

Summary
Tumor development relies upon essential contributions from the tumor microenvironment and host immune alterations. These contributions may inform the plasma proteome in a manner that could be exploited for cancer diagnosis and prognosis. In this study, we employed a systems biology approach to characterize the plasma proteome response in the inducible HER2/neu mouse model of breast cancer during tumor induction, progression and regression. Mass spectrometry data derived from ∼ 1.6 million spectra identified protein networks involved in wound healing, microenvironment and metabolism that coordinately changed during tumor development. The observed alterations developed prior to cancer detection, increased progressively with tumor growth, and reverted toward baseline with tumor regression. Gene expression and immunohistochemical analyses suggested that the cancer-associated plasma proteome was derived from transcriptional responses in the non-cancerous host tissues as well as the developing tumor. The proteomic signature was distinct from a non-specific response to inflammation. Overall, the developing tumor simultaneously engaged a number of innate physiological processes, including wound repair, immune response, coagulation and complement cascades, tissue remodeling and metabolic homeostasis that were all detectable in plasma. Our findings offer an integrated view of tumor development with relevance to plasma-based strategies to detect and diagnose cancer.
Introduction
The tumor microenvironment and host factors play a major role in the establishment, progression, and metastatic dissemination of cancer (1). Stromal fibroblasts, myofibroblasts, endothelial cells, and immune cells provide growth factors, proteases, and angiogenic potential to support tumor growth (2, 3). Dendritic cells and recruited T and B cells, T-regs, macrophages, and other myeloid-derived suppressor cells, contribute both positively and negatively to multiple stages of tumor progression (4–7). Tumors can also recruit progenitor cells from the bone marrow to promote angiogenesis and to establish pre-metastatic niches in distal organs such as the lung (8–10). To date, these tumor-host interactions have been primarily studied at the cellular and tissue levels, and it remains unclear to what extent they contribute to the changes in the plasma proteome, particularly at early stages of tumor development. Proteomic advances currently allow in-depth quantitative profiling of biological fluids such as plasma spanning more than six logs of protein abundance (11, 12). Application of proteomics to mouse models provides a number of distinct advantages to interrogate the systems biology of cancer (13, 14).
We utilized a conditional transgenic Neu-induced mouse model of mammary cancer to determine the extent and source of changes in the plasma proteome at predetermined stages of tumor development (15). Neu is an activated rat homolog of the human ErbB2 (epidermal growth factor receptor 2) gene. ErbB2 is a receptor tyrosine kinase amplified and overexpressed in over 25% of human breast cancers and signaling from this oncogene is a central driver in breast cancer development. When induced with doxycycline, bitransgenic MMTV-rtTA/TetO-NeuNT mice synchronously develop invasive mammary carcinomas that recapitulate the morphologic, pathologic, and molecular features of ErbB2 positive human breast cancer (16). Doxycycline withdrawal results in transgene de-induction and tumor regression, mimicking responses of tumors to targeted therapy. We applied in-depth quantitative proteomic profiling to this model and identified a set of proteins that changed in relative abundance in plasma during tumor induction, progression, and regression. Database mining, cross referencing proteomic data with transcript profiles, and confirmatory cell-type localization studies revealed that tumor cells, the microenvironment, and systemic responses contribute to the dynamic changes in the plasma proteome with tumor development and progression.
Materials and Methods
Tissue collection
Bitransgenic FVB MMTV-rtTA/TetO-NeuNT “case” and monotransgenic FVB TetO-NeuNT “control” mice (15) were paired at weaning and maintained in the same cage. Doxycycline (2 mg/ml) was added to drinking water starting at 8 weeks of age. Mice were palpated every other day to detect mammary tumor growth. Each pair of case and control mice was euthanized on the same day by CO2 inhalation and plasma was collected as described (13).
Plasma Proteomic Analysis
The plasma proteomics workflow has been previously described (17). Briefly, case and control plasma pools (250-315 μL/pool) from 5-11 mice were immunodepleted (3 abundant proteins, MS-3, Agilent). Samples were concentrated and reduced with DTT. Isotopic intact protein labeling of cysteine residues was performed with control samples receiving 12C-acrylamide (Fluka), and case samples receiving 13C-acrylamide (Cambridge Isotope Laboratories). Case and control pools were combined for 2D-HPLC (Shimadzu) protein separation. Anion-exchange chromatography (Poros HQ/10, Applied Biosystems) used an 8-step elution (0-1000 mM NaCl). Anion-exchange fractions were separated by reversed-phase chromatography (Poros R2/10). Lyophilized fractions were digested with trypsin and 96 fractions per tumor stage were analyzed by a LTQ-Orbitrap (Thermo) mass spectrometer and NanoLC-1D (Eksigent). Online reversed-phase peptide separation and MS/MS of the 5 most abundant +2 or +3 ions (m/z 400-1800) was performed.
Each tumor stage analysis yielded ∼400,000 spectra which were searched against the mouse IPI database (v.3.48) using X!Tandem with comet scoring (18–21). PeptideProphet and ProteinProphet (22) were used for validation and protein inference. Quantification was performed by Q3 (23). Identifications with PeptideProphet probability >0.2 were submitted to ProteinProphet. Proteins were filtered using <5% error rate. Protein ratios were computed from peptides with PeptideProphet probability >0.75. Proteins with multiple IPIs and single gene name, were combined (log2 average of peptides) to give one ratio per gene name.
For proteins with multiple paired MS events of heavy and light acrylamide, a two-tailed Students t-test was used to calculate a p-value for the mean case/control ratio of the protein. Secondly, the probability for the case/control ratio for each MS event was calculated from the distribution of ratios in control/control experiments where the same sample was labeled with heavy and light acrylamide. Based on the control/control experiments, if the p-value for each individual event was <0.05, the overall protein ratio was considered significant.
