


Abstract
An emerging theme in transforming growth factor-β (TGF-β) signalling is the association of the Smad proteins with diverse groups of transcriptional regulatory proteins. Several Smad cofactors have been identified to date but the diversity of TGF-β effects on gene transcription suggests that interactions with other co-regulators must occur. In these studies we addressed the possible interaction of Smad proteins with the myocyte enhancer-binding factor 2 (MEF2) transcriptional regulators. Our studies indicate that Smad2 and 4 (Smad2/4) complexes cooperate with MEF2 regulatory proteins in a GAL4-based one-hybrid reporter gene assay. We have also observed in vivo interactions between Smad2 and MEF2A using co-immunoprecipitation assays. This interaction is confirmed by glutathione S-transferase pull-down analysis. Immunofluorescence studies in C2C12 myotubes show that Smad2 and MEF2A co-localise in the nucleus of multinuclear myotubes during differentiation. Interestingly, phospho-acceptor site mutations of MEF2 that render it unresponsive to p38 MAP kinase signalling abrogate the cooperativity with the Smads suggesting that p38 MAP Kinase-catalysed phosphorylation of MEF2 is a prerequisite for the Smad–MEF2 interaction. Thus, the association between Smad2 and MEF2A may subserve a physical link between TGF-β signalling and a diverse array of genes controlled by the MEF2 cis element.
INTRODUCTION
Members of the transforming growth factor-β (TGF-β) superfamily are multifunctional cytokines that mediate a diversity of cellular responses encompassing growth and differentiation, apoptosis, control of extracellular matrix production and inflammatory responses (1–5). The properties of TGF-β in mediating such a diversity of cellular responses result from their ability to regulate the expression of a multitude of target genes. Therefore, a central question for understanding TGF-β signalling concerns how signals are transduced to the transcriptional machinery in the nucleus so that the cell can mount an appropriate re-programming of gene expression. Initially, the TGF-β pathway is activated by a diverse superfamily of ligands (6–9). Ligand binding by the type II (TβR-II) receptor leads to multimerisation with the type I receptor (TβR-I) (10–13). TβR-II has a constitutively active serine/threonine kinase domain and, upon binding, TGF-β transphosphorylates TβR-I in its glycine/serine-rich domain (14). Activation of the TβR-I serine/threonine kinase activity phosphorylates downstream effector molecules and propagates intracellular transduction of TGF-β signals (5,10,15).
Although the mechanisms for large-scale transcriptional control by TGF-β are not yet well defined, a family of proteins responsible for transducing signals from the activated receptor to the nucleus has been extensively characterised. The Smad protein family, consisting of at least eight members, functions as a central mediator of the transcriptional effects of TGF-β signalling in vertebrates, insects and nematodes (16,17). The name Smad originates from a contraction of the Drosophila protein, MAD (mothers against decapapentaplegic), the first downstream component of serine/threonine kinase receptor signalling to be identified and the Caenorhabditis elegans proteins, sma-2, 3 and 4 (named after a mutant with a phenotype of small body size) (18–21). The Smad proteins are grouped into three classes based on their function: (i) the receptor-activated R-Smads (Smads 1, 2, 3, 5 and 8), of which Smads 2 and 3 are TGF-β and activin responsive and Smads 1, 5 and 8 are BMP2/4 responsive; (ii) the central or common mediator Co-Smads (Smad4), which form heteromeric complexes with R-Smads; and (iii) the antagonistic Smads (Smads 6 and 7), which negatively regulate TGF-β family pathways by blocking receptor-mediated phosphorylation of receptor-activated Smads or sequestering Smad4 (22–26).
Smads contain two Mad-homology domains (MH1 and MH2) separated by a central linker region rich in serine, threonine and proline residues (18–21,27). The intracellular propagation of the TGF-β signal involves the protein SARA (Smad anchor for receptor activation), which contains a lipid-binding FYVE (double zinc finger) domain that interacts directly with the MH2 domains of Smad2 and Smad3. SARA presents Smad2 to the TGF-β receptor, a mechanism that is enhanced once TGF-β signalling is initiated. After being phosphorylated by the type I receptor, Smad2 dissociates from SARA, binds to Smad4 and translocates to the nucleus (28–31). The nuclear Smad complex subsequently activates transcriptional responses of TGF-β activated target genes. Therefore, TGF-β signalling can be viewed as a two-step process which comprises of a ligand-mediated assembly of an activated receptor complex, followed by propagation of the signal in the form of nuclear-localised Smads, which integrate into diverse multi-subunit transcription complexes.
Transcriptional control by the Smad proteins has proven to be inherently complex involving interaction with DNA and transcriptional co-regulators. For example, the Mix.2 gene is induced during the early stages of Xenopus development by activin and requires activation of an activin response element (ARE) by a Smad2–Smad4–FAST-1 complex. FAST-1 is a winged helix transcription factor that physically interacts with, and requires Smads through a C-terminal Smad interacting domain. Smads have also been shown to directly bind to DNA, and crystal structure analysis has shown that the MH1 domain of Smad3 can interact with two different DNA target sites: the 8 bp palindromic DNA sequence, called the Smad binding element (SBE, 5′-AGTATGTCTAGACTGA); and the 4 bp Smad box (SB, 5′-GTCT) (32–36). Clearly, the 4 bp Smad binding site is insufficient to confer precise responses to TGF-β signals. Interestingly, some activin responsive genes, such as the goosecoid promoter, contain elements that do not bear much sequence similarity to the Xenopus Mix.2 ARE (37). Members of the Mix family of paired–like homeodomain transcription factors, Mixer and Milk, as well as the FAST-2 transcription factor, have also been implicated in the recruitment of Smad2 to the goosecoid promoter (38,39). Other studies addressing the requirement for DNA binding of Smads for TGF-β signalling are also equivocal. For example, even though phosphorylation-dependent binding of a Smad3/4 complex is observed on the TGF-β responsive 3TP-Lux reporter gene (derived from the PAI 1 gene promoter), deletion of the Smad binding site does not interfere with TGF-β-dependent transcriptional activation (29,40–42). In Drosophila the MAD protein can bind to a G–C-rich target sequence in the quadrant enhancer of vestigial (vg); however, this binding is of very low affinity suggesting that the primary DNA binding activity of the complex is not provided by MAD (32). Smad3 can also bind Fos/Jun dimers and an AP-1 site in the collagenase I promoter to enhance transcriptional activation of this gene (43). These observations are therefore consistent with the view that, when activated and localised to the nucleus, Smads can form promiscuous interactions with multiple transcription factor complexes bound to different target sequences (44). The emerging model is therefore centred on combinatorial interactions of the Smads with other sequence-specific DNA binding proteins, which allow not only signal divergence but also enhanced specificity in target gene activation (38,45–53). Therefore, characterizing the full array of Smad interacting transcriptional co-regulators will be vital in understanding the mechanisms of transcriptional control by TGF-β signalling.
Myocyte enhancer-binding factor 2 (MEF2) proteins were originally identified as important regulators of gene expression in cardiac and skeletal muscle (54–58). Vertebrate MEF2 genes (A to D) encode nuclear phosphoproteins belonging to the MADS (MCM1, agamous, deficiens, serum response factor) superfamily of DNA binding proteins. The MEF2 proteins bind as homo- and heterodimers to a cis element with the consensus (C/T)TA(A/T)4TA(G/A) via interactions between their N-terminal MADS/MEF2 domain and the major groove of DNA (59,60). This binding site is found in the control region of numerous cardiac and skeletal muscle-specific genes (56,61–64). MEF2 transcripts are widely expressed and these proteins are now known to play a role in differentiation and apoptosis in neurons, T cells, cardiac myocytes and skeletal muscle cells (65–71). Whilst the N-terminus of the MEF2s is a highly conserved DNA binding/dimerisation domain, the more divergent C-termini of the MEF2s are known to function as transcription activation domains (TADs) and the TADs are known to be targeted by MAP Kinase signalling cascades (56,72–76).
Since MEF2 transcription factors fulfil a crucial role in transcriptional control in a variety of TGF-β responsive cell types and are known to interact with heterogeneous partners, we tested the possibility that MEF2 could cooperate with Smads in response to TGF-β signalling. We report that Smad2 can physically associate with MEF2 proteins in vivo and that this interaction leads to a cooperative activation of MEF2 transcriptional activity. Our data suggest that the Smad–MEF2 interaction depends on phosphorylation of the MEF2 transactivation domain by p38 MAP kinase signalling, thus indicating a cooperative intersection of the TGF-β and p38 MAP kinase signalling cascades on a single transcriptional regulator.
