower than the number of endogenous proteins likely to be present (Takamori et al.2006). Thus it seems unlikely that the appearance of these reporters in a resting compartment results from miss-targeting. Similarly, the fact that the most abundant synaptic vesicle proteins display similar partitioning argues that this is likely a bona-fide property of synaptic vesicles. Since we are measuring recycling vesicles relative to the total pool, it should be noted that, for example, a “silent” bouton does not necessarily mean a low fraction of vesicles recycling (i.e., a bouton that has a few vesicles recycling, although silent, will have a large fraction recycling if the total pool number is also very small; see Figure 2a).
Our results are in reasonable agreement with other approaches for examining recycling vesicles based on photoconversion of FM 1-43 and electron microscopy (Harata et al.2001). However, FM photoconversion experiments do not distinguish whether non-stained vesicles are ones that fail to recycle or recycle in a manner that excludes dye labeling (i.e., kiss and run fusion). Since our method relies only on the exchange of protons within vesicles with the extracellular medium to track exocytosis, we conclude that resting vesicles are indeed ones that do not fuse with the plasma membrane. Our approach also allows analyzing synaptic vesicle recycling in a vast number of synapses simultaneously, in living specimens, and in real-time. We believe that the methods introduced in this work should add substantially in determining how various modulators of presynaptic function regulate recycling pool sizes.
Our photobleaching experiments indicate that both recycling and resting vesicle pools appear to exchange between synaptic boutons. Even at intermediate time points in the photobleaching recovery, the relative size of the recycling and resting pools (as judged by the peak of the NH4Cl compared to the peak of the stimulus response) appears constant. This suggests that the two pools are always closely linked, perhaps even constantly interchanging within a given synapse over these time scales. Recently it was demonstrated that FM 1-43-labeled and thus by definition recycling vesicles, exchange between boutons (Darcy et al.2006). Our results compliment those studies and demonstrate that this sharing of vesicles extends to non-recycling vesicles as well. Here our experiments do not distinguish between possible recovery by local interchange with a recycling pool (which itself can recover by surface diffusional exchange of spH or the slower intracellular exchange) or long range mobility of resting vesicles. It is interesting to point out that the time scale of recovery we see is considerably slower than the time scale of movement of recycling vesicle between boutons observed by Darcy et al.
In Drosophila neuromuscular junctions, two spatially segregated pools of vesicles have been characterized; one located in the periphery and the other in the central region of presynaptic terminals (Kuromi and Kidokoro 1998). Interestingly, the central one was shown to participate in recycling only after high frequency stimulation or activation of protein kinase A (Kuromi and Kidokoro 2000). Results using an approach very similar to ours with pHluorin and alkaline trapping has also been used in drosophila, which confirmed the existence of a synaptic vesicle pool that could only successfully be mobilized following endocytic block in the shibire mutant at non-permissive temperatures (Poskanzer and Davis 2004), which had previously been observed using other methods (Delgado et al.2000). It seems that there exists a pool of vesicles in a “deep reserve” state that can be switched to participate in recycling and is conserved across species for regulating synaptic transmission. It is important to note, however, that we did not find any obvious spatial distinction of the two pools in our experiments (compare panels ii and iii in Figure 1a). Although it is true that our system does not provide high enough resolution to detect fine spatial distributions, electron micrographs of FM photoconversion experiments suggest that recycling and resting vesicles are distributed at random in mammalian CNS (Harata et al.2001).
In conclusion, we show that a varying fraction of synaptic vesicles remain in a resting state (i.e., do not participate in recycling) during prolonged stimulation at CNS synapses. Although in a resting state, it is tempting to consider that part of this pool could be tapped to undergo recycling, that would imply that synapses have a large reserve capacity that might be used to modulate presynaptic function. It is also tempting to speculate that this resting pool might not only serve to regulate synaptic efficacy in individual boutons, but also be exchanged between synapses under some conditions. At present we do not know what physiological signal transduction pathways might normally control the balance of recycling and resting pools, however, the methods used here should prove useful in exploring how and when this “untapped” synaptic resource is utilized.
