

Abstract
Inspired by studies demonstrating mate-choice copying effects in non-human species, recent studies of attractiveness judgements suggest that social learning also influences human preferences. In the first part of our article, we review evidence for social learning effects on preferences in humans and other animals. In the second part, we present new empirical evidence that social learning not only influences the attractiveness of specific individuals, but can also generalize to judgements of previously unseen individuals possessing similar physical traits. The different conditions represent different populations and, once a preference arises in a population, social learning can lead to the spread of preferences within that population. In the final part of our article, we discuss the theoretical basis for, and possible impact of, biases in social learning whereby individuals may preferentially copy the choices of those with high status or better access to critical information about potential mates. Such biases could mean that the choices of a select few individuals carry the greatest weight, rapidly generating agreement in preferences within a population. Collectively, these issues suggest that social learning mechanisms encourage the spread of preferences for certain traits once they arise within a population and so may explain certain cross-cultural differences.
Keywords: social transmission, facial attractiveness, copy, learning, prestige bias, culture
1. Introduction
Individuals often learn from others and selection for social learning mechanisms may occur when there are costs to acquiring accurate behavioural information via individual learning [1]. In terms of mate choices and preferences, using the judgement of others may be beneficial if it allows an individual to assess potential mates more quickly and efficiently than through individual trial and error. Indeed, there are potentially very large costs if individuals choose their mates badly, with desertion, infidelity or violence posing very real risks [2]. In this article, we review the evidence that social learning influences mate preferences in both humans and non-human species (§2), present evidence that the effects of social learning can generalize to judgements of previously unseen individuals and discuss how generalized learning is potentially related to between-population variation in attraction (§§3 and 4), and discuss both the theoretical basis for and possible benefits associated with biases in the effects of social learning on attraction in humans (§5). More fundamentally, our aim is to highlight the utility of considering the effects of social learning in order to gain new insights into the processes that shape human mate preferences and human variability in preference.
2. social learning and mate preferences in non-human species and humans
(a). Mate-choice copying in non-human species: a brief review
Mate-choice copying has been observed among females in a number of different non-human species [3–6], including fish [7–10] and bird species [11–13]. Such studies have generally shown that when females observe another female (the model) to be paired with one of two males (the targets), they are subsequently more likely to prefer the target male they had seen paired with the model over the male that was not paired with the model. These effects suggest that social learning may play an important role in female mate choices and preferences in non-human species. The effects of social learning on mate choice in non-human species have generally been demonstrated in the laboratory (e.g. [7,8,11–13]), but similar effects have been found when experiments are conducted in the wild (e.g. [10]). One early example of social learning in the field comes from Höglund et al. [14], who used stuffed female dummies to examine copying effects in sage grouse. They demonstrated that female grouse mated preferentially with the male that appeared to have other females, which were in fact stuffed models, in his territory.
While most of the research described above has focused on the choices of females for males, males in some non-human species also appear to copy the mate choices of other males. In species where males are often the chosen rather than the choosy sex (e.g. in the lek breeding bird species described above), males may not need to mate-choice copy. In other mating systems, however, benefits to males may lead to copying. For example, male sailfin mollies copy the apparent preferences of other males [10,15]. In this species, copying may be advantageous because of a short period of sexual receptivity in females, leading to a need for males to efficiently identify females who are receptive [10,15].
(b). Social learning and human mate preferences
Inspired by work on non-human species, recent research also suggests social learning may influence human mate preferences. While some research has shown that the presence of wedding rings on men did not increase women's preferences for those men [16], other studies have found that images of men labelled as married were more attractive than those labelled as single [17] and that women rate men as more desirable when they are shown surrounded by women than when they are shown alone or with other men [18]. Another study has shown that women prefer pictures of men that had been previously seen paired with images of other women who were looking at the face with smiling (i.e. positive) expressions compared with pictures of men that had been seen paired with images of women with neutral (i.e. relatively negative) expressions [19]. Women therefore do appear to mimic the attitude of other women to particular men.
Alongside partnership status, simple presence and expressions of attitude towards the male, the physical traits of the observed model may also play a role in social transmission of preference. Previous studies have shown that men and women are influenced in their judgements of attractiveness by the apparent choice of attractive members of the same sex. Sigall & Landy [20] used real individuals to show that positive characteristics are attributed more frequently to men who are paired with attractive rather than unattractive women. In this way, they show that an attractive partner may ‘radiate beauty’. Such a phenomenon suggests a more sophisticated form of mate-choice copying, whereby women can use the attractiveness of a partner that a man can acquire in order to judge the man's own attractiveness. We discuss the idea of ‘bias’ in transmission of preference in more detail in §5 of this article. Another study using images that were presented with a fictitious partner has shown that both men and women find a face paired with an attractive partner to be more attractive than one paired with an unattractive partner for a long-term but not a short-term relationship [21]. Other studies have also demonstrated a similar effect for women judging male attractiveness [22]. Effects specific to long-term preferences in humans suggest that social information is being used to infer non-physical traits that make a target a good long-term partner, such as resources or intelligence, which may be difficult to determine from physical appearance alone.
(c). The benefits of learning
Mate-choice copying (usually in females) has been proposed to be adaptive when there is a cost, such as time or energy, to evaluating the quality of potential mates or when discriminating between the quality of potential mates is difficult [23]. In this way, social transmission may allow individuals to assess a potential mate quickly and efficiently and perhaps helps individuals learn what to look for in a mate. In humans, there are many aspects to a partner other than their physical traits that may be valued, and others' choices may be used to infer positive or negative traits, such as behaviour, resources or intelligence, that are difficult to infer from physical appearance alone. These are the sort of traits we might expect to be important for long-term but not necessarily short-term relationships.
In humans, as most individuals will partner during their lives [24], indiscriminately valuing men with partners is unlikely to be a useful mechanism for identifying high-quality partners. Humans bring two factors to a mating relationship: direct benefits such as resources for parental investment or a willingness to invest in children and indirect benefits such as high fertility or potential heritable genetic benefits (e.g. genes for high-quality immune systems). Social information may be more useful for judging direct benefits, given that such information is less likely to be available from physical appearance than is information about indirect benefits. In other species without male parental care, mate-choice copying most probably occurs because individuals are able to acquire information about the association between physical characteristics and the genetic quality of a prospective mate [10]. The results of Little et al. [21], which demonstrate that men are as influenced as women are by the choices of other men, also differ from effects seen in some non-human species where males actively avoid mating with a female after they have seen her interacting with other males [25]. We note here that humans may be different from other species owing to both men and women highly valuing positive personality traits in long-term partners [26]. Importantly, such traits may be usefully inferred by examining the attractiveness of a person's partner. In fact, we might predict that in any species with monogamous relationships or biparental care, there will be pressure on both males and females to choose partners with qualities that are not necessarily signalled by physical appearance, leading to social information being both useful and used. While avoidance of recently mated individuals may be of concern to males in species where male investment and relationship length are limited, evidence of recent mating may not be such an issue for species that might use social information for long-term mate-choice decisions.
(d). Copying directed at specific individuals and the impact of social learning
To date, work on humans has focused on the effects of social learning on perceptions of specific target individuals. In other words, these studies have only investigated how certain models increase attractiveness of their paired individual targets. There are, however, obvious disadvantages to being attracted to individuals who are already partnered, particularly in a species like humans where pairs can remain in committed long-term relationships. Although humans do pursue short-term strategies and extra-pair copulations [27], the effects of social learning on preferences in humans appear to be more focused on long-term preferences than short-term preferences [21]. The potential of mate desertion or mate poaching [28], however, would allow social learners to take advantage of other people's choices. Generalization, whereby individuals learn about the traits of those chosen and find those traits attractive in other individuals, also provides a solution to this problem. Generalization is also probably an important component in the spread of preferences for certain traits through a population. Thus, generalization would be important in generating cross-cultural variation in preferences. We discuss these issues further and present evidence for generalization of social learning in §§3 and 4.
3. cultural variation in preference and the spread of preference for traits: extending social learning beyond the observed pairs
Studies of human attractiveness have documented several traits that are proposed to be attractive across individuals and cultures, potentially reflecting species-wide ‘universal’ preferences. These include preferences for facial traits such as masculinity and symmetry [29,30]. In addition to these general preferences, studies and reviews have pointed to how individual and cultural differences in preferences are, or can also be, consistent with evolutionary predictions [31–34].
While considerable agreement is found in judgements of facial attractiveness within a particular culture, as well as across different cultures (e.g. [35]; see [36] for a meta-analytic review), there may also be differences in the relative attractiveness of certain traits between cultures. Darwin [37], for example, was struck by cultural differences in attractiveness criteria, such as preferences for skin colour, body hair, body fat and practices such as lip ornamentation and tooth filing. Such convictions are supported by early cross-cultural work by Ford & Beach [38], who catalogued differences between cultures in preferences for various aspects of female physique and also by more recent work documenting systematic cross-cultural variation in the importance of physical traits [39,40] and in preferences for facial masculinity [41,42]. Likewise, variation is seen in symmetry preferences across cultures; while individuals from the UK and the Hadza in Tanzania both demonstrate preferences for symmetry in faces, preferences for face symmetry are stronger in the Hadza [29].
Agreement within a culture and differences between cultures could, at least in part, be the result of learning about attractiveness by examining the choices of others in the population. Copying the traits of the choices of others, rather than their preference for a specific individual, could lead to the spread of agreement on which individuals and traits are attractive within a culture. Because arbitrary choices can arise in different cultures, social learning can lead to cultures valuing different traits and the individuals who possess those traits. Of course, arbitrary choice is not itself adaptive, but could be the result of adaptive mechanisms operating on neutral traits. Using the judgement of others may then allow the copying of both adaptively important traits and also other arbitrary traits. Social learning may also influence evolutionarily relevant traits in adaptive ways, acting to tune individuals into traits that are locally adaptive. For example, there is much variation in masculinity preferences, and this has been proposed to link to evolutionarily relevant trade-offs in choosing masculine partners [30,43]. If masculinity is more or less valuable under certain conditions, then social learning may provide a mechanism that promotes following locally adaptive choices. In this way, social learning mechanisms may: (i) reinforce relatively culturally invariant preferences for traits that universally predict adaptive outcomes, (ii) lead to culturally variant preferences for traits for which the adaptive value depends on ecological conditions, and (iii) encourage the spread of preferences for traits that do not appear to serve an adaptive function once they arise within a population. We note that there are also likely biases towards preferences for certain traits, such as symmetry, and these preferences may appear irrespective of learning, though learning may impact on them.
Research on preferences for body traits highlights potentially adaptive variation in attractiveness judgements. For example, there appears to be a positive association between body fat and prestige in the South Pacific [44,45]. In contrast, in cultures such as the UK and the USA, where food is abundant, individuals show strong preferences for low-weight, or thin, individuals [46,47]. These findings are in line with other preference studies where fatness is preferred in cultures where food is less abundant [48–50] and in individuals of low socio-economic status within cultures [51]. Potentially, as body fat, at least in women, is preferred in cultures that are at greater risk of food shortage [50], environmental conditions of subsistence living may place more emphasis on weight as a valued trait in partners. Individual differences are not necessarily restricted to comparisons among cultures but have also been reported within cultures. In line with ideas of food shortage, those in less affluent contexts prefer heavier women than those in more affluent contexts [51].
In previous studies, differences in weight preferences for female bodies were compared between Caucasian individuals from the UK and Zulu individuals from South Africa [52]. South African Zulus prefer higher body mass indexes (BMIs) than people in the UK. Such preferences appear to be the result of learning the norm of attractiveness, as Zulus who moved to the UK have preferences more similar to Caucasian individuals and UK residents of African origin who were born and raised in the UK also have similar preferences to Caucasian individuals [52]. In this way, preferences for weight need not be innate but may be adaptable to the environment in which individuals find themselves. The mechanism for learning what is attractive within a particular culture remains unclear, but social learning is a prime candidate.
(a). Generalization in social learning
Previous studies on humans have focused on copying-like effects on preferences for specific individuals. In other words, they have shown that a particular individual becomes more attractive when paired with an attractive partner. This is also seen in the non-human animal literature, where copying has been defined as simply the acceptance or rejection of potential mates on the basis of their having been accepted or rejected by others (e.g. [53,54]). Copying effects, however, can be more general, with females showing preferences for novel males that are physically similar to males observed with other females [9,12,13]. This generalization of preferences for certain traits, and not just preferences for specific individuals, is important as it allows social transmission to have more wide-reaching consequences on the evolution of certain anatomical traits and preferences [55].
Social transmission of mate choice can have important consequences for sexual selection [55]. Darwin [37] laid out the first notions of how evolution of traits by preference could occur. A common example of sexual selection is self-reinforcing, or ‘runaway’, selection [56], which provides one account for the evolution of traits and preferences. After a preference for any particular trait has arisen, such as a preference for large noses, females begin to reproduce with males in possession of large noses to produce offspring with both genes for large noses (in males) and genes for a preference for large noses (in females). A feedback loop between genes for traits and preferences could produce stronger preferences and ever more elaborate expression of traits. The initial preference could come from a sensory disposition evolved for another purpose [57], and hence may not necessarily serve a function, or because the preferred trait is associated with either phenotypic or genotypic quality [58]. Modelling work has shown that social transmission of preferences in humans can result in a directional pressure on both traits and preferences within populations, and this could potentially account for genetically based phenotypic variation between cultures [55].
There is another problem, alluded to earlier, with copying the choices of others. The problem is that the specific person by definition is already partnered and, at least in humans, the partnership could be long lasting, thus limiting the benefits of social copying. Generalized preferences for the traits of individuals who are chosen, rather than specific individuals themselves, means that copying effects will not necessarily lead to the problem of increasing your preference for a person who already has a partner, thus avoiding direct competition in competing for partnered individuals and increasing the adaptive benefits of social learning of preferences.
(b). Rationale for the current experiment
In our experiment, we extended previous research demonstrating that social learning can influence preferences for specific individuals to examine whether copying-like effects extend to judgements of novel faces that share the facial traits of members of the opposite sex that were observed paired with attractive partners. The experiment is designed to address at a small scale what could happen in different human populations. Our experimental conditions can be regarded as separate human populations that are exposed to choices of other individuals within their population. While previous studies demonstrate attraction to specific individuals based on social learning, learning about the attractiveness of traits in general opens the possibility of social learning having much more wide-ranging effects on the preferences of a population.
(i). Participants
Participants were 53 heterosexual adults (21 women, 32 men, mean age = 30.6 years, s.d. = 10.9). All participants were volunteers visiting an online testing site and were selected for being heterosexual and between the ages of 16 and 60. Participants were largely white (white: 77.4%, Asian: 9.5%, Hispanic: 3.8%, black: 1.5%, other: 7.5%). The study was approved by the ethics committee in the Department of Psychology, University of Stirling. The study was run over the Internet and participants were volunteers visiting a research-based website (www.alittlelab.com).
(ii). Consent and participation
Participants were given a description of the study prior to filling in the questionnaire and instructed that by continuing to the next page they were consenting to take part in the study and that they were free to drop out at any point. Participants were unsupervised and undertook the test at their own computer.
(iii). Stimuli
The base stimuli were 10 composite male and 10 composite female faces. Each stimulus was composed of five randomly selected male or female images from a set of 50 young adult male and 50 young adult female facial photographs that had been taken under standard lighting conditions with neutral facial expressions. The composite faces were created using specially designed software. Key locations (179 points) were manually marked around the main features (e.g. eyes, nose and mouth) and the outline of each face (e.g. jaw line, hair line). The average location of each feature point in the five faces for each composite was then calculated. The features of the individual faces were then warped to the relevant average shape before superimposing the images to produce a photographic quality result (this technique has been used to create composite images in previous studies: [59–61]). The composite images were made perfectly symmetrical by combining them with their mirror image, creating images symmetrical in both shape and colour.
Eye spacing was manipulated by transforming all images relative to a pair of face images, one original image and one image where all the points delineating the eyes had been moved outwards. The distance change in eye-spacing distance from original (measured from the centre of the eye) in the presented images was 21 pixels for each individual face, either wider or narrower. The same transform was applied to all starting images, ensuring the wide and narrow eye-spacing images differed from the real starting images in an identical manner but in opposite directions. This procedure for manipulating eye spacing in face images has been used in previous studies.
Attractive and unattractive versions of the same base faces were manufactured using a masculine/feminine transform, following our previous work [21]. Femininity is reliably associated with the attractiveness of female faces [62,63]. While masculinity in male faces is not so consistently associated with attractiveness across different studies using different sets of stimuli, previous studies using the same stimuli as those used here have established that masculine versions are, on average, preferred over feminine versions in the male face set [21,64]. For reasons of clarity, when referring to model images, we refer to masculine male faces and feminine female faces as ‘attractive’ and feminine male faces and masculine female faces as ‘unattractive’. Composites were transformed on a masculine/feminine dimension using the linear difference between a composite of 50 males and 50 females following the technique reported in Perrett et al. [62]. Using the shape difference between male and female composites, the vector of masculinity/femininity can be parametrized allowing manipulation along the vector, described here as a percentage of the distance between male and female. Transforms represented 50 per cent plus or minus the difference between these two composites. Examples of manipulated stimuli can be seen in figure 1.
Figure 1.

Examples of stimuli pairs used in the experiment reported in §§3 and 4 of our article. In condition A, narrow eye spacing was paired with attractive partners and wide eye spacing with unattractive partners. In condition B, narrow eye spacing was paired with unattractive partners and wide eye spacing with attractive partners.
(iv). Procedure
Briefly, we pre-tested participants' preference for eye spacing in opposite-sex faces, and then exposed participants to pairs of faces in which novel, opposite-sex target individuals were paired with attractive or unattractive partners (the attractiveness of the partner depended on the eye spacing of the target) before repeating the test of preference for eye spacing. We compared pre- with post-test scores to determine whether the eye spacing that was paired with attractive partners increased in attractiveness. Details of this procedure are given below.
Participants were presented with a short questionnaire assessing age, sex and sexual orientation and were then given a pre-test for eye-spacing preference. They were presented with five novel face pairs (five male pairs for women and five female pairs for men), which comprised a wide-eyed and narrow-eyed version of the same composite, and were asked to choose which face they thought was most attractive for a long-term relationship. A long-term relationship was specified because previous research has demonstrated that social learning effects on face preferences are greater when women assess men's attractiveness for long-term relationship contexts than for short-term relationship contexts [21]. Clicking a button under the image selected it as more attractive and moved onto the next trial.
Following this pre-test were exposure trials, where participants were shown 10 pairs of male and female faces and were told that the person on the right (model) was the partner of the person on the left (target). Participants were randomly allocated to one of two exposure conditions. In condition (or population) A, narrow eye spacing was paired with attractive partners and wide eye spacing with unattractive partners. In condition (or population) B, narrow eye spacing was paired with unattractive partners and wide eye spacing with attractive partners. Different faces were used in the exposure test than were used in the pre- and post-tests.
After this exposure, we measured post-test masculinity preference by again presenting the five face pairs from the pre-test. Preference for eye spacing was recorded in both the pre- and post-test eye-spacing preference tests as a percentage of times participants chose the wide-eyed image of the pair. All image pairs in each set of trials were presented in a random order.
4. Results
The dependent variable ‘change in wide eye-spacing preference’ was calculated by subtracting the pre-exposure preference for wide eye spacing from the post-exposure preference. Positive scores therefore indicate preferences for wide eye spacing that increased after exposure and negative scores indicate preferences for wide eye spacing that decreased after exposure. Generalization of social learning would be evidenced by scores that were greater for participants who saw wide eye spacing paired with attractive partners than for participants who saw wide eye spacing paired with unattractive partners.
A univariate ANOVA was carried out with change in wide eye-spacing preference as the dependent variable and condition (wide eye spacing paired with attractive faces, wide eye spacing paired with unattractive faces) and sex of participant (male, female) as between-participant factors. This revealed a significant effect of condition (F1,49 = 8.73, p = 0.005, 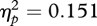 ), no significant effect of sex of participant (F1,49 = 0.06, p = 0.813,
), no significant effect of sex of participant (F1,49 = 0.06, p = 0.813, 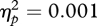 ) and no significant interaction between sex of participant and condition (F1,49 = 0.85, p = 0.362,
) and no significant interaction between sex of participant and condition (F1,49 = 0.85, p = 0.362, 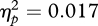 ). The main effect of condition reflects the predicted effect that preferences for wide eye spacing were higher after observing wide eye spacing paired with attractive models and were lower when wide eye spacing was paired with unattractive models. Indeed, in both conditions, preferences for wide eye spacing changed from pre-test to post-test, increasing after exposure to wide eye spacing paired with attractive partners (t28 = 1.82, p = 0.079, d = 0.69) and decreasing after exposure to wide eye spacing paired with unattractive partners (t23 = −2.43, p = 0.023, d = 1.01). Mean changes in eye-spacing preference by condition can be seen in figure 2.
). The main effect of condition reflects the predicted effect that preferences for wide eye spacing were higher after observing wide eye spacing paired with attractive models and were lower when wide eye spacing was paired with unattractive models. Indeed, in both conditions, preferences for wide eye spacing changed from pre-test to post-test, increasing after exposure to wide eye spacing paired with attractive partners (t28 = 1.82, p = 0.079, d = 0.69) and decreasing after exposure to wide eye spacing paired with unattractive partners (t23 = −2.43, p = 0.023, d = 1.01). Mean changes in eye-spacing preference by condition can be seen in figure 2.
Figure 2.

Mean (±s.e. of mean) change in preference for wide eye spacing from pre-test to post-test for the experiment reported in §§3 and 4 of our article. Exposure to faces with narrow eye spacing paired with attractive partners decreased preferences for wide eye spacing while exposure to faces with wide eye spacing paired with attractive partners increased preferences for wide eye spacing. White bar, narrow-attractive; grey bar, wide-attractive.
(a). Summary of the current experiment
Our experiment demonstrated that general preferences for face traits in opposite-sex faces are influenced by the pairing of an attractive/unattractive face with other opposite-sex faces in possession of that trait. This finding extends previous research by demonstrating that the effects of social learning on attractiveness judgements generalize to judgments of novel individuals. When targets with wide eye spacing were paired with an attractive model and targets with narrow eye spacing were paired with an unattractive model, preferences for wide eye spacing increased in both men and women compared with the preferences of participants who were exposed to the opposite pairing.
(b). Generalizing social learning and cross-cultural differences in preference
Generalization, as demonstrated above, greatly increases the influence social learning can potentially have across different cultures as it means certain traits, not certain individuals, can rapidly become preferred in a population. If we consider our two experimental conditions as different human populations, then population A came to prefer narrow-spaced eyes and population B came to prefer wide-spaced eyes after only minimal exposure to pairing individuals with those traits with attractive partners.
If attractive individuals in a population exhibit some consistency in preferences for certain traits, then the effect seen in our experiment can lead to selection for the preferred traits and increased frequency of genes for such preferences. This would be in line with runaway selection described earlier, as even weak transmission biases can generate population-specific traits and preferences [55]. In this way, social learning could create consistency in traits and preferences within a specific culture and also generate cross-cultural differences once differences arise. Such transfer effects may allow for the social transmission of mate-choice preferences to influence the evolution of certain anatomical traits and preferences [55].
Copying the choices of others can be an important part of sexual selection [23]. Mathematical models of mate choice have suggested that social transmission of mate preferences can contribute to sexual selection for male traits [55,65,66]. In all three models, depending on the strength of copying effects, mate-choice copying can cause directional selection, meaning that copying is capable of reducing variation in traits present in a population and causing novel traits, and preferences for these traits, to increase in frequency in a population [55,65,66].
Overall, our findings demonstrate the plausibility of cultural inheritance of mating preferences in humans driving sexual selection consistent with the model proposed by Laland [55]. Future studies could address whether the social learning effects seen here are observable in other cultures as well as document the extent of cross-cultural variation in mate preferences/physical traits and their genetic basis, which may be the result of such learning.
Our findings are consistent with a sophisticated copying-like process, whereby individuals copy the choices of attractive individuals (as the attractive model increased attraction to their partner). This may represent a biased form of copying whereby individuals are most inclined to follow the choices of attractive people in mate choice. Generalization increases the chances that social learning can drive large changes between cultures, but the impact of social learning could be further increased if there were bias in who is learned from. If a select few are attended to, then the choices of few can lead the preferences of many if social learning generalizes. In §5, we address the possibility of such bias in learning.
5. biases in social learning
In the last part of this review, we turn our attention to factors that might increase the impact of social learning within a population. Certain individuals may have a greater impact than others on social learning, leading to greater conformity to a single norm more quickly. This can occur if there is a bias to attend to information from certain individuals. Such biases in social learning may play important roles in generating and maintaining diversity in preferences both within a population and between populations.
Once social learning evolved, it would pay to be selective about who to learn from because some models will be more successful than others [67]. This has led to the notion that there are mechanisms promoting differential attention to particular models [68,69], so-called ‘model-based biases’ (following [1]). Three aspects of the model that might promote social learning are cues of prestige, success and skill [69]. Learners should preferentially pay attention to people in possession of these cues because selective social learners would then have an advantage over those who were not selective. Indeed, it has been argued that prestige evolved from social learning strategies to identify appropriate models from which to learn [67]. The possession of prestige ensures that prestigious individuals are listened to and have influence. Although the source of prestige can be unclear to observers, if others consider someone to be prestigious, it might be assumed he or she is a useful model for social learning. For example, age and skill are often associated with prestige; in hunter–gatherer societies, skilled individuals have higher status [70] and older individuals are generally seen as people to be respected [71]. While prestige is indirectly inferred through others' behaviour towards the prestigious individual, other cues to useful models are more directly observable. For example, one can observe potential models actively engaged in a particular task, such as hunting, and gauge their skill in order to decide who to learn from [69]. Even without direct observation of skill, observations of an individual's apparent success can also provide cues to their value as a model [69]. In the example of hunting, this might mean the number of animals killed. Thus, bias can be influenced by cues ranging from direct observation at particular tasks to inferred prestige.
Prestige, skill and success certainly seem important in social learning. As noted by Henrich & McElreath [69], Rogers [72] argued that the diffusion of innovations is strongly influenced by ‘local opinion leaders.’ These opinion leaders are generally high in local social status, well respected and widely connected [72]. There are other lines of research that also suggest that social learning is biased towards prestigious or successful models. Several studies demonstrate that social learning is biased in some way. For example, studies have shown that participants are more likely to follow the choices of competent and/or successful individuals [73–77]. Other studies have demonstrated that participants are more likely to follow the behaviour of prestigious individuals [78–80]. Bias has also been seen in the transmission of managerial styles [81,82] and in naturalistic studies [83]. These findings suggest a bias towards competent individuals when such individuals are observed to be successful on particular tasks or even towards those who are of high status/prestige. In non-human species, bias has been observed in fish species whereby young guppies are more likely to copy the mate choice of older models than younger models [8,84]. Older guppies are also less likely to be influenced by the choices of younger females [8]. As age is related to experience, this suggests a sophisticated social learning bias involving trade-offs between personal and public information use (e.g. [85,86]) that could lead to more adaptive learning.
Evidence for biases in the effects of social learning on attractiveness judgements in humans is relatively scarce, but is undoubtedly an important issue for future research. Moreover, what little evidence currently exists for such biases in humans is certainly encouraging, suggesting that tests for such biases may well yield interesting effects. For example, both previous research [21] and the experiment reported in §§3 and 4 of the current article suggest that people are more likely to be influenced by the choices of attractive individuals than unattractive individuals. Because attractive individuals are likely to be most able to be selective of partners, it can be inferred that the partner whom they have chosen possesses attractive qualities that may not be physical. A physically attractive partner may then act as an indirect advertisement of quality. Additionally, the finding that women's preferences for men who were being looked at by other women increased more if the other women were smiling than if they had neutral expressions [19] demonstrates that the model's apparent satisfaction with the target influences social learning. Thus, our previous and current studies already demonstrate some evidence for biased social learning in human mate preferences. Previous authors have also speculated that preferences for body weight may involve social learning [51] and social learning based on observing how weight is associated with prestige is a plausible mechanism for changes in preferences seen when individuals move between cultures [52]. There are other likely biases in human social learning and we expect individuals would be most influenced by others who have greatest access to information, demonstrate success and/or have high prestige.
6. Summary and conclusions
As is clear from §2 of our article, the attractiveness literature has recently seen a steady accumulation of compelling evidence that social learning influences human mate preferences. Adding to this research, our new experiment (reported in §§3 and 4 of this article) demonstrated that social learning effects may extend beyond the specific individuals observed to novel individuals possessing similar physical traits, greatly extending both the scope and power of social learning effects. Section 5 highlighted the importance of considering the potential impact of biases in social learning, whereby individuals may preferentially copy the choices of individuals with high status or better access to critical information about potential mates. Such biases may provide further insight into the importance of social learning for human mate preferences and could mean that the choices of a select few individuals carry the greatest weight, rapidly generating agreement in preferences within a population and allowing social learning to be more influential. Social learning mechanisms encourage the spread of preferences for certain traits once they arise within a population and so may explain certain cross-cultural differences either by driving locally adaptive choice or by selection for arbitrary traits that are relatively neutral to selection. Together, these data and arguments highlight the potential power of social learning in human mate preferences and how understanding social learning processes can help in understanding human mate preferences.
Acknowledgements
A.C.L. is supported by a Royal Society University Research Fellowship. B.C.J. and L.M.D. are supported by ESRC grant RES-000-22-2498. C.A.C. is supported by ESRC grant RES-062-23-1634. Empirical study was designed and run by A.C.L. We thank B. P. Tiddeman and D. I. Perrett for the use of their software.
Footnotes
One contribution of 14 to a Theme Issue ‘Evolution and human behavioural diversity’.
References
- 1.Richerson P. J., Boyd R. 2005. Not by genes alone: how culture transformed human evolution. Chicago, IL: University of Chicago Press [Google Scholar]
- 2.Daly M., Wilson M. 1988. Evolutionary social-psychology and family homicide. Science 242, 519–524 10.1126/science.3175672 (doi:10.1126/science.3175672) [DOI] [PubMed] [Google Scholar]
- 3.Brown G. R., Fawcett T. W. 2005. Sexual selection: copycat mating in birds. Curr. Biol. 15, R626–R628 10.1016/j.cub.2005.08.005 (doi:10.1016/j.cub.2005.08.005) [DOI] [PubMed] [Google Scholar]
- 4.Dugatkin L. A. 2000. The imitation factor: evolution beyond the gene. New York, NY: Free Press [Google Scholar]
- 5.Galef B. G., Laland K. N. 2005. Social learning in animals: empirical studies and theoretical models. Bioscience 55, 489–499 10.1641/0006-3568(2005)055[0489:SLIAES]2.0.CO;2 (doi:10.1641/0006-3568(2005)055[0489:SLIAES]2.0.CO;2) [DOI] [Google Scholar]
- 6.White D. J. 2004. Influences of social learning on mate-choice decisions. Learn. Behav. 32, 105–113 [DOI] [PubMed] [Google Scholar]
- 7.Dugatkin L. A., Godin J. G. J. 1992. Reversal of female mate choice by copying in the guppy (Poecilia reticulata). Proc. R. Soc. Lond. B 249, 179–184 10.1098/rspb.1992.0101 (doi:10.1098/rspb.1992.0101) [DOI] [PubMed] [Google Scholar]
- 8.Dugatkin L. A., Godin J. G. J. 1993. Female mate copying in the guppy (Poecilia reticulata)—age-dependent effects. Behav. Ecol. 4, 289–292 10.1093/beheco/4.4.289 (doi:10.1093/beheco/4.4.289) [DOI] [Google Scholar]
- 9.Godin J. G. J., Herdman E. J. E., Dugatkin L. A. 2005. Social influences on female mate choice in the guppy, Poecilia reticulata: generalized and repeatable trait-copying behaviour. Anim. Behav. 69, 999–1005 10.1016/j.anbehav.2004.07.016 (doi:10.1016/j.anbehav.2004.07.016) [DOI] [Google Scholar]
- 10.Witte K., Ryan M. J. 2002. Mate choice copying in the sailfin molly, Poecilia latipinna, in the wild. Anim. Behav. 63, 943–949 10.1006/anbe.2001.1982 (doi:10.1006/anbe.2001.1982) [DOI] [Google Scholar]
- 11.Galef B. G., White D. J. 1998. Mate-choice copying in Japanese quail, Coturnix coturnix japonica. Anim. Behav. 55, 545–552 [DOI] [PubMed] [Google Scholar]
- 12.Swaddle J. P., Cathey M. G., Correll M., Hodkinson B. P. 2005. Socially transmitted mate preferences in a monogamous bird: a non-genetic mechanism of sexual selection. Proc. R. Soc. B 272, 1053–1058 10.1098/rspb.2005.3054 (doi:10.1098/rspb.2005.3054) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.White D. J., Galef B. G. 2000. ‘Culture’ in quail: social influences on mate choices of female Coturnix japonica. Anim. Behav. 59, 975–979 10.1006/anbe.1999.1402 (doi:10.1006/anbe.1999.1402) [DOI] [PubMed] [Google Scholar]
- 14.Hoglund J., Alatalo R. V., Gibson R. M., Lundberg A. 1995. Mate-choice copying in black grouse. Anim. Behav. 49, 1627–1633 10.1016/0003-3472(95)90085-3 (doi:10.1016/0003-3472(95)90085-3) [DOI] [Google Scholar]
- 15.Schlupp I., Ryan M. J. 1997. Male sailfin mollies (Poecilia latipinna) copy the mate choice of other males. Behav. Ecol. 8, 104–107 10.1093/beheco/8.1.104 (doi:10.1093/beheco/8.1.104) [DOI] [Google Scholar]
- 16.Uller T., Johansson L. C. 2003. Human mate choice and the wedding ring effect—are married men more attractive? Hum. Nat. 14, 267–276 [DOI] [PubMed] [Google Scholar]
- 17.Eva K. W., Wood T. J. 2006. Are all the taken men good? An indirect examination of mate-choice copying in humans. Can. Med. Assoc. J. 175, 1573–1574 10.1503/cmaj.061367 (doi:10.1503/cmaj.061367) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hill S. E., Buss D. M. 2008. The mere presence of opposite-sex others on judgments of sexual and romantic desirability: opposite effects for men and women. Pers. Soc. Psychol. Bull. 34, 635–647 10.1177/0146167207313728 (doi:10.1177/0146167207313728) [DOI] [PubMed] [Google Scholar]
- 19.Jones B. C., DeBruine L. M., Little A. C., Burriss R. P., Feinberg D. R. 2007. Social transmission of face preferences among humans. Proc. R. Soc. B 274, 899–903 10.1098/rspb.2006.0205 (doi:10.1098/rspb.2006.0205) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sigall H., Landy D. 1973. Radiating beauty—effects of having a physically attractive partner on person perception. J. Pers. Soc. Psychol. 28, 218–224 10.1037/h0035740 (doi:10.1037/h0035740) [DOI] [Google Scholar]
- 21.Little A. C., Burriss R. P., Jones B. C., DeBruine L. M., Caldwell C. A. 2008. Social influence in human face preference: men and women are influenced more for long-term than short-term attractiveness decisions. Evol. Hum. Behav. 29, 140–146 10.1016/j.evolhumbehav.2007.11.007 (doi:10.1016/j.evolhumbehav.2007.11.007) [DOI] [Google Scholar]
- 22.Waynforth D. 2007. Mate choice copying in humans. Hum. Nat. 18, 264–271 [DOI] [PubMed] [Google Scholar]
- 23.Wade M. J., Pruett-Jones S. G. 1990. Female copying increases the variance in male mating success. Proc. Natl Acad. Sci. USA 87, 5749–5753 10.1073/pnas.87.15.5749 (doi:10.1073/pnas.87.15.5749) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Vandenberg S. G. 1972. Assortative mating or who marries whom? Behav. Genet. 2, 127–157 10.1007/BF01065686 (doi:10.1007/BF01065686) [DOI] [PubMed] [Google Scholar]
- 25.White D. J., Galef B. G. 2000. Differences between the sexes in direction and duration of response to seeing a potential sex partner mate with another. Anim. Behav. 59, 1235–1240 10.1006/anbe.1999.1431 (doi:10.1006/anbe.1999.1431) [DOI] [PubMed] [Google Scholar]
- 26.Buss D. M., Schmitt D. P. 1993. Sexual strategies theory: an evolutionary perspective on human mating. Psychol. Rev. 100, 204–232 10.1037/0033-295X.100.2.204 (doi:10.1037/0033-295X.100.2.204) [DOI] [PubMed] [Google Scholar]
- 27.Simpson J. A., Gangestad S. W. 1991. Individual differences in sociosexuality: evidence for convergent and discriminant validity. J. Pers. Indiv. Differ. 60, 870–883 [DOI] [PubMed] [Google Scholar]
- 28.Bleske A. L., Shackleford T. K. 2001. Poaching, promiscuity, and deceit: combating mating rivalry in same-sex friendships. Pers. Relat. 8, 407–424 10.1111/j.1475-6811.2001.tb00048.x (doi:10.1111/j.1475-6811.2001.tb00048.x) [DOI] [Google Scholar]
- 29.Little A. C., Apicella C. L., Marlowe F. W. 2007. Preferences for symmetry in human faces in two cultures: data from the UK and the Hadza, an isolated group of hunter–gatherers. Proc. R. Soc. B 274, 3113–3117 10.1098/rspb.2007.0895 (doi:10.1098/rspb.2007.0895) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Thornhill R., Gangestad S. W. 1999. Facial attractiveness. Trends Cogn. Sci. 3, 452–460 10.1016/S1364-6613(99)01403-5 (doi:10.1016/S1364-6613(99)01403-5) [DOI] [PubMed] [Google Scholar]
- 31.DeBruine L. M., et al. 2006. Correlated preferences for facial masculinity and ideal or actual partner's masculinity. Proc. R. Soc. B 273, 1355–1360 10.1098/rspb.2005.3445 (doi:10.1098/rspb.2005.3445) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Jones B. C., DeBruine L. M., Perrett D. I., Little A. C., Feinberg D. R., Smith M. J. L. 2008. Effects of menstrual cycle phase on face preferences. Arch. Sexual Behav. 37, 78–84 10.1007/s10508-007-9268-y (doi:10.1007/s10508-007-9268-y) [DOI] [PubMed] [Google Scholar]
- 33.Little A., Perrett D. 2002. Putting beauty back in the eye of the beholder. Psychologist 15, 28–32 [Google Scholar]
- 34.Little A. C., Burt D. M., Penton-Voak I. S., Perrett D. I. 2001. Self-perceived attractiveness influences human female preferences for sexual dimorphism and symmetry in male faces. Proc. R. Soc. Lond. B 268, 39–44 10.1098/rspb.2000.1327 (doi:10.1098/rspb.2000.1327) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Cunningham M. R., Roberts A. R., Barbee A. P., Druen P. B. 1995. ‘Their ideas of beauty are, on the whole, the same as ours’: consistency and variability in the cross-cultural perception of female attractiveness. J. Pers. Soc. Psychol. 68, 261–279 10.1037/0022-3514.68.2.261 (doi:10.1037/0022-3514.68.2.261) [DOI] [Google Scholar]
- 36.Langlois J. H., Kalakanis L., Rubenstein A. J., Larson A., Hallamm M., Smoot M. 2000. Maxims or myths of beauty? A meta-analytic and theoretical review. Psychol. Bull. 126, 390–423 10.1037/0033-2909.126.3.390 (doi:10.1037/0033-2909.126.3.390) [DOI] [PubMed] [Google Scholar]
- 37.Darwin C. 1871. The descent of man, and selection in relation to sex. London, UK: John Murray [Google Scholar]
- 38.Ford C. S., Beach F. A. 1951. Patterns of sexual behaviour. New York, NY: Harper & Row [Google Scholar]
- 39.Gangestad S. W., Buss D. M. 1993. Pathogen prevalence and human mate preferences. Ethol. Sociobiol. 14, 89–96 10.1016/0162-3095(93)90009-7 (doi:10.1016/0162-3095(93)90009-7) [DOI] [Google Scholar]
- 40.Gangestad S. W., Haselton M. G., Buss D. M. 2006. Evolutionary foundations of cultural variation: evoked culture and mate preferences. Psychol. Inquiry 17, 75–95 10.1207/s15327965pli1702_1 (doi:10.1207/s15327965pli1702_1) [DOI] [Google Scholar]
- 41.DeBruine L. M., Jones B. C., Crawford J. R., Welling L. M., Little A. C. 2010. The health of a nation predicts their mate preferences: cross-cultural variation in women's preferences for masculinized male faces. Proc. R. Soc. B 277, 2405–2410 10.1098/rspb.2009.2184 (doi:10.1098/rspb.2009.2184) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Penton-Voak I. S., Jacobson A., Trivers R. 2004. Populational differences in attractiveness judgements of male and female faces: comparing British and Jamaican samples. Evol. Hum. Behav. 25, 355–370 [Google Scholar]
- 43.Little A. C., Jones B. C., Penton-Voak I. S., Burt D. M., Perrett D. I. 2002. Partnership status and the temporal context of relationships influence human female preferences for sexual dimorphism in male face shape. Proc. R. Soc. Lond. B 269, 1095–1100 10.1098/rspb.2002.1984 (doi:10.1098/rspb.2002.1984) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Brewis A. A., McGarvey S. T. 2000. Body image, body size, and Samoan ecological and individual modernization. Ecol. Food Nutr. 39, 105–120 10.1080/03670244.2000.9991609 (doi:10.1080/03670244.2000.9991609) [DOI] [Google Scholar]
- 45.Wilkinson J. Y., Bentovim D. I., Walker M. K. 1994. An insight into the personal and cultural significance of weight and shape in large Samoan women. Int. J. Obes. 18, 602–606 [PubMed] [Google Scholar]
- 46.Seifert T. 2005. Anthropomorphic characteristics of centerfold models: trends towards slender figures over time. Int. J. Eat. Disord. 37, 271–274 10.1002/eat.20086 (doi:10.1002/eat.20086) [DOI] [PubMed] [Google Scholar]
- 47.Tovee M. J., Reinhardt S., Emery J. L., Cornelissen P. L. 1998. Optimum body-mass index and maximum sexual attractiveness. Lancet 352, 548. 10.1016/S0140-6736(05)79257-6 (doi:10.1016/S0140-6736(05)79257-6) [DOI] [PubMed] [Google Scholar]
- 48.Anderson J. L., Crawford C. B., Nadeau J., Lindberg T. 1992. Was the Duchess of Windsor right—a cross-cultural review of the socioecology of ideals of female body shape. Ethol. Sociobiol. 13, 197–227 10.1016/0162-3095(92)90033-Z (doi:10.1016/0162-3095(92)90033-Z) [DOI] [Google Scholar]
- 49.Marlowe F., Wetsman A. 2001. Preferred waist-to-hip ratio and ecology. Pers. Indiv. Differ. 30, 481–489 10.1016/S0191-8869(00)00039-8 (doi:10.1016/S0191-8869(00)00039-8) [DOI] [Google Scholar]
- 50.Wetsman A., Marlowe F. 1999. How universal are preferences for female waist-to-hip ratios? Evidence from the Hadza of Tanzania. Evol. Hum. Behav. 20, 219–228 10.1016/S1090-5138(99)00007-0 (doi:10.1016/S1090-5138(99)00007-0) [DOI] [Google Scholar]
- 51.Swami V., Tovee M. J. 2005. Female physical attractiveness in Britain and Malaysia: a crosscultural study. Body Image 2, 115–128 10.1016/j.bodyim.2005.02.002 (doi:10.1016/j.bodyim.2005.02.002) [DOI] [PubMed] [Google Scholar]
- 52.Tovee M. J., Swami V., Furnham A., Mangalparsad R. 2006. Changing perceptions of attractiveness as observers are exposed to a different culture. Evol. Hum. Behav. 27, 443–456 10.1016/j.evolhumbehav.2006.05.004 (doi:10.1016/j.evolhumbehav.2006.05.004) [DOI] [Google Scholar]
- 53.Pruett-Jones S. 1992. Independent versus nonindependent mate choice—do females copy each other. Am. Nat. 140, 1000–1009 [DOI] [PubMed] [Google Scholar]
- 54.Westneat D. F., Walters A., McCarthy T. M., Hatch M. I., Hein W. K. 2000. Alternative mechanisms of nonindependent mate choice. Anim. Behav. 59, 467–476 10.1006/anbe.1999.1341 (doi:10.1006/anbe.1999.1341) [DOI] [PubMed] [Google Scholar]
- 55.Laland K. N. 1994. Sexual selection with a culturally transmitted mating preference. Theor. Popul. Biol. 45, 1–15 10.1006/tpbi.1994.1001 (doi:10.1006/tpbi.1994.1001) [DOI] [PubMed] [Google Scholar]
- 56.Fisher R. A. 1930. The genetical theory of natural selection. Oxford, UK: Clarendon Press [Google Scholar]
- 57.Ryan M. J., Rand S. 1990. The sensory basis of sexual selection for complex calls in the Tungara frog, Physalaemus pustulosus (sexual selection for sensory exploitation). Evolution 44, 305–314 10.2307/2409409 (doi:10.2307/2409409) [DOI] [PubMed] [Google Scholar]
- 58.Andersson M. 1994. Sexual selection. Princeton, NJ: Princeton University Press [Google Scholar]
- 59.Little A. C., Hancock P. J. 2002. The role of masculinity and distinctiveness on the perception of attractiveness in human male faces. Br. J. Psychol. 93, 451–464 10.1348/000712602761381349 (doi:10.1348/000712602761381349) [DOI] [PubMed] [Google Scholar]
- 60.Benson P. J., Perrett D. I. 1993. Extracting prototypical facial images from exemplars. Perception 22, 257–262 10.1068/p220257 (doi:10.1068/p220257) [DOI] [PubMed] [Google Scholar]
- 61.Tiddeman B. P., Burt D. M., Perrett D. I. 2001. Prototyping and transforming facial texture for perception research. IEEE Comput. Graphic. Appl. 21, 42–50 10.1109/38.946630 (doi:10.1109/38.946630) [DOI] [Google Scholar]
- 62.Perrett D. I., Lee K. J., Penton-Voak I. S., Rowland D. R., Yoshikawa S., Burt D. M., Henzi S. P., Castles D. L., Akamatsu S. 1998. Effects of sexual dimorphism on facial attractiveness. Nature 394, 884–887 10.1038/29772 (doi:10.1038/29772) [DOI] [PubMed] [Google Scholar]
- 63.Rhodes G. 2006. The evolutionary psychology of facial beauty. Annu. Rev. Psychol. 57, 199–226 10.1146/annurev.psych.57.102904.190208 (doi:10.1146/annurev.psych.57.102904.190208) [DOI] [PubMed] [Google Scholar]
- 64.Little A. C., Mannion H. 2006. Viewing attractive or unattractive same-sex individuals changes self-rated attractiveness and face preferences in women. Anim. Behav. 72, 981–987 10.1016/j.anbehav.2006.01.026 (doi:10.1016/j.anbehav.2006.01.026) [DOI] [Google Scholar]
- 65.Agrawal A. F. 2001. The evolutionary consequences of mate copying on male traits. Behav. Ecol. Sociobiol. 51, 33–40 10.1007/s002650100401 (doi:10.1007/s002650100401) [DOI] [Google Scholar]
- 66.Kirkpatrick M., Dugatkin L. A. 1994. Sexual selection and the evolutionary effects of copying mate choice. Behav. Ecol. Sociobiol. 34, 443–449 10.1007/BF00167336 (doi:10.1007/BF00167336) [DOI] [Google Scholar]
- 67.Henrich J., Gil-White F. J. 2001. The evolution of prestige—freely conferred deference as a mechanism for enhancing the benefits of cultural transmission. Evol. Hum. Behav. 22, 165–196 10.1016/S1090-5138(00)00071-4 (doi:10.1016/S1090-5138(00)00071-4) [DOI] [PubMed] [Google Scholar]
- 68.Boyd R., Richerson P. J. 1985. Culture and the evolutionary process. Chicago, IL: University of Chicago Press [Google Scholar]
- 69.Henrich J., McElreath R. 2007. Dual inheritance theory: the evolution of human cultural capacities and cultural evolution. In Oxford handbook of evolutionary psychology (eds Dunbar R., Barrett L.). Oxford, UK: Oxford University Press [Google Scholar]
- 70.Kelly R. L. 1995. The foraging spectrum: diversity in hunter-gatherer lifeways. Washington, DC: Smithsonian Institution Press [Google Scholar]
- 71.Silverman P., Maxwell R. J. 1978. How do I respect thee—let me count ways—deference towards elderly men and women. Behav. Sci. Res. 13, 91–108 [Google Scholar]
- 72.Rogers E. M. 1995. Diffusion of innovations. New York, NY: Free Press [Google Scholar]
- 73.Baron R. A. 1970. Attraction toward model and models competence as determinants of adult imitative behavior. J. Pers. Soc. Psychol. 14, 345–351 [DOI] [PubMed] [Google Scholar]
- 74.Greenfeld N., Kuznicki J. T. 1975. Implied competence, task complexity, and imitative behavior. J. Soc. Psychol. 95, 251–261 10.1080/00224545.1975.9918711 (doi:10.1080/00224545.1975.9918711) [DOI] [Google Scholar]
- 75.Kroll Y., Levy H. 1992. Further tests of the separation theorem and the capital-asset pricing model. Am. Econ. Rev. 82, 664–670 [Google Scholar]
- 76.Offerman T., Sonnemans J. 1998. Learning by experience and learning by imitating successful others. J. Econ. Behav. Organ. 34, 559–575 10.1016/S0167-2681(97)00109-1 (doi:10.1016/S0167-2681(97)00109-1) [DOI] [Google Scholar]
- 77.Rosenbaum M. E., Tucker I. F. 1962. Competence of model and learning of imitation and nonimitation. J. Exp. Psychol. 63, 183–190 10.1037/h0042363 (doi:10.1037/h0042363) [DOI] [PubMed] [Google Scholar]
- 78.Bauer G. P., Schlottmann R. S., Bates J. V., Masters M. A. 1983. Effect of state and trait anxiety and prestige of model on imitation. Psychol. Rep. 52, 375–382 [DOI] [PubMed] [Google Scholar]
- 79.Eckel C. C., Wilson R. K. 2007. Social learning in coordination games: does status matter? Exp. Econ. 10, 317–329 10.1007/s10683-007-9185-x (doi:10.1007/s10683-007-9185-x) [DOI] [Google Scholar]
- 80.Ryckman R. M., Sherman M. F., Rodda W. C. 1972. Locus of control and expertise relevance as determinants of changes in opinion about student activism. J. Soc. Psychol. 88, 107–114 [Google Scholar]
- 81.Weiss H. M. 1977. Subordinate imitation of supervisor behavior—role of modeling in organizational socialization. Organ. Behav. Hum. Perform. 19, 89–105 10.1016/0030-5073(77)90056-3 (doi:10.1016/0030-5073(77)90056-3) [DOI] [Google Scholar]
- 82.Weiss H. M., Suckow K., Rakestraw T. L. 1999. Influence of modeling on self-set goals: direct and mediated effects. Hum. Perform. 12, 89–114 [Google Scholar]
- 83.Mullen B., Copper C., Driskell J. E. 1990. Jaywalking as a function of model behavior. Pers. Soc. Psychol. Bull. 16, 320–330 10.1177/0146167290162012 (doi:10.1177/0146167290162012) [DOI] [Google Scholar]
- 84.Amlacher J., Dugatkin L. A. 2005. Preference for older over younger models during mate-choice copying in young guppies. Ethol. Ecol. Evol. 17, 161–169 10.1080/08927014.2005.9522605 (doi:10.1080/08927014.2005.9522605) [DOI] [Google Scholar]
- 85.Kendal R. L., Coolen I., van Bergen Y., Laland K. N. 2005. Trade-offs in the adaptive use of social and asocial learning. Adv. Study Behav. 35, 333–379 10.1016/S0065-3454(05)35008-X (doi:10.1016/S0065-3454(05)35008-X) [DOI] [Google Scholar]
- 86.Laland K. N. 2004. Social learning strategies. Learn. Behav. 32, 4–14 [DOI] [PubMed] [Google Scholar]