

Abstract
The switch from producing vegetative structures (branches and leaves) to producing reproductive structures (flowers) is a crucial developmental transition that significantly affects the reproductive success of flowering plants. In Arabidopsis, this transition is in large part controlled by the meristem identity regulator LEAFY (LFY). The molecular mechanisms by which LFY orchestrates a precise and robust switch to flower formation is not well understood. Here, we show that the direct LFY target LATE MERISTEM IDENTITY2 (LMI2) has a role in the meristem identity transition. Like LFY, LMI2 activates AP1 directly; moreover, LMI2 and LFY interact physically. LFY, LMI2 and AP1 are connected in a feed-forward and positive feedback loop network. We propose that these intricate regulatory interactions not only direct the precision of this crucial developmental transition in rapidly changing environmental conditions, but also contribute to its robustness and irreversibility.
Keywords: APETALA1, LEAFY, LMI2, Meristem identity transition, Reproductive development, Arabidopsis
INTRODUCTION
Flowering plants transition through a number of distinct developmental phases in their lifecycle. During each phase, different organs are generated from a group of cells at the flanks of the shoot apical meristem, which form the primordia (Steeves and Sussex, 1989). Developmental phase transitions have been studied extensively in the plant model system Arabidopsis thaliana (Araki, 2001; Blazquez et al., 2006; Poethig, 2003; Steeves and Sussex, 1989). During the vegetative phase, the primordia give rise to a series of leaves, which form the basal rosette. During the early reproductive phase, the shoot apical meristem grows upward (bolts) and the newly formed primordia develop into secondary inflorescence branches subtended by cauline leaves. Finally, after the meristem identity transition, the first flowers are formed.
The precise timing of flower formation is crucial for reproductive fitness, as plants must ensure that the energy and resources accumulated during the vegetative phase are optimally allocated to the production of offspring (Roux et al., 2006). Plants rely on both environmental and endogenous cues to fine-tune the onset of reproductive development (Araki, 2001; Koornneef et al., 1998; Simpson et al., 1999). These signals modulate the level and activity of flowering-time regulators, which initiate the reproductive phase and induce expression of the meristem identity genes (Amasino, 2010; Baurle and Dean, 2006; Kobayashi and Weigel, 2007; Komeda, 2004; Turck et al., 2008). The meristem identity regulators then trigger formation of the first flower (Blazquez et al., 2006; Liu et al., 2009a; Parcy, 2005).
Two key meristem identity regulators in Arabidopsis are the plant-specific transcription factor LEAFY (LFY) and the MADS box transcription factor APETALA1 (AP1). LFY is considered to be a central meristem identity regulator because lfy null mutants cause a very dramatic delay in the meristem identity transition (Huala and Sussex, 1992; Weigel et al., 1992). Moreover, LFY upregulation in the initiating primordia flanking the shoot apical meristem is one of the first steps in the regulatory cascade that leads to the meristem identity transition (Blazquez et al., 1997; Hempel et al., 1997). LFY executes its meristem identity role in part by activating AP1 expression directly (Parcy et al., 1998; Wagner et al., 1999; William et al., 2004). AP1 upregulation marks commitment to flower formation (Blazquez et al., 1997; Bowman et al., 1993; Hempel et al., 1997; Liu et al., 2007; Mandel and Yanofsky, 1995; Yu et al., 2004). AP1 promotes floral fate by upregulating floral identity pathways and by repressing inflorescence identity pathways (Ferrandiz et al., 2000; Kaufmann et al., 2010; Liljegren et al., 1999; Liu et al., 2007; Yu et al., 2004). Two LFY-independent pathways can also upregulate AP1: one involves the photoperiod flowering-time regulators, FLOWERING LOCUS T (FT) and FD, and the other involves components of the age-sensing flowering-time pathway, the SBP transcription factors (Abe et al., 2005; Wang et al., 2009; Wigge et al., 2005; Yamaguchi et al., 2009). In agreement with this, simultaneous loss-of-function mutations in both LFY and AP1 results in plants that essentially lack flowers (Bowman et al., 1993; Huala and Sussex, 1992; Schultz and Haughn, 1993; Weigel et al., 1992).
Although the meristem identity transition is a key developmental switch, our understanding of the events that lead from LFY upregulation to flower formation is still incomplete. Previously, we used a genomic approach to define direct targets of LFY during the meristem identity transition (William et al., 2004). This approach identified the meristem identity regulators and direct LFY targets CAULIFLOWER (CAL), a close AP1 homolog, and LATE MERISTEM IDENTITY1 (LMI1), a class I HD-Zip transcription factor (Saddic et al., 2006; William et al., 2004). Another direct LFY target identified was AtMYB17 (William et al., 2004). AtMYB17 is a member of the R2R3 class of MYB transcription factors, which have important roles in many processes in plants, including cell fate specification, metabolism, and biotic and abiotic stress responses (Dubos et al., 2010; Kranz et al., 1998; Martin and Paz-Ares, 1997; Stracke et al., 2001). The Arabidopsis homologs of AtMYB17, AtMYB16 (MIXTA) and AtMYB106 (NOECK), have been reported to function in the determination of cell shape in the petal epidermis and in the repression of trichome branching (Baumann et al., 2007; Jakoby et al., 2008). The biological function of AtMYB17 is not understood. Here, we show a role for AtMYB17 in the meristem identity transition upstream of AP1; based on these findings, we renamed this gene LATE MERISTEM IDENTITY2 (LMI2).
MATERIALS AND METHODS
Plant lines, growth and LMI2 rescue construct
T-DNA insertion lines were obtained from the SALK collection (Alonso and Stepanova, 2003) and twice backcrossed to Columbia (wild type). lfy and ap1 alleles used were described previously (Saddic et al., 2006; Yamaguchi et al., 2009). lfy-2 and lfy-10 carry the same lesion (Schultz and Haughn, 1993; Weigel et al., 1992) and were used interchangeably. For all genotyping primers, see Table S2 in the supplementary material.
All plant growth was in inductive photoperiod. Seeds were stratified for seven days at 4°C and either grown in white fluorescent lights at 22°C in soil in long-day conditions (16 hours light, 8 hours dark; 110 μmol/m2s) for experiments involving phenotyping and inflorescences, or on plates (0.5× MS media) in long-day conditions for three days followed by growth in continuous light (90 μmol/m2s) for seedling experiments.
For genomic rescue, the LMI2 locus including 2150 bp upstream of the translational start site was PCR amplified, sequenced and Gateway cloned into pGWB1 (Nakagawa et al., 2007). The resulting construct was transformed into lmi2-2 lfy-10 plants. A representative pLMI2:LMI2 lmi2-2 lfy-10 transgenic line was characterized further. For all cloning primers see Table S3 in the supplementary material.
Semi-quantitative and quantitative PCR
Developmental age was determined based on number of days of growth and adjusted by developmental stage (emergence and size of true leaves) (Saddic et al., 2006). RNA was extracted from entire seedlings except for the study of LMI2 mis-expression in lmi2-1 mutants. RNA purification, reverse transcription and qRT-PCR were described previously (Yamaguchi et al., 2009). All real-time RT-PCR experiments were normalized over the ubiquitously expressed EIF4A gene (AT3G13920). The mean and s.e.m. were calculated for each biological replicate using three technical replicates. One representative experiment is shown. See Table S4 in the supplementary material for qRT-PCR primers used.
β-Glucuronidase (GUS) assays
Upstream and downstream intergenic regions (2150 bp upstream of the translation start site and 2699 bp downstream of the translation termination site) were PCR amplified, sequenced and cloned into pBI101 (Clontech, Mountain View, CA, USA). Wild-type plants (Col) were transformed and a representative transgenic line was characterized. To investigate the role of LFY on LMI2:GUS expression, LMI2:GUS was crossed to lfy-9, 35S:LFY-GR in Ler (Wagner et al., 1999), and Ler (wild type). GUS assays were performed as described by Saddic et al. and Yamaguchi et al. (Saddic et al., 2006; Yamaguchi et al., 2005) using seven-day-old seedlings or 1-2 cm bolted primary inflorescences. For transient induction assays, seven-day-old F1 seedlings (LMI2:GUS × Ler or LMI2:GUS × 35S:LFY-GR) were incubated overnight with 10 μM dexamethasone at room temperature as previously described (Wagner et al., 1999) prior to GUS staining. Whole-mount samples and histological sections were visualized using an Olympus SZX12 dissecting or an Olympus BX51 compound microscope.
The LMI2:GUS reporter showed ectopic expression in the L1 layer of stems, petioles and leaves not detected by LMI2 in situ hybridization analyses. This might be due to missing cis regulatory elements located in LMI2 introns (Liu et al., 2007; Oh et al., 2009; Sieburth and Meyerowitz, 1997).
In situ hybridization
For the LMI2 antisense and sense probes, the genic region downstream of the MYB DNA binding domain was used. The AP1 in situ probe contained the genic region downstream of the MADS box. The constructs were PCR amplified, cloned into pGEM T-easy (LMI2) and pGEM-T (AP1; Promega, Madison, WI, USA), and sequenced. Sense and antisense LMI2 probes were digested with SalI and transcribed with the T7 polymerase, whereas the antisense AP1 probe was transcribed using the T7 polymerase following digestion with EcoRI. The Riboprobe Combination System (Promega) and DIG RNA labeling mix (Roche, Branchburg, NJ, USA) were used for probe synthesis. In situ hybridization was performed as described by Long and Barton (Long and Barton, 1998).
Chromatin immunoprecipitation (ChIP)
The pLMI2:LMI2 rescue construct excluding the translation termination codon was Gateway cloned into pGWB13 (Nakagawa et al., 2007). pLMI2:LMI2-HA was transformed into lmi2-2 plants followed by testing for rescue. For ChIP, 300 mg tissue from eleven-day-old seedlings of a representative line were used with 3 μg/sample or 4 μg/sample of anti-HA antibody [sc-805 (Santa Cruz, Santa Cruz, CA, USA) or 12CA5 (Roche), respectively] using published procedures (Kwon et al., 2005; William et al., 2004). LMI2 occupancy on genomic DNA was calculated by computing the enrichment over the respective input and normalized over lmi2-2. The mean and s.e.m. were calculated using at least three technical replicates; one representative biological replicate is shown. For ChIP-qPCR primers see Table S5 in the supplementary material.
Glutathione-S-transferase (GST) Pull-down
The LFY coding region was amplified and inserted between the EcoRI and NotI sites into pGEX-5X-1 (GE Healthcare, Piscataway, NJ, USA). The fusion protein was expressed in Escherichia coli (AD494). After induction with 0.1 mM IPTG at 37°C for one hour, cells were harvested by centrifugation and resuspended in ice-cold PBS containing 1 mM EDTA, 1 mM PMSF, 1 mg/ml lysozyme and 1% Triton X-100. Following a 20 minute incubation at room temperature, the cell lysate was cleared by centrifugation. Protein extracts were incubated with Sepharose 4B slurry (GE Healthcare) at 4°C for one hour. The beads were washed five times with PBS containing 1 mM EDTA and 1 mM PMSF. The protein-bound beads were used directly for pull-down assays. In vitro transcription and translation of LFY, LMI2 and NCa (1-464 amino acid fragment of the chromatin remodeling ATPase SYD) (Wagner and Meyerowitz, 2002) and the pull-down assay were performed as previously described (Sang et al., 2005).
Yeast 2-hybrid
LMI2N consisted of the N-terminal protein coding region of LMI2, including the MYB domain and the subgroup 9 motif, whereas LMI2C contained the remainder protein coding region of LMI2. The LMI2 fragments were amplified and inserted between the SalI and NotI sites of pDBLeu (Invitrogen, Carlsbad, CA, USA). The coding region of LFY was amplified and Gateway cloned into pDEST22 (Invitrogen).
pDBLeu-LMI2N or LMI2C bait constructs were co-transformed into yeast (PJ69-4A) with either pDEST22-LFY or pDEST alone. After transformation, cells were plated on –Trp –Leu/SD media. Double transformants were grown in –Trp –Leu/SD liquid media overnight, adjusted for equal cell density, serially diluted (10–1-10–4) and spotted on –Trp –Leu –His/SD plates.
Bimolecular fluorescence complementation
LMI2N and LMI2C fragments were inserted into pENTR3C (Invitrogen) and Gateway cloned into pCL113 (pBATL). The coding region of LFY was cloned into pCL112 (pBATL) to create the nYFP. p35S::2xmCherry was cloned into pEarley102 (Earley et al., 2006). The control protein (NCb: TDY1-NLS in pCL113) was previously described (Ma et al., 2009). Constructs were transformed into onion epidermal cells using the PDS-1000/He Biolistic Particle Delivery System (BioRad, Hercules, CA, USA) as described by Ma et al. (Ma et al., 2009). Protein interactions were observed using an Olympus MVX10 fluorescent microscope.
RESULTS
LMI2 regulates the meristem identity transition
To elucidate the role of LMI2 in the meristem identity transition, we analyzed three T-DNA insertion alleles (Alonso and Stepanova, 2003): lmi2-1, lmi2-2 and lmi2-3 (Fig. 1A). In lmi2-1, the T-DNA insertion was located in the promoter region (116 bp from the transcription start site), whereas the insertions in lmi2-2 and lmi2-3 were located in the conserved MYB DNA binding domain (Fig. 1A). All three T-DNA insertions caused deletions in the LMI2 locus ranging in size from 4 to 41 bp (Fig. 1A).
Fig. 1.

lmi2 mutants cause a meristem identity phenotype. (A) Map of the LMI2 locus. Gray boxes, exons; gray line, 5′ and 3′ UTR and introns; dark green boxes, MYB DNA binding domain; light green box, conserved amino acid motif found in LMI2 and its homologs (Kranz et al., 1998; Stracke et al., 2001); triangles, T-DNA insertions. The lines connecting each T-DNA to the sequence denote the size of the deletion caused by each insertion. P1-P4, primers used for RT-PCR. (B) Semi-quantitative RT-PCR of LMI2 expression performed on nine-day-old seedlings for each T-DNA insertion line and Col (wild type). Primers used (see A for location) are indicated at the left. The EUKARYOTIC TRANSLATION INITIATION FACTOR 4A (EIF4A) gene was used as an internal control. (C) Number of secondary inflorescences formed in lmi2 single mutants compared with Col (wild type). (D) lfy-10 and lmi2-2 lfy-10 mutant phenotypes, with close-ups of the inflorescence apices (insets). Arrow indicates a secondary inflorescence subtended by a cauline leaf. Arrowheads indicate lateral organs formed in lmi2-2 lfy-10 and lfy-10 at a comparable stage. Scale bar: 1 cm. (E) Number of secondary inflorescences of lmi2-2 lfy-10 compared with lfy-10 (weak), lfy-9 (intermediate) and lfy-1 (strong) alleles. (F) qRT-PCR of LMI2 expression in thirteen-day-old lfy-10, pLMI2:LMI2 lmi2-2 lfy-10 and lmi2-2 lfy-10 seedlings. (G) Number of secondary inflorescences formed in pLMI2:LMI2 lmi2-2 lfy-10 compared with lfy-10 and lmi2-2 lfy-10. **P<10–3 (lmi2-1 compared with Col); ***P<10–9 (lmi2-2 and lmi2-2 lfy-10 compared with Col and lfy-10, respectively); one-tailed Student's t-test. All values represent mean ± s.e.m.
Both lmi2-2 and lmi2-3 expressed RNA upstream of the T-DNA insertion, suggesting that they are not RNA-null alleles (Fig. 1B). However, we did not detect LMI2 expression in either the lmi2-2 or the lmi2-3 mutant using primers flanking the T-DNA insertions (Fig. 1B). Hence, these insertions probably give rise to a truncated non-functional LMI2 protein lacking part of the conserved DNA binding domain. The lmi2-1 mutant, however, expressed elevated levels of LMI2 RNA (Fig. 1B). As the T-DNA insertion in lmi2-1 is located in the promoter region, it is likely that this insertion generates a full length LMI2 transcript. Nonetheless, our combined data (see below) suggests that lmi2-1 is a loss-of-function allele. Because lmi2-2 and lmi2-3 have similar T-DNA insertion sites, we chose to focus on the lmi2-1 and lmi2-2 alleles.
We assessed the timing of the meristem identity transition in lmi2 mutants compared with wild type by counting the number of secondary inflorescences and cauline leaves formed prior to the formation of the first flower (Saddic et al., 2006; Yamaguchi et al., 2009). Flowering time was measured by counting the number of rosette leaves (see Table S1 in the supplementary material) (Yamaguchi et al., 2009). lmi2-2 displayed a statistically significant increase in the number of cauline leaves and secondary inflorescences formed compared with wild type in five independent experiments (Table 1; Fig. 1C), suggesting that LMI2 plays a non-redundant role in the meristem identity transition. lmi2-1 exhibited a more subtle delay in the meristem identity transition that differed significantly from wild type in only some of the experiments performed (Table 1; Fig. 1C).
Table 1.
Meristem identity phenotypes of lmi2 mutants

All three lmi2 alleles significantly enhanced the meristem identity phenotype of the weak lfy-10 mutant in at least six independent experiments (Table 1; Fig. 1D,E). lmi2-2 lfy-10 double mutants showed the strongest meristem identity delay, essentially phenocopying the lfy-1 null mutant (Fig. 1E). In addition, in the lfy-10 background, lmi2-2, and to a lesser extent lmi2-1, caused a delay in the meristem identity transition in heterozygotes (see Fig. S1 in the supplementary material). Hence, LMI2 is a dosage-sensitive gene, at least under conditions when LFY activity is impaired. LFY itself is also dosage dependent (Blazquez et al., 1997; Okamuro et al., 1996), highlighting the sensitivity of this pathway to the level of both regulators. Finally, lmi2-2 and lmi2-1 displayed a subtle delay in flowering time (see Table S1 in the supplementary material) both as single mutants and in the lfy-10 genetic background.
We next tested whether the mutations in LMI2 caused the delay in the meristem identity transition by performing phenotypic rescue. Transformation of lmi2-2 lfy-10 with a genomic copy of LMI2 (pLMI2:LMI2) restored LMI2 expression to a level similar to that observed in lfy-10 (Fig. 1F). In addition, pLMI2:LMI2 fully rescued the enhanced meristem identity defects of lmi2-2 lfy-10 relative to lfy-10 (Fig. 1G).
To test whether LMI2 has additional LFY-independent roles during the meristem identity transition, we crossed the lmi2-2 allele to the lfy-1 null mutant. lmi2-2 significantly enhanced the meristem identity transition defect of lfy-1 (Table 1) indicating that LMI2 acts both downstream of and in parallel to LFY in this pathway. This is similar to AP1, which also acts downstream of and in parallel to LFY (Bowman et al., 1993).
LMI2 is expressed in the inflorescence meristem, in young floral primordia and in flowers
We first examined the expression of LMI2 during the meristem identity transition using a bacterial β-glucuronidase (GUS) transcriptional reporter. In nine-day-old wild-type seedlings, LMI2:GUS was expressed in the center of the rosette close to the shoot apex (Fig. 2A) in a pattern roughly similar to that of pLFY:GUS (Fig. 2E). In the inflorescence, the LMI2:GUS reporter was expressed in the meristem proper and in young floral primordia, as well as in the carpels of older flowers (Fig. 2B-D; see Fig. S2A in the supplementary material). By contrast, as previously reported (Blazquez et al., 1997), pLFY:GUS expression was absent from the meristem proper but was observed in young floral primordia as well as in older flower primordia (Fig. 2F,G; see Fig. S2B in the supplementary material). In addition, both LMI2:GUS and pLFY:GUS were strongly expressed in secondary inflorescences (Fig. 2D,H). Thus, LMI2:GUS and pLFY:GUS have overlapping, but not identical, expression patterns during reproductive development.
Fig. 2.

LMI2 is expressed in the initiating floral primordia and in developing flowers. (A-H) Expression of LMI2:GUS (A-D) and pLFY:GUS (E-H). Scale bars: 1 mm. (A,E) Nine-day-old seedlings. (B,F) Young (1 cm bolt) primary inflorescences. (C,G) Higher magnification of the shoot apices shown in B and F. Black arrowheads point to stage 1 floral primordia (p1) and asterisks indicate the shoot apical meristem. (D,H) GUS reporter expression in flowers and secondary inflorescences formed on 1 cm bolt primary inflorescences. Black arrowheads point to secondary inflorescences. (I-N) LMI2 expression based on in situ hybridization. Scale bars: 100 μm. Numbers indicate the developmental stage of young floral primordia (Smyth et al., 1990). Expression in wild type (I,K-N) and lfy-1 (J). Tissues assayed were: primary inflorescence apices (1 cm bolt; I-K), developing flowers at stage 4 (L), stage 7 (M) and stage 6 (asterisk), as well as stage 8 (N). Sense probe control is shown in K.
LMI2:GUS expression was reduced in the shoot apex of intermediate lfy-9 mutants compared with wild-type seedlings (see Fig. S2C,D in the supplementary material). Conversely, steroid treatment of an inducible version of LFY, LFY-GR (William et al., 2004), resulted in elevated LMI2:GUS expression in seedlings; this was not observed in steroid treated wild-type seedlings expressing LMI2:GUS (see Fig. S2E,F in the supplementary material). Therefore, LFY acts on LMI2 cis regulatory elements present in this reporter construct, consistent with in vivo LFY binding to this locus (Winter et al., 2011).
We next examined endogenous LMI2 expression by in situ hybridization. LMI2 was expressed throughout the shoot apical meristem of primary inflorescences, with the highest expression observed in the young flower primordia (Fig. 2I). LMI2 expression was reduced, but not absent, in the young flower primordia of lfy-1 null mutant apices (Fig. 2J). No signal was observed using a sense probe (Fig. 2K). The residual LMI2 expression in lfy-1 is consistent with our genetic data that revealed an LFY-independent role for LMI2 in addition to its function downstream of LFY.
Subsequent to the meristem identity transition, LMI2 was expressed in stage 2 to stage 4 flowers (Fig. 2L; data not shown) (Smyth et al., 1990) and in the developing stamens and carpels of older flowers from stage 6 to stage 8 (Fig. 2M,N). Eventually, in stage 8 flowers, LMI2 expression decreased in the developing stamens but persisted in the carpels (Fig. 4N).
Fig. 4.

LMI2 is required for proper activation of AP1 expression. (A) AP1, CAL and FUL expression in lfy-10 and lmi2-2 lfy-10 seedlings based on qRT-PCR at days 9, 11, 13 and 15. Values represent mean ± s.e.m. (B-E) AP1 expression based on in situ hybridization of eleven-day-old wild-type (Col; B) and thirteen-day-old lfy-10 (C) and lmi2-2 lfy-10 (D,E) seedlings. Scale bars: 100 μm. (F) Map of the AP1 locus. Light purple boxes, 5′ and 3′ UTRs; dark purple boxes, exons; black lines, introns and intergenic regions; asterisks, predicted plant MYB binding sites with a score exceeding the threshold score (Steffens et al., 2004); horizontal bars, regions amplified in ChIP q-PCR. Binding sites of known regulators of AP1 are shown below the locus (see text for details). (G) Rescue of AP1 expression in eleven-day-old pLMI2:LMI2-HA lmi2-2 seedlings. (H) ChIP-qPCR in eleven-day-old lmi2-2 and pLMI2:LMI2-HA lmi2-2 seedlings to assess LMI2 binding to AP1 regulatory regions. Immunoprecipitated DNA is represented as fold enrichment relative to the lmi2-2 control. Values shown are mean ± s.e.m. The heterochromatic TA3 retrotransposon (Konieczny et al., 1991) served as a negative ChIP control.
lmi2-1 acts as a loss-of-function allele
lmi2-1 displayed elevated LMI2 expression in seedlings based on semi-quantitative RT-PCR (Fig. 1B), yet behaved as a loss-of-function allele (Table 1; Fig. 1C; see Fig. S1 in the supplementary material). Moreover, the defect in lmi2-1 lfy-10 was rescued by pLMI2:LMI2 (Fig. 3A). In contrast to the wild type, LMI2 expression was undetectable in lmi2-1 shoot apices and young flower primordia (Fig. 3B,C), similar to the sense control (Fig. 3D). Thus, in lmi2-1 mutants, LMI2 is absent from the initiating floral primordia, where it is required for the meristem identity transition. This suggests that the increased LMI2 levels observed by RT-PCR could be due to ectopic LMI2 expression. Indeed, whereas LMI2 expression was very low in the roots and leaves of nine-day-old wild-type seedlings, it was strongly expressed in these tissues in lmi2-1 (a 40-fold and 400-fold increase, respectively; Fig. 3E). Based on our combined findings, we conclude that the T-DNA insertion in lmi2-1 apparently disrupts the LMI2 promoter, causing loss of LMI2 expression in the shoot apical meristem and in the young flower primordia. At the same time, the insertion causes ectopic and elevated LMI2 expression, perhaps from a promoter located in the T-DNA insertion.
Fig. 3.

The T-DNA insertion in lmi2-1 causes misregulation of LMI2. (A) The number of secondary inflorescences in pLMI2:LMI2 lmi2-1 lfy-10 compared with lfy-10 and lmi2-1 lfy-10. P-values for one-tailed Student's t-test are indicated. (B,C) LMI2 expression based on in situ hybridization in wild-type (B) and lmi2-1 (C) 1 cm bolt primary inflorescences. (D) LMI2 sense probe control. Scale bars: 100 μm. (E) qRT-PCR analysis of LMI2 expression in roots (including hypocotyl), leaves (including cotyledons) and whole seedlings from nine-day-old wild type (Col) and lmi2-1 mutants. Values are mean ± s.e.m.
LMI2 is required for proper AP1 upregulation
To place LMI2 in the meristem identity pathway, we examined the expression of the direct LFY targets AP1, CAL, LMI1 to LMI5, and that of another meristem identity regulator, FRUITFULL (FUL) (Ferrandiz et al., 2000; Wagner et al., 1999; William et al., 2004) in lfy-10 single mutants compared with lmi2-2 lfy-10 double mutants during the meristem identity transition (Fig. 4A, see Fig. S3A in the supplementary material). We conducted a time-course experiment spanning time points prior to, during and immediately subsequent to the meristem identity transition for all genotypes tested (Fig. 4A,B-E) (William et al., 2004; Yamaguchi et al., 2009). Although we did not observe a reduction in the expression of LMI1, LMI3 or LMI5, we observed a subtle reduction in the expression of LMI4 and a pronounced (approximately fourfold) reduction in the expression of AP1 in lmi2-2 lfy-10 compared with lfy-10 at day 13 (Fig. 4A; see Fig. S3A in the supplementary material). Indeed, AP1 expression was induced more slowly in the double mutant compared with lfy-10 (Fig. 4A). By contrast, induction of CAL and FUL expression was very similar in lfy-10 and lmi2-2 lfy-10 plants, suggesting that the observed defect in AP1 upregulation is specific. AP1 expression was also reduced in lmi2-2/+ lfy-10 plants relative to lfy-10 mutants (see Fig. S3B in the supplementary material), consistent with the observed dosage sensitivity of LMI2, as well as in lmi2-2 single mutant seedlings compared with wild type (Fig. 4G). Our combined data suggest that LMI2 acts upstream of AP1.
lfy null mutants cause a delay, but not a loss in AP1 expression; AP1 is expressed in the flowers that eventually form in these mutants (Ruiz-Garcia et al., 1997; Wagner et al., 1999). Likewise, based on qRT-PCR, AP1 is upregulated in lmi2-2 lfy-10, reaching expression levels similar to those observed in lfy-10 at day 15 (Fig. 4A), when flower patterning is initiated (see Fig. S3C in the supplementary material).
We next examined AP1 upregulation in wild-type, lfy-10 and lmi2-2 lfy-10 seedlings using in situ hybridization. By day 13, all three genotypes had initiated the first flowers. AP1 expression was much reduced in stage 1 or 2 flower primordia in thirteen-day-old lmi2-2 lfy-10 and the lfy-10 mutants relative to wild type (Fig. 4B-E; data not shown). In addition, AP1 expression levels were slightly more reduced in developing flower primordia of lmi2-2 lfy-10 compared with lfy-10 (Fig. 4C-E), and in the double mutants especially in the shoot meristem proximal region of stage 2 flower primordia (Fig. 4D,E).
To test whether LMI2 can regulate AP1 expression directly, we scanned the AP1 locus for the presence of plant MYB binding sites using AthaMap (http://www.athamap.de/) (Steffens et al., 2004). Eight predicted MYB binding sites were found in the 5′ upstream region; two in the introns and one in the first exon of AP1 (Fig. 4F). We next examined whether LMI2 binds to AP1 regulatory regions in vivo by anti-HA chromatin immunoprecipitation (ChIP) followed by qPCR using plants expressing a HA-tagged genomic version of LMI2 driven from its own promoter (pLMI2:LMI2-HA). The LMI2-HA fusion protein is biologically active, as pLMI2:LMI2-HA lmi2-2 rescued the reduced AP1 expression observed in lmi2-2 mutants (Fig. 4G). LMI2-HA was recruited to the AP1 promoter and bound to region six of AP1, which is very close to the known or predicted binding sites of other regulators of AP1, including LFY (Fig. 4F,H) (Abe et al., 2005; Parcy et al., 1998; Wang et al., 2009; Wigge et al., 2005; William et al., 2004; Winter et al., 2011; Xu et al., 2010; Yamaguchi et al., 2009). By contrast, we did not see enrichment of LMI2-HA relative to control lmi2-2 plants in the remaining regions of the AP1 locus, suggesting that the binding of LMI2 at region six is specific (Fig. 4H). Taken together, our data suggest that LMI2 directly activates AP1 expression during the meristem identity transition.
To test whether LMI2 acts solely to induce AP1 or whether it regulates other factors during the meristem identity transition, we crossed lmi2-2 to the strong ap1-10 mutant and examined the timing of the meristem identity transition. We did not observe an increase in the number of secondary inflorescences in lmi2-2 ap1-10 compared with ap1-10. There was, however, a significant increase in the number of cauline leaves produced in lmi2-2 ap1-10 compared with ap1-10 (Table 2). AP1 does not play a significant role in cauline leaf suppression during the floral transition (Bowman et al., 1993; Schultz and Haughn, 1993). Thus, like LFY (Liljegren et al., 1999), LMI2 functions through an AP1-independent pathway to suppress cauline leaf formation. We conclude that LMI2 acts both upstream of and in parallel to AP1 during the meristem identity transition.
Table 2.
Meristem identity phenotypes of lmi2 ap1 mutants

Interactions between LMI2 and LFY
LMI2 binds very close to the known LFY binding site in the AP1 locus (one putative LMI2 binding site in region six is 6 bp downstream of the LFY binding site; data not shown) (Winter et al., 2011). Hence, LMI2 and LFY might interact physically. Indeed, based on pull-down assays, LMI2 interacted with GST-LFY (Fig. 5A). Full length LFY protein homodimerized, as previously proposed (Hames et al., 2008), serving as a positive control. A negative control protein (see Materials and methods for details) did not interact with GST-LFY, confirming the specificity of the observed interactions (Fig. 5A).
Fig. 5.

LMI2 interacts physically with LFY. (A) In vitro GST-pull-down assay. GST-tagged LFY protein incubated with in vitro translated LFY, LMI2 and a negative control peptide (NCa). 5% input is shown. Input and pull-down (PD) were run on the same gel, spaces between lanes denote irrelevant samples removed from the gel image. Molecular weight markers (kDa) are indicated on the right. (B) Yeast two-hybrid assay. Growth of yeast transformed with pDBLeu-LMI2N bait construct and pDEST22-LFY or pDEST22 alone (EV) on –Trp –Leu/SD plates (–WL) or –Trp –Leu –His/SD plates (–WLH). (C) Interaction of LMI2N and LMI2C with LFY based on bimolecular fluorescence complementation (BiFC). Left: 35S:2XmCherry transformation control. Right: protein interactions. Positive control: nY:LFY and cY:LFY. Negative control: nY:LFY and cY:NCb.