MATERIALS AND METHODS
Transcriptional response assays
C2C12 myoblasts (mb) were grown in growth medium: Dulbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum (FBS) in 60-mm dishes. At densities of 60% confluence, cells were transiently transfected with pSV-β-galactosidase (pSV-βgal), as a marker of transfection efficiency, and the indicated constructs using the standard calcium phosphate–DNA precipitation method (77,78). The empty expression vectors pMT2 and pCMV5B were used to bring the total DNA to 15 µg per transfection. pMT2, pCMV5B and GAL4 constructs have been described previously (14,27,30,74,79–81). The growth medium of transfected C2C12 mb was replaced after 24 h by medium containing 1% FBS or 5% horse serum, as indicated, and supplemented with either 2 ng/ml TGF-β (R&D Systems) or an equal volume of the TGF-β diluent [0.1% bovine serum albumin, 4 mM HCl]. Seventy-two hours after transfection, cellular extracts were prepared and the luciferase activity of each extract was measured using a Berthold Lumat luminometer as described by the manufacturer (Promega).
Western immunoblotting
C2C12 mb were plated at 60% confluence in growth media. Twenty-four hours later, the medium was replaced with 5% horse serum (differentiation medium) and either 2 ng/ml TGF-β or an equal volume of the TGF-β diluent. Cells were collected after 2, 4 and 6 days. To examine the levels of endogenous Smad2 in C2C12 cells, equal amounts of cell extract were resolved by 10% SDS–PAGE and electrophoretically transferred to Nitroplus nitro-cellulose transfer membrane (MSI). Immunoblots were probed with anti-Smad2 antibody (Signal Transduction) at a 1:500 dilution and analysed using Western Chemiluminescence Reagent (Du Pont NEN) to detect specific secondary antibodies as described previously (77,78).
Immunoprecipitation
COS cells were transiently transfected with the indicated constructs using the calcium phosphate–DNA precipitation method. C2C12 mb and myotubes (mt) were grown in growth medium for 2 days and differentiation medium for 4 days, respectively. Whole cell extracts were lysed with the NP-40 lysis buffer (150 mM NaCl, 1% NP-40, 50 mM Tris, pH 8.0, 1 mM sodium orthovanadate, 1 mM phenylmethylsulfonyl fluoride, 1 mM dithiothreitol and protease inhibitors). Supernatants were subjected to immunoprecipitation with either anti-MEF2A, anti-Smad2 (Signal Transduction Labs) or anti-phospho-Smad2 (Upstate Biotech.) antibodies. Precipitates were analysed by immunoblotting as described previously (78). To check for non-specific interactions, negative control assays were performed in the same manner, using normal mouse immunoglobin (IgG) antibody (Santa Cruz, sc-2025). Anti-MEF2A antibody was kindly provided by R. Prywes and used at 1:1000 dilution. Anti-Smad2 and anti-phospho-Smad2 antibodies were used at 1:500 dilutions. Normal mouse IgG antibodies were used at dilutions of 1:1000.
Glutathione S-transferase (GST) pull-down assays
Aliquots of 1 µg hMEF2C-pGem 7zF(+) and hMEF2A-pGem 7zF(+) (Promega) were transcribed, translated and labelled with [35S]methionine according to the manufacturer’s instructions (MBI Fermentas). The constructs: hMEF2C–pGem 7zF(+) (1–435 amino acids) and hMEF2A–pGem 7zF(+) (1–507 amino acids) have been described previously (56,76). Aliquots of 2 and 2.5 µl of in vitro translated MEF2A and MEF2C products, respectively, were incubated in 90 µl of binding buffer (50 mM Tris pH 7.6, 150 mM NaCl, 5 mM EDTA, 0.1% Tween-20 and 10 µg/ml of the following protease inhibitors: aprotinin, leupeptin and pepstatin) with ~5 µg of either GST or GST–Smad2 fusion proteins immobilised on glutathione–agarose beads (Sigma). The mixtures were gently mixed for 1 h at room temperature. The beads were then washed twice with washing buffer (50 mM Tris pH 7.6, 450 mM NaCl, 5 mM EDTA, 0.1% Tween-20). Proteins retained on beads were boiled in 10 µl of sodium dodecyl sulfate (SDS) sample buffer (New England Biolabs) and subjected to 10% SDS–polyacrylamide gel electrophoresis (PAGE). The gels were stained with Coomassie blue dye (GelCode, Pierce) to show the amount of loaded GST fusion proteins. The presence of labelled MEF2 proteins was detected by autoradiography. The bacterial expression construct for GST–Smad2 consists of a full-length clone of Smad2 (1–467 amino acids) fused into the pGEX4T-1 vector (Pharmacia) as described elsewhere (83,84).
Subcellular localisation by immunofluorescence confocal microscopy
To detect the localisation of MEF2A in the presence of Flag-tagged dominant negative Smad2, MADR2(3SA), C2C12 mb were transiently transfected with the indicated constructs using lipofectamine (Gibco), grown in growth medium for 3 days, fixed with methanol and probed with antibodies raised against the Flag epitope, MEF2A or Smad2. To detect endogenous levels of MEF2A, Smad1, Smad2, phosphorylated Smad2 and Smad3 in C2C12 mt; mb were grown in differentiation medium for 4 days replacing the medium every 2 days. At this point, mt were either fixed for immunofluorescence or exposed to TGF-β (+TGF-β) for 24 h. Fixation and reaction with the primary and secondary antibodies was performed as described previously (78). Localisation of endogenous Smad1, Smad2, phosphorylated Smad2, Smad3 and MEF2A were detected using isoform-specific antibodies.
RESULTS
TGF-β responsive co-modulators: Smad2 and Smad4 enhance the activity of MEF2A and MEF2C transcription factors
To determine if the Smad proteins regulate MEF2 activity, the effects of exogenous Smad proteins on MEF2 transcriptional activity were analysed using GAL4-based one-hybrid reporter assays (Fig. 1). C2C12 cells, which are known to be TGF-β responsive, were used to determine whether the Smad–MEF2 cooperativity was ligand-sensitive and could be stimulated through endogenous TGF-β receptors. C2C12 cells were co-transfected with expression vectors for Smad2, 3, 4, GAL4–MEF2A, GAL4–MEF2C and GAL4(DBD) as indicated (Fig. 1A and B). Twenty-four hours after transfection, the growth medium of C2C12 mb was replaced by medium containing 1% FBS and supplemented with either 2 ng/ml TGF-β or an equal volume of the TGF-β diluent. One percent FBS is known to contain 0.1 ng/ml TGF-β, thus the comparison studied was between low (0.1 ng/ml) and high levels (2.1 ng/ml) of TGF-β. The over-expression of Smad2 and 4 (Smad2/4) was found to increase the activities of both GAL4–MEF2A and GAL4–MEF2C in high TGF-β conditions. Additional experiments were carried out in COS cells (data not shown), and the effects observed by over-expressing TGF-β responsive Smads with GAL4–MEF2A or GAL4–MEF2C were comparable to experiments performed in C2C12 cells.
Figure 1.

TGF-β responsive co-modulators, Smad2 and 4, increase the activities of both GAL4–MEF2A and GAL4–MEF2C. (A and B) C2C12 cells were co-transfected with pCMV5B–TβRI(T204D), pCMV5B–Smad2, pCMV5B–Smad4, 5×GAL4-luc, GAL4(DBD), GAL4–MEF2A, GAL4–MEF2C and pSV-βgal as indicated and treated with (+) or without (–) 2 ng/ml of TGF-β in 1% FBS. The transcriptional activation domains of both MEF2 isoforms are fused to the DNA binding domain (DBD) of GAL4 to form GAL4–MEF2A (91–507 amino acids) and GAL4–MEF2C (87–442 amino acids). GAL4(DBD) contains the GAL4 DBD only. The reporter plasmid, 5×GAL4-luc, contains five copies of the GAL4 binding site linked to the adenovirus EIB promoter and the firefly luciferase gene. After 72 h, β-galactosidase activities were measured and used to normalise luciferase activity values. Two or more sets of assays were performed in both COS and C2C12 cells with comparable results. Each data point is a mean of triplicate samples from single experiments and the error bars represent the standard error of the mean (SEM).
In TGF-β responsive C2C12 cells, the cooperativity of Smad2 and 4 with GAL4–MEF2A and GAL4–MEF2C is ligand and receptor sensitive.
