

Abstract
Gastric cancer is the most common malignancy of the gastrointestinal tract in East Asian populations and the second most frequent cause of cancer-related mortality in the world. While previous studies have investigated the genetic factors involved in gastric carcinogenesis, there still exist relatively few studies that have investigated the genetic traits associated with the risk of gastric precancerous conditions. In this paper we will review the biology and genetic polymorphisms involved in the genesis of gastric precancerous conditions reported to date and discuss the future prospects of this field of study. The associations of gastric precancerous conditions with polymorphisms in the cytotoxin-associated gene A-related genes (e.g. PTPN11 G/A at intron 3, rs2301756), those in the genes involved in host immunity against Helicobacter pylori (H. pylori) infection (e.g. TLR4 +3725G/C, rs11536889) or polymorphisms of the genes essential for the development/ differentiation of the gastric epithelial cells (e.g. RUNX3 T/A polymorphism at intron 3, rs760805) have been reported to date. Genetic epidemiological studies of the associations between H. pylori-induced gastric precancerous conditions and other gene polymorphisms in these pathways as well as polymorphisms of the genes involved in other pathways like oxidative DNA damage repair pathways would provide useful evidence for the individualized prevention of these H. pylori-induced gastric precancerous conditions.
Keywords: Helicobacter pylori, Gastric cancer, Single nucleotide polymorphisms, Genetic predisposition to disease, Gastric precancerous conditions
INTRODUCTION
Gastric cancer is the most common malignancy of the gastrointestinal tract in East Asian populations and the second most frequent cause of cancer-related mortality in the world[1,2]. Helicobacter pylori (H. pylori) infection has been established as a major risk factor for developing gastric cancer and its precursor lesions by numerous epidemiological studies[3,4]. More than 50% of the world population is infected with this bacterium[5]. Most case-control and cohort studies have shown that the risk of patients with H. pylori infection for developing gastric cancer is from two- to six-fold[6]. Moreover, some of the trials on H. pylori eradication revealed that cure of its infection reduces the development of gastric cancer in high risk populations[7,8]. Meanwhile, accumulated evidence indicates that there are three steps in gastric carcinogenesis: H. pylori infection, development of gastric precancerous conditions and carcinogenesis[9] (Figure 1). Severe gastric atrophy (GA) and corpus-predominant gastritis, intestinal metaplasia (IM) and dysplasia are well recognized as predominant predispositions to gastric cancer[10,11]. The extent of these gastric damages due to H. pylori infection seems to vary from one subject to another, suggesting the existence of some genetic factors that play important roles in determining the long-term outcome of H. pylori infection. While previous studies have investigated the genetic factors involved in gastric carcinogenesis[12,13] or H. pylori infection[14], the number of the reports that examined the roles of genetic factors in each step of gastric carcinogenesis was limited. Especially, few studies investigated the genetic traits associated with the risk of gastric precancerous conditions which would potentially be of significance for the prevention of gastric cancer itself. In this paper we will review the biology and genetic polymorphisms involved in the genesis of gastric precancerous conditions reported to date and discuss the future prospects in this field of study.
Figure 1.

Steps in Helicobacter pylori-related gastric cancer. H. pylori: Helicobacter pylori; NOD1: Nucleotide-binding oligomerization domain protein 1; PTPN11: Protein-tyrosine phosphatase non receptor-type 11; RUNX3: Runt-related gene 3; cagPAI: cag pathogenicity island.
EPIDEMIOLOGY OF GASTRIC ATROPHY
GA is supposed to be a result of inflammation induced by H. pylori infection[15-21]. In epidemiologic studies, serum pepsinogens (PGs) have been used as a marker of GA[22] because it is easily available with a less invasive method. Several lifestyle factors like salty food intake[23], low light-colored vegetable intake[15,21], low vitamin C[16] and high starch intake[24] have been shown to be risk factors for GA among subjects with and without H. pylori infection. A recent study with 1071 H. pylori-infected Japanese revealed that those who take too much rice, miso soup, cod roe, and cuttlefish, representative Japanese traditional food, were at higher risk of developing GA[25]. Frequent rice intake was shown to increase the risk of atrophic gastritis in another study with 291 H. pylori-infected Japanese Brazilians[26]. A double-blinded randomized controlled intervention study in Japan demonstrated that 500 mg of vitamin C supplementation for 5 years prevented the decrease in average PGI/II ratio slightly without reduction of H. pylori seropositive percentage[27].
GENES AND POLYMORPHISMS INVOLVED IN GASTRIC PRECANCEROUS CONDITIONS
Identifying candidate genes for genetic predisposition to a gastric precancerous condition is a major challenge that stems from a profound understanding of the etiology of this condition. p53 mutations are shown to be detected during the stages of GA and metaplasia[28]. The deregulation of the “hummingbird phenotype” induction in the gastric epithelial cells through cytotoxin-associated gene A (CagA)-src homology 2 domain-containing protein tyrosine phosphatase-2 (SHP-2) interaction[29] or the disruption of epithelial cell polarity through the CagA-Partitioning-defective 1 (PAR1) interaction[30] might possibly play important roles in the formation of gastric precancerous conditions. Also, like in other gastrointestinal carcinogenesis, TGF-β signaling and the subsequent inflammatory process might play essential roles in the genesis of these conditions[31]. These hypothetical biological mechanisms underlying the genesis of gastric precancerous conditions need to be investigated further in future research.
Although biological mechanisms involved in the genesis of gastric precancerous conditions remain largely unclear, they seem to involve both direct effects by the virulence factors of H. pylori and indirect effects derived from pro-inflammatory immune response by the host[28].
Cag pathogenicity island-related genes and their polymorphisms
The former effects of H. pylori virulence factors may include those induced by H. pylori induced virulence factor CagA[32-34]. CagA is a 120 to 145-kDa H. pylori protein encoded by the cagA gene[35,36] which is localized at one end of the cag pathogenicity island (cagPAI), a 40-kb DNA segment considered to be horizontally transfected to the H. pylori genome[37,38]. CagA is delivered from H. pylori bacterium into host cell cytoplasms through the type IV secretion system[39] and undergoes tyrosine phosphorylation[40]. In the injected gastric epithelial cells, CagA induces cellular spreading and elongation, called the hummingbird phenotype, which is thought to play important roles in H. pylori-induced gastric carcinogenesis. In this CagA-dependent morphologic transformation of gastric epithelial cells, a key molecule src homology 2 domain-containing protein tyrosine phosphatase-2 (SHP-2) is required[41]. Binding of tyrosine phosphorylated CagA to the SH2 domains of SHP-2 causes a conformational change in SHP-2 itself that leads to aberrantly activated SHP-2 phosphatase. SHP-2 plays a major role in intracellular signaling provoked by various growth factors, hormones or cytokines and is widely expressed in both embryonic and adult tissues[30,42]. SHP-2 is required for full activation of the Ras-MAP kinase cascade in response to growth factor-receptor interaction and plays an important role in cell morphogenesis as well as cell motility[43] which might partly explain the mechanism for the formation of hummingbird phenotype.
Meanwhile, CagA is shown to disrupt the tight junctions and causes loss of epithelial apical-basolateral polarity through the specific interaction of CagA with partitioning-defective-1 (PAR1)/ microtubule affinity-regulating kinase-2 (MARK2)[30,34] (Figure 2). PAR1b is localized to the basolateral membrane in normal polarized epithelial cells while atypical protein kinase C (aPKC) complex is localized specifically to the apical membrane. When CagA is delivered and injected into normal polarized gastric epithelial cells, CagA inhibits the kinase activity of PAR1b by binding directly to its kinase domain which subsequently leads to junctional and polarity defects followed by the disorganization of the epithelial monolayer[30]. PAR1b exists as a homodimer in the cells and two CagA proteins bind to a PAR1b dimer, also essential for stable CagA-SHP2 interaction.
Figure 2.
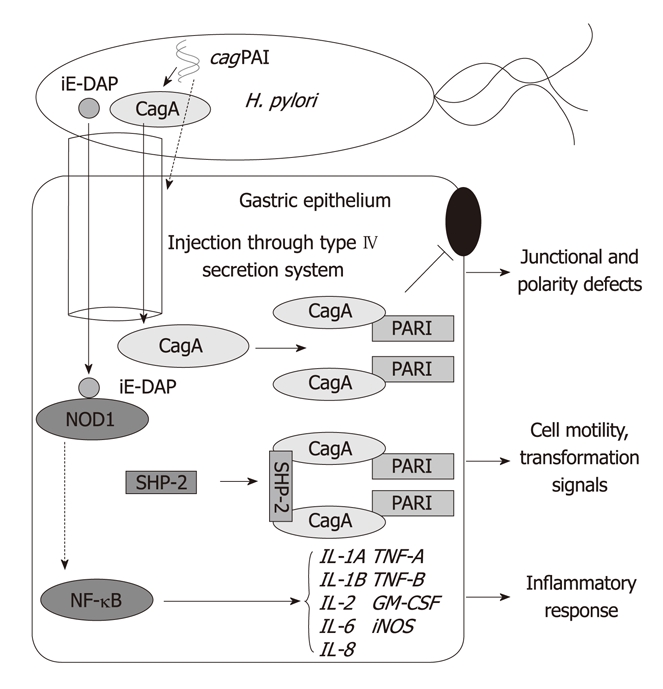
Changes in the gastric epithelial cells due to the activation of Helicobacter pylori cag pathogenicity island-related molecules. H. pylori: Helicobacter pylori; iE-DAP: γ-D-glutamyl-meso-diaminopimelic acid; iNOS: Inducible nitric oxide synthase; NF-κB, Nuclear factor κB; NOD1: Nucleotide-binding oligomerization domain protein 1; PAR1: Partitioning-defective-1; SHP-2: Src homology 2 domain-containing protein tyrosine phosphatase-2; cagPAI: cag pathogenicity island.
Recently, a cytosolic pattern recognition receptor, nucleotide-binding oligomerization domain protein 1 (NOD1), was found to respond to peptidoglycan delivered by H. pylori cagPAI[44]. NOD1 is known to sense the essential γ-D-glutamyl-meso-diaminopimelic acid (iE-DAP) dipeptide which is uniquely contained in peptidoglycan of all gram negative and certain gram-positive bacteria[45].