Based on yeast two-hybrid assays, the N-terminal half of LMI2 (LMI2N) showed a weak, but reproducible interaction with LFY (Fig. 5B). The C-terminal domain of LMI2 (LMI2C) also interacted with LFY in yeast (data not shown). This interaction was more difficult to observe because, as previously reported (Zhang et al., 2009), this domain of LMI2 displays transcriptional activation activity. Finally, bimolecular fluorescence complementation (BiFC) was used to test for an in vivo interaction between LMI2 and LFY. Both the LMI2N and, to a lesser extent, LMI2C interacted with LFY (Fig. 5C). Again, LFY interacted with itself. By contrast, a negative control protein did not interact with LFY, suggesting the observed interactions were specific. The combined data suggest that LFY and LMI2 can form heterodimers.
During the floral transition, LFY and AP1 act in a positive feedback regulatory loop (Ferrandiz et al., 2000; Kaufmann et al., 2010; Liljegren et al., 1999). In light of this, we examined whether LMI2 can also feedback to regulate LFY. Indeed, LFY levels were reduced in lmi2-2 lfy-10 compared with lfy-10 mutants throughout the meristem identity transition (Fig. 6A). Furthermore, LFY levels were reduced in eleven-day-old lmi2-2 seedlings compared with wild type (Col) (Fig. 6B). To determine whether the reduction in LFY in lmi2 mutants was an indirect consequence of reduced AP1 expression in these mutants or whether LMI2 directly regulated LFY levels, we used ChIP to examine LMI2 binding to LFY regulatory regions. We tested binding of LMI2 to three predicted MYB binding sites in the 5′ upstream regulatory region: two sites in exon one and two sites in the second intron of LFY (Fig. 6C). We did not see binding of LMI2-HA to the promoter or intron regions of LFY, but we did observe a subtle enrichment at region four in exon one (Fig. 6D). Although one other LFY regulator has previously been shown to bind this region (Yamaguchi et al., 2009), further experiments are needed to determine whether the feedback from LMI2 to LFY is direct.
Fig. 6.

LMI2 regulates LFY expression by positive feedback during the floral transition. (A) LFY expression based on qRT-PCR in lfy-10 and lmi2-2 lfy-10 seedlings at days 9, 11, 13 and 15. (B) LFY expression in eleven-day-old wild-type (Col) and lmi2-2 seedlings. (C) LFY Locus. Light red boxes, 5′ and 3′ UTRs; dark red boxes, exons; black lines, introns and intergenic regions; asterisks, predicted plant MYB binding sites (see Fig. 4F). (D) ChIP q-PCR to test for LMI2-HA binding to LFY regulatory loci. See Fig. 4H for details on the ChIP analysis. Values shown are mean ± s.e.m.
DISCUSSION
LMI2 is a meristem identity regulator downstream of LFY
We show here that the direct LFY target and MYB transcription factor LMI2 is required for correct timing of the meristem identity transition in Arabidopsis. LMI2 was identified by two independent genomic approaches as a direct LFY-regulated and LFY-bound target during meristem identity transition (William et al., 2004; Winter et al., 2011). Notably, unlike two other known meristem identity regulator mutants (cal and lmi1) (Bowman et al., 1993; Saddic et al., 2006), lmi2 single mutants displayed a statistically significant delay in the meristem identity transition, suggesting a central role for this transcription factor in the timing of flower formation. Thus far, only one other direct LFY target has a non-redundant role in this vital developmental transition: AP1 (Bowman et al., 1993; Weigel et al., 1992).
Additional roles for LMI2 at other stages of reproductive development
The observed LMI2 expression pattern suggests that LMI2 might have a broad role in reproductive development. Like many flowering-time regulators (Abe et al., 2005; Hempel et al., 1997; Lee et al., 2000; Wigge et al., 2005), LMI2 was expressed in the shoot apex, and LMI2 controls the timing of bolting. In addition, both LFY and LMI2 were expressed in older flower primordia. Unlike lfy mutants (Huala and Sussex, 1992; Weigel et al., 1992), lmi2 mutants did not display noticeable floral homeotic defects nor did they enhance the floral homeotic defects of weak lfy mutants (data not shown), suggesting that LMI2 might have a different role in flower development.
LMI2 directly activates AP1 to promote floral fate
AP1 upregulation signals commitment to flower formation and, therefore, must be tightly controlled for proper timing of the meristem identity transition (Bowman et al., 1993; Kaufmann et al., 2010; Mandel et al., 1992; Wellmer and Riechmann, 2010). Here, we provide evidence that LMI2 directly upregulates AP1 expression during the meristem identity transition. The effect of LMI2 on AP1 expression is specific and is not due to a general delay in phase transitions, because accumulation of other meristem identity regulators, such as FUL or CAL (Bowman et al., 1993; Ferrandiz et al., 2000), are not altered in lmi2-2 lfy-10 mutants. LMI2 induction precedes that of AP1 and both are expressed in stage 1 floral primordia, where AP1 directs flower development (this study) (Liljegren et al., 1999; Mandel et al., 1992; Schmid et al., 2005). LMI2 binds to a region of the AP1 locus also occupied by many other transcription factors in vivo, including LFY (Wang et al., 2009; William et al., 2004; Xu et al., 2010; Yamaguchi et al., 2009), thus defining a critical AP1 cis regulatory module (Jeziorska et al., 2009; Wilczynski and Furlong, 2010).
LMI2 and LFY interact physically
The LMI2 and LFY binding sites on the AP1 promoter are very close to each other and, based on three independent assays, the LMI2 and LFY proteins interact physically. MYB proteins are known to interact with other transcription factors to regulate gene expression (Li et al., 2009; Shin et al., 2007; Zimmermann et al., 2004). LFY also interacts with cofactors, including at least one other downstream target, to regulate gene expression (Chae et al., 2008; Lenhard et al., 2001; Liu et al., 2009b; Lohmann et al., 2001; Winter et al., 2011). For example, LFY directly upregulates the floral homeotic regulator SEPALLATA3 (SEP3) and, in turn, these two factors interact physically to activate the class B and C floral homeotic genes (Liu et al., 2009b; Winter et al., 2011).
Based on the recent finding that LFY acts as both a direct transcriptional activator and repressor (Parcy et al., 2002; William et al., 2004; Winter et al., 2011), it seems likely that cofactors modulate the effect of LFY on gene expression. Consistent with this idea, LFY alone is unable to activate gene expression from the AP1 promoter in yeast: it can only act as a transcriptional activator in this system when fused to a strong activation domain (Parcy et al., 1998; Winter, 2011). It is likely that LFY also needs a co-activator for AP1 induction in vivo. LMI2 is a good candidate for this LFY co-activator: it has strong transactivation activity based on yeast assays, it is induced by LFY prior to AP1 upregulation and can form heterodimers with LFY (this study) (Blazquez et al., 1997; Hempel et al., 1997; Schmid et al., 2005; Schmid et al., 2003; Zhang et al., 2009). Moreover, the temporal delay in the formation of the first flower is very similar in ap1 and lmi2 single mutants (this study) (Xu et al., 2010); thus, LMI2 might be sufficient for LFY-dependent activation of AP1 expression. However, we cannot rule out that other LFY co-factors contribute to this process.
The LFY, LMI2 and AP1 regulatory network might contribute to an abrupt and robust meristem identity transition
The observed interactions between LFY, LMI2 and AP1 represent a coherent feed-forward loop (Fig. 7) (Alon, 2007), a regulatory circuit with crucial roles in control of developmental processes in many organisms (Alon, 2007; Mangan et al., 2003; Shen-Orr et al., 2002). The type of coherent feed-forward loop observed here serves as a persistence detector for inductive signal(s) and as a temporal delay element (Alon, 2007). Thus, transient inductive cues that cause a temporary increase in LFY, but not in LMI2, will delay LFY-dependent upregulation of AP1.
Fig. 7.
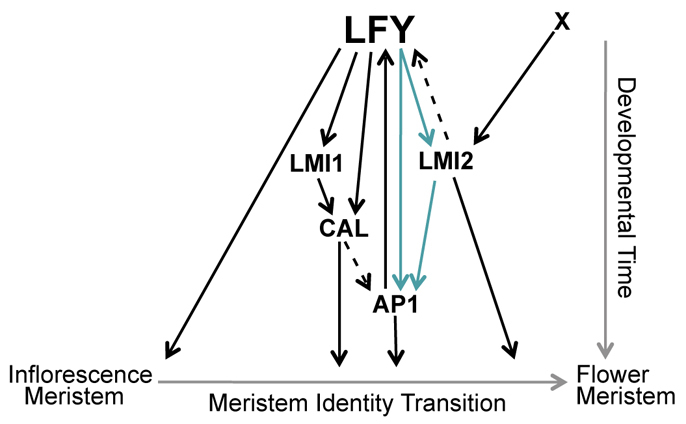
Meristem identity pathway downstream of LFY. The LFY transcription factor directly activates multiple downstream factors during the meristem identity transition, including CAL, LMI1, AP1 and LMI2 (Saddic et al., 2006; William et al., 2004). LMI2 is also upregulated by another factor, `X', in a pathway parallel to LFY. LFY, LMI2 and AP1 act in a feed-forward loop (blue arrows) to initiate the meristem identity transition, and LMI2 and AP1 positively feedback to LFY (this study) (Kaufmann et al., 2010). Interactions, which could be indirect or direct, are indicated by dashed arrows.
This finding is consistent with prior observations. For example, LFY upregulation is directed by environmental cues, such as changes in day length or ambient temperature (Amasino, 2010; Kobayashi and Weigel, 2007; Liu et al., 2009a); these stimuli are inherently noisy inputs, yet the transition to flower formation is abrupt in Arabidopsis, without formation of intermediate structures (Parcy, 2005). In addition, as discussed above, AP1 induction is delayed with respect to that of LFY and LMI2, and is reduced in both single mutants. Finally, as predicted by the feed-forward loop model, LMI2 was a haplo-insufficient, rate-limiting factor for AP1 induction downstream of LFY, at least under conditions when LFY activity was compromised.
In addition to the feed-forward loop uncovered here, LFY directs at least two additional coherent feed-forward loops, one of which is also linked to the meristem identity transition and involves the direct LFY targets LMI1 and CAL (Fig. 7) (Kaufmann et al., 2009; Liu et al., 2009b; Saddic et al., 2006; William et al., 2004; Winter et al., 2011). Among these feed-forward loops involving LFY, the LFY-LMI2-AP1 feed-forward loop stands out as it alone comprises three regulators that have non-redundant roles in the process they regulate; hence, it might represent a crucial regulatory module in the meristem identity transition.
In Arabidopsis, the meristem identity transition is not only precise (it occurs after formation of a defined number of secondary inflorescences subtended by cauline leaves), but also robust (no reversion from flower to inflorescence fate is observed) (Amasino, 2010; Blazquez et al., 2006; Liu et al., 2009a; Tooke et al., 2005). As outlined above, the LFY-LMI2-AP1 feed-forward loop is likely to contribute to the precision of this developmental transition; its robustness, however, might be due to positive feedback (Alon, 2007). Indeed, a positive direct feedback from AP1 to LFY has recently been described (Kaufmann et al., 2010; Liljegren et al., 1999). We show here that LMI2 also positively regulates LFY: LFY expression was reduced in lmi2-2 single and double mutants. This reduction of LFY expression could be an indirect effect, triggered by the reduced AP1 expression levels observed in lmi2-2 mutants. However, the positive feedback might, in part, be direct as LMI2 was weakly recruited to the LFY locus. The observed enhancement of the ap1 mutant meristem identity defect by lmi2 is consistent with this hypothesis. It is likely that the AP1 and possible LMI2 feedback loops keep the LFY-LMI2-AP1 feed-forward loop active after full AP1 upregulation has been achieved. Indeed, AP1 directly downregulates upstream activators of itself and of LFY (Kaufmann et al., 2010), providing further support for the idea that the combined feed-forward and feedback loop is self-maintained.
It will be of interest to examine these regulatory interactions in other flowering plant species. In light of this question, we note that LMI2 separated from its closest homologs, the MIXTA/MYB16 and MYB106 genes, before the split of the monocots from the eudicots ∼100 million years ago (Baumann et al., 2007). This raises the possibility that the function of LMI2 in reproductive development evolved early in the flowering plant lineage and might be conserved in other angiosperm species.
Supplementary Material
Acknowledgments
We are grateful to David M. Braun for the BiFC negative control protein and to Jennifer Pfluger for the yeast two-hybrid and BiFC LFY constructs. We thank Kim Gallagher, John Wagner, Matthew Willmann and Cara M. Winter for critical comments and suggestions on this manuscript. This research was funded by NSF grant IOS 0849298 to D.W. and a NIH Training Grant 5-T32-HD007516 (Developmental Biology) as well as a University of Pennsylvania, School of Arts and Sciences, Dissertation Research Fellowship to J.J.P. Deposited in PMC for release after 12 months.
Footnotes
Competing interests statement
The authors declare no competing financial interests.
Supplementary material
Supplementary material for this article is available at http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.063073/-/DC1
References
- Abe M., Kobayashi Y., Yamamoto S., Daimon Y., Yamaguchi A., Ikeda Y., Ichinoki H., Notaguchi M., Goto K., Araki T. (2005). FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 309, 1052-1056 [DOI] [PubMed] [Google Scholar]
- Alon U. (2007). Network motifs: theory and experimental approaches. Nat. Rev. Genet. 8, 450-461 [DOI] [PubMed] [Google Scholar]
- Alonso J. M., Stepanova A. N. (2003). T-DNA mutagenesis in Arabidopsis. Methods Mol. Biol. 236, 177-188 [DOI] [PubMed] [Google Scholar]
- Amasino R. (2010). Seasonal and developmental timing of flowering. Plant J. 61, 1001-1013 [DOI] [PubMed] [Google Scholar]
- Araki T. (2001). Transition from vegetative to reproductive phase. Curr. Opin. Plant Biol. 4, 63-68 [DOI] [PubMed] [Google Scholar]
- Baumann K., Perez-Rodriguez M., Bradley D., Venail J., Bailey P., Jin H., Koes R., Roberts K., Martin C. (2007). Control of cell and petal morphogenesis by R2R3 MYB transcription factors. Development 134, 1691-1701 [DOI] [PubMed] [Google Scholar]
- Baurle I., Dean C. (2006). The timing of developmental transitions in plants. Cell 125, 655-664 [DOI] [PubMed] [Google Scholar]
- Blazquez M. A., Soowal L. N., Lee I., Weigel D. (1997). LEAFY expression and flower initiation in Arabidopsis. Development 124, 3835-3844 [DOI] [PubMed] [Google Scholar]
- Blazquez M. A., Ferrandiz C., Madueno F., Parcy F. (2006). How floral meristems are built. Plant Mol. Biol. 60, 855-870 [DOI] [PubMed] [Google Scholar]
- Bowman J. L., Alvarez J., Weigel D., Meyerowitz E. M., Smyth D. R. (1993). Control of flower development In Arabidopsis thaliana By Apetala1 and interacting genes. Development 119, 721-743 [Google Scholar]
- Chae E., Tan Q. K., Hill T. A., Irish V. F. (2008). An Arabidopsis F-box protein acts as a transcriptional co-factor to regulate floral development. Development 135, 1235-1245 [DOI] [PubMed] [Google Scholar]
- Dubos C., Stracke R., Grotewold E., Weisshaar B., Martin C., Lepiniec L. (2010). MYB transcription factors in Arabidopsis. Trends Plant Sci. 15, 573-581 [DOI] [PubMed] [Google Scholar]
- Earley K. W., Haag J. R., Pontes O., Opper K., Juehne T., Song K., Pikaard C. S. (2006). Gateway-compatible vectors for plant functional genomics and proteomics. Plant J. 45, 616-629 [DOI] [PubMed] [Google Scholar]
- Ferrandiz C., Gu Q., Martienssen R., Yanofsky M. F. (2000). Redundant regulation of meristem identity and plant architecture by FRUITFULL, APETALA1 and CAULIFLOWER. Development 127, 725-734 [DOI] [PubMed] [Google Scholar]
- Hames C., Ptchelkine D., Grimm C., Thevenon E., Moyroud E., Gerard F., Martiel J. L., Benlloch R., Parcy F., Muller C. W. (2008). Structural basis for LEAFY floral switch function and similarity with helix-turn-helix proteins. EMBO J. 27, 2628-2637 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hempel F. D., Weigel D., Mandel M. A., Ditta G., Zambryski P. C., Feldman L. J., Yanofsky M. F. (1997). Floral determination and expression of floral regulatory genes in Arabidopsis. Development 124, 3845-3853 [DOI] [PubMed] [Google Scholar]
- Huala E., Sussex I. M. (1992). LEAFY interacts with floral homeotic genes to regulate Arabidopsis floral development. Plant Cell 4, 901-913 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jakoby M. J., Falkenhan D., Mader M. T., Brininstool G., Wischnitzki E., Platz N., Hudson A., Hulskamp M., Larkin J., Schnittger A. (2008). Transcriptional profiling of mature Arabidopsis trichomes reveals that NOECK encodes the MIXTA-like transcriptional regulator MYB106. Plant Physiol. 148, 1583-1602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeziorska D. M., Jordan K. W., Vance K. W. (2009). A systems biology approach to understanding cis-regulatory module function. Semin. Cell Dev. Biol. 20, 856-862 [DOI] [PubMed] [Google Scholar]
- Kaufmann K., Muino J. M., Jauregui R., Airoldi C. A., Smaczniak C., Krajewski P., Angenent G. C. (2009). Target genes of the MADS transcription factor SEPALLATA3: integration of developmental and hormonal pathways in the Arabidopsis flower. PLoS Biol. 7, e1000090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaufmann K., Wellmer F., Muino J. M., Ferrier T., Wuest S. E., Kumar V., Serrano-Mislata A., Madueno F., Krajewski P., Meyerowitz E. M., et al. (2010). Orchestration of floral initiation by APETALA1. Science 328, 85-89 [DOI] [PubMed] [Google Scholar]
- Kobayashi Y., Weigel D. (2007). Move on up, it's time for change mobile signals controlling photoperiod-dependent flowering. Genes Dev. 21, 2371-2384 [DOI] [PubMed] [Google Scholar]
- Komeda Y. (2004). Genetic regulation of time to flower in Arabidopsis thaliana. Annu. Rev. Plant Biol. 55, 521-535 [DOI] [PubMed] [Google Scholar]
- Konieczny A., Voytas D. F., Cummings M. P., Ausubel F. M. (1991). A superfamily of Arabidopsis thaliana retrotransposons. Genetics 127, 801-809 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koornneef M., Alonso-Blanco C., Peeters A. J., Soppe W. (1998). Genetic control of flowering time in Arabidopsis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49, 345-370 [DOI] [PubMed] [Google Scholar]
- Kranz H. D., Denekamp M., Greco R., Jin H., Leyva A., Meissner R. C., Petroni K., Urzainqui A., Bevan M., Martin C., et al. (1998). Towards functional characterisation of the members of the R2R3-MYB gene family from Arabidopsis thaliana. Plant J. 16, 263-276 [DOI] [PubMed] [Google Scholar]
- Kwon C. S., Chen C., Wagner D. (2005). WUSCHEL is a primary target for transcriptional regulation by SPLAYED in dynamic control of stem cell fate in Arabidopsis. Genes Dev. 19, 992-1003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee H., Suh S. S., Park E., Cho E., Ahn J. H., Kim S. G., Lee J. S., Kwon Y. M., Lee I. (2000). The AGAMOUS-LIKE 20 MADS domain protein integrates floral inductive pathways in Arabidopsis. Genes Dev. 14, 2366-2376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lenhard M., Bohnert A., Jurgens G., Laux T. (2001). Termination of stem cell maintenance in Arabidopsis floral meristems by interactions between WUSCHEL and AGAMOUS. Cell 105, 805-814 [DOI] [PubMed] [Google Scholar]
- Li L., Yu X., Thompson A., Guo M., Yoshida S., Asami T., Chory J., Yin Y. (2009). Arabidopsis MYB30 is a direct target of BES1 and cooperates with BES1 to regulate brassinosteroid-induced gene expression. Plant J. 58, 275-286 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liljegren S. J., Gustafson-Brown C., Pinyopich A., Ditta G. S., Yanofsky M. F. (1999). Interactions among APETALA1, LEAFY, and TERMINAL FLOWER1 specify meristem fate. Plant Cell 11, 1007-1018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu C., Zhou J., Bracha-Drori K., Yalovsky S., Ito T., Yu H. (2007). Specification of Arabidopsis floral meristem identity by repression of flowering time genes. Development 134, 1901-1910 [DOI] [PubMed] [Google Scholar]
- Liu C., Thong Z., Yu H. (2009a). Coming into bloom: the specification of floral meristems. Development 136, 3379-3391 [DOI] [PubMed] [Google Scholar]
- Liu C., Xi W., Shen L., Tan C., Yu H. (2009b). Regulation of floral patterning by flowering time genes. Dev. Cell 16, 711-722 [DOI] [PubMed] [Google Scholar]
- Lohmann J. U., Hong R. L., Hobe M., Busch M. A., Parcy F., Simon R., Weigel D. (2001). A molecular link between stem cell regulation and floral patterning in Arabidopsis. Cell 105, 793-803 [DOI] [PubMed] [Google Scholar]
- Long J. A., Barton M. K. (1998). The development of apical embryonic pattern in Arabidopsis. Development 125, 3027-3035 [DOI] [PubMed] [Google Scholar]
- Ma Y., Slewinski T. L., Baker R. F., Braun D. M. (2009). Tie-dyed1 encodes a novel, phloem-expressed transmembrane protein that functions in carbohydrate partitioning. Plant Physiol. 149, 181-194 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mandel M. A., Yanofsky M. F. (1995). A gene triggering flower formation in Arabidopsis. Nature 377, 522-524 [DOI] [PubMed] [Google Scholar]
- Mandel M. A., Gustafson-Brown C., Savidge B., Yanofsky M. F. (1992). Molecular characterization of the Arabidopsis floral homeotic gene APETALA1. Nature 360, 273-277 [DOI] [PubMed] [Google Scholar]
- Mangan S., Zaslaver A., Alon U. (2003). The coherent feedforward loop serves as a sign-sensitive delay element in transcription networks. J. Mol. Biol. 334, 197-204 [DOI] [PubMed] [Google Scholar]
- Martin C., Paz-Ares J. (1997). MYB transcription factors in plants. Trends Genet. 13, 67-73 [DOI] [PubMed] [Google Scholar]
- Nakagawa T., Kurose T., Hino T., Tanaka K., Kawamukai M., Niwa Y., Toyooka K., Matsuoka K., Jinbo T., Kimura T. (2007). Development of series of gateway binary vectors, pGWBs, for realizing efficient construction of fusion genes for plant transformation. J. Biosci. Bioeng. 104, 34-41 [DOI] [PubMed] [Google Scholar]
- Oh E., Kang H., Yamaguchi S., Park J., Lee D., Kamiya Y., Choi G. (2009). Genome-wide analysis of genes targeted by PHYTOCHROME INTERACTING FACTOR 3-LIKE5 during seed germination in Arabidopsis. Plant Cell 21, 403-419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okamuro J. K., den Boer B. G., Lotys-Prass C., Szeto W., Jofuku K. D. (1996). Flowers into shoots: photo and hormonal control of a meristem identity switch in Arabidopsis. Proc. Natl. Acad. Sci. USA 93, 13831-13836 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parcy F. (2005). Flowering: a time for integration. Int. J. Dev. Biol. 49, 585-593 [DOI] [PubMed] [Google Scholar]
- Parcy F., Nilsson O., Busch M. A., Lee I., Weigel D. (1998). A genetic framework for floral patterning. Nature 395, 561-566 [DOI] [PubMed] [Google Scholar]
- Parcy F., Bomblies K., Weigel D. (2002). Interaction of LEAFY, AGAMOUS and TERMINAL FLOWER1 in maintaining floral meristem identity in Arabidopsis. Development 129, 2519-2527 [DOI] [PubMed] [Google Scholar]
- Poethig R. S. (2003). Phase change and the regulation of developmental timing in plants. Science 301, 334-336 [DOI] [PubMed] [Google Scholar]
- Roux F., Touzet P., Cuguen J., Le Corre V. (2006). How to be early flowering: an evolutionary perspective. Trends Plant Sci. 11, 375-381 [DOI] [PubMed] [Google Scholar]
- Ruiz-Garcia L., Madueno F., Wilkinson M., Haughn G., Salinas J., Martinez-Zapater J. M. (1997). Different roles of flowering-time genes in the activation of floral initiation genes in Arabidopsis. Plant Cell 9, 1921-1934 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saddic L. A., Huvermann B., Bezhani S., Su Y., Winter C. M., Kwon C. S., Collum R. P., Wagner D. (2006). The LEAFY target LMI1 is a meristem identity regulator and acts together with LEAFY to regulate expression of CAULIFLOWER. Development 133, 1673-1682 [DOI] [PubMed] [Google Scholar]
- Sang Y., Li Q. H., Rubio V., Zhang Y. C., Mao J., Deng X. W., Yang H. Q. (2005). N-terminal domain-mediated homodimerization is required for photoreceptor activity of Arabidopsis CRYPTOCHROME 1. Plant Cell 17, 1569-1584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmid M., Uhlenhaut N. H., Godard F., Demar M., Bressan R., Weigel D., Lohmann J. U. (2003). Dissection of floral induction pathways using global expression analysis. Development 130, 6001-6012 [DOI] [PubMed] [Google Scholar]
- Schmid M., Davison T. S., Henz S. R., Pape U. J., Demar M., Vingron M., Scholkopf B., Weigel D., Lohmann J. U. (2005). A gene expression map of Arabidopsis thaliana development. Nat. Genet. 37, 501-506 [DOI] [PubMed] [Google Scholar]
- Schultz E. A., Haughn G. W. (1993). Genetic analysis of the floral induction process (FLIP) in Arabidopsis. Development 119, 745-765 [Google Scholar]
- Shen-Orr S. S., Milo R., Mangan S., Alon U. (2002). Network motifs in the transcriptional regulation network of Escherichia coli. Nat. Genet. 31, 64-68 [DOI] [PubMed] [Google Scholar]
- Shin R., Burch A. Y., Huppert K. A., Tiwari S. B., Murphy A. S., Guilfoyle T. J., Schachtman D. P. (2007). The Arabidopsis transcription factor MYB77 modulates auxin signal transduction. Plant Cell 19, 2440-2453 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sieburth L. E., Meyerowitz E. M. (1997). Molecular dissection of the AGAMOUS control region shows that cis elements for spatial regulation are located intragenically. Plant Cell 9, 355-365 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson G. G., Gendall A. R., Dean C. (1999). When to switch to flowering. Annu. Rev. Cell Dev. Biol. 15, 519-550 [DOI] [PubMed] [Google Scholar]
- Smyth D. R., Bowman J. L., Meyerowitz E. M. (1990). Early flower development in Arabidopsis. Plant Cell 2, 755-767 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steeves T. A., Sussex I. M. (1989). Patterns in Plant Development. Cambridge: Cambridge University Press; [Google Scholar]
- Steffens N. O., Galuschka C., Schindler M., Bulow L., Hehl R. (2004). AthaMap: an online resource for in silico transcription factor binding sites in the Arabidopsis thaliana genome. Nucleic Acids Res. 32, D368-D372 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stracke R., Werber M., Weisshaar B. (2001). The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 4, 447-456 [DOI] [PubMed] [Google Scholar]
- Tooke F., Ordidge M., Chiurugwi T., Battey N. (2005). Mechanisms and function of flower and inflorescence reversion. J. Exp. Bot. 56, 2587-2599 [DOI] [PubMed] [Google Scholar]
- Turck F., Fornara F., Coupland G. (2008). Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Annu. Rev. Plant Biol. 59, 573-594 [DOI] [PubMed] [Google Scholar]
- Wagner D., Meyerowitz E. M. (2002). SPLAYED, a novel SWI/SNF ATPase homolog, controls reproductive development in Arabidopsis. Curr. Biol. 12, 85-94 [DOI] [PubMed] [Google Scholar]
- Wagner D., Sablowski R. W., Meyerowitz E. M. (1999). Transcriptional activation of APETALA1 by LEAFY. Science 285, 582-584 [DOI] [PubMed] [Google Scholar]
- Wang J. W., Czech B., Weigel D. (2009). miR156-regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana. Cell 138, 738-749 [DOI] [PubMed] [Google Scholar]
- Weigel D., Alvarez J., Smyth D. R., Yanofsky M. F., Meyerowitz E. M. (1992). LEAFY controls floral meristem identity in Arabidopsis. Cell 69, 843-859 [DOI] [PubMed] [Google Scholar]
- Wellmer F., Riechmann J. L. (2010). Gene networks controlling the initiation of flower development. Trends Genet. 26, 519-527 [DOI] [PubMed] [Google Scholar]
- Wigge P. A., Kim M. C., Jaeger K. E., Busch W., Schmid M., Lohmann J. U., Weigel D. (2005). Integration of spatial and temporal information during floral induction in Arabidopsis. Science 309, 1056-1059 [DOI] [PubMed] [Google Scholar]
- Wilczynski B., Furlong E. E. (2010). Dynamic CRM occupancy reflects a temporal map of developmental progression. Mol. Syst. Biol. 6, 383 [DOI] [PMC free article] [PubMed] [Google Scholar]
- William D. A., Su Y., Smith M. R., Lu M., Baldwin D. A., Wagner D. (2004). Genomic identification of direct target genes of LEAFY. Proc. Natl. Acad. Sci. USA 101, 1775-1780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winter C. M., Austin R. S., Blanvillain-Baufume S., Reback M. A., Monniaux M., Wu M. F., Sang Y., Yamaguchi A., Yamaguchi N., Parker J. E., et al. (2011). LEAFY target genes reveal floral regulatory logic, cis motifs, and a link to biotic stimulus response. Dev. Cell 20, 430-443 [DOI] [PubMed] [Google Scholar]
- Xu M., Hu T., McKim S. M., Murmu J., Haughn G. W., Hepworth S. R. (2010). Arabidopsis BLADE-ON-PETIOLE1 and 2 promote floral meristem fate and determinacy in a previously undefined pathway targeting APETALA1 and AGAMOUS-LIKE24. Plant J. 63, 974-989 [DOI] [PubMed] [Google Scholar]
- Yamaguchi A., Kobayashi Y., Goto K., Abe M., Araki T. (2005). TWIN SISTER OF FT (TSF) acts as a floral pathway integrator redundantly with FT. Plant Cell Physiol. 46, 1175-1189 [DOI] [PubMed] [Google Scholar]
- Yamaguchi A., Wu M. F., Yang L., Wu G., Poethig R. S., Wagner D. (2009). The microRNA-regulated SBP-Box transcription factor SPL3 is a direct upstream activator of LEAFY, FRUITFULL, and APETALA1. Dev. Cell 17, 268-278 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu H., Ito T., Wellmer F., Meyerowitz E. M. (2004). Repression of AGAMOUS-LIKE 24 is a crucial step in promoting flower development. Nat. Genet. 36, 157-161 [DOI] [PubMed] [Google Scholar]
- Zhang Y., Cao G., Qu L. J., Gu H. (2009). Characterization of Arabidopsis MYB transcription factor gene AtMYB17 and its possible regulation by LEAFY and AGL15. J. Genet. Genomics 36, 99-107 [DOI] [PubMed] [Google Scholar]
- Zimmermann I. M., Heim M. A., Weisshaar B., Uhrig J. F. (2004). Comprehensive identification of Arabidopsis thaliana MYB transcription factors interacting with R/B-like BHLH proteins. Plant J. 40, 22-34 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.