In order to determine whether ligand-activated TGF-β receptors can regulate the effects of the Smads on GAL4–MEF2 activity, C2C12 cells were co-transfected with either wild-type TGF-β receptors or a dominant negative TGF-β receptor (Fig. 2A–D). The dominant negative receptor, TβRII(K277R), contains a substitution of lysine for arginine in its ATP-binding site and is still able to form a ligand-bound heterotetramer receptor complex but is unable to phosphorylate TβR-I (80). The enhancement of GAL4–MEF2 activity by Smad2/4 was found to be increased by the over-expression of TGF-β receptor complexes and was negated by expression of a dominant negative receptor, TβRII(K277R) (Fig. 2A and C). It was also found that exogenous TGF-β-activated receptors and Smad2/4 increased the activities of both GAL4–MEF2A and GAL4–MEF2C. Alternatively, TGF-β-activated wild-type receptors in combination with Smad3 and 4, repressed the activity of GAL4–MEF2A and GAL4–MEF2C (Fig. 2B and D).
Figure 2.

In C2C12 cells, the activities of both GAL4–MEF2A and GAL4–MEF2C are enhanced by the over-expression of Smad2 and 4 with wild-type TGF-β receptors (TβR-I and -II), but not in the presence of a TGF-β dominant negative receptor, TβRII(K277R). (A–D) C2C12 cells were co-transfected with pCMV5B–TβRII(K277R), pCMV5B–TβR-I, pCMV5B–TβR-II, pCMV5B–Smad2, pCMV5B–Smad3, pCMV5B–Smad4, 5×GAL4-luc, GAL4(DBD), GAL4–MEF2A(91–507), GAL4–MEF2C(87–442) and pSV-βgal as indicated. Twenty-four hours after transfection, growth medium was replaced with 1% FBS ± 2 ng/ml of TGF-β. Cells were harvested 72 h after transfection for luciferase assays. β-galactosidase activities were used to normalise for transfection efficiency. Two or more sets of experiments were performed with comparable results. Each data point is the mean of triplicate samples from single experiments and the error bars represent the SEM.
Smad2 and MEF2A are complexed together in C2C12 mt
COS cells were co-transfected with expression vectors for Smad2, 4, MEF2A and the constitutively-active type I TGF-β receptor, TβRI(T204D), and cellular extracts were used for co-immunoprecipitation (Fig. 3A and B). COS cells express low levels of TGF-β receptors and therefore exhibit little background from endogenous TGF-β signalling (30). To reconstitute TGF-β signalling, COS cells were co-transfected with a constitutively-active form of the TGF-β type I receptor, TβRI(T204D). An interaction between Smad2 and MEF2A was detected by probing a blot containing immunoprecipitated MEF2A with the anti-Smad2 antibody. The reverse experiment was also done and MEF2A was detected in the Smad2 immunoprecipitate. To determine whether a physical interaction exists between endogenous Smad2 and MEF2A in cultured cells, Smad2 was immunoprecipitated from both mb and mt and probed with an anti-MEF2A antibody (Fig. 3C). MEF2A was detected in Smad2 immunoprecipitate from mt extracts. Conversely, Smad2 was detected in MEF2A immunoprecipitate from mt but not from mb (data not shown). In addition, a negative control assay was done using normal mouse IgG antibody instead of anti-Smad2 antibody showing that MEF2A does not bind protein Sepharose A gel or mouse IgG antibodies non-specifically (Fig. 3D). Detection of an association between endogenous Smad2 and MEF2A indicates that endogenous MEF2 activity in mt may be regulated by Smad2 and TGF-β.
Figure 3.

A physical interaction between endogenous Smad2 and MEF2A in C2C12 mt was detected through co-immunoprecipitation. (A and B) COS cells were transiently transfected with pCMV5B–TβRI(T204D), pCMV5B–Smad2, pCMV5B–Smad4 and pMT2–MEF2A as indicated. Cell lysates were immunoprecipitated with antibodies raised against either MEF2A (α-MEF2A), or Smad2 (α-Smad2). (A) An immunoblot probed by an α-Smad2 antibody contains: 15 µl of anti-MEF2A immunoprecipitate (α-MEF2A ip) in lanes 1 and 3; 15 µl of anti-Smad2 immunoprecipitate (α-Smad2 ip) in lanes 5 and 7; and 30 µg of C2C12 cell extract in lane 9. The arrow indicates Smad2 protein. (B) An immunoblot probed by an α-MEF2A antibody contains: 15 µl of α-Smad2 ip in lane 1; 1 µl of α-Smad2 ip in lane 2; 20 µg of C2C12 cell extract in lane 4. Lane 3 was empty (C) An immunoblot probed by an α-MEF2A antibody contains: a molecular weight marker in lane 1; 1 µl of α-MEF2A ip from myoblast lysate (mb) in lane 2; 1 µl of α-MEF2A ip from myotube lysate (mt) in lane 3; 20 µl of α-Smad2 ip from mb in lane 5; and 20 µl of α-Smad2 ip from mt in lane 6. (D) A negative control for the co-immunoprecipitation assays was performed. An immunoblot probed with an α-MEF2A antibody contains: 20 µl of α-mouse IgG ip from mt lysate in lane 1; 50 µg of mt cellular extract (mt) in lane 3; 1 µl of mt lysate in lane 5 (used for α-Smad2 ip in lane 7); 20 µl of α-Smad2 ip from mt in lane 7; 15 µl of pre-cleared supernatant from α-Smad2 ip (lane 7) in lane 9; and a molecular weight marker in lane 10. Lanes 2, 4, 6 and 8 were empty. (B–D) The arrow indicates MEF2A protein. (A–D) The arrowheads indicate IgG recognised by the goat anti-rabbit secondary antibody. The bands below the IgG are non-specific. (E) C2C12 mb were plated at 25% confluence and 24 h later the medium was replaced with differentiation medium with (+) or without (–) 2 ng/ml TGF-β. After 2, 4 and 6 days (d), protein expression was analysed by loading 20 µg of each extract on a SDS–polyacrylamide gel and western blotting using an anti-Smad2 antibody. Jurkats cell extract (JC) was used as a control.
Smad2 is expressed at equal levels in C2C12 mb and mt
It has been previously shown that C2C12 cells express Smad1, 2, 4 and 5 mRNAs and that their expression levels are not altered by treatment with TGF-β (85). To further extend these data, an analysis was done over a 6 day time course during the differentiation programme from mb to mt. Thus, in order to determine if the level of Smad2 protein changes during myogenesis, extracts were examined by western blotting at 2, 4 and 6 days after C2C12 mb had been transferred to differentiation medium with or without TGF-β (Fig. 3E). Smad2 expression was found to remain constant over the time course, and did not change with the addition of TGF-β.
In vitro association of Smad2 with both MEF2A and MEF2C is detected in GST pull-down assays
To further confirm the interaction between Smad2 and MEF2 proteins, we carried out a GST pull-down assay using immobilised GST and GST–Smad2 fusion proteins and [35S]methionine-labelled MEF2 (Fig. 4). In vitro translated and 35S-labelled MEF2A or MEF2C were incubated with immobilised GST or GST–Smad2. After washing, the beads were boiled in SDS buffer and the supernatants analysed by SDS–PAGE to determine the presence and amount of associated MEF2 proteins. As seen in Figure 4, MEF2A proteins (lane 4) and MEF2C proteins (lane 7) were retained by GST–Smad2, but not by GST (lanes 3 and 6). These data further confirm a physical association between Smad2 and MEF2A.
Figure 4.
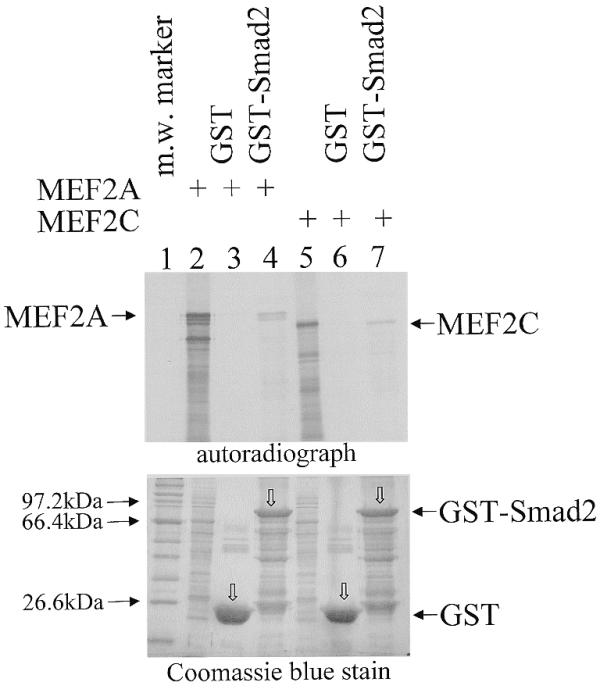
MEF2 proteins associate with GST–Smad2 in vitro. (Top) In a GST pull-down assay, in vitro translated [35S]methionine-labelled MEF2A (2 µl) or MEF2C (2.5 µl) was mixed with 5 µg of GST (lanes 3 and 6) or GST–Smad2 (lanes 4 and 7) immobilised on glutathione–agarose beads. After washing, 35S-labelled bound proteins were analysed by SDS–PAGE and autoradiography. Aliquots of 0.4 µl MEF2A (lane 2) and 0.5 µl MEF2C (lane 5) in vitro translation reactions were included as references. The arrows indicate the position of MEF2A or MEF2C. (Bottom) Coomassie blue staining of the gel shows that comparable amount of the GST or GST–Smad2 proteins were loaded. The open arrows (lanes 3, 4, 6 and 7) indicate the position of the corresponding GST or GST–Smad2 fusion proteins.