As the precise relationship of gastric precancerous conditions like GA and IM with these cagPAI-associated molecules is largely left unknown, further investigations are required to clarify the roles of these cagPAI-related molecules in the genesis of gastric precancerous conditions.
The studies on the associations between genotypes and GA among the infected were relatively limited (Table 1)[46-59]. In the majority of these studies, serum PGs were measured for the diagnosis of gastric mucosal atrophy where gastric mucosal atrophy was grouped into “none” (PG I >70 ng/mL or PG I/PG II > 3), “mild” (PG I ≤ 70 ng/mL and PG I/PG II ≤ 3, excluding “severe” cases) or “severe” (PG I ≤ 30 ng/mL and PG I/PG II ≤ 2)[46,47,50,52,54-56,58], while the diagnosis of GA was done based on the endoscopic findings in the rest of the studies[48,49,51,53,58,60].
Table 1.
Polymorphisms reported on the associations with gastric atrophy among Helicobacter pylori seropositives as well as odds ratio and/or gastric atrophy percent.
| Polymorphism | Function | Rs No. | Subjects [Ref.] | OR and/or GA% |
| IL-1B C-31T | 5'UTR | rs1143627 | 253 Japanese[104] | CC (54%),CT (52%),TT (56%) |
| IL-1B C-31T | 5'UTR | rs1143627 | 455 Jpn.Brazil.[59] | CC, CT: 0.61,TT: 0.58 CC (36%), CT (31%), TT (21%) |
| IL-1B C-31T | 5'UTR | rs1143627 | 1328Venezuelan[94] | TT, CT: 1.01, CC: 0.91 |
| IL-2 T-330G | 5'UTR | rs2069762 | 244 Japanese[52] | GG ,TG: 1.64, TT: 2.78 aGG (38%), TG (50%), TT (62%) |
| IL-4 C-33T | 5'UTR | rs2070874 | 249 Japanese[52] | CC, CT: 2.47, TT: 1.80 CC (38%), CT (60%), TT (53%) |
| IL-4 T-590C | 5'UTR | rs2243250 | 1301 Venezuelan[53] | TT, CT: 0.82, CC: 0.81 |
| IL-4R C-3223T | 5'UTR | rs2057768 | 1301 Venezuelan[53] | CC, CT: 0.97, TT: 1.01 |
| IL-4R A398G | Non-synonymous (Ile50Val) | rs1805010 | 1301 Venezuelan[53] | AA, AG: 1.14, GG: 1.52a |
| IL-6 G-174C | 5'UTR | rs1800795 | 1315 Venezuelan[94] | GG, CG: 0.98, CC: 0.57 |
| IL-8 T-251A | 5'UTR | rs4073 | 1347 Venezuelan[94] | TT, AT: 0.98, AA: 1.07 |
| IL-10 G-1082A | 5'UTR | rs1800896 | 1301 Venezuelan[53] | GG, AG: 1.05, AA: 1.14 |
| IL-13 C-1111T | 5'UTR | rs1800925 | 248 Japanese[52] | CC, CT+TT: 0.41 aCC (59%), CT+TT (45%) |
| TNF-A T-1031C | 5'UTR | rs4647198 | 455 Jpn.Brazil.[105] | CC (29%),TC (33%), TT (34%) |
| TNF-A C-857T | 5'UTR | rs1799724 | 456 Jpn.Brazil.[105] | CC (32%), CT (36%), TT (39%) |
| TNF-A-1031&-857 | 5'UTR | rs1799724 | 455 Jpn.Brazil.[105] | CC&CC (29%), TT&CC (33%), TC&CT (43%), TT&TT (39%) |
| TNF-A G-308A | 5'UTR | rs1800629 | 1327 Venezuelan[106] | GG, AG+AA: 1.27 |
| MCP1 G-2518A | 5'UTR | rs1024611 | 1311 Venezuelan[94] | AA, AG: 1.02, GG: 1.18 |
| PTPN11 G/A at intron3 | Intron | rs2301756 | 248 Japanese[46] | GG, GA: 0.70, AA: 0.09 aGG (59%),GA (49%),AA (11%) |
| PTPN11 G/A at intron3 | Intron | rs2301756 | 979 Japanese[47]b | GG, GA+AA: 0.62c aGG (22%),GA+AA (15%)c |
| NOD1 G796A | Non-synonymous (Glu266Lys) | rs2075820 | 150 Turks[48]e | GG, GA: 13.35a, AA: 34.2a |
| TLR4 A+896G | Non-synonymous (Asp299Gly) | rs4986790 | 103 Caucasians[49] | AA, AG: 11.0* AA (36%), AG (87%)d |
| TLR4 A+896G | Non-synonymous (Asp299Gly) | rs4986790 | 717 Venezuelan[94] | GlyGly/AspGly, AspAsp: 1.53 |
| TLR4 G+3725C | 3'UTR | rs11536889 | 980 Japanese[50]b | GG, GC+CC: 1.33 GG( 18%), GC+CC (22%)c |
| CD14 C-159T(C-260T) | 5'UTR | rs2569190 | 717 Venezuelan[94] | CC, CT+TT: 1.06 |
| PRKCH rs3783799 G/A | Intron | rs3783799 | 1638 Japanese[54] | GG, GA: 0.99, AA: 2.37a |
| iNOS C150T | Non-synonymous (Ser608Leu) | rs2297518 | 250 Japanese[55] | CC, CT+TT: 0.75 |
| RUNX3 T/A at intron3 | Intron | rs760805 | 938 Japanese[56]b | TT, TA: 1.51a, AA:1.59a TT(46%), TA (56%), AA (56%) |
| HSP70-2 A/B (A1267G) | Synonymous (Gln351Gln) | rs1061581 | 137 Japanese[57] | BB (45%), AA+AB (67%)c |
| FAS G-1377A | 5'UTR | rs2234767 | 109 Taiwanese[85] | No association |
| FAS A-670G | 5'UTR | rs1800682 | 109 Taiwanese[85] | No association |
| FASL T-844C | 5'UTR | rs763110 | 109 Taiwanese[85] | TT, TC+CC: 9.4a |
| PGC Ins/Del | Ins/Del | (unknown) | 86 Chinese[68] | DD (90%), others (50%)a |
| NQO1 C609T | Non-synonymous (Pro187Ser) | rs1800682 | 396 Japanese[95] | TT, CT: 1.25, CC: 1.23 |
| GSTM1 | Ins/Del | (unknown) | 396 Japanese[95] | Null, Present: 1.35 |
| GSTT1 | Ins/Del | rs71748309 | 396 Japanese[95] | Null, Present: 0.87 |
| ACE Ins/Del | Ins/Del | rs1799752 | 271 Japanese[107] | II, ID: 1.12, DD: 0.99 |
| RANTES C-471T | 5'UTR | rs2107538 | 344 Germans[108] | No association |
| SDHC JST173800 C/G | 3'UTR | rs3813632 | 249 Japanese[109] | CC, GC: 1.26, GG: 0.51 CC (53%), GC (59%), GG (38%) |
| IFNGR1 G-611A | 5'UTR | rs1327474 | 805 Portuguese[110] | GG, GA: 1.2, AA: 1.2 |
| IFNGR1 C-56T | 5'UTR | rs2234711 | 814 Portuguese[110] | CC, CT: 1.4, TT: 1.3 |
aStatistically significant (P < 0.05); bNumber of H. pylori infected subjects (H. pylori seropositive subjects or subjects with gastric atrophy); cOR for/percentages of subjects with severe gastric atrophy; dThere was no subject with GG genotype; eCagA-positive subjects, H. pylori: Helicobacter pylori; GA: Gastric atrophy; OR: Odds ratio.
Protein tyrosine phosphatase, non-receptor type, 11 G/A at intron 3 (rs2301756): The protein tyrosine phosphatase, non-receptor type, 11 (PTPN11) G/A polymorphism at intron 3 (rs2301756) is a G-to-A single nucleotide substitution at 223 bp upstream of exon 4 in the PTPN11 gene encoding SHP-2 at chromosome 12q24.1. The biological function of this polymorphism has not yet been reported. The first dataset showed that one (11.1%) out of 9 infected individuals with the AA genotype had GA while 134 (56.1%) among 239 infected with the G allele had atrophy[60]. Our recent report of 1636 non-cancer Japanese subjects demonstrated that the risk of severe GA was significantly reduced for those with at least one A allele of this PTPN11 G/A polymorphism at intron 3 (OR = 0.62, 95%CI: 0.42-0.90), confirming the association of this PTPN11 gene polymorphism with the risk of gastric precancerous conditions in H. pylori-infected subjects[46]. If the polymorphism is functional or linked to a functional one, the association can be biologically explained by the difference in the strength of signal transduction through the CagA-SHP2 complex. According to the NCBI dbSNP, the frequencies of the G allele of rs2301756, high risk allele for GA, is 0.802 among 1484 Japanese and 0.917 among 48 Chinese while it was 0.348 among 46 African American and 0.064 among 46 Caucasians, indicating that Japanese and Chinese become high risk ethnic groups through CagA-positive H. pylori infection if the hypothesis that the G allele confers stronger signals via the CagA-SHP2 interaction is true.
Nucleotide-binding oligomerization domain protein 1 G796A (E266K): A recent report revealed that the carriage of the NOD1 G796A mutation increases the susceptibility for GA strikingly: OR = 34.2 in NOD1 796AA and OR = 13.35 in NOD1 796GA compared to subjects with NOD1 796GG[48].
Immune related genes and their polymorphisms
For the latter effects of pro-inflammatory immune response by the hosts, TLR4 recognizes lipopolysaccharide (LPS) of gram-negative bacteria and is proved to play important roles in H. pylori infection through the interaction of macrophage/monocyte TLR4 with H. pylori LPS[61,62]. The initial recognition of LPS and subsequent signaling by TLR4 is supported by several accessory proteins: LPS first binds to lipopolysaccharide-binding protein (LBP) which works as an opsonin for CD14 which then acts as a catalyst for the binding of LPS to MD-2[63]. Then the signal induced by LPS/MD-2/TLR4 complex is transmitted through myeloid differentiation factor 88 (MyD88), interleukin (IL)-1 receptor associated kinase (IRAK), tumor necrosis factor (TNF) receptor-associated factor 6 (TRAF6) and inhibitory κB kinase (IKK) to nuclear factor (NF)-κB, leading to the production of pro-inflammatory cytokines such as IL-1A, IL-1B, IL-6 or TNF-A[64] (Figure 3). Meanwhile, the human immune system is also balanced by the anti-inflammatory cytokines like IL-10, IL-4 or IL-13 which are controlled by regulatory T cells[65]. In these inflammatory processes, increased expression of inducible nitric oxide synthase (iNOS) is shown to play important roles in the production of oxygen radicals whereas overexpression of cyclooxygenase-2 (COX-2) is demonstrated to contribute to the proliferation of the gastric epithelium through the up-regulation of cell-cycles as well as to the propagation of gastric inflammation via the prostaglandin pathways[66]. The induction of iNOS is also supposed to be modulated by the activity of protein kinase C-eta (PRKCH) via the phosphorylation of NF-κB or activator protein-1 (AP-1)[67,68].
Figure 3.
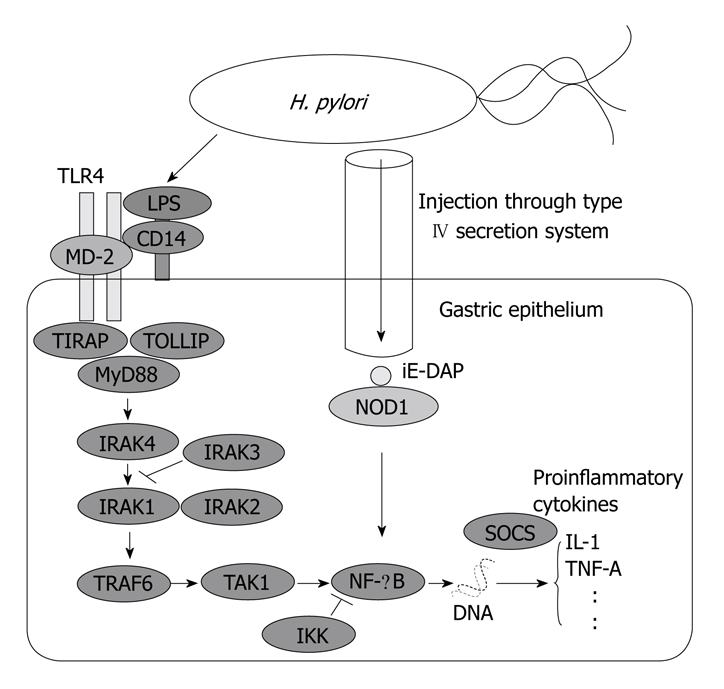
Signal pathways from Helicobacter pylori to cytokine gene expression through innate immunity. H. pylori: Helicobacter pylori; IKK: Inhibitory κB kinase; iE-DAP: γ-D-glutamyl-meso-diaminopimelic acid; IRAK: Interleukin 1 receptor-associated kinase; MyD88: Myeloid differentiation factor 88; NF-κB, Nuclear factor κB; NOD1: Nucleotide-binding oligomerization domain protein 1; SOCS: Suppressor of cytokine signaling; TOLLIP: Toll-interacting protein; TIRAP: TIR domain-containing adaptor protein.
Oxidative DNA damage is also supposed to play important roles in the pathogenesis of H. pylori-induced gastric mucosal damage where 8-OHdG is a potential sensitive marker of DNA oxidation[69]. The damaged bases in DNA are mainly repaired by the base excision repair (BER) system; the accumulation of 8-Hydroxy-2’-deoxyguanosine (8-OHdG) or 2-hydroxyadenine (2-OH-A) in DNA is prevented by the co-operation of mutT human homolog-1 (MTH1), 8-hydroxyguanine DNA glycosylase (OGG1) and mutY human homolog (MUTYH)[70]. The number of studies that investigated the contribution of these molecules involved in the inflammatory response, such as innate immune response, oxygen radical production, oxidative DNA damage repair processes, together with cell-cycle regulation and/or cell proliferation in the genesis of H. pylori-induced gastric precancerous conditions, is also limited, requiring further biological investigations in the near future.
TLR4 polymorphisms (+896 A/G, rs4986790; +3725G/C, rs11536889): One study in Caucasians showed that the TLR4 +896 A/G polymorphism was associated with the risk of GA where the TLR4 +896 G carriers had an 11-fold increased risk of GA with hypochlorhydria[49]. A subsequent Japanese study also clarified the possible association between another genetic variation in TLR4 gene, the TLR4 +3725G/C polymorphism (rs11536889) and the risk of severe GA in Japanese[50], suggesting the significance of genetic variations in host innate immunity due to TLR4 polymorphisms also in East Asian populations.
CD14 C-159T polymorphism: There is one single nucleotide polymorphism in the promoter region of the CD14 gene, CD14 C-159T polymorphism, which is critical for CD14 expression[71]. A recent study by one Japanese group demonstrated that CD14 promoter -159TT and T carrier were associated with lower risk of GA in H. pylori-infected subjects who were 61 years or older[51].
IL-2 T-330G and IL-13 C-1111T polymorphisms: IL-2 T-330G polymorphism was demonstrated to be a functional polymorphism[72] with higher IL-2 production in GG genotype than in TT genotype[73]. Those with TT genotype were shown to be at a higher risk of GA[52], less frequent in Asians (38% out of 29 individuals) than in Caucasians (51% out of 199 individuals)[74].
IL-13 gene in chromosome 5q31 has several polymorphisms; at least 3 polymorphisms at the promoter region, 2 polymorphisms at intron 1, Arg130Gln and 4 polymorphisms at 3’ UTR of exon 4 have been reported[75]. The -1111TT genotype was shown to harbor increased binding ability of nuclear proteins and was also reported to be associated with asthma[75,76]. As for the risk of GA, -1111TT was found to be a low risk genotype[52]. The biological mechanism involved was not yet clarified.
IL-4R C-332T (rs1805010) polymorphism: One study of Venezuelan subjects revealed that those with homozygotes with the low activity allele (GG) of the A398G polymorphism in the IL-4R gene (rs1805010) had a modestly increased risk of GA (OR = 1.52, 95%CI: 1.05-2.21)[53], suggesting the role of genetic variability in the anti-inflammatory mediators in the genesis of H. pylori-induced gastric precancerous conditions.
Inducible nitric oxide synthase C150T (rs2297518) and PRKCH rs3783799 G/A polymorphisms: PRKCH is shown to be involved in oxidative stress by activating iNOS and nitric oxide production[67]. The associations of the polymorphisms in these two genes [iNOS C150T (rs2297518) and PRKCH rs3783799 G/A polymorphisms] with the risk of GA were investigated in the Japanese population which revealed that those with PRKCH rs3783799 AA genotype were at significantly higher risk of severe GA (OR = 2.37, 95%CI: 1.11-5.05)[54] while there were no significant association between the iNOS C150T polymorphism and risk of GA[55].
Other miscellaneous genes and their polymorphisms
Recently, it was reported that the loss expression of sonic hedgehog (Shh), a regulatory gene essential for developmental patterning, and aberrant expressions of caudal-type homeobox transcription factor 2 (CDX2), a master regulatory gene of intestinal development and differentiation, in H. pylori-induced atrophic gastritis are the early events correlated with the occurrence of IM which can be reversible by the eradication of H. pylori. In accordance with these findings, CDX2 expression has been demonstrated to be associated with intestinal phenotypes in gastric cancers[77].
Another important tumor suppressor gene in intestinal-type gastric cancer is runt-related gene 3 (RUNX3) encoding a subunit of polyomavirus enhancer binding protein 2[78], since expression of RUNX3 is greatly reduced in IMs in human stomachs[79] and RUNX3-/- mouse gastric epithelial cells have a potential to differentiate into CDX-2 positive intestinal type cells[80]. Li et al[78] and Levanon et al[81] reported that the gastric mucosa of RUNX3 null mice showed hyperplasia, indicating that loss of RUNX3 leads to gastric carcinogenesis in humans. Consistent with this, an analysis of RUNX3 in human stomach cancer cell lines and primary human tumors revealed hemizygosity in 40% of the tumors examined and silencing by promoter hypermethylation in 60% of the tumors; this figure increased up to 90% in the advanced stage tumors. It is shown that the RUNX3-/- mouse gastric mucosa exhibits hyperplasias due to the stimulated proliferation and suppressed apoptosis in the cells, suggesting that RUNX3 is an attractive candidate as a tumor suppressor of gastric cancer. The CpG island of RUNX3 P2 promoter is hypermethylated in human and mouse gastric cancer cell lines and in primary human tumors[78,82], also suggesting the tumor suppressor function of RUNX3 in the etiology of stomach cancer.
Heat-shock protein (HSP) 70 plays essential roles in cellular response to a variety of environmental stresses by acting as molecular chaperons in the folding of newly synthesized proteins in cells and assist in the folding of damaged proteins[83]. HSP expression in the gastric mucosa is shown to be attenuated by H. pylori infection and aspirin intake and one HSP inducer geranylgeranylacetone (GGA) reportedly protects gastric mucosa from iNOS induced by H. pylori infection[84], suggesting that HSP has important roles in protecting gastric mucosa against H. pylori or aspirin induced injuries. Gastric carcinogenesis can also be regarded as a multistep process that initiates with the disregulation of normal controls of apoptosis and cell proliferation in which FAS receptor-ligand system is shown to be a key regulator of apoptosis[85].
Pepsinogen C (PGC), alternatively called pepsinogen II or gastricsin, an inactive precursor of pepsin C, is an aspartic protease specifically produced by the gastric chief cells, cardiac cells, pylori cells and Brunner’s glands from late infant stages to the adulthood period. PGC is considered to be a differentiation marker of gastric epithelium whose changes in expression may reflect the severity of gastric mucosal damage[60].
Runt-related gene 3 T/A polymorphism at intron 3 (rs760805): Among H. pylori seropositive subjects, we found a significant association between RUNX3 rs760805 polymorphism and the risk of GA with the age- and sex-adjusted OR of 1.51 (95%CI: 1.11-2.05, P = 0.008) in TA, 1.59 (95%CI: 1.08-2.33, P = 0.019) in AA and 1.53 (95%CI: 1.14-2.05, P = 0.004) in TA+AA compared with TT genotype[56]. This finding was in accordance with the recent biological report that RUNX3 expression correlated with chief cell differentiation in human gastric cancers[86].
Heat-shock protein 70-2 A/B (A1267G) polymorphism: It is shown that the AA genotype of HSP 70-2 A/B polymorphism (PsiI polymorphism, corresponding to A1267G polymorphism) had the highest level of mRNA expression compared with the other genotypes (AB or BB). Recently one Japanese group reported that the BB genotype of HSP 70-2 gene is significantly associated with the reduced risk of severe GA in H. pylori infected older subjects[57], indicating the importance of this HSP polymorphism in the genesis of H. pylori-induced gastric precancerous conditions. In a recent study, polymorphisms in HSP 70 genes along with TNF polymorphisms showed a significant severity-dose-response as risk markers from precancerous lesions to gastric cancer in Mexican population, presumably because of their association with the intense and sustained inflammatory response[87].
FASL T-844C polymorphism: Lately, one study group in Taiwan investigated the relationship between precancerous gastric lesions and polymorphisms in the promoter regions of the death pathway genes FAS and FASL (FAS G-1377A, FAS A-670G and FASL T-844C) in 109 H. pylori-infected Taiwanese individuals and found that FASL -844 C allele significantly increased the risk of atrophy in the gastric corpus, with an adjusted OR of 5.0 (95%CI: 1.5-6.8)[85].
Pepsinogen C ins/del polymorphism: A recent study among Chinese demonstrated that subjects with PGC del/del genotype were at significantly higher risk of atrophic gastritis (OR = 3.11; 95%CI: 1.44-6.71) and H. pylori-seropositive subjects with PGC del/del genotype had significantly elevated risk of atrophic gastritis (OR = 11.16; 95%CI: 1.37-90.84) with the interaction of 6.48[58], suggesting the positive link between PGC gene polymorphism and H. pylori-induced GA.
Genes and polymorphisms for advanced precancerous conditions
Advanced precancerous conditions like IM or dysplasia develops in some part of H. pylori infected subjects. One Chinese study demonstrated no significant differences in genotype frequencies of CYP2E1, GSTM1, GSTP1, GSTT1, ALDH2 and ODC between those with mild chronic atrophic gastritis including 29.7% H. pylori negative subjects and those with deep IM or dysplasia with 20.2% of H. pylori negative subjects, but found significant interaction between CYP2E1 DraI genotypes and smoking[88]. Another study in Germany revealed that carriers of both of IL-1B -511T and IL-1RN 2rpt alleles relative to subjects lacking IL-1B -511T or/and IL-1RN 2rpt alleles had significantly increased risk for the development of atrophic gastritis, IM and severe inflammation[89].
Recently, one Japanese study group investigated extensively into the genetic polymorphisms associated with the risk of gastric advanced precancerous conditions. They reported that the risk of IM among H. pylori seropositive individuals was significantly associated with the polymorphisms of COMT Val158Met[90], cyclin D1 (CCND1) G870A[91], p22PHOX C242T[92], VEGF G1612A at 3’-UTR[93] and HSP70-2 A1267G[57] while it was not associated with VEGF C936T at 3’-UTR[93]. A recent report by a Turkish group also revealed the significant association of risk of antral IM with G796A (E266K) polymorphism in the NOD1 gene encoding a cytosolic receptor to peptidoglycan delivered by cagPAI; the risk was strikingly increased in those with AA genotype (OR = 39.76) and also significantly increased in those with GA genotype (OR = 2.71)[48]. One group in the US investigated the associations of risk for H. pylori-induced gastric precancerous conditions with the polymorphisms in the genes involved in host-bacterial interaction, (IL-1B C-31T, IL-6 G-174C, IL-8 T-251A, MCP-1 G-2518A and TNF G-308A), bacterial LPS signaling (CD14 C-260T, TLR4 Asp299Gly, NOD2 del 3020 ins C and NOD2 Gly908Arg) and anti-inflammatory cytokine signaling (IL-10 G-1082A, IL-4 T-590C, IL-4R C-3223T and IL-4R A398G) among Venezuelan subjects[53,94,95], in which they found the associations between risk of dysplasia and IL-8 T-251A polymorphism (OR = 2.00 for AA, 1.33 for AT; P for trend = 0.02) or between that of IM and CD14 C-260T (OR = 1.45 for CT, 1.45 for TT; P for trend = 0.025) or IL-10 G-1082A (OR = 1.34 for AG, 1.50 for AA; P for trend = 0.055) polymorphisms. Another group in Taiwan investigated the associations of the risk of precancerous gastric lesions and polymorphisms in the promoter regions of the death pathway genes FAS and FASL (FAS G-1377A, FAS A-670G and FASL T-844C) in 109 H. pylori-infected Taiwanese individuals where they found significantly increased risk of IM in the antrum for those with FAS -1377 A allele with the adjusted OR of 0.3 (95%CI: 0.1-0.9)[85].
FUTURE PROSPECTS
Intriguing genes for future investigation
Although the G allele of PTPN11 may be a part of the genetic traits to develop GA via signal transduction from CagA, there seems to be other genetic traits involved in this process. CagA binds several molecules, Grb2 which transduces the signal to Ras-MAP kinase pathway causing cell proliferation, c-Met hepatocyte growth factor (HGF) receptor which have a role of cell proliferation and motility, zona occludens-1 (ZO-1), a tight-junction protein, and PAR1/MARK kinase which has an essential role in epithelial cell polarity[30,96-99]. Although no studies have been conducted, functional polymorphisms of these molecules might also be possible candidates for the genetic traits of GA.
There seem to be other intriguing candidate molecules as possible susceptible factors. A recent study that examined the associations of 75 haplotype-tagging SNPs in genes in the TLR signaling pathway with pertussis toxin (PT)-IgG titers demonstrated that antibody response to pertussis vaccination was significantly associated with the polymorphisms in CD14, TLR4, toll-interacting protein (TOLLIP), TIR domain-containing adaptor protein (TIRAP), interleukin 1 receptor-associated kinase 3 (IRAK3), interleukin 1 receptor-associated kinase 4 (IRAK4), TIR domain-containing adaptor molecule 1 (TICAM1), and tumor necrosis factor ligand superfamily, member 4 (TNFSF4)[100]. Considering the crucial role of TLR4 pathways in the genesis of H. pylori-induced gastric cancer, it would be of interest to investigate the involvement of these polymorphisms in the three steps of H. pylori-induced gastric carcinogenesis. Meanwhile, although some previous studies have shown the essential roles of the polymorphisms of DNA BER genes (OGG1, MUTYH and MTH1) in H. pylori-related gastric carcinogenesis[101,102], few studies have investigated their roles in the genesis of H. pylori-induced gastric precancerous conditions. There are several other genes reported to be underlying the genesis of H. pylori-induced gastric precancerous conditions. Ornithine decarboxylase (ODC), the first and rate-limiting enzyme, is shown to be up-regulated by H. pylori with strong expression in atrophic and IM areas. In a recent Japanese study, CDX2 expression was observed in patients with chronic gastritis closely associated with IM while some other genes like mucin 1 (MUC1), p27 or p53 are also shown to be implicated in the genesis of IM[103]. The associations of these gene polymorphisms with the risk of H. pylori-induced gastric premalignant lesions have also not yet been examined. Further investigations are expected to investigate the significance of the polymorphisms of these genes in H. pylori-induced gastric precancerous conditions. Furthermore, haplotype analyses as a gene-specific approach to find novel functional polymorphisms in the genes involved or genome-wide association studies (GWAS) as a comprehensive approach to detect novel candidate gene polymorphisms strongly associated with disease risk should also be conducted to provide useful evidence for the individualized prevention of H. pylori-induced gastric precancerous conditions. By identifying the full genetic risk profile for H. pylori-induced gastric precancerous conditions, we will be able to target the population at risk and subsequently direct eradication therapy and closer follow-up to the affected individuals.
Recommendation for stepwise risk evaluation
Although a large number of studies have been reported concerning genetic traits associated with gastric cancer risk, few studies investigated which step of H. pylori-related gastric carcinogenesis (H. pylori infection, GA and gastric cancer) the genetic traits examined has effects on[9]. These stepwise evaluations of H. pylori-related gastric carcinogenesis provide us with more precise and detailed information about the genes involved in each step of H. pylori related gastric carcinogenesis which would help us establish the effective way of the individualized prevention against H. pylori-induced gastric cancer in the near future. We have already conducted this stepwise evaluation in previous reports[46,47,50,59] and expect that forthcoming studies by other groups would also be conducted in this way to improve the quality of the studies so that we can put the obtained results into practice for effective gastric cancer prevention.
It would also be relevant to claim that H. pylori-induced gastric precancerous conditions can be histologically subdivided into two lesions, GA and more advanced precancerous conditions of IM; thus the stepwise evaluation should be conducted with four steps, i.e. dividing the third step of gastric carcinogenesis from GA into more precise categories of IM/dysplasia from GA and gastric carcinogenesis from IM/dysplasia as implicated in Figure 1. In addition, the investigation of the interaction between host genetic factors and dietary factors like salty food intake, low light-colored vegetable intake, low vitamin C and starch intake on the risk of H. pylori-induced gastric precancerous condition might provide clues for effective ways of individualized gastric cancer prevention. We think this is our future assignment and more profound collaborations between epidemiologists, pathologists, gastroenterologists and nutritionists would be required to accomplish these idealistic goals.
Other remaining questions
In addition to the points raised above, the roles of the candidate genes in the genesis of gastric precancerous conditions in each histological subtype of gastric cancer (diffuse type or intestinal type) need to be investigated. Also, as the underlying molecular biological mechanisms are largely unclear as mentioned earlier, molecular epidemiological studies should keep up with the advance of the biological research in the field of gastric precancerous conditions in the future.
CONCLUSION
While recent epidemiological studies revealed the important roles of polymorphisms in the cagPAI-related genes and genes involved in immune response or development/differentiation of gastric epithelial cells in the genesis of H. pylori-induced gastric precancerous conditions as discussed in this editorial review (Figure 1), the fields of genetic epidemiological study regarding H. pylori-induced gastric precancerous conditions are still left relatively uninvestigated in spite of its substantial significance for the prevention of gastric cancer. Association studies between the risk of H. pylori-induced gastric precancerous conditions and polymorphisms of other genes in cagPAI-related pathways, innate immunity or oxidative DNA damage repair pathways would potentially provide useful evidence for the individualized prevention of these H. pylori-induced gastric precancerous conditions.
Further investigation of the association of these polymorphisms with risk of H. pylori-induced gastric precancerous conditions together with the elucidation of the biological roles of these molecules would be required for the confirmation of recent evidence and realization of practical individualized prevention of H. pylori-induced gastric cancer in the near future.
Footnotes
Supported by A Grant-in-Aid for Scientific Research from the Japanese Ministry of Education, Culture, Sports, Science and Technology.
Peer reviewers: Ying-Yan Yu, MD, PhD, Professor, School of Medicine, Shanghai Jiaotong University, Shanghai 200025, China; Rama Devi Mittal, PhD, Associate Professor, Department of Urology, Sanjay Gandhi Post Graduate, Institute of Medical Sciences, Rae Bareli Road, Lucknow 226014, India
S- Editor Wang JL L- Editor Roemmele A E- Editor Yang C
References
- 1.Parkin DM, Whelan SL, Ferlay J, Raymond L, Young J. Cancer incidence in five continents. Vol VII. Lyon: IARC Press; 1997. [Google Scholar]
- 2.Parkin DM, Pisani P, Ferlay J. Estimates of the worldwide incidence of 25 major cancers in 1990. Int J Cancer. 1999;80:827–841. doi: 10.1002/(sici)1097-0215(19990315)80:6<827::aid-ijc6>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- 3.Hamilton SR, Aaltonen LA. Pathology and genetics of tumours of the digestive system. WHO classification of tumours. Lyon, France: IARC Press; 2000. [Google Scholar]
- 4.Tsuji S, Tsujii M, Murata H, Nishida T, Komori M, Yasumaru M, Ishii S, Sasayama Y, Kawano S, Hayashi N. Helicobacter pylori eradication to prevent gastric cancer: underlying molecular and cellular mechanisms. World J Gastroenterol. 2006;12:1671–1680. doi: 10.3748/wjg.v12.i11.1671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Danesh J. Helicobacter pylori infection and gastric cancer: systematic review of the epidemiological studies. Aliment Pharmacol Ther. 1999;13:851–856. doi: 10.1046/j.1365-2036.1999.00546.x. [DOI] [PubMed] [Google Scholar]
- 6.Eslick GD, Lim LL, Byles JE, Xia HH, Talley NJ. Association of Helicobacter pylori infection with gastric carcinoma: a meta-analysis. Am J Gastroenterol. 1999;94:2373–2379. doi: 10.1111/j.1572-0241.1999.01360.x. [DOI] [PubMed] [Google Scholar]
- 7.Fukase K, Kato M, Kikuchi S, Inoue K, Uemura N, Okamoto S, Terao S, Amagai K, Hayashi S, Asaka M. Effect of eradication of Helicobacter pylori on incidence of metachronous gastric carcinoma after endoscopic resection of early gastric cancer: an open-label, randomised controlled trial. Lancet. 2008;372:392–397. doi: 10.1016/S0140-6736(08)61159-9. [DOI] [PubMed] [Google Scholar]
- 8.Wong BC, Lam SK, Wong WM, Chen JS, Zheng TT, Feng RE, Lai KC, Hu WH, Yuen ST, Leung SY, et al. Helicobacter pylori eradication to prevent gastric cancer in a high-risk region of China: a randomized controlled trial. JAMA. 2004;291:187–194. doi: 10.1001/jama.291.2.187. [DOI] [PubMed] [Google Scholar]
- 9.Hamajima N, Naito M, Kondo T, Goto Y. Genetic factors involved in the development of Helicobacter pylori-related gastric cancer. Cancer Sci. 2006;97:1129–1138. doi: 10.1111/j.1349-7006.2006.00290.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Correa P. A human model of gastric carcinogenesis. Cancer Res. 1988;48:3554–3560. [PubMed] [Google Scholar]
- 11.Uemura N, Okamoto S, Yamamoto S, Matsumura N, Yamaguchi S, Yamakido M, Taniyama K, Sasaki N, Schlemper RJ. Helicobacter pylori infection and the development of gastric cancer. N Engl J Med. 2001;345:784–789. doi: 10.1056/NEJMoa001999. [DOI] [PubMed] [Google Scholar]
- 12.El-Omar EM, Rabkin CS, Gammon MD, Vaughan TL, Risch HA, Schoenberg JB, Stanford JL, Mayne ST, Goedert J, Blot WJ, et al. Increased risk of noncardia gastric cancer associated with proinflammatory cytokine gene polymorphisms. Gastroenterology. 2003;124:1193–1201. doi: 10.1016/s0016-5085(03)00157-4. [DOI] [PubMed] [Google Scholar]
- 13.Figueiredo C, Machado JC, Pharoah P, Seruca R, Sousa S, Carvalho R, Capelinha AF, Quint W, Caldas C, van Doorn LJ, et al. Helicobacter pylori and interleukin 1 genotyping: an opportunity to identify high-risk individuals for gastric carcinoma. J Natl Cancer Inst. 2002;94:1680–1687. doi: 10.1093/jnci/94.22.1680. [DOI] [PubMed] [Google Scholar]
- 14.Hamajima N, Hishida A. Genetic traits for the persistence of Helicobacter pylori infection. Per Med. 2010;7:249–262. doi: 10.2217/pme.10.14. [DOI] [PubMed] [Google Scholar]
- 15.Tsugane S, Kabuto M, Imai H, Gey F, Tei Y, Hanaoka T, Sugano K, Watanabe S. Helicobacter pylori, dietary factors, and atrophic gastritis in five Japanese populations with different gastric cancer mortality. Cancer Causes Control. 1993;4:297–305. doi: 10.1007/BF00051331. [DOI] [PubMed] [Google Scholar]
- 16.Fontham ET, Ruiz B, Perez A, Hunter F, Correa P. Determinants of Helicobacter pylori infection and chronic gastritis. Am J Gastroenterol. 1995;90:1094–1101. [PubMed] [Google Scholar]
- 17.Kuipers EJ, Uyterlinde AM, Peña AS, Roosendaal R, Pals G, Nelis GF, Festen HP, Meuwissen SG. Long-term sequelae of Helicobacter pylori gastritis. Lancet. 1995;345:1525–1528. doi: 10.1016/s0140-6736(95)91084-0. [DOI] [PubMed] [Google Scholar]
- 18.Ozasa K, Kurata JH, Higashi A, Hayashi K, Inokuchi H, Miki K, Tada M, Kawai K, Watanabe Y. Helicobacter pylori infection and atrophic gastritis: a nested case-control study in a rural town in Japan. Dig Dis Sci. 1999;44:253–256. doi: 10.1023/a:1026633913154. [DOI] [PubMed] [Google Scholar]
- 19.Kuwahara Y, Kono S, Eguchi H, Hamada H, Shinchi K, Imanishi K. Relationship between serologically diagnosed chronic atrophic gastritis, Helicobacter pylori, and environmental factors in Japanese men. Scand J Gastroenterol. 2000;35:476–481. doi: 10.1080/003655200750023723. [DOI] [PubMed] [Google Scholar]
- 20.Namekata T, Miki K, Kimmey M, Fritsche T, Hughes D, Moore D, Suzuki K. Chronic atrophic gastritis and Helicobacter pylori infection among Japanese Americans in Seattle. Am J Epidemiol. 2000;151:820–830. doi: 10.1093/oxfordjournals.aje.a010282. [DOI] [PubMed] [Google Scholar]
- 21.Shibata K, Moriyama M, Fukushima T, Une H, Miyazaki M, Yamaguchi N. Relation of Helicobacter pylori infection and lifestyle to the risk of chronic atrophic gastritis: a cross-sectional study in Japan. J Epidemiol. 2002;12:105–111. doi: 10.2188/jea.12.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Samloff IM, Varis K, Ihamaki T, Siurala M, Rotter JI. Relationships among serum pepsinogen I, serum pepsinogen II, and gastric mucosal histology. A study in relatives of patients with pernicious anemia. Gastroenterology. 1982;83:204–209. [PubMed] [Google Scholar]
- 23.Chen VW, Abu-Elyazeed RR, Zavala DE, Ktsanes VK, Haenszel W, Cuello C, Montes G, Correa P. Risk factors of gastric precancerous lesions in a high-risk Colombian population. I. Salt. Nutr Cancer. 1990;13:59–65. doi: 10.1080/01635589009514045. [DOI] [PubMed] [Google Scholar]
- 24.Kato I, Vivas J, Plummer M, Lopez G, Peraza S, Castro D, Sanchez V, Cano E, Andrade O, Garcia R, et al. Environmental factors in Helicobacter pylori-related gastric precancerous lesions in Venezuela. Cancer Epidemiol Biomarkers Prev. 2004;13:468–476. [PubMed] [Google Scholar]
- 25.Montani A, Sasazuki S, Inoue M, Higuchi K, Arakawa T, Tsugane S. Food/nutrient intake and risk of atrophic gastritis among the Helicobacter pylori-infected population of northeastern Japan. Cancer Sci. 2003;94:372–377. doi: 10.1111/j.1349-7006.2003.tb01449.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ito LS, Oba-Shinjo SM, Marie SK, Uno M, Shinjo SK, Hamajima N, Tajima K, Tominaga S. Lifestyle factors associated with atrophic gastritis among Helicobacter pylori-seropositive Japanese-Brazilians in São Paulo. Int J Clin Oncol. 2003;8:362–368. doi: 10.1007/s10147-003-0355-3. [DOI] [PubMed] [Google Scholar]
- 27.Sasazuki S, Sasaki S, Tsubono Y, Okubo S, Hayashi M, Kakizoe T, Tsugane S. The effect of 5-year vitamin C supplementation on serum pepsinogen level and Helicobacter pylori infection. Cancer Sci. 2003;94:378–382. doi: 10.1111/j.1349-7006.2003.tb01450.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Takaishi S, Wang TC. Providing AID to p53 mutagenesis. Nat Med. 2007;13:404–406. doi: 10.1038/nm0407-404. [DOI] [PubMed] [Google Scholar]
- 29.Higashi H, Nakaya A, Tsutsumi R, Yokoyama K, Fujii Y, Ishikawa S, Higuchi M, Takahashi A, Kurashima Y, Teishikata Y, et al. Helicobacter pylori CagA induces Ras-independent morphogenetic response through SHP-2 recruitment and activation. J Biol Chem. 2004;279:17205–17216. doi: 10.1074/jbc.M309964200. [DOI] [PubMed] [Google Scholar]
- 30.Saadat I, Higashi H, Obuse C, Umeda M, Murata-Kamiya N, Saito Y, Lu H, Ohnishi N, Azuma T, Suzuki A, et al. Helicobacter pylori CagA targets PAR1/MARK kinase to disrupt epithelial cell polarity. Nature. 2007;447:330–333. doi: 10.1038/nature05765. [DOI] [PubMed] [Google Scholar]
- 31.Hong S, Lee HJ, Kim SJ, Hahm KB. Connection between inflammation and carcinogenesis in gastrointestinal tract: focus on TGF-beta signaling. World J Gastroenterol. 2010;16:2080–2093. doi: 10.3748/wjg.v16.i17.2080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hatakeyama M. Helicobacter pylori and gastric carcinogenesis. J Gastroenterol. 2009;44:239–248. doi: 10.1007/s00535-009-0014-1. [DOI] [PubMed] [Google Scholar]
- 33.Hatakeyama M. SagA of CagA in Helicobacter pylori pathogenesis. Curr Opin Microbiol. 2008;11:30–37. doi: 10.1016/j.mib.2007.12.003. [DOI] [PubMed] [Google Scholar]
- 34.Hatakeyama M. Linking epithelial polarity and carcinogenesis by multitasking Helicobacter pylori virulence factor CagA. Oncogene. 2008;27:7047–7054. doi: 10.1038/onc.2008.353. [DOI] [PubMed] [Google Scholar]
- 35.Covacci A, Censini S, Bugnoli M, Petracca R, Burroni D, Macchia G, Massone A, Papini E, Xiang Z, Figura N. Molecular characterization of the 128-kDa immunodominant antigen of Helicobacter pylori associated with cytotoxicity and duodenal ulcer. Proc Natl Acad Sci USA. 1993;90:5791–5795. doi: 10.1073/pnas.90.12.5791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tummuru MK, Cover TL, Blaser MJ. Cloning and expression of a high-molecular-mass major antigen of Helicobacter pylori: evidence of linkage to cytotoxin production. Infect Immun. 1993;61:1799–1809. doi: 10.1128/iai.61.5.1799-1809.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Censini S, Lange C, Xiang Z, Crabtree JE, Ghiara P, Borodovsky M, Rappuoli R, Covacci A. cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors. Proc Natl Acad Sci USA. 1996;93:14648–14653. doi: 10.1073/pnas.93.25.14648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Akopyants NS, Clifton SW, Kersulyte D, Crabtree JE, Youree BE, Reece CA, Bukanov NO, Drazek ES, Roe BA, Berg DE. Analyses of the cag pathogenicity island of Helicobacter pylori. Mol Microbiol. 1998;28:37–53. doi: 10.1046/j.1365-2958.1998.00770.x. [DOI] [PubMed] [Google Scholar]
- 39.Segal ED, Cha J, Lo J, Falkow S, Tompkins LS. Altered states: involvement of phosphorylated CagA in the induction of host cellular growth changes by Helicobacter pylori. Proc Natl Acad Sci USA. 1999;96:14559–14564. doi: 10.1073/pnas.96.25.14559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Asahi M, Azuma T, Ito S, Ito Y, Suto H, Nagai Y, Tsubokawa M, Tohyama Y, Maeda S, Omata M, et al. Helicobacter pylori CagA protein can be tyrosine phosphorylated in gastric epithelial cells. J Exp Med. 2000;191:593–602. doi: 10.1084/jem.191.4.593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Higashi H, Tsutsumi R, Muto S, Sugiyama T, Azuma T, Asaka M, Hatakeyama M. SHP-2 tyrosine phosphatase as an intracellular target of Helicobacter pylori CagA protein. Science. 2002;295:683–686. doi: 10.1126/science.1067147. [DOI] [PubMed] [Google Scholar]
- 42.Tartaglia M, Niemeyer CM, Shannon KM, Loh ML. SHP-2 and myeloid malignancies. Curr Opin Hematol. 2004;11:44–50. doi: 10.1097/00062752-200401000-00007. [DOI] [PubMed] [Google Scholar]
- 43.Neel BG, Gu H, Pao L. The 'Shp'ing news: SH2 domain-containing tyrosine phosphatases in cell signaling. Trends Biochem Sci. 2003;28:284–293. doi: 10.1016/S0968-0004(03)00091-4. [DOI] [PubMed] [Google Scholar]
- 44.Viala J, Chaput C, Boneca IG, Cardona A, Girardin SE, Moran AP, Athman R, Mémet S, Huerre MR, Coyle AJ, et al. Nod1 responds to peptidoglycan delivered by the Helicobacter pylori cag pathogenicity island. Nat Immunol. 2004;5:1166–1174. doi: 10.1038/ni1131. [DOI] [PubMed] [Google Scholar]
- 45.Chamaillard M, Hashimoto M, Horie Y, Masumoto J, Qiu S, Saab L, Ogura Y, Kawasaki A, Fukase K, Kusumoto S, et al. An essential role for NOD1 in host recognition of bacterial peptidoglycan containing diaminopimelic acid. Nat Immunol. 2003;4:702–707. doi: 10.1038/ni945. [DOI] [PubMed] [Google Scholar]
- 46.Goto Y, Ando T, Yamamoto K, Tamakoshi A, El-Omar E, Goto H, Hamajima N. Association between serum pepsinogens and polymorphismof PTPN11 encoding SHP-2 among Helicobacter pylori seropositive Japanese. Int J Cancer. 2006;118:203–208. doi: 10.1002/ijc.21338. [DOI] [PubMed] [Google Scholar]
- 47.Hishida A, Matsuo K, Goto Y, Naito M, Wakai K, Tajima K, Hamajima N. Associations of a PTPN11 G/A polymorphism at intron 3 with Helicobactor pylori seropositivity, gastric atrophy and gastric cancer in Japanese. BMC Gastroenterol. 2009;9:51. doi: 10.1186/1471-230X-9-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kara B, Akkiz H, Doran F, Bayram S, Erken E, Gumurdullu Y, Sandikci M. The significance of E266K polymorphism in the NOD1 gene on Helicobacter pylori infection: an effective force on pathogenesis? Clin Exp Med. 2010;10:107–112. doi: 10.1007/s10238-009-0077-6. [DOI] [PubMed] [Google Scholar]
- 49.Hold GL, Rabkin CS, Chow WH, Smith MG, Gammon MD, Risch HA, Vaughan TL, McColl KE, Lissowska J, Zatonski W, et al. A functional polymorphism of toll-like receptor 4 gene increases risk of gastric carcinoma and its precursors. Gastroenterology. 2007;132:905–912. doi: 10.1053/j.gastro.2006.12.026. [DOI] [PubMed] [Google Scholar]
- 50.Hishida A, Matsuo K, Goto Y, Mitsuda Y, Hiraki A, Naito M, Wakai K, Tajima K, Hamajima N. Toll-like receptor 4 +3725 G/C polymorphism, Helicobacter pylori seropositivity, and the risk of gastric atrophy and gastric cancer in Japanese. Helicobacter. 2009;14:47–53. doi: 10.1111/j.1523-5378.2009.00659.x. [DOI] [PubMed] [Google Scholar]
- 51.Tahara T, Arisawa T, Shibata T, Hirata I, Nakano H. Association of polymorphism of TLR4 and CD14 genes with gastroduodenal diseases in Japan. Inflammopharmacology. 2007;15:124–128. doi: 10.1007/s10787-006-1567-8. [DOI] [PubMed] [Google Scholar]
- 52.Togawa S, Joh T, Itoh M, Katsuda N, Ito H, Matsuo K, Tajima K, Hamajima N. Interleukin-2 gene polymorphisms associated with increased risk of gastric atrophy from Helicobacter pylori infection. Helicobacter. 2005;10:172–178. doi: 10.1111/j.1523-5378.2005.00308.x. [DOI] [PubMed] [Google Scholar]
- 53.Kato I, Canzian F, Franceschi S, Plummer M, van Doorn LJ, Lu Y, Gioia-Patricola L, Vivas J, Lopez G, Severson RK, et al. Genetic polymorphisms in anti-inflammatory cytokine signaling and the prevalence of gastric precancerous lesions in Venezuela. Cancer Causes Control. 2006;17:1183–1191. doi: 10.1007/s10552-006-0060-4. [DOI] [PubMed] [Google Scholar]
- 54.Goto Y, Hishida A, Matsuo K, Tajima K, Morita E, Naito M, Wakai K, Hamajima N. PRKCH gene polymorphism is associated with the risk of severe gastric atrophy. Gastric Cancer. 2010;13:90–94. doi: 10.1007/s10120-009-0542-7. [DOI] [PubMed] [Google Scholar]
- 55.Goto Y, Ando T, Naito M, Goto H, Hamajima N. Inducible nitric oxide synthase polymorphism is associated with the increased risk of differentiated gastric cancer in a Japanese population. World J Gastroenterol. 2006;12:6361–6365. doi: 10.3748/wjg.v12.i39.6361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hishida A, Matsuo K, Goto Y, Mitsuda Y, Naito M, Wakai K, Tajima K, Hamajima N. Significant association of RUNX3 T/A polymorphism at intron 3 (rs760805) with the risk of gastric atrophy in Helicobacter pylori seropositive Japanese. J Gastroenterol. 2009;44:1165–1171. doi: 10.1007/s00535-009-0118-7. [DOI] [PubMed] [Google Scholar]
- 57.Tahara T, Shibata T, Arisawa T, Nakamura M, Yoshioka D, Okubo M, Maruyama N, Kamano T, Kamiya Y, Fujita H, et al. The BB genotype of heat-shock protein (HSP) 70-2 gene is associated with gastric pre-malignant condition in H. pylori-infected older patients. Anticancer Res. 2009;29:3453–3458. [PubMed] [Google Scholar]
- 58.Sun LP, Guo XL, Zhang Y, Chen W, Bai XL, Liu J, Yuan Y. Impact of pepsinogen C polymorphism on individual susceptibility to gastric cancer and its precancerous conditions in a Northeast Chinese population. J Cancer Res Clin Oncol. 2009;135:1033–1039. doi: 10.1007/s00432-008-0539-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Uno M, Ito LS, Oba-Shinjo SM, Marie SK, Shinjo SK, Hamajima N. Possible association of interleukin 1B C-31T polymorphism among Helicobacter pylori seropositive Japanese Brazilians with susceptibility to atrophic gastritis. Int J Mol Med. 2004;14:421–426. [PubMed] [Google Scholar]
- 60.Samloff IM. Pepsinogens I and II: purification from gastric mucosa and radioimmunoassay in serum. Gastroenterology. 1982;82:26–33. [PubMed] [Google Scholar]
- 61.El-Omar EM, Ng MT, Hold GL. Polymorphisms in Toll-like receptor genes and risk of cancer. Oncogene. 2008;27:244–252. doi: 10.1038/sj.onc.1210912. [DOI] [PubMed] [Google Scholar]
- 62.Maeda S, Akanuma M, Mitsuno Y, Hirata Y, Ogura K, Yoshida H, Shiratori Y, Omata M. Distinct mechanism of Helicobacter pylori-mediated NF-kappa B activation between gastric cancer cells and monocytic cells. J Biol Chem. 2001;276:44856–44864. doi: 10.1074/jbc.M105381200. [DOI] [PubMed] [Google Scholar]
- 63.Ferwerda B, McCall MB, Verheijen K, Kullberg BJ, van der Ven AJ, Van der Meer JW, Netea MG. Functional consequences of toll-like receptor 4 polymorphisms. Mol Med. 2008;14:346–352. doi: 10.2119/2007-00135.Ferwerda. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Knuefermann P, Nemoto S, Baumgarten G, Misra A, Sivasubramanian N, Carabello BA, Vallejo JG. Cardiac inflammation and innate immunity in septic shock: is there a role for toll-like receptors? Chest. 2002;121:1329–1336. doi: 10.1378/chest.121.4.1329. [DOI] [PubMed] [Google Scholar]
- 65.Opal SM, DePalo VA. Anti-inflammatory cytokines. Chest. 2000;117:1162–1172. doi: 10.1378/chest.117.4.1162. [DOI] [PubMed] [Google Scholar]
- 66.Wambura C, Aoyama N, Shirasaka D, Sakai T, Ikemura T, Sakashita M, Maekawa S, Kuroda K, Inoue T, Ebara S, et al. Effect of Helicobacter pylori-induced cyclooxygenase-2 on gastric epithelial cell kinetics: implication for gastric carcinogenesis. Helicobacter. 2002;7:129–138. doi: 10.1046/j.1083-4389.2002.00069.x. [DOI] [PubMed] [Google Scholar]
- 67.Pham TN, Rahman P, Tobin YM, Khraishi MM, Hamilton SF, Alderdice C, Richardson VJ. Elevated serum nitric oxide levels in patients with inflammatory arthritis associated with co-expression of inducible nitric oxide synthase and protein kinase C-eta in peripheral blood monocyte-derived macrophages. J Rheumatol. 2003;30:2529–2534. [PubMed] [Google Scholar]
- 68.Pham TN, Brown BL, Dobson PR, Richardson VJ. Protein kinase C-eta (PKC-eta) is required for the development of inducible nitric oxide synthase (iNOS) positive phenotype in human monocytic cells. Nitric Oxide. 2003;9:123–134. doi: 10.1016/j.niox.2003.09.006. [DOI] [PubMed] [Google Scholar]
- 69.Farinati F, Cardin R, Bortolami M, Nitti D, Basso D, de Bernard M, Cassaro M, Sergio A, Rugge M. Oxidative DNA damage in gastric cancer: CagA status and OGG1 gene polymorphism. Int J Cancer. 2008;123:51–55. doi: 10.1002/ijc.23473. [DOI] [PubMed] [Google Scholar]
- 70.Nakabeppu Y, Tsuchimoto D, Ichinoe A, Ohno M, Ide Y, Hirano S, Yoshimura D, Tominaga Y, Furuichi M, Sakumi K. Biological significance of the defense mechanisms against oxidative damage in nucleic acids caused by reactive oxygen species: from mitochondria to nuclei. Ann N Y Acad Sci. 2004;1011:101–111. doi: 10.1007/978-3-662-41088-2_11. [DOI] [PubMed] [Google Scholar]
- 71.Zhang DE, Hetherington CJ, Tan S, Dziennis SE, Gonzalez DA, Chen HM, Tenen DG. Sp1 is a critical factor for the monocytic specific expression of human CD14. J Biol Chem. 1994;269:11425–11434. [PubMed] [Google Scholar]
- 72.Williams TM, Eisenberg L, Burlein JE, Norris CA, Pancer S, Yao D, Burger S, Kamoun M, Kant JA. Two regions within the human IL-2 gene promoter are important for inducible IL-2 expression. J Immunol. 1988;141:662–666. [PubMed] [Google Scholar]
- 73.Hoffmann SC, Stanley EM, Darrin Cox E, Craighead N, DiMercurio BS, Koziol DE, Harlan DM, Kirk AD, Blair PJ. Association of cytokine polymorphic inheritance and in vitro cytokine production in anti-CD3/CD28-stimulated peripheral blood lymphocytes. Transplantation. 2001;72:1444–1450. doi: 10.1097/00007890-200110270-00019. [DOI] [PubMed] [Google Scholar]
- 74.Hoffmann SC, Stanley EM, Cox ED, DiMercurio BS, Koziol DE, Harlan DM, Kirk AD, Blair PJ. Ethnicity greatly influences cytokine gene polymorphism distribution. Am J Transplant. 2002;2:560–567. doi: 10.1034/j.1600-6143.2002.20611.x. [DOI] [PubMed] [Google Scholar]
- 75.Howard TD, Whittaker PA, Zaiman AL, Koppelman GH, Xu J, Hanley MT, Meyers DA, Postma DS, Bleecker ER. Identification and association of polymorphisms in the interleukin-13 gene with asthma and atopy in a Dutch population. Am J Respir Cell Mol Biol. 2001;25:377–384. doi: 10.1165/ajrcmb.25.3.4483. [DOI] [PubMed] [Google Scholar]
- 76.van der Pouw Kraan TC, van Veen A, Boeije LC, van Tuyl SA, de Groot ER, Stapel SO, Bakker A, Verweij CL, Aarden LA, van der Zee JS. An IL-13 promoter polymorphism associated with increased risk of allergic asthma. Genes Immun. 1999;1:61–65. doi: 10.1038/sj.gene.6363630. [DOI] [PubMed] [Google Scholar]
- 77.Shiotani A, Kamada T, Yamanaka Y, Manabe N, Kusunoki H, Hata J, Haruma K. Sonic hedgehog and CDX2 expression in the stomach. J Gastroenterol Hepatol. 2008;23 Suppl 2:S161–S166. doi: 10.1111/j.1440-1746.2008.05406.x. [DOI] [PubMed] [Google Scholar]
- 78.Li QL, Ito K, Sakakura C, Fukamachi H, Inoue K, Chi XZ, Lee KY, Nomura S, Lee CW, Han SB, et al. Causal relationship between the loss of RUNX3 expression and gastric cancer. Cell. 2002;109:113–124. doi: 10.1016/s0092-8674(02)00690-6. [DOI] [PubMed] [Google Scholar]
- 79.Oshimo Y, Oue N, Mitani Y, Nakayama H, Kitadai Y, Yoshida K, Ito Y, Chayama K, Yasui W. Frequent loss of RUNX3 expression by promoter hypermethylation in gastric carcinoma. Pathobiology. 2004;71:137–143. doi: 10.1159/000076468. [DOI] [PubMed] [Google Scholar]
- 80.Fukamachi H, Ito K, Ito Y. Runx3-/- gastric epithelial cells differentiate into intestinal type cells. Biochem Biophys Res Commun. 2004;321:58–64. doi: 10.1016/j.bbrc.2004.06.099. [DOI] [PubMed] [Google Scholar]
- 81.Levanon D, Brenner O, Otto F, Groner Y. Runx3 knockouts and stomach cancer. EMBO Rep. 2003;4:560–564. doi: 10.1038/sj.embor.embor868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Waki T, Tamura G, Sato M, Terashima M, Nishizuka S, Motoyama T. Promoter methylation status of DAP-kinase and RUNX3 genes in neoplastic and non-neoplastic gastric epithelia. Cancer Sci. 2003;94:360–364. doi: 10.1111/j.1349-7006.2003.tb01447.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Becker J, Craig EA. Heat-shock proteins as molecular chaperones. Eur J Biochem. 1994;219:11–23. doi: 10.1007/978-3-642-79502-2_2. [DOI] [PubMed] [Google Scholar]
- 84.Yeo M, Park HK, Kim DK, Cho SW, Kim YS, Cho SY, Paik YK, Hahm KB. Restoration of heat shock protein70 suppresses gastric mucosal inducible nitric oxide synthase expression induced by Helicobacter pylori. Proteomics. 2004;4:3335–3342. doi: 10.1002/pmic.200400951. [DOI] [PubMed] [Google Scholar]
- 85.Hsu PI, Lu PJ, Wang EM, Ger LP, Lo GH, Tsay FW, Chen TA, Yang HB, Chen HC, Lin WS, et al. Polymorphisms of death pathway genes FAS and FASL and risk of premalignant gastric lesions. Anticancer Res. 2008;28:97–103. [PubMed] [Google Scholar]
- 86.Ogasawara N, Tsukamoto T, Mizoshita T, Inada KI, Ban H, Kondo S, Takasu S, Ushijima T, Ito K, Ito Y, et al. RUNX3 expression correlates with chief cell differentiation in human gastric cancers. Histol Histopathol. 2009;24:31–40. doi: 10.14670/HH-24.31. [DOI] [PubMed] [Google Scholar]
- 87.Partida-Rodríguez O, Torres J, Flores-Luna L, Camorlinga M, Nieves-Ramírez M, Lazcano E, Perez-Rodríguez M. Polymorphisms in TNF and HSP-70 show a significant association with gastric cancer and duodenal ulcer. Int J Cancer. 2010;126:1861–1868. doi: 10.1002/ijc.24773. [DOI] [PubMed] [Google Scholar]
- 88.You WC, Hong JY, Zhang L, Pan KF, Pee D, Li JY, Ma JL, Rothman N, Caporaso N, Fraumeni JF Jr, et al. Genetic polymorphisms of CYP2E1, GSTT1, GSTP1, GSTM1, ALDH2, and ODC and the risk of advanced precancerous gastric lesions in a Chinese population. Cancer Epidemiol Biomarkers Prev. 2005;14:451–458. doi: 10.1158/1055-9965.EPI-04-0311. [DOI] [PubMed] [Google Scholar]
- 89.Rad R, Prinz C, Neu B, Neuhofer M, Zeitner M, Voland P, Becker I, Schepp W, Gerhard M. Synergistic effect of Helicobacter pylori virulence factors and interleukin-1 polymorphisms for the development of severe histological changes in the gastric mucosa. J Infect Dis. 2003;188:272–281. doi: 10.1086/376458. [DOI] [PubMed] [Google Scholar]
- 90.Tahara T, Shibata T, Nakamura M, Yoshioka D, Okubo M, Maruyama N, Kamano T, Kamiya Y, Fujita H, Nagasaka M, et al. COMT gene Val158Met polymorphism influences the severity of intestinal metaplasia in H. pylori infected older subjects. Hepatogastroenterology. 2009;56:411–415. [PubMed] [Google Scholar]
- 91.Tahara T, Shibata T, Yamashita H, Hirata I, Arisawa T. Effect of cyclin D1 (CCND1) polymorphism on gastric premalignant condition. Clin Chem Lab Med. 2008;46:1696–1701. doi: 10.1515/CCLM.2008.340. [DOI] [PubMed] [Google Scholar]
- 92.Tahara T, Shibata T, Wang F, Nakamura M, Okubo M, Yoshioka D, Sakata M, Nakano H, Hirata I, Arisawa T. Association of polymorphism of the p22PHOX component of NADPH oxidase in gastroduodenal diseases in Japan. Scand J Gastroenterol. 2009;44:296–300. doi: 10.1080/00365520701702348. [DOI] [PubMed] [Google Scholar]
- 93.Tahara T, Arisawa T, Shibata T, Nakamura M, Yamashita H, Yoshioka D, Okubo M, Maruyama N, Kamano T, Kamiya Y, et al. Effect of polymorphisms in the 3'-untranslated region (3'-UTR) of VEGF gene on gastric pre-malignant condition. Anticancer Res. 2009;29:485–489. [PubMed] [Google Scholar]
- 94.Kato I, Canzian F, Plummer M, Franceschi S, van Doorn LJ, Vivas J, Lopez G, Lu Y, Gioia-Patricola L, Severson RK, et al. Polymorphisms in genes related to bacterial lipopolysaccharide/peptidoglycan signaling and gastric precancerous lesions in a population at high risk for gastric cancer. Dig Dis Sci. 2007;52:254–261. doi: 10.1007/s10620-006-9303-1. [DOI] [PubMed] [Google Scholar]
- 95.Goto Y, Hamajima N, Honda H, Matsuo K, Yamamoto K, Tamakoshi A, Ando T, Goto H. Association between Helicobacter pylori seropositivity and NAD(P)H:quinone oxidoreductase 1 (NQO1) C609T polymorphism observed in outpatients and health checkup examinees. Gastric Cancer. 2005;8:12–17. doi: 10.1007/s10120-004-0308-1. [DOI] [PubMed] [Google Scholar]
- 96.Mimuro H, Suzuki T, Tanaka J, Asahi M, Haas R, Sasakawa C. Grb2 is a key mediator of helicobacter pylori CagA protein activities. Mol Cell. 2002;10:745–755. doi: 10.1016/s1097-2765(02)00681-0. [DOI] [PubMed] [Google Scholar]
- 97.Churin Y, Al-Ghoul L, Kepp O, Meyer TF, Birchmeier W, Naumann M. Helicobacter pylori CagA protein targets the c-Met receptor and enhances the motogenic response. J Cell Biol. 2003;161:249–255. doi: 10.1083/jcb.200208039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Tsutsumi R, Higashi H, Higuchi M, Okada M, Hatakeyama M. Attenuation of Helicobacter pylori CagA x SHP-2 signaling by interaction between CagA and C-terminal Src kinase. J Biol Chem. 2003;278:3664–3670. doi: 10.1074/jbc.M208155200. [DOI] [PubMed] [Google Scholar]
- 99.Amieva MR, Vogelmann R, Covacci A, Tompkins LS, Nelson WJ, Falkow S. Disruption of the epithelial apical-junctional complex by Helicobacter pylori CagA. Science. 2003;300:1430–1434. doi: 10.1126/science.1081919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Kimman TG, Banus S, Reijmerink N, Reimerink J, Stelma FF, Koppelman GH, Thijs C, Postma DS, Kerkhof M. Association of interacting genes in the toll-like receptor signaling pathway and the antibody response to pertussis vaccination. PLoS One. 2008;3:e3665. doi: 10.1371/journal.pone.0003665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Tsukino H, Hanaoka T, Otani T, Iwasaki M, Kobayashi M, Hara M, Natsukawa S, Shaura K, Koizumi Y, Kasuga Y, et al. hOGG1 Ser326Cys polymorphism, interaction with environmental exposures, and gastric cancer risk in Japanese populations. Cancer Sci. 2004;95:977–983. doi: 10.1111/j.1349-7006.2004.tb03186.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Goto M, Shinmura K, Yamada H, Tsuneyoshi T, Sugimura H. OGG1, MYH and MTH1 gene variants identified in gastric cancer patients exhibiting both 8-hydroxy-2'-deoxyguanosine accumulation and low inflammatory cell infiltration in their gastric mucosa. J Genet. 2008;87:181–186. doi: 10.1007/s12041-008-0028-0. [DOI] [PubMed] [Google Scholar]
- 103.Nardone G, Rocco A, Malfertheiner P. Review article: helicobacter pylori and molecular events in precancerous gastric lesions. Aliment Pharmacol Ther. 2004;20:261–270. doi: 10.1111/j.1365-2036.2004.02075.x. [DOI] [PubMed] [Google Scholar]
- 104.Katsuda N, Hamajima N, Matsuo K, Saito T, Ito LS, Inoue M, Takezaki T, Tajima K, Tominaga S. [Association between the interleukin 1B (C-31T) polymorphism and Helicobacter pylori infection in health checkup examinees] Nippon Koshu Eisei Zasshi. 2001;48:604–612. [PubMed] [Google Scholar]
- 105.Atsuta Y, Ito LS, Oba-Shinjo SM, Uno M, Shinjo SK, Marie SK, Goto Y, Hamajima N. Associations of TNF-A-1031TT and -857TT genotypes with Helicobacter pylori seropositivity and gastric atrophy among Japanese Brazilians. Int J Clin Oncol. 2006;11:140–145. doi: 10.1007/s10147-005-0549-y. [DOI] [PubMed] [Google Scholar]
- 106.Kato I, van Doorn LJ, Canzian F, Plummer M, Franceschi S, Vivas J, Lopez G, Lu Y, Gioia-Patricola L, Severson RK, et al. Host-bacterial interaction in the development of gastric precancerous lesions in a high risk population for gastric cancer in Venezuela. Int J Cancer. 2006;119:1666–1671. doi: 10.1002/ijc.21979. [DOI] [PubMed] [Google Scholar]
- 107.Goto Y, Ando T, Nishio K, Ishida Y, Kawai S, Goto H, Hamajima N. The ACE gene polymorphism is associated with the incidence of gastric cancer among H. pylori seropositive subjects with atrophic gastritis. Asian Pac J Cancer Prev. 2005;6:464–467. [PubMed] [Google Scholar]
- 108.Hellmig S, Mascheretti S, Fölsch U, Schreiber S. Functional promotor polymorphism in RANTES gene does not influence the clinical course of Helicobacter pylori infection. J Gastroenterol Hepatol. 2005;20:405–408. doi: 10.1111/j.1440-1746.2005.03606.x. [DOI] [PubMed] [Google Scholar]
- 109.Goto Y, Ando T, Naito M, Goto H, Hamajima N. No association of an SDHC gene polymorphism with gastric cancer. Asian Pac J Cancer Prev. 2006;7:525–528. [PubMed] [Google Scholar]
- 110.Canedo P, Corso G, Pereira F, Lunet N, Suriano G, Figueiredo C, Pedrazzani C, Moreira H, Barros H, Carneiro F, et al. The interferon gamma receptor 1 (IFNGR1) -56C/T gene polymorphism is associated with increased risk of early gastric carcinoma. Gut. 2008;57:1504–1508. doi: 10.1136/gut.2007.143578. [DOI] [PubMed] [Google Scholar]