Acknowledgments
We would like to thank Dr. James Rothman for kindly providing the super-ecliptic synapto-pHluorin and tagmin-pHluorin constructs, Robert Edwards and Susan Voglemeier for the vGlut-1-pHluorin construct, members of the Ryan lab for useful discussions, and Wayne Yan and Ricky Kwan for excellent technical assistance. Data using vGlut-pHluorin and tagmin-pHluorin were kindly provided by Sung Yung Kim and Pablo Ariel. Sethu Sankaranarayanan kindly provided some of the data using spH. This work was supported by NIH (NS24992) a McKnight Technological Innovations in Neuroscience Award and an Irma T. Hirshcl career scientist award to T.A.R.
References
- Balaji J, Ryan TA. Single-vesicle imaging reveals that synaptic vesicle exocytosis and endocytosis are coupled by a single stochastic mode. Proc Natl Acad Sci U S A. 2007;104(51):20576–81. doi: 10.1073/pnas.0707574105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Betz WJ, Angleson JK. The synaptic vesicle cycle. Annu Rev Physiol. 1998;60:347–63. doi: 10.1146/annurev.physiol.60.1.347. [DOI] [PubMed] [Google Scholar]
- Cousin MA, Nicholls DG. Synaptic vesicle recycling in cultured cerebellar granule cells: role of vesicular acidification and refilling. J Neurochem. 1997;69(5):1927–35. doi: 10.1046/j.1471-4159.1997.69051927.x. [DOI] [PubMed] [Google Scholar]
- Darcy KJ, Staras K, Collinson LM, Goda Y. Constitutive sharing of recycling synaptic vesicles between presynaptic boutons. Nat Neurosci. 2006;9(3):315–21. doi: 10.1038/nn1640. [DOI] [PubMed] [Google Scholar]
- de Lange RP, de Roos AD, Borst JG. Two modes of vesicle recycling in the rat calyx of Held. J Neurosci. 2003;23(31):10164–73. doi: 10.1523/JNEUROSCI.23-31-10164.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delgado R, Maureira C, Oliva C, Kidokoro Y, Labarca P. Size of vesicle pools, rates of mobilization, and recycling at neuromuscular synapses of a Drosophila mutant, shibire. Neuron. 2000;28(3):941–53. doi: 10.1016/s0896-6273(00)00165-3. [DOI] [PubMed] [Google Scholar]
- Dobrunz LE, Stevens CF. Heterogeneity of release probability, facilitation, and depletion at central synapses. Neuron. 1997;18(6):995–1008. doi: 10.1016/s0896-6273(00)80338-4. [DOI] [PubMed] [Google Scholar]
- Elmqvist D, Quastel DM. A quantitative study of end-plate potentials in isolated human muscle. J Physiol. 1965;178(3):505–29. doi: 10.1113/jphysiol.1965.sp007639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez-Alfonso T, Kwan R, Ryan TA. Synaptic vesicles interchange their membrane proteins with a large surface reservoir during recycling. Neuron. 2006;51(2):179–86. doi: 10.1016/j.neuron.2006.06.008. [DOI] [PubMed] [Google Scholar]
- Fernandez-Alfonso T, Ryan TA. The efficiency of the synaptic vesicle cycle at central nervous system synapses. Trends Cell Biol. 2006;16(8):413–20. doi: 10.1016/j.tcb.2006.06.007. [DOI] [PubMed] [Google Scholar]
- Granseth B, Odermatt B, Royle SJ, Lagnado L. Clathrin-mediated endocytosis is the dominant mechanism of vesicle retrieval at hippocampal synapses. Neuron. 2006;51(6):773–86. doi: 10.1016/j.neuron.2006.08.029. [DOI] [PubMed] [Google Scholar]
- Harata N, Ryan TA, Smith SJ, Buchanan J, Tsien RW. Visualizing recycling synaptic vesicles in hippocampal neurons by FM 1-43 photoconversion. Proc Natl Acad Sci U S A. 2001;98(22):12748–53. doi: 10.1073/pnas.171442798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuromi H, Kidokoro Y. Two distinct pools of synaptic vesicles in single presynaptic boutons in a temperature-sensitive Drosophila mutant, shibire. Neuron. 1998;20(5):917–25. doi: 10.1016/s0896-6273(00)80473-0. [DOI] [PubMed] [Google Scholar]
- Kuromi H, Kidokoro Y. Tetanic stimulation recruits vesicles from reserve pool via a cAMP-mediated process in Drosophila synapses. Neuron. 2000;27(1):133–43. doi: 10.1016/s0896-6273(00)00015-5. [DOI] [PubMed] [Google Scholar]
- Lee HC, Epel D. Changes in intracellular acidic compartments in sea urchin eggs after activation. Dev Biol. 1983;98(2):446–54. doi: 10.1016/0012-1606(83)90374-3. [DOI] [PubMed] [Google Scholar]
- Lemasters JJ, Chacon E, Ohata H, Harper IS, Nieminen AL, Tesfai SA, Herman B. Measurement of electrical potential, pH, and free calcium ion concentration in mitochondria of living cells by laser scanning confocal microscopy. Methods Enzymol. 1995;260:428–44. doi: 10.1016/0076-6879(95)60156-2. [DOI] [PubMed] [Google Scholar]
- Li Z, Burrone J, Tyler WJ, Hartman KN, Albeanu DF, Murthy VN. Synaptic vesicle recycling studied in transgenic mice expressing synaptopHluorin. Proc Natl Acad Sci U S A. 2005;102(17):6131–6. doi: 10.1073/pnas.0501145102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Z, Murthy VN. Visualizing postendocytic traffic of synaptic vesicles at hippocampal synapses. Neuron. 2001;31(4):593–605. doi: 10.1016/s0896-6273(01)00398-1. [DOI] [PubMed] [Google Scholar]
- Miesenbock G, De Angelis DA, Rothman JE. Visualizing secretion and synaptic transmission with pH-sensitive green fluorescent proteins. Nature. 1998;394(6689):192–5. doi: 10.1038/28190. [DOI] [PubMed] [Google Scholar]
- Mukherjee S, Ghosh RN, Maxfield FR. Endocytosis. Physiol Rev. 1997;77(3):759–803. doi: 10.1152/physrev.1997.77.3.759. [DOI] [PubMed] [Google Scholar]
- Murthy VN, Sejnowski TJ, Stevens CF. Heterogeneous release properties of visualized individual hippocampal synapses. Neuron. 1997;18(4):599–612. doi: 10.1016/s0896-6273(00)80301-3. [DOI] [PubMed] [Google Scholar]
- Murthy VN, Stevens CF. Reversal of synaptic vesicle docking at central synapses. Nat Neurosci. 1999;2(6):503–7. doi: 10.1038/9149. [DOI] [PubMed] [Google Scholar]
- Neher E. Vesicle pools and Ca2+ microdomains: new tools for understanding their roles in neurotransmitter release. Neuron. 1998;20(3):389–99. doi: 10.1016/s0896-6273(00)80983-6. [DOI] [PubMed] [Google Scholar]
- Pieribone VA, Shupliakov O, Brodin L, Hilfiker-Rothenfluh S, Czernik AJ, Greengard P. Distinct pools of synaptic vesicles in neurotransmitter release. Nature. 1995;375(6531):493–7. doi: 10.1038/375493a0. [DOI] [PubMed] [Google Scholar]
- Poskanzer KE, Davis GW. Mobilization and fusion of a non-recycling pool of synaptic vesicles under conditions of endocytic blockade. Neuropharmacology. 2004;47(5):714–23. doi: 10.1016/j.neuropharm.2004.07.026. [DOI] [PubMed] [Google Scholar]
- Ryan TA. Inhibitors of myosin light chain kinase block synaptic vesicle pool mobilization during action potential firing. J Neurosci. 1999;19(4):1317–23. doi: 10.1523/JNEUROSCI.19-04-01317.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryan TA, Li L, Chin LS, Greengard P, Smith SJ. Synaptic vesicle recycling in synapsin I knock-out mice. J Cell Biol. 1996;134(5):1219–27. doi: 10.1083/jcb.134.5.1219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryan TA, Smith SJ. Vesicle pool mobilization during action potential firing at hippocampal synapses. Neuron. 1995;14(5):983–9. doi: 10.1016/0896-6273(95)90336-4. [DOI] [PubMed] [Google Scholar]
- Sankaranarayanan S, De Angelis D, Rothman JE, Ryan TA. The use of pHluorins for optical measurements of presynaptic activity. Biophys J. 2000;79(4):2199–208. doi: 10.1016/S0006-3495(00)76468-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sankaranarayanan S, Ryan TA. Real-time measurements of vesicle-SNARE recycling in synapses of the central nervous system. Nat Cell Biol. 2000;2(4):197–204. doi: 10.1038/35008615. [DOI] [PubMed] [Google Scholar]
- Sankaranarayanan S, Ryan TA. Calcium accelerates endocytosis of vSNAREs at hippocampal synapses. Nat Neurosci. 2001;4(2):129–36. doi: 10.1038/83949. [DOI] [PubMed] [Google Scholar]
- Stevens CF, Tsujimoto T. Estimates for the pool size of releasable quanta at a single central synapse and for the time required to refill the pool. Proc Natl Acad Sci U S A. 1995;92(3):846–9. doi: 10.1073/pnas.92.3.846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudhof TC. The synaptic vesicle cycle revisited. Neuron. 2000;28(2):317–20. doi: 10.1016/s0896-6273(00)00109-4. [DOI] [PubMed] [Google Scholar]
- Takamori S, Holt M, Stenius K, Lemke EA, Gronborg M, Riedel D, Urlaub H, Schenck S, Brugger B, Ringler P. Molecular anatomy of a trafficking organelle. Cell. 2006;127(4):831–46. doi: 10.1016/j.cell.2006.10.030. others. [DOI] [PubMed] [Google Scholar]
- Voglmaier SM, Kam K, Yang H, Fortin DL, Hua Z, Nicoll RA, Edwards RH. Distinct endocytic pathways control the rate and extent of synaptic vesicle protein recycling. Neuron. 2006;51(1):71–84. doi: 10.1016/j.neuron.2006.05.027. [DOI] [PubMed] [Google Scholar]
- von Gersdorff H, Matthews G. Depletion and replenishment of vesicle pools at a ribbon-type synaptic terminal. J Neurosci. 1997;17(6):1919–27. doi: 10.1523/JNEUROSCI.17-06-01919.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Gersdorff H, Matthews G. Electrophysiology of synaptic vesicle cycling. Annu Rev Physiol. 1999;61:725–52. doi: 10.1146/annurev.physiol.61.1.725. [DOI] [PubMed] [Google Scholar]
- West AE, Neve RL, Buckley KM. Targeting of the synaptic vesicle protein synaptobrevin in the axon of cultured hippocampal neurons: evidence for two distinct sorting steps. J Cell Biol. 1997;139(4):917–27. doi: 10.1083/jcb.139.4.917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wienisch M, Klingauf J. Vesicular proteins exocytosed and subsequently retrieved by compensatory endocytosis are nonidentical. Nature Neuroscience. 2006;9(8):1019–1027. doi: 10.1038/nn1739. [DOI] [PubMed] [Google Scholar]
- Wu LG, Betz WJ. Kinetics of synaptic depression and vesicle recycling after tetanic stimulation of frog motor nerve terminals. Biophys J. 1998;74(6):3003–9. doi: 10.1016/S0006-3495(98)78007-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Q, Petersen CC, Nicoll RA. Effects of reduced vesicular filling on synaptic transmission in rat hippocampal neurones. J Physiol 525 Pt. 2000;1:195–206. doi: 10.1111/j.1469-7793.2000.t01-1-00195.x. [DOI] [PMC free article] [PubMed] [Google Scholar]