Phosphorylated Smad2 co-localises with MEF2A in the nucleus of C2C12 mt
It is likely that, as in other cellular systems, TGF-β receptors are activated by TGF-β and are able to phosphorylate both Smad2 and 3, allowing them to bind to Smad4 and migrate to the nucleus. In contrast, Smad1 is not TGF-β sensitive. To determine the cellular localisation of Smad proteins at different stages of muscle development, cells were immunostained with isoform-specific antibodies. Endogenous MEF2A, Smad1, phosphorylated Smad2 and 3 were detected in mt in the presence of exogenous TGF-β (Fig. 5). MEF2A is strongly expressed in the nuclei of mt, and the addition of TGF-β does not alter either localisation or expression levels of MEF2A. In contrast, Smad1 remained cytoplasmic in both the absence and presence of TGF-β. Interestingly, however, we observed differential regulation of Smad2 and 3 in differentiating mt. Without the addition of exogenous TGF-β, phosphorylated Smad2 translocated to the nucleus in differentiating mt. Conversely, Smad3 remained cytoplasmic in differentiating mt, but did localise to the nucleus when cells were treated with TGF-β. These data suggest that Smad2–MEF2A cooperativity may exist in later stages of muscle development. These data also suggest that Smad2 and 3 are differentially regulated during muscle cell differentiation.
Figure 5.
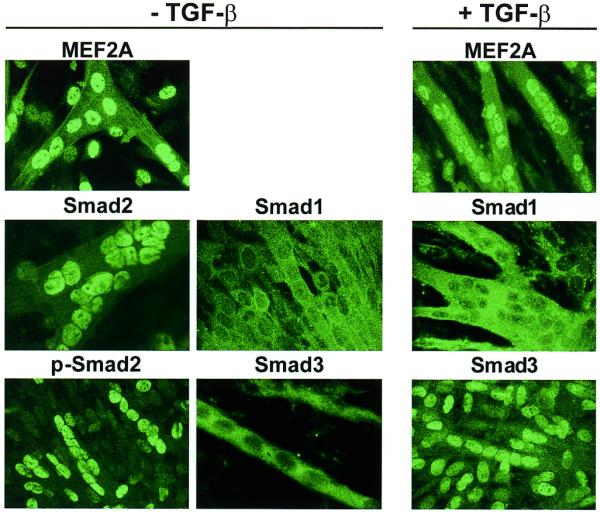
Localisation of MEF2A, Smad1, Smad2, phosphorylated-Smad2 and Smad3 in cultured muscle cells. C2C12 mb were grown in differentiation media in the absence of exogenous TGF-β (–TGF-β) until myotubes formed after 4 days (left and middle panels). At this point, myotubes were either fixed for immunofluorescence or exposed to differentiation medium + 2 ng/ml TGF-β (+TGF-β) for 24 h (right panels). Mt were fixed and immunostained using specific antibodies raised against MEF2A, Smad1, Smad2, phospho-Smad2 or Smad3.
MEF2A is not retained in the cytoplasm by TGF-β signalling
In previous studies by De Angelis et al., TGF-β was found to induce the re-localisation of MEF2C to the cytoplasm of myogenic cells (86). To determine whether MEF2A localisation is dependent on TGF-β signalling through Smad2, C2C12 mb were co-transfected with green fluorescent protein (GFP) and a dominant negative Smad2, MADR2(3SA) (Fig. 6). The phosphorylation site of MADR2(3SA) has three serines substituted with alanines and is known to be able to bind to TGF-β receptors, but cannot be phosphorylated (30). After 2 days in growth medium containing 1 ng/ml TGF-β, it was found that MEF2A is nuclear, while the dominant negative Smad2 is cytoplasmic. This suggests that the MEF2–Smad interaction only occurs when the Smad2 is targeted to the nucleus.
Figure 6.
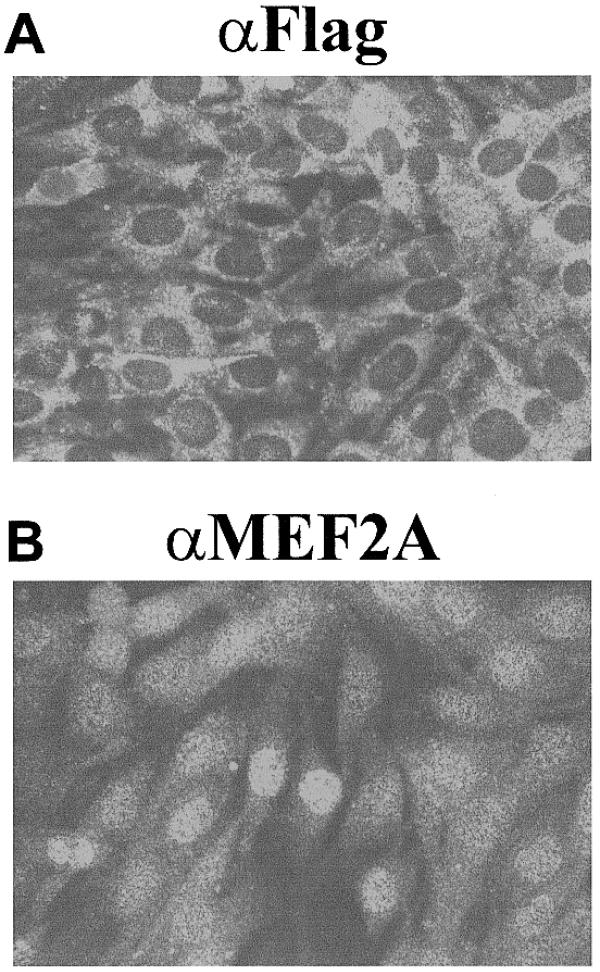
MEF2A is not retained in the cytoplasm by TGF-β signalling. (A and B) C2C12 mb were co-transfected with Flag–pCMVB–MADR2(3SA) and cytomegalovirus–eGFP using lipofectamine. The expression of GFP was used to determine which cells had been transfected. Cells were fixed with methanol and immunostained using either an anti-Flag (αFlag) antibody or an anti-MEF2A antibody (αMEF2A).
The p38 MAPK phosphorylation sites (Ser387, Thr293, Thr300) of MEF2C are necessary for Smad2/4 cooperativity
There has been evidence to suggest that there is cross-talk between TGF-β and p38 MAPK signalling (51). In addition, it is known that p38 MAPK phosphorylation of MEF2A and MEF2C TADs modulates MEF2 activity (73,74,76). Therefore, we tested whether MEF2 mutants that are not phosphorylated by p38 MAPK can still cooperate with the Smads (Fig. 7B). COS cells were transfected with expression vectors for a constitutively-active TGF-β receptor TβRI(T204D), Smad2, 3, 4, GAL4–MEF2C, GAL4–MEF2C(S387A) and GAL4–MEF2C(T293,300A). The MEF2C mutations constructed by Han et al., GAL4–MEF2C(S387A) and GAL4–MEF2C(T293,300A), are known to bind DNA and activate transcription of GAL4 reporter genes to the same degree as wild-type GAL4–MEF2C when co-transfected with p38 MAPK (and constitutively active MKK3) and ERK5/BMK1 (and constitutively active MEK5), respectively (74,87). It was found that enhancement of GAL4–MEF2C activity by Smad2/4 only occurred if Ser387, Thr293 and Thr300 are intact, indicating that phosphorylation of MEF2C by either p38 MAPK or ERK5/BMK1 is necessary for Smad–MEF2 cooperativity.
Figure 7.

The Smad2–MEF2 interaction is dependent on p38 MAPK phosphorylation sites in MEF2C and in mt, the endogenous MEF2 activity is increased by the addition of TGF-β. (A) COS cells were co-transfected with pSV-βgal, pCMV5B–Smad2, pCMV5B–Smad3, pCMV5B–Smad4, pCMV5B–TβRI(T204D), GAL4(DBD), GAL4–MEF2C(87–442), GAL4–MEF2C(S387A), GAL4–MEF2C(T293,300A) and 5×GAL4-luc as indicated. The construct GAL4–MEF2C(S387A) contains a replacement of serine 387 for alanine and GAL4–MEF2C(T293,300A) contains the exchange of two threonines for alanines at positions 293 and 300. Cells were harvested 72 h after transfection for luciferase assays. (B and C) C2C12 cells were co-transfected with pSV-βgal, pMT2, pMT2-MEF2A, pMEF2-luc, GAL4–MEF2A, GAL4–MEF2C, GAL4(DBD) and 5×GAL4-luc as indicated. Twenty-four hours later the medium was replaced with growth medium. One day later, growth medium was replaced with differentiation medium, which was replaced every 2 days. Mt formed after 4 days, at which point the medium was replaced with differentiation medium and either 2 ng/ml TGF-β (+) or an equal volume of TGF-β diluent (–). Cells were harvested after 24 h for luciferase assays. (A–C) β-galactosidase activities were used to normalise for transfection efficiency. Two or more sets of experiments were performed with comparable results. Each data point is a mean of triplicate samples from single experiments and the error bars represent the SEM.
DISCUSSION
In this report, we document a novel interaction between the MEF2 transcription factors and the Smad downstream effectors of the TGF-β signalling pathway. The MEF2–Smad interaction may confer a unique TGF-β-regulated sub-programme of MEF2-dependent gene expression. Since TGF-β superfamily members are potent regulators of mesodermally-derived cells, this interaction could subserve a variety of subcellular processes. In addition, these data indicate that the MAPK and TGF-β signalling pathways intersect at the level of the MEF2 proteins, suggesting the possibility of cross-regulation between the two signalling pathways. Emerging recognition of the association of the Smads with different transcriptional co-factors, such as MEF2, could, in part, underlie the high degree of divergence in TGF-β-activated programmes of gene expression. Moreover, this could also explain why the responses to TGF-β are highly dependent on cellular context, in that the host cell-specific complement of Smad interacting transcriptional regulators will largely determine the outcome of TGF-β signalling pathway activation. Our observations suggest that Smads and MEF2 function in concert in response to TGF-β signalling but, in addition, both have other binding partners, which presumably have discrete functions. Therefore, there may be considerable complexity in determining competition between heterologous binding partners. Consistent with this view, our studies indicate that only a fraction of the MEF2 pool is complexed with the Smads.
Integration of TGF-β signalling with MAP kinase signalling pathways
Our observation that mutation of the MEF2 phospho-acceptor sites on MEF2C renders it incapable of cooperating with the Smads is intriguing. In the case of SH2 domain interactions with phosphotyrosines on receptors or cytoplasmic proteins, SH2 domains bind to target phosphopeptides with high affinity (Kd = 10–100 nm) whereas they have virtually no affinity for the unphosphorylated peptide motif. Thus, the majority of the binding energy for the interaction is derived from the tyrosine phosphorylated peptides. In the case of the Smad–MEF2 interaction, based on our current data, our hypothesis is that the Smads interact with the serine/threonine phosphorylated C-terminus of MEF2 but have no affinity for the unphosphorylated form. Thus, the phosphorylation of MEF2 by p38 MAP kinase and ERK5/BMK1 may precede or stabilise the Smad–MEF2 interaction. This model infers that signalling to MEF2 requires phosphorylation and translocation of the Smads and also MAP kinase catalysed phosphorylation of the MEF2 transactivation domain.
There is already evidence that the Ras- and the Smad-mediated pathways can interact at different levels, indicating the possibility of reciprocal regulation between ligand–receptor complexes activating these pathways (1). In support of this idea, it has been shown that the TGF-β-associated kinase (TAK1) can also activate the MKK3–p38 MAP kinase pathway (50,51). In addition, Smad3 has been shown to be inactivated through phosphorylation in the proline-rich linker domain by epidermal growth factor stimulation via the Erk MAP kinases (88). Of note is a recent report that ATF-2, a p38 MAP kinase target, has also been shown to require signalling by p38 MAPK to be TGF-β responsive (50,51).
Role for TGF-β family members in the genetic hierarchy controlling MEF2
Perhaps the clearest view of the genetic role of the MEF2 factors is in Drosophila, where homozygous null mutations in the single D-MEF2 gene result in a complete abrogation of the differentiation programme for cardiac, visceral and skeletal muscle (89,90). In the genetic hierarchy leading to D-MEF2 expression, the tinman gene is a key regulator of D-MEF2 expression and its deletion extinguishes D-MEF2 expression (91). Interestingly, the tinman gene is enhanced by the Drosophila TGF-β family member decapentaplegic (DPP). Therefore, it is possible that DPP signalling could initiate the cascade leading to D-MEF2 expression while also activating transcriptional co-regulators in the form of the MAD genes. A vertebrate homologue of tinman exists, NKx2.5, which is involved in the differentiation programme of cardiomyocytes (92,93). Our data suggesting a direct physical association between the MEF2 and Smad proteins, and also the genetic evidence suggesting the importance of MEF2 and DPP in embryonic muscle specification, raise several important issues. It will be interesting to determine if DPP signalling in Drosophila also activates MAD co-regulators for D-MEF2 function. Moreover, based on the role of DPP in Drosophila, there may be a TGF-β family member that activates NKx2.5 expression in vertebrates and this could lead to the expression and modulation of MEF2 activity. Clearly, elucidation of these questions may lead to important observations concerning the role of TGF-β signalling in muscle gene regulation.
Regulation of muscle gene expression by TGF-β signalling
Skeletal muscle cells are known to be highly responsive to TGF-β signalling. In these studies we show that the endogenous MEF2 proteins are co-localised with the Smad proteins in the nuclei of differentiated mt. These data suggest a possible role for TGF-β signalling in the control of gene expression in differentiated muscle cells. Conversely, in mb, prior to the initiation of differentiation, the Smads are cytoplasmic and MEF2 proteins are not expressed at high levels (Z.A.Quinn, unpublished data). It is known that if mb are stimulated with exogenous TGF-β this inhibits the differentiation programme, which leads to the nuclear accumulation of Smads (present study). The mechanism of mb inhibition seems to be mediated by transcriptional down-regulation of the MyoD gene and constitutive MyoD expression can rescue myogenesis in the face of TGF-β signalling (Z.A.Quinn, unpublished data). Taken together, these observations suggest that once the differentiation programme is initiated and MEF2, functioning as a Smad co-regulator, is expressed, the inhibitory effect of TGF-β is lost and the association of the Smads with the MEF2 proteins promotes rather than inhibits the differentiation programme. One property of the MEF2–Smad interaction is that the Smads and MEF2 are spatially separated binding partners in which juxtaposition by regulated re-location allows the interaction to occur; the cooperative association thus being formed between the nuclear localised Smads and the phosphorylated MEF2 protein. A recent study has provided evidence that the subcellular localisation of MEF2C can be modified by TGF-β signalling and that a possible cytoplasmic tethering factor might be involved (86). In our studies we have no evidence to suggest that MEF2A is regulated in this manner. First, TGF-β treatment does not change the nuclear localisation of MEF2A and second, a Smad2 dominant negative that is permanently retained in the cytoplasm does not change this pattern. Therefore, the expression of MEF2 proteins may be a key switch in converting Smad-dependent TGF-β signalling in muscle cells from an inhibitory to a stimulatory differentiation signal.
In summary, investigation of transcriptional activation by the Smads has revealed a high degree of complexity involving multiple proteins and diverse promoter elements. In this report we identify a novel interaction between the Smad proteins and the MEF2 transcriptional regulatory proteins that could mediate transcriptional responses to TGF-β in a variety of cell types.
Acknowledgments
ACKNOWLEDGEMENT
This work was supported by a grant from The Medical Research Council of Canada to J.C.M.
References
- 1.Massague J. and Chen,Y.-G. (2000) Controlling TGF-beta signaling. Genes Dev., 14, 627–644. [PubMed] [Google Scholar]
- 2.Reddi A.H. (1997) BMPs: actions in flesh and bone. Nature Med., 3, 837–839. [DOI] [PubMed] [Google Scholar]
- 3.Zhang Y. and Derynck,R. (1999) Regulation of Smad signalling by protein associations and signalling crosstalk. Trends Cell Biol., 9, 274–279. [DOI] [PubMed] [Google Scholar]
- 4.ten Dijke P. and Heldin,C.H. (1999) Signal transduction. An anchor for activation [news]. Nature, 397, 109, 111. [DOI] [PubMed] [Google Scholar]
- 5.Wrana J.L. (2000) Regulation of Smad activity. Cell, 100, 189–192. [DOI] [PubMed] [Google Scholar]
- 6.Hill C.S. (1996) Signalling to the nucleus by members of the transforming growth factor-beta (TGF-beta) superfamily. Cell Signal ., 8, 533–544. [DOI] [PubMed] [Google Scholar]
- 7.Massague J. (1996) TGF-beta Signaling: Receptors, Transducers and Mad Proteins. Cell, 85, 947–950. [DOI] [PubMed] [Google Scholar]
- 8.Padgett R.W., Savage,C. and Das,P. (1997) Genetic and Biochemical Analysis of TGF-beta Signal Transduction. Cytokine Growth Factor Rev., 8, 1–9. [DOI] [PubMed] [Google Scholar]
- 9.Kingsley D.M. (1994) The TGF-beta superfamily: new members, new receptors and new genetic test of function in different organisms. Genes Dev., 8, 133–146. [DOI] [PubMed] [Google Scholar]
- 10.Wrana J.L., Attisano,L., Carcamo,J., Zentella,A., Doody,J., Laiho,M., Wang,X.F. and Massague,J. (1992) TGF beta signals through a heteromeric protein kinase receptor complex. Cell, 71, 1003–1014. [DOI] [PubMed] [Google Scholar]
- 11.Attisano L., Carcamo,J., Ventura,F., Weis,F.M., Massague,J. and Wrana,J.L. (1993) Identification of human activin and TGF beta type I receptors that form heteromeric kinase complexes with type II receptors. Cell, 75, 671–680. [DOI] [PubMed] [Google Scholar]
- 12.Ebner R., Chen,R.H., Lawler,S., Zioncheck,T. and Derynck,R. (1993) Determination of type I receptor specificity by the type II receptors for TGF-beta or activin. Science, 262, 900–902. [DOI] [PubMed] [Google Scholar]
- 13.Ebner R., Chen,R.H., Shum,L., Lawler,S., Zioncheck,T.F., Lee,A., Lopez,A.R. and Derynck,R. (1993) Cloning of a type I TGF-beta receptor and its effect on TGF-beta binding to the type II receptor. Science, 260, 1344–1348. [DOI] [PubMed] [Google Scholar]
- 14.Wieser R., Wrana,J.L. and Massague,J. (1995) GS domain mutations that constitutively activate T beta R-I, the downstream signaling component in the TGF-beta receptor complex. EMBO J., 14, 2199–2208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wrana J.L., Attisano,L., Wieser,R., Ventura,F. and Massague,J. (1994) Mechanism of activation of the TGF-beta receptor. Nature, 370, 341–347. [DOI] [PubMed] [Google Scholar]
- 16.Wrana J.L. and Attisano,L. (2000) The smad pathway [In Process Citation]. Cytokine Growth Factor Rev., 11, 5–13. [DOI] [PubMed] [Google Scholar]
- 17.Massague J. and Wotton,D. (2000) Transcriptional control by the TGF-beta/Smad signaling system. EMBO J., 19, 1745–1754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Raftery L.A., Twombly,V., Wharton,K. and Gelbart,W.M. (1995) Genetic screens to identify elements of the decapentaplegic signaling pathway in Drosophila. Genetics, 139, 241–254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sekelsky J.J., Newfeld,S.J., Raftery,L.A., Chartoff,E.H. and Gelbart,W.M. (1995) Genetic characterization and cloning of mothers against dpp, a gene required for decapentaplegic function in Drosophila melanogaster. Genetics, 139, 1347–1358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liu F., Hata,A., Baker,J.C., Doody,J., Carcamo,J., Harland,R.M. and Massague,J. (1996) A human Mad protein acting as a BMP-regulated transcriptional activator [see comments]. Nature, 381, 620–623. [DOI] [PubMed] [Google Scholar]
- 21.Savage C., Das,P., Finelli,A.L., Townsend,S.R., Sun,C.Y., Baird,S.E. and Padgett,R.W. (1996) Caenorhabditis elegans genes sma-2, sma-3 and sma-4 define a conserved family of transforming growth factor beta pathway components. Proc. Natl Acad. Sci. USA, 93, 790–794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lagna G., Hata,A., Hemmati-Brivanlou,A. and Massague,J. (1996) Partnership between DPC4 and SMAD proteins in TGF-beta signalling pathways. Nature, 383, 832–836. [DOI] [PubMed] [Google Scholar]
- 23.Zhang Y., Musci,T. and Derynck,R. (1997) The tumor suppressor Smad4/DPC 4 as a central mediator of Smad function. Curr. Biol., 7, 270–276. [DOI] [PubMed] [Google Scholar]
- 24.Heldin C.H., Miyazono,K. and ten Dijke,P. (1997) TGF-beta signalling from cell membrane to nucleus through SMAD proteins. Nature, 390, 465–471. [DOI] [PubMed] [Google Scholar]
- 25.Attisano L. and Wrana,J.L. (1998) Mads and Smads in TGF beta signalling. Curr. Opin. Cell Biol., 10, 188–194. [DOI] [PubMed] [Google Scholar]
- 26.Kretzschmar M. and Massague,J. (1998) SMADs: mediators and regulators of TGF-beta signaling. Curr. Opin. Genet. Dev., 8, 103–111. [DOI] [PubMed] [Google Scholar]
- 27.Hoodless P.A., Haerry,T., Abdollah,S., Stapleton,M., O’Connor,M.B., Attisano,L. and Wrana,J.L. (1996) MADR1, a MAD-related protein that functions in BMP2 signaling pathways. Cell, 85, 489–500. [DOI] [PubMed] [Google Scholar]
- 28.Carcamo J., Weis,F.M., Ventura,F., Wieser,R., Wrana,J.L., Attisano,L. and Massague,J. (1994) Type I receptors specify growth-inhibitory and transcriptional responses to transforming growth factor beta and activin. Mol. Cell. Biol., 14, 3810–3821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Liu X., Sun,Y., Constantinescu,S.N., Karam,E., Weinberg,R.A. and Lodish,H.F. (1997) Transforming growth factor beta-induced phosphorylation of Smad3 is required for growth inhibition and transcriptional induction in epithelial cells. Proc. Natl Acad. Sci. USA, 94, 10669–10674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Macias-Silva M., Abdollah,S., Hoodless,P.A., Pirone,R., Attisano,L. and Wrana,J.L. (1996) MADR2 is a substrate of the TGFbeta receptor and its phosphorylation is required for nuclear accumulation and signaling. Cell, 87, 1215–1224. [DOI] [PubMed] [Google Scholar]
- 31.Tsukazaki T., Chiang,T.A., Davison,A.F., Attisano,L. and Wrana,J.L. (1998) SARA, a FYVE domain protein that recruits Smad2 to the TGFbeta receptor. Cell, 95, 779–791. [DOI] [PubMed] [Google Scholar]
- 32.Kim J., Johnson,K., Chen,H.J., Carroll,S. and Laughon,A. (1997) Drosophila Mad binds to DNA and directly mediates activation of vestigial by Decapentaplegic. Nature, 388, 304–308. [DOI] [PubMed] [Google Scholar]
- 33.Shi Y., Wang,Y.F., Jayaraman,L., Yang,H., Massague,J. and Pavletich,N.P. (1998) Crystal structure of a Smad MH1 domain bound to DNA: insights on DNA binding in TGF-beta signaling. Cell, 94, 585–594. [DOI] [PubMed] [Google Scholar]
- 34.Qin B., Lam,S.S. and Lin,K. (1999) Crystal structure of a transcriptionally active Smad4 fragment. Structure Fold Des., 7, 1493–1503. [DOI] [PubMed] [Google Scholar]
- 35.Johnson K., Kirkpatrick,H., Comer,A., Hoffmann,F.M. and Laughon,A. (1999) Interaction of Smad complexes with tripartite DNA-binding sites. J. Biol. Chem., 274, 20709–20716. [DOI] [PubMed] [Google Scholar]
- 36.Dennler S., Huet,S. and Gauthier,J.M. (1999) A short amino-acid sequence in MH1 domain is responsible for functional differences between Smad2 and Smad3. Oncogene, 18, 1643–1648. [DOI] [PubMed] [Google Scholar]
- 37.Watabe T., Kim,S., Candia,A., Rothbacher,U., Hashimoto,C., Inoue,K. and Cho,K.W. (1995) Molecular mechanisms of Spemann’s organizer formation: conserved growth factor synergy between Xenopus and mouse. Genes Dev., 9, 3038–3050. [DOI] [PubMed] [Google Scholar]
- 38.Labbe E., Silvestri,C., Hoodless,P.A., Wrana,J.L. and Attisano,L. (1998) Smad2 and Smad3 positively and negatively regulate TGF beta-dependent transcription through the forkhead DNA-binding protein FAST2. Mol. Cell, 2, 109–120. [DOI] [PubMed] [Google Scholar]
- 39.Germain S., Howell,M., Esslemont,G.M. and Hill,C.S. (2000) Homeodomain and winged-helix transcription factors recruit activated Smads to distinct promoter elements via a common Smad interaction motif. Genes Dev., 14, 435–451. [PMC free article] [PubMed] [Google Scholar]
- 40.Chen Y., Lebrun,J.J. and Vale,W. (1996) Regulation of transforming growth factor beta- and activin-induced transcription by mammalian Mad proteins. Proc. Natl Acad. Sci. USA, 93, 12992–12997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhang Y., Feng,X., We,R. and Derynck,R. (1996) Receptor-associated Mad homologues synergize as effectors of the TGF-beta response. Nature, 383, 168–172. [DOI] [PubMed] [Google Scholar]
- 42.Nakao A., Imamura,T., Souchelnytskyi,S., Kawabata,M., Ishisaki,A., Oeda,E., Tamaki,K., Hanai,J., Heldin,C.H., Miyazono,K. and ten Dijke,P. (1997) TGF-beta receptor-mediated signalling through Smad2, Smad3 and Smad4. EMBO J., 16, 5353–5362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhang Y., Feng,X.H. and Derynck,R. (1998) Smad3 and Smad4 cooperate with c-Jun/c-Fos to mediate TGF-beta-induced transcription. Nature, 394, 909–913. [DOI] [PubMed] [Google Scholar]
- 44.Song C.Z., Tian,X. and Gelehrter,T.D. (1999) Glucocorticoid receptor inhibits transforming growth factor-beta signaling by directly targeting the transcriptional activation function of Smad3. Proc. Natl Acad. Sci. USA, 96, 11776–11781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yahata T., de Caestecker,M.P., Lechleider,R.J., Andriole,S., Roberts,A.B., Isselbacher,K.J. and Shioda,T. (2000) The MSG1 non-DNA-binding transactivator binds to the p300/CBP coactivators, enhancing their functional link to the Smad transcription factors. J. Biol. Chem., 275, 8825–8834. [DOI] [PubMed] [Google Scholar]
- 46.Akiyoshi S., Inoue,H., Hanai,J., Kusanagi,K., Nemoto,N., Miyazono,K. and Kawabata,M. (1999) c-Ski acts as a transcriptional co-repressor in transforming growth factor-beta signaling through interaction with smads. J. Biol. Chem., 274, 35269–35277. [DOI] [PubMed] [Google Scholar]
- 47.Shen X., Hu,P.P., Liberati,N.T., Datto,M.B., Frederick,J.P. and Wang,X.F. (1998) TGF-beta-induced phosphorylation of Smad3 regulates its interaction with coactivator p300/CREB-binding protein. Mol. Biol. Cell, 9, 3309–3319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nishihara A., Hanai,J.I., Okamoto,N., Yanagisawa,J., Kato,S., Miyazono,K. and Kawabata,M. (1998) Role of p300, a transcriptional coactivator, in signalling of TGF-beta. Genes Cells, 3, 613–623. [DOI] [PubMed] [Google Scholar]
- 49.Janknecht R., Wells,N.J. and Hunter,T. (1998) TGF-beta-stimulated cooperation of smad proteins with the coactivators CBP/p300. Genes Dev., 12, 2114–2119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sano Y., Harada,J., Tashiro,S., Gotoh-Mandeville,R., Maekawa,T. and Ishii,S. (1999) ATF-2 is a common nuclear target of Smad and TAK1 pathways in transforming growth factor-beta signaling. J. Biol. Chem., 274, 8949–8957. [DOI] [PubMed] [Google Scholar]
- 51.Hanafusa H., Ninomiya-Tsuji,J., Masuyama,N., Nishita,M., Fujisawa,J., Shibuya,H., Matsumoto,K. and Nishida,E. (1999) Involvement of the p38 mitogen-activated protein kinase pathway in transforming growth factor-beta-induced gene expression. J. Biol. Chem., 274, 27161–27167. [DOI] [PubMed] [Google Scholar]
- 52.Zhang Y. and Derynck,R. (2000) Transcriptional regulation of the TGF-β-inducible mouse germ-line Ig α constant region gene by functional cooperation of Smad, CREB and AML family members. J. Biol. Chem., 275, 16979–16985. [DOI] [PubMed] [Google Scholar]
- 53.Chen Y.G., Hata,A., Lo,R.S., Wotton,D., Shi,Y., Pavletich,N. and Massague,J. (1998) Determinants of specificity in TGF-beta signal transduction. Genes Dev., 12, 2144–2152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Bassel-Duby R., Hernandez,M.D., Gonzalez,M.A., Krueger,J.K. and Williams,R.S. (1992) A 40-kilodalton protein binds specifically to an upstream sequence element essential for muscle-specific transcription of the human myoglobin promoter. Mol. Cell. Biol., 12, 5024–5032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Leifer D., Krainc,D., Yu,Y.T., McDermott,J., Breitbart,R.E., Heng,J., Neve,R.L., Kosofsky,B., Nadal-Ginard,B. and Lipton,S.A. (1993) MEF2C, a MADS/MEF2-family transcription factor expressed in a laminar distribution in cerebral cortex. Proc. Natl Acad. Sci. USA, 90, 1546–1550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Yu Y.T., Breitbart,R.E., Smoot,L.B., Lee,Y., Mahdavi,V. and Nadal-Ginard,B. (1992) Human myocyte-specific enhancer factor 2 comprises a group of tissue-restricted MADS box transcription factors. Genes Dev., 6, 1783–1798. [DOI] [PubMed] [Google Scholar]
- 57.Breitbart R.E., Liang,C.S., Smoot,L.B., Laheru,D.A., Mahdavi,V. and Nadal-Ginard,B. (1993) A fourth human MEF2 transcription factor, hMEF2D, is an early marker of the myogenic lineage. Development, 118, 1095–1106. [DOI] [PubMed] [Google Scholar]
- 58.Edmondson D.G., Cheng,T.C., Cserjesi,P., Chakraborty,T. and Olson,E.N. (1992) Analysis of the myogenin promoter reveals an indirect pathway for positive autoregulation mediated by the muscle-specific enhancer factor MEF-2. Mol. Cell. Biol., 12, 3665–3677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Black B.L. and Olson,E.N. (1998) Transcriptional control of muscle development by myocyte enhancer factor-2 (MEF2) proteins. Annu. Rev. Cell. Dev. Biol., 14, 167–196. [DOI] [PubMed] [Google Scholar]
- 60.Olson E.N. and Klein,W.H. (1998) Muscle minus myoD. Dev. Biol., 202, 153–156. [DOI] [PubMed] [Google Scholar]
- 61.Ross R.S., Navankasattusas,S., Harvey,R.P. and Chien,K.R. (1996) An HF-1a/HF-1b/MEF-2 combinatorial element confers cardiac ventricular specificity and established an anterior-posterior gradient of expression. Development, 122, 1799–1809. [DOI] [PubMed] [Google Scholar]
- 62.Lee Y., Nadal-Ginard,B., Mahdavi,V. and Izumo,S. (1997) Myocyte-specific enhancer factor 2 and thyroid hormone receptor associate and synergistically activate the alpha-cardiac myosin heavy-chain gene. Mol. Cell. Biol., 17, 2745–2755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kuisk I.R., Li,H., Tran,D. and Capetanaki,Y. (1996) A single MEF2 site governs desmin transcription in both heart and skeletal muscle during mouse embryogenesis. Dev. Biol., 174, 1–13. [DOI] [PubMed] [Google Scholar]
- 64.Chambers A.E., Logan,M., Kotecha,S., Towers,N., Sparrow,D. and Mohun,T.J. (1994) The RSRF/MEF2 protein SL1 regulates cardiac muscle-specific transcription of a myosin light-chain gene in Xenopus embryos. Genes Dev., 8, 1324–1334. [DOI] [PubMed] [Google Scholar]
- 65.Bruneau B.G., Bao,Z.Z., Tanaka,M., Schott,J.J., Izumo,S., Cepko,C.L., Seidman,J.G. and Seidman,C.E. (2000) Cardiac expression of the ventricle-specific homeobox gene Irx4 is modulated by Nkx2-5 and dHand. Dev. Biol., 217, 266–277. [DOI] [PubMed] [Google Scholar]
- 66.Bi W., Drake,C.J. and Schwarz,J.J. (1999) The transcription factor MEF2C-null mouse exhibits complex vascular malformations and reduced cardiac expression of angiopoietin 1 and VEGF. Dev. Biol., 211, 255–267. [DOI] [PubMed] [Google Scholar]
- 67.Lin Q., Schwarz,J., Bucana,C. and Olson,E.N. (1997) Control of mouse cardiac morphogenesis and myogenesis by transcription factor MEF2C. Science, 276, 1404–1407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Lyons G.E., Micales,B.K., Schwarz,J., Martin,J.F. and Olson,E.N. (1995) Expression of mef2 genes in the mouse central nervous system suggests a role in neuronal maturation. J. Neurosci., 15, 5727–5738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Edmondson D.G., Lyons,G.E., Martin,J.F. and Olson,E.N. (1994) Mef2 gene expression marks the cardiac and skeletal muscle lineages during mouse embryogenesis. Development, 120, 1251–1263. [DOI] [PubMed] [Google Scholar]
- 70.Swanson B.J., Jack,H.M. and Lyons,G.E. (1998) Characterization of myocyte enhancer factor 2 (MEF2) expression in B and T cells: MEF2C is a B cell-restricted transcription factor in lymphocytes. Mol. Immunol., 35, 445–458. [DOI] [PubMed] [Google Scholar]
- 71.Buchberger A. and Arnold,H.H. (1999) The MADS domain containing transcription factor cMef2a is expressed in heart and skeletal muscle during embryonic chick development. Dev. Genes Evol., 209, 376–381. [DOI] [PubMed] [Google Scholar]
- 72.Youn H.D., Sun,L., Prywes,R. and Liu,J.O. (1999) Apoptosis of T cells mediated by Ca2+-induced release of the transcription factor MEF2. Science, 286, 790–793. [DOI] [PubMed] [Google Scholar]
- 73.Zhao M., New,L., Kravchenko,V.V., Kato,Y., Gram,H., di Padova,F., Olson,E.N., Ulevitch,R.J. and Han,J. (1999) Regulation of the MEF2 family of transcription factors by p38. Mol. Cell. Biol., 19, 21–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Han J., Jiang,Y., Li,Z., Kravchenko,V.V. and Ulevitch,R.J. (1997) Activation of the transcription factor MEF2C by the MAP kinase p38 in inflammation. Nature, 386, 296–299. [DOI] [PubMed] [Google Scholar]
- 75.Yang C.C., Ornatsky,O.I., McDermott,J.C., Cruz,T.F. and Prody,C.A. (1998) Interaction of myocyte enhancer factor 2 (MEF2) with a mitogen-activated protein kinase, ERK5/BMK1. Nucleic Acids Res., 26, 4771–4777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Ornatsky O.I., Cox,D.M., Tangirala,P., Andreucci,J.J., Quinn,Z.A., Wrana,J.L., Prywes,R., Yu,Y.T. and McDermott,J.C. (1999) Post-translational control of the MEF2A transcriptional regulatory protein. Nucleic Acids Res., 27, 2646–2654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Ornatsky O.I., Andreucci,J.J. and McDermott,J.C. (1997) A dominant-negative form of transcription factor MEF2 inhibits myogenesis. J. Biol. Chem., 272, 33271–33278. [DOI] [PubMed] [Google Scholar]
- 78.Ornatsky O.I. and McDermott,J.C. (1996) MEF2 protein expression, DNA binding specificity and complex composition, and transcriptional activity in muscle and non-muscle cells. J. Biol. Chem., 271, 24927–24933. [DOI] [PubMed] [Google Scholar]
- 79.Attisano L., Wrana,J.L., Montalvo,E. and Massague,J. (1996) Activation of signalling by the activin receptor complex. Mol. Cell. Biol., 16, 1066–1073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wieser R., Attisano,L., Wrana,J.L. and Massague,J. (1993) Signaling activity of transforming growth factor beta type II receptors lacking specific domains in the cytoplasmic region. Mol. Cell. Biol., 13, 7239–7247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Hayashi H., Abdollah,S., Qiu,Y., Cai,J., Xu,Y.Y., Grinnell,B.W., Richardson,M.A., Topper,J.N., Gimbrone,M.A.J., Wrana,J.L. and Falb,D. (1997) The MAD-related protein Smad7 associates with the TGFbeta receptor and functions as an antagonist of TGFbeta signaling. Cell, 89, 1165–1173. [DOI] [PubMed] [Google Scholar]
- 82.McDermott J.C., Cardoso,M.C., Yu,Y.T., Andres,V., Leifer,D., Krainc,D., Lipton,S.A. and Nadal-Ginard,B. (1993) hMEF2C gene encodes skeletal muscle- and brain-specific transcription factors. Mol. Cell Biol., 13, 2564–2577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Abdollah S., Macias-Silva,M., Tsukazaki,T., Hayashi,H., Attisano,L. and Wrana,J.L. (1997) TbetaRI phosphorylation of Smad2 on Ser465 and Ser467 is required for Smad2-Smad4 complex formation and signaling. J. Biol. Chem., 272, 27678–27685. [DOI] [PubMed] [Google Scholar]
- 84.Eppert K., Scherer,S.W., Ozcelik,H., Pirone,R., Hoodless,P., Kim,H., Tsui,L.C., Bapat,B., Gallinger,S., Andrulis,I.L., Thomsen,G.H., Wrana,J.L. and Attisano,L. (1996) MADR2 maps to 18q21 and encodes a TGFbeta-regulated MAD-related protein that is functionally mutated in colorectal carcinoma. Cell, 86, 543–552. [DOI] [PubMed] [Google Scholar]
- 85.Yamamoto N., Akiyama,S., Katagiri,T., Namiki,M., Kurokawa,T. and Suda,T. (1997) Smad1 and smad5 act downstream of intracellular signalings of BMP-2 that inhibits myogenic differentiation and induces osteoblast differentiation in C2C12 myoblasts. Biochem. Biophys. Res. Commun., 238, 574–580. [DOI] [PubMed] [Google Scholar]
- 86.De Angelis L., Borghi,S., Melchionna,R., Berghella,L., Baccarani-Contri,M., Parise,F., Ferrari,S. and Cossu,G. (1998) Inhibition of myogenesis by transforming growth factor beta is density-dependent and related to the translocation of transcription factor MEF2 to the cytoplasm. Proc. Natl Acad. Sci. USA, 95, 12358–12363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kato Y., Kravchenko,V., Tapping,R., Han,J., Ulevitch,R. and Lee,J. (1997) BMK1/ERK5 regulates serum-induced early gene expression through transcription factor MEF2C. EMBO J., 16, 7054–7066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Kretzschmar M., Doody,J. and Massague,J. (1997) Opposing BMP and EGF signalling pathways converge on the TGF-beta family mediator Smad1. Nature, 389, 618–622. [DOI] [PubMed] [Google Scholar]
- 89.Ranganayakulu G., Zhao,B., Dokidis,A., Molkentin,J.D., Olson,E.N. and Schulz,R.A. (1995) A series of mutations in the D-MEF2 transcription factor reveal multiple functions in larval and adult myogenesis in Drosophila. Dev. Biol., 171, 169–181. [DOI] [PubMed] [Google Scholar]
- 90.Bour B.A., O’Brien,M.A., Lockwood,W.L., Goldstein,E.S., Bodmer,R., Taghert,P.H., Abmayr,S.M. and Nguyen,H.T. (1995) Drosophila MEF2, a transcription factor that is essential for myogenesis. Genes Dev., 9, 730–741. [DOI] [PubMed] [Google Scholar]
- 91.Gajewski K., Kim,Y., Lee,Y.M., Olson,E.N. and Schulz,R.A. (1997) D-mef2 is a target for Tinman activation during Drosophila heart development. EMBO J., 16, 515–522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Gajewski K., Kim,Y., Choi,C.Y. and Schulz,R.A. (1998) Combinatorial control of Drosophila mef2 gene expression in cardiac and somatic muscle cell lineages. Dev. Genes Evol., 208, 382–392. [DOI] [PubMed] [Google Scholar]
- 93.Cripps R.M., Zhao,B. and Olson,E.N. (1999) Transcription of the myogenic regulatory gene Mef2 in cardiac, somatic and visceral muscle cell lineages is regulated by a Tinman-dependent core enhancer. Dev. Biol., 215, 420–430. [DOI] [PubMed] [Google Scholar]