Breast cancer cell line profiling
The cell line methods are previously described (24). MCF7, MDA-MB-231, and SKBR3 cell lines were obtained from ATCC, cultured for less than 6 months, and were not reauthenticated. Cells were cultured for 7 passages in SILAC DMEM (Invitrogen, dialyzed FBS, 13C-lysine) (25). After 48 hours, secreted proteins were obtained from media. For total cell extract, cells were sonicated and centrifuged to remove debris. For cell surface proteins, cells were biotinylated, extracted, sonicated, and captured by an avidin column. 1 mg of each of secreted, total cell extract, and cell surface proteins were reduced and alkylated with iodoacetamide and fractionated by reversed-phase chromatography. Fractions were digested with trypsin, and subjected to LC-MS/MS as described above.
Immunoassays
Rules Based Medicine RodentMAP v2.0 Luminex immunoassays measured 58 proteins in plasmas. Plasma levels of Igfbp5, Timp1, Lcn2, Lgals1, and Ccl8 were measured by enzyme-linked immunosorbent assays (ELISA) (R&D Systems) according to manufacturer's protocol.
Immunohistochemistry
Tissues were processed and stained with Calr (Abcam), Lgals1 (R&D Systems), Ltf (Abgent), S100a9 (NovusBio), Pdlim1 (Abcam), smooth muscle actin (Thermo), Timp1 (Abbiotec), Vasp (Abcam) or Igfbp5 (Santa Cruz) antibodies using a standard three step technique (26). Collagen staining was done using the Sigma Trichrome Kit. The University of Washington Histology Core performed CD3 and F4/80 staining. Twelve tumors were stained. For each antibody and all tumors showed similar staining patterns.
Results
Histological evaluation of mammary tumor induction, progression, and regression
Cohorts of MMTV-rtTA/TetO-NeuNT “case” and TetO-NeuNT “control” mice were sacrificed at four time points during tumor development (Fig. 1A). After five weeks on doxycycline and prior to clinically detectable lesions, mammary duct hyperplasia, intraepithelial neoplasia, and microinvasive carcinoma was apparent (Fig. 1B). By 8-12 weeks, 0.5 cm tumors consisted of hyperplastic nodules of intraepithelial neoplasia and areas of adenocarcinoma. By 14-16 weeks, 1 cm breast tumors were classified as mammary adenocarcinomas with areas of local invasion. Neoplastic cells were supported by vascular stroma and fibrous connective tissue indicating a desmoplastic reaction. Inflammatory cell infiltrates included lymphocytes, neutrophils, macrophages, plasma cells, and mast cells. Doxycycline was withdrawn from mice bearing ∼1 cm tumors and plasma samples were taken three days later, when tumors had visibly decreased in size. Histological evaluation showed regressing mammary trees with ductal ectasia, inflammation, and fibrosis. Plasma from case and control mice at each of these four tumor stages was pooled and subjected to differential isotopic labeling, extensive protein fractionation, and LC-MS/MS analysis of tryptic digests from individual fractions (17). The same plasma pools were analyzed using Luminex/Rules Based Medicine, which measured 58 proteins representing cytokines, growth factors, and other signaling proteins.
Figure 1. Histologic and plasma proteomic analysis of HER2/neu mammary tumor progression.
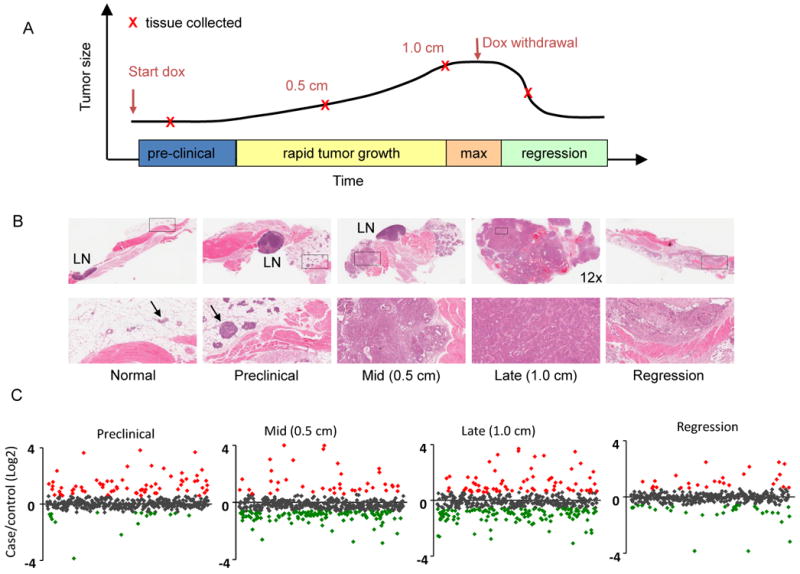
(A) Samples were collected from litter-matched case and control mice at indicated stages. (B) H&E staining of mammary tissue from bitransgenic case mice at the preclinical, 0.5 cm diameter, 1.0 cm diameter tumor stages and during tumor regression. Top row: 26× magnification unless otherwise indicated. Bottom row: 130× magnification of boxed sections from the top row. LN=lymph node. Arrows indicate breast epithelial cells within the mammary fat pads of normal and preclinical sections. (C) Case/control ratios of all quantified plasma proteins at all four tumor stages. Red indicates proteins increased >1.5 fold and green indicates proteins decreased >1.5 fold in plasma from case vs. control mice (p<0.05).
Changes in the plasma proteome prior to clinical evidence of cancer
Plasma samples from preclinical tumor-bearing mice yielded 49 proteins with increased and 17 with decreased concentrations in case compared to control mice (>1.5-fold change, p<0.05) (Fig. 1C and Supplementary Table S1). These proteins were grouped by function, biological process, and cellular localization. Proteins involved in acute phase response, immune cell signaling, including cytokines, chemokines, and macrophage and neutrophil signaling proteins, and cytoskeletal and extracellular matrix (ECM) function were increased (Fig. 2). Response to wounding was the most significant biological process altered in the proteome of mice with preclinical neoplasia (Supplementary Fig. S1A). Analysis of the subset of proteins known to be extracellular identified the acute phase response and the coagulation and complement systems as the most significantly altered pathways (Supplementary Figs. S1B and S2). Among the altered plasma proteins with known intracellular localization, those involved in glycolysis, fatty acid, and amino acid metabolism predominated (Supplementary Table S2).
Figure 2. Wound healing response components: acute phase, immune cell, and extracellular matrix proteins.

Plasma proteins that increased in tumor-bearing mice fell into major functional categories. (A) acute phase response proteins, mainly liver-derived, (B) proteins from tumor infiltrating immune cells (macrophages and neutrophils), and (C) proteins involved in cell movement and tumor microenvironment, e.g. cytoskeletal, extracellular matrix, and tumor stroma proteins. Red indicates case/control ratios >1.5 (p<0.05), black indicates no change between case and control plasma, and white indicates the proteins were not measured. For the breast cancer cell lines, red indicates the protein was identified by proteomic profiling and white indicates the protein was not identified. Unless otherwise indicated, levels were measured by LC-MS. *measured by Luminex/Rules Based Medicine. **measured by both LC-MS and Luminex/Rules Based Medicine.
Of the 49 plasma proteins that increased in relative abundance from mice with preclinical neoplasia, 22 were identified in proteomic profiling experiments from three human breast cancer cell lines, MCF7, MDA-MB-231, and SKBR3 (Supplementary Table S2). SKBR3 cells overexpress ErbB2 and have been utilized extensively to study ErbB2 signaling (27, 28). Immunohistochemical (IHC) staining of preclinical tumor sections showed cytoplasmic and/or nuclear localization in tumor cells for all five proteins for which antibodies were available (Fig. 3) suggesting that release from tumor cell turnover could contribute to increased plasma levels of these proteins. Calreticulin (Calr) is a calcium binding chaperone protein reported to have prognostic significance in breast cancer based on tissue expression (29). Galectin-1 (Lgals1) is induced by ErbB2 (30), binds to Hras and mediates cell transformation (31). Lactotransferrin (Ltf) is an iron-binding protein that is secreted by breast tissue. Calgranulin B (S100A9) is a calcium binding protein present in a wide range of cells and overexpressed in neutrophils, monocytes, and tumor cells (32). The Pdz and lim domain protein (Pdlim1) associates with actin filaments and stress fibers.
Figure 3. IHC staining of selected proteins identified in plasma in normal breast and preclinical breast lesions.
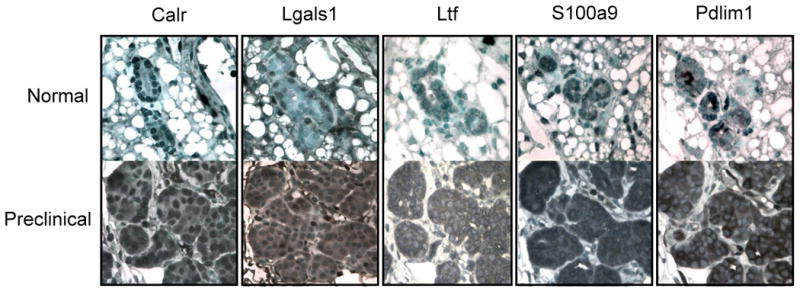
Calreticulin (Calr) shows some nuclear staining in normal breast epithelial and strong nuclear and cytoplasmic staining in preclinical tumor cells. Galectin-1 (Lgals1) shows some nuclear staining in normal and strong nuclear and cytoplasmic staining in preclinical tumor cells. Lactotransferrin (Ltf) shows some cytoplasmic staining in normal and strong cytoplasmic staining in tumor cells. Calgranulin B (S100a9) shows some nuclear staining in normal and strong nuclear and cytoplasmic staining in tumor cells. Pdz and lim domain protein 1 (Pdlim1) shows some cytoplasmic staining in normal and strong cytoplasmic staining in the tumor cells. 60× magnification.
To determine the extent to which protein changes detected at this early time point may be specifically associated with tumor development, we cross referenced this data to similar plasma proteomic profiling from mouse models of subacute and chronic inflammation and angiogenesis (33). Of the 49 proteins with increased concentration in plasma from mice with early stage breast cancer, 12 were increased in the plasma of one or more of the confounding mouse models (Supplementary Table S2), indicating these 12 proteins are not breast cancer specific. Levels of S100a9, Pdlim, Ltf, Calr, and Lgals1 that were detected in primary tumors by IHC (Fig. 2) were not increased in the confounder models, thereby increasing their potential cancer specificity (see Supplementary Table S2 for additional examples). Interestingly, a greater percentage (∼50%) of proteins with decreased concentration in preclinical plasmas were decreased in confounding condition models indicating reduced proteins are less likely to be cancer specific.
Thus, even at a relatively early stage of tumor development, changes were observed in the plasma proteome reflecting contributions from tumor cells, the microenvironment, and the host response. Proteins involved in functions relevant to wound healing, inflammation, metabolism, and ECM remodeling predominated.
Plasma proteome changes associated with tumor progression
To identify plasma protein changes associated with tumor progression, we profiled the plasma proteome of mice bearing 0.5 cm and 1 cm breast tumors. The overall numbers of proteins that showed altered plasma levels in case vs. control mice progressively increased with tumor growth (Fig. 1C). In mice bearing 0.5 cm tumors, 38 proteins showed a significant increase and 105 a decrease in abundance, while in mice bearing 1.0 cm tumors, 70 proteins were increased and 114 were decreased (>1.5, p<0.05) (Supplementary Table S2). As in the preclinical samples, proteins that increased in mid and late stage tumor-bearing mice included those identified in proteomic profiling of human breast cancer cells, as well as proteins involved in the acute phase response, complement, and coagulation cascades, chemokines, macrophage and neutrophil signaling and metabolic proteins (Fig. 2 and Supplementary Fig. S2). A number of cytoskeletal, extracellular matrix, and stromal associated proteins including Loxl1, Tnc, Mmp9, Timp1, Dsg2, Fbln2, Col15a1, Pxdn, Prg4, and Vwf increased in their levels. Interestingly only 1/30 of these tissue remodeling proteins was identified as increased in plasma from the confounder mouse models (Fig. 2), suggesting a substantial contribution of the stroma to plasma protein changes associated with tumor development.
It is noteworthy that a greater proportion of plasma proteins were reduced in abundance, rather than increased in plasma from mice with mid and advanced stage tumors (Fig. 1C). Decreased proteins that were unique to tumor-bearing mice included metabolic proteins and ten components of the proteasome (Psma1, Psma2, Psma4, Psma7, Psmb1, Psmb10, Psmb6, Psmc5, Psmd4, and Psme1) (Supplementary Table S2). Approximately a quarter of the proteins with reduced levels in mice with clinically apparent breast cancer were decreased in plasma from mice with inflammation or angiogenesis (Fig. 4). These data indicate that decreased protein abundance associated with cancer can be attributed to metabolic changes, reduced proteasomal components, as well as host responses that are not cancer specific.
Figure 4. Correlation of plasma protein abundance between tumor stages.

Shown are case/control ratios for plasma proteins that were quantified in both 0.5 and 1.0 cm tumor stages. Green and red data points indicate proteins identified as increased or decreased in the plasma from the confounder models, respectively.
We next examined the extent and potential contribution of altered gene expression in both primary tumor and non tumor host tissues to changes in the plasma proteome. Gene expression profiling was performed on blood buffy coat, thymus, spleen, liver, and breast tumors from mice with 1 cm tumors, using an independent cohort of case and control mice (34). Of the 31 proteins that were increased in plasma from tumor-bearing mice and identified in proteomic profiling of breast cancer cells, 25 (81%) showed a corresponding increase in mRNA expression in HER2/neu breast tumors as compared to normal breast tissue (Supplementary Fig. S3). Ten of these proteins (Flna, Vasp, Cnn2, Coro1a, Cap1, Pxdn, Dsg2, Tpm4, Pfn1, Cotl1, Zyx, and Ctsd) are involved in cytoskeletal function or ECM remodeling, processes that are necessary for tumor cell invasion. None of these proteins increased in concentration in plasma from the confounder models. The concordance between protein levels in plasma, protein expression in breast cancer cells, and mRNA expression in HER2/neu-driven tumors for these proteins indicates a contribution of tumor cells to their increased levels in plasma.
To determine the contributions of the tumor microenvironment to increased levels of proteins in plasma, we used IHC to examine the cellular and stromal landscape of HER2/neu mammary tumors. In addition to tumor cells, HER2/neu tumors contained macrophage, neutrophil, and lymphocyte infiltrates, fibroblasts, extensive collagen deposition, and prominent blood vessels (Fig. 5). Plasma proteins that could be derived from tumor cell interactions with these recruited cell types and the local environment are also displayed according to function (immunity, ECM, cytoskeletal, and angiogenesis). Note the examples of Timp1 and Vasp that were elevated in plasma and expressed in tumor cells.
Figure 5. HER2/neu mammary tumor microenvironment schematic.
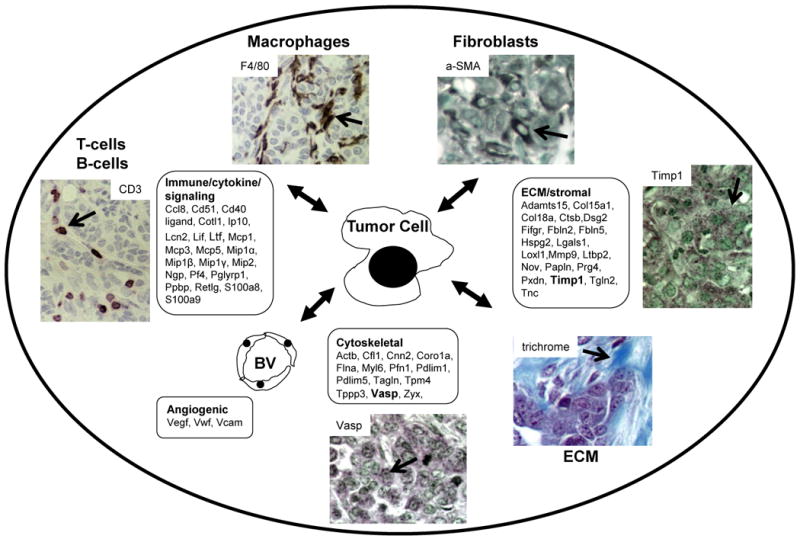
In addition to the tumor cells, Her2/neu tumors contain cancer associated macrophages (F4/80), cancer associated fibroblasts (α-smooth muscle actin (aSMA)), lymphocytes (CD3), blood vessels (not shown), and extensive collagen deposition (trichrome). Also shown are IHC results of HER2/neu tumors for Timp1 and Vasp which were elevated in plasma from tumor-bearing mice. Additional proteins elevated in plasma from tumor-bearing mice are listed according to functional categories (boxes).
For 14 proteins that were increased in plasma of tumor-bearing mice and that are known to be involved in myeloid cell function, we compared the corresponding mRNA expression levels in thymus, spleen and blood buffy coat fractions which are rich in white blood cells. Of these 14 proteins, 13 had increased levels of mRNA expression levels in one or more of these tissues from tumor-bearing vs. control mice (Supplementary Fig. S4). Lipocalin 2 (Lcn2), S100a9, S100a8, neutrophilic granule protein (Ngp), myeloperoxidase (Mpo), and peptidoglycan recognition protein (Pglyrp1) were the most significantly increased in both plasma and immune cell compartments, suggesting the increased levels of these proteins in plasma reflect an immune cell response to the developing tumor. S100a9 and Lcn2 are examples of proteins identified as increased in both immune cells and tumors. A set of 27 genes showed significantly increased mRNA expression in the thymus in case vs. control mice (p<0.05) (34). Proteins corresponding to seven of the 27 genes were increased in the plasma suggesting a contribution of distal immunologic tissues to their increased plasma levels.
Reversion of the plasma proteome toward baseline after oncogene de-induction and tumor regression
To identify plasma protein changes associated with early stage tumor regression, we quantitated protein changes in plasma from mice three days after doxycycline withdrawal and oncogene de-induction (Fig. 1). At this time point only 24 proteins had increased levels and 28 decreased levels in case vs. control mice (Supplementary Table S2). Many proteins increased in tumor-bearing mice were undetected or returned to baseline levels after oncogene de-induction. Of the twelve that remained elevated, eight were immune cell or acute phase proteins (Hp, C8g, Ccl8, EG214403, Mpo, Cd40l, Orm3, and Hpx). Changes in acute phase proteins and complement have also been observed in plasma from epirubicin/docetaxel treated breast cancer (35).
Comparative analysis of the plasma proteome at different tumor stages
Comparison of the plasma proteome from mice with 0.5 cm and 1.0 cm tumors revealed a significant positive correlation (Pearson correlation coefficient=0.67) in relative protein abundance indicating many of the same proteins are either reduced or increased at both tumor stages (Fig. 4). Some proteins were quantified at all four tumor stages. Papln, Ltf, Igfbp5, S100a8, Lrg1, Pcsk9, and Ccl8 are examples of proteins that showed progressive increases in relative abundance during tumor progression and diminished to near control levels after tumor regression (Fig. 6). Of these, Ltf and Igfbp5 were expressed in tumor cells as assessed by IHC. A set of proteins progressively decreased in their levels during tumor development, but rebounded after tumor regression (Fig. 6). These proteins are mainly involved in acute phase response and many were also decreased in the confounder inflammation models. Pathway analysis was used to assess the biologic processes behind these dynamic changes in the plasma proteome that accompanied tumor development. Proteins involved in wound healing, the complement pathway, and metabolism were altered at multiple tumor stages (Supplementary Fig. S2).
Figure 6. Dynamics of protein changes with tumor establishment, progression, and regression.
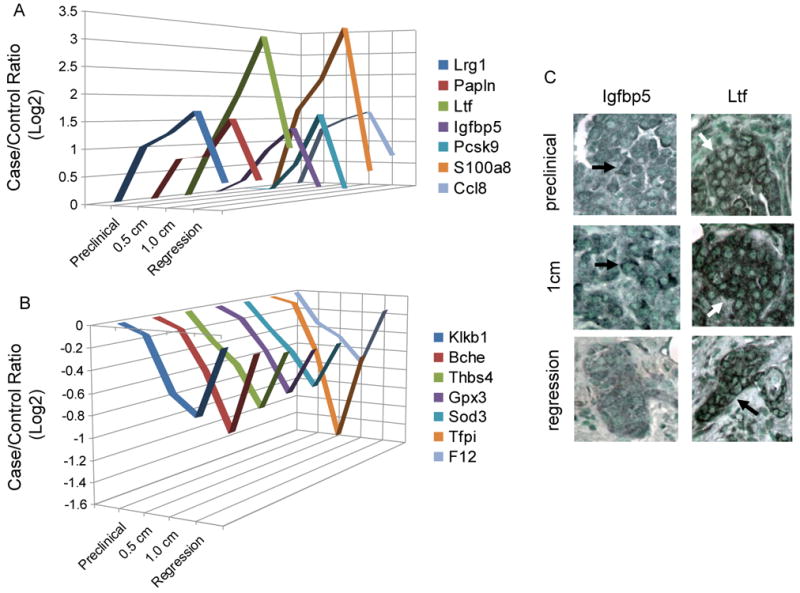
(A) Proteins increased with tumor development and decreased after regression. (B) Proteins decreased with tumor development and increased after regression. (C) IHC revealed Ltf and Igfbp5 staining in tumor cells.
Confirmation of LC-MS/MS detected changes in protein abundance by independent antibody-based methods
Four proteins that span a wide range of concentrations in normal plasma and that exhibited altered plasma levels in tumor-bearing mice based on LC-MS/MS analysis, were subjected to validation using ELISAs applied to samples from independent cohorts of mice. Protein concentrations of Igfbp5, Lcn2, Timp1, and Ccl8 were found to progressively increase during tumor development and return to normal after regression, concordant with MS data (Fig. 7). We also quantified levels of 37 proteins using bead-based assays applied to plasma samples from all four tumor stages. Thirteen of these showed increased levels in case vs. control mice for at least one tumor stage (Supplementary Table S3). These included Cd40 ligand, monocyte and macrophage signaling proteins, and stromal proteins Mmp-9, Timp1, and Vwf. Seven of these proteins were quantified by LC-MS/MS and again there was substantial agreement between the two measurement platforms (Supplementary Fig. S5). Thus, two different antibody-based methods confirmed the LC-MS/MS results. Collectively these results illuminate the dynamic changes in the plasma proteome that track tumor evolution.
Figure 7. ELISA protein measurements in an independent mouse cohort.

Plasma levels of (A) Insulin-like growth factor binding protein 5 (Igfbp5), (B) Lipocalin-2 (Lcn2), (C) Metallopeptidase inhibitor-1 (Timp1), and (D) C-C motif chemokine 8 (Ccl8) as determined by ELISA in a set of mouse plasmas independent of those used for LC-MS/MS experiments. For each time point n=6 controls and n=6 cases. Statistical significance was determined by two-tailed Student's t-test *p<0.05, **p<0.01, and ***p<0.0001.
Discussion
The cancer associated plasma proteome is derived from the developing tumor, the tumor microenvironment, and host tissues
In this study, we present a dynamic analysis of the plasma proteome that accompanies HER2/neu-induced mammary tumor establishment, malignant progression, and regression. Our findings show that the plasma proteome increasingly deviates from normal as tumors develop and progress. This presented as an escalation in the total number of altered proteins as well as a progressive increase or decrease in the relative abundance of a number of specific proteins with advancing tumor stage. Most of these tumor associated protein changes rapidly reverted upon oncogene down-regulation and incipient tumor regression. Thus, the plasma proteome responds very early, i.e. shortly after tumor induction, changes both qualitatively and quantitatively during tumor progression, and rapidly reverts to normal following oncogene down-regulation and tumor regression.
We used several methods to ascertain the specificity and tissue(s) of origin of the plasma protein signature that occurred in the context of breast tumor development. First, comparisons with mouse models of inflammation and angiogenesis were used to identify those proteins that were not cancer specific. Approximately 25% of the elevated and 50% of the reduced proteins overlapped between tumor and confounder plasma proteomes. This indicates that the proteins that decrease in abundance in tumor-bearing mice are more likely attributable to a host response that is not specific to cancer. In the future, analysis of other confounder models such as wound healing or tissue specific inflammatory conditions will be useful to further refine the cancer specific signature. Second, we compared plasma proteins identified in tumor-bearing mice to proteomic and gene expression analyses of human breast cancer cells and HER2/neu-induced breast tumors. Among all the proteins that were increased in plasma from tumor-bearing mice, over half were identified in the proteomic profiling of human breast cancer cell lines, and approximately 80% of this concordant set showed a corresponding increase in gene expression in HER2/neu breast tumors. Third, a subset of those proteins found to be altered in the plasma of tumor-bearing mice was expressed in tumor cells as detected by IHC. These findings point to the tumor itself as the likely source of a subset of proteins found to be altered in plasma. In addition to the secretion of proteins by tumor cells, tumor cell lysis may also lead to detectable levels of cancer-specific proteins in the blood. Finally, microarray gene expression analysis of known inflammatory proteins identified a correlation between protein levels in plasma and gene expression in distal immunologic tissues such as thymus and spleen. Collectively, these findings demonstrate that changes in the plasma proteome that arise during cancer development derive from multiple sources: non-specific systemic reactions, e.g. acute phase proteins such as those produced by the liver; other host responses that are more specific to breast cancer, but not directly tumor-derived, e.g. proteins involved in immune cell recruitment and signaling, and metabolic pathways; and proteins that originate directly from the tumor and the local microenvironment (Fig. 5). Some proteins are expressed in multiple tissues (e.g. S100a9, Lcn2, metabolic and other housekeeping proteins) and unambiguous ascertainment of tissue of origin is more difficult.
Plasma proteome dynamics reflect breast cancer biology
After only five weeks of HER2/neu oncogene activity, extensive neoplastic changes were noted throughout the mammary ductal tree. Although the lesions were clinically undetectable, significant changes in the plasma proteome were already apparent, and these reflected acute phase responses, macrophage and neutrophil trafficking, ECM remodeling, metabolism, and potentially tumor-derived proteins. A significant fraction of these early plasma proteome alterations were not seen in models of inflammation and so are more likely to be associated with early stage cancer. A number of the candidate tumor-derived proteins, such as Igfbp5, Ltf, Timp1, Lgals1, Mmp9, Vwf, Calr, and Pdlim1, that were increased in preclinical plasma were also seen in tumor sections and/or breast cancer cell lines based on IHC or gene expression changes. A subset of these was confirmed to be elevated in plasma from other cohorts of mice and by independent antibody-based methods. Further, the plasma levels of some of these proteins increased with tumor size and rapidly fell after oncogene de-induction indicating they effectively track tumor growth. This establishes both the sensitivity of our method to detect tumor associated proteins very early in the neoplastic process, and also the ability to monitor protein levels over the course of tumor development and during tumor regression.
Notable histopathological changes that occurred during tumor progression included extensive remodeling of the tumor microenvironment, in particular recruitment of immune cells, including T and B cells, macrophages, and neutrophils. This presentation resembles a wound bed and implies that the tumors actively attract these cells from the circulation. In support of this conclusion, plasma levels of multiple macrophage signaling proteins, including monocyte chemoattractant proteins (Mcps), macrophage inflammatory proteins (Mips), and other chemokines, calgranulins, and lipocalin 2, were increased. Many of these proteins were elevated even at the earliest tumor stage and progressively increased in abundance with tumor growth. Once recruited to the tumor site, tumor associated macrophages (TAMs) secrete Mmp2, Mmp9, TGFβ, cathepsins, and other factors that degrade and remodel matrix (36) and promote angiogenesis, tumor cell invasion, and metastasis (37). Our analysis showing elevated plasma levels of Mmp9, Ctsb, and numerous stromal components associated with TAMs extends the reach of this network to the organismal level.
Cancer associated fibroblasts (CAFs) are another significant component of the HER2/neu mammary tumor microenvironment (Fig. 5). These “activated” fibroblasts express smooth muscle actin (38) and support tumor growth, deposit ECM, and promote an inflammatory response by secreting factors such as collagens, tenascin C, matrix metalloproteinases, and chemokines (2, 3). Tumor cells, in turn, must reorganize cytoskeletal structures, break down the basement membrane, and degrade ECM components in order to invade, intravasate, and metastasize. The levels of many proteins related to these processes, including ECM, stromal, and cytoskeletal proteins, were increased in plasma, particularly from mice with late stage tumors. Together this shows that, in addition to non-specific host response proteins and tumor-derived proteins, alterations in signaling and effector pathways regulating tumor/TAM and tumor/CAF interactions are prominent features of the cancer associated plasma proteome. These features bear similarities to a wound response (39), which has recently been described to parallel tumor initiation (40), and indicate the extent to which the tumor co-opts these host responses. This establishes the plasma proteome as a sensitive biomonitor of the ongoing interaction between the developing tumor and the host.
Altered plasma levels of enzymes involved in glucose, lipid, and amino acid metabolism were observed at all three tumor stages. Liver transcriptional profiling identified altered expression of similar metabolic enzymes in case vs. control mice (34) indicating the presence of a tumor triggers systemic metabolic dysregulation, and this initiates very early during tumor establishment. It will be interesting to determine the mechanistic similarities between this response and cachexia which is an altered systemic metabolic response to the presence of late stage cancer (41).
In summary, this study highlights the value of integrating in-depth proteomic, transcriptomic, and tissue-wide approaches as a means to illuminate the systems biology of cancer. The identification of plasma protein networks and transcriptional responses in distal normal tissues of tumor-bearing mice indicates the host senses and responds to neoplastic growth to a greater degree than previously appreciated. The developing tumor simultaneously engages a number of innate physiological processes including wound repair, the immune system, the blood coagulation and complement cascades, and metabolic homeostasis. This extensive conversation initiates very early, involves many cell types and tissues, accurately tracks tumor development over time, and mirrors the underlying biology. Further mining of the cancer associated plasma proteome and other host tissue responses will likely identify additional regulators, mediators, and biomarkers of the evolving tumor, its microenvironment, and the systemic response to cancer. Extension of this approach to other models of cancer as well as human studies will establish its utility for early detection, diagnosis, and prognosis.
Supplementary Material
Acknowledgments
We thank the Allen Foundation Biomarker Project team, Drs. Aebersold, Anderson, Diamandis, and Smith for advice.
GRANT SUPPORT: This work was funded by the Paul G. Allen Family Foundation, NCI Mouse Models of Human Cancer Consortium, and Canary Foundation.
References
- 1.Polyak K, Kalluri R. The role of the microenvironment in mammary gland development and cancer. Cold Spring Harb Perspect Biol. 2010;2:a003244. doi: 10.1101/cshperspect.a003244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kalluri R, Zeisberg M. Fibroblasts in cancer. Nat Rev Cancer. 2006;6:392–401. doi: 10.1038/nrc1877. [DOI] [PubMed] [Google Scholar]
- 3.Mueller MM, Fusenig NE. Friends or foes - bipolar effects of the tumour stroma in cancer. Nat Rev Cancer. 2004;4:839–49. doi: 10.1038/nrc1477. [DOI] [PubMed] [Google Scholar]
- 4.Grivennikov SI, Greten FR, Karin M. Immunity, inflammation, and cancer. Cell. 2010;140:883–99. doi: 10.1016/j.cell.2010.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.de Visser KE, Eichten A, Coussens LM. Paradoxical roles of the immune system during cancer development. Nat Rev Cancer. 2006;6:24–37. doi: 10.1038/nrc1782. [DOI] [PubMed] [Google Scholar]
- 6.Dunn GP, Old LJ, Schreiber RD. The immunobiology of cancer immunosurveillance and immunoediting. Immunity. 2004;21:137–48. doi: 10.1016/j.immuni.2004.07.017. [DOI] [PubMed] [Google Scholar]
- 7.Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation. Nature. 2008;454:436–44. doi: 10.1038/nature07205. [DOI] [PubMed] [Google Scholar]
- 8.Hiratsuka S, Watanabe A, Sakurai Y, Akashi-Takamura S, Ishibashi S, Miyake K, et al. The S100A8-serum amyloid A3-TLR4 paracrine cascade establishes a pre-metastatic phase. Nat Cell Biol. 2008;10:1349–55. doi: 10.1038/ncb1794. [DOI] [PubMed] [Google Scholar]
- 9.Kaplan RN, Riba RD, Zacharoulis S, Bramley AH, Vincent L, Costa C, et al. VEGFR1-positive haematopoietic bone marrow progenitors initiate the pre-metastatic niche. Nature. 2005;438:820–7. doi: 10.1038/nature04186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wels J, Kaplan RN, Rafii S, Lyden D. Migratory neighbors and distant invaders: tumor-associated niche cells. Genes Dev. 2008;22:559–74. doi: 10.1101/gad.1636908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Faca VM, Song KS, Wang H, Zhang Q, Krasnoselsky AL, Newcomb LF, et al. A mouse to human search for plasma proteome changes associated with pancreatic tumor development. PLoS Med. 2008;5:e123. doi: 10.1371/journal.pmed.0050123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hanash SM, Pitteri SJ, Faca VM. Mining the plasma proteome for cancer biomarkers. Nature. 2008;452:571–9. doi: 10.1038/nature06916. [DOI] [PubMed] [Google Scholar]
- 13.Kelly-Spratt KS, Kasarda AE, Igra M, Kemp CJ. A mouse model repository for cancer biomarker discovery. J Proteome Res. 2008;7:3613–8. doi: 10.1021/pr800210b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hung KE, Faca V, Song K, Sarracino DA, Richard LG, Krastins B, et al. Comprehensive proteome analysis of an Apc mouse model uncovers proteins associated with intestinal tumorigenesis. Cancer Prev Res (Phila Pa) 2009;2:224–33. doi: 10.1158/1940-6207.CAPR-08-0153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Moody SE, Sarkisian CJ, Hahn KT, Gunther EJ, Pickup S, Dugan KD, et al. Conditional activation of Neu in the mammary epithelium of transgenic mice results in reversible pulmonary metastasis. Cancer Cell. 2002;2:451–61. doi: 10.1016/s1535-6108(02)00212-x. [DOI] [PubMed] [Google Scholar]
- 16.Cardiff RD, Anver MR, Gusterson BA, Hennighausen L, Jensen RA, Merino MJ, et al. The mammary pathology of genetically engineered mice: the consensus report and recommendations from the Annapolis meeting. Oncogene. 2000;19:968–88. doi: 10.1038/sj.onc.1203277. [DOI] [PubMed] [Google Scholar]
- 17.Pitteri SJ, Faca VM, Kelly-Spratt KS, Kasarda AE, Wang H, Zhang Q, et al. Plasma proteome profiling of a mouse model of breast cancer identifies a set of up-regulated proteins in common with human breast cancer cells. J Proteome Res. 2008;7:1481–9. doi: 10.1021/pr7007994. [DOI] [PubMed] [Google Scholar]
- 18.Rauch A, Bellew M, Eng J, Fitzgibbon M, Holzman T, Hussey P, et al. Computational Proteomics Analysis System (CPAS): an extensible, open-source analytic system for evaluating and publishing proteomic data and high throughput biological experiments. J Proteome Res. 2006;5:112–21. doi: 10.1021/pr0503533. [DOI] [PubMed] [Google Scholar]
- 19.Craig R, Beavis RC. TANDEM: matching proteins with tandem mass spectra. Bioinformatics. 2004;20:1466–7. doi: 10.1093/bioinformatics/bth092. [DOI] [PubMed] [Google Scholar]
- 20.MacLean B, Eng JK, Beavis RC, McIntosh M. General framework for developing and evaluating database scoring algorithms using the TANDEM search engine. Bioinformatics. 2006;22:2830–2. doi: 10.1093/bioinformatics/btl379. [DOI] [PubMed] [Google Scholar]
- 21.Kersey PJ, Duarte J, Williams A, Karavidopoulou Y, Birney E, Apweiler R. The International Protein Index: an integrated database for proteomics experiments. Proteomics. 2004;4:1985–8. doi: 10.1002/pmic.200300721. [DOI] [PubMed] [Google Scholar]
- 22.Keller A, Nesvizhskii AI, Kolker E, Aebersold R. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal Chem. 2002;74:5383–92. doi: 10.1021/ac025747h. [DOI] [PubMed] [Google Scholar]
- 23.Faca V, Coram M, Phanstiel D, Glukhova V, Zhang Q, Fitzgibbon M, et al. Quantitative analysis of acrylamide labeled serum proteins by LC-MS/MS. J Proteome Res. 2006;5:2009–18. doi: 10.1021/pr060102+. [DOI] [PubMed] [Google Scholar]
- 24.Faca VM, Ventura AP, Fitzgibbon MP, Pereira-Faca SR, Pitteri SJ, Green AE, et al. Proteomic analysis of ovarian cancer cells reveals dynamic processes of protein secretion and shedding of extra-cellular domains. PLoS One. 2008;3:e2425. doi: 10.1371/journal.pone.0002425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ong SE, Mann M. A practical recipe for stable isotope labeling by amino acids in cell culture (SILAC) Nat Protoc. 2006;1:2650–60. doi: 10.1038/nprot.2006.427. [DOI] [PubMed] [Google Scholar]
- 26.Bailey SL, Gurley KE, Hoon-Kim K, Kelly-Spratt KS, Kemp CJ. Tumor suppression by p53 in the absence of Atm. Mol Cancer Res. 2008;6:1185–92. doi: 10.1158/1541-7786.MCR-07-2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Le XF, Arachchige-Don AS, Mao W, Horne MC, Bast RC., Jr Roles of human epidermal growth factor receptor 2, c-jun NH2-terminal kinase, phosphoinositide 3-kinase, and p70 S6 kinase pathways in regulation of cyclin G2 expression in human breast cancer cells. Mol Cancer Ther. 2007;6:2843–57. doi: 10.1158/1535-7163.MCT-07-0109. [DOI] [PubMed] [Google Scholar]
- 28.Yang S, Raymond-Stintz MA, Ying W, Zhang J, Lidke DS, Steinberg SL, et al. Mapping ErbB receptors on breast cancer cell membranes during signal transduction. J Cell Sci. 2007;120:2763–73. doi: 10.1242/jcs.007658. [DOI] [PubMed] [Google Scholar]
- 29.Lwin ZM, Guo C, Salim A, Yip GW, Chew FT, Nan J, et al. Clinicopathological significance of calreticulin in breast invasive ductal carcinoma. Mod Pathol. 2010 doi: 10.1038/modpathol.2010.173. [DOI] [PubMed] [Google Scholar]
- 30.Mackay A, Jones C, Dexter T, Silva RL, Bulmer K, Jones A, et al. cDNA microarray analysis of genes associated with ERBB2 (HER2/neu) overexpression in human mammary luminal epithelial cells. Oncogene. 2003;22:2680–8. doi: 10.1038/sj.onc.1206349. [DOI] [PubMed] [Google Scholar]
- 31.Paz A, Haklai R, Elad-Sfadia G, Ballan E, Kloog Y. Galectin-1 binds oncogenic H-Ras to mediate Ras membrane anchorage and cell transformation. Oncogene. 2001;20:7486–93. doi: 10.1038/sj.onc.1204950. [DOI] [PubMed] [Google Scholar]
- 32.Gebhardt C, Nemeth J, Angel P, Hess J. S100A8 and S100A9 in inflammation and cancer. Biochem Pharmacol. 2006;72:1622–31. doi: 10.1016/j.bcp.2006.05.017. [DOI] [PubMed] [Google Scholar]
- 33.Kelly-Spratt KS, Pitteri SJ, Gurley KE, Liggitt D, Chin A, Kennedy J, et al. Plasma proteome profiles associated with inflammation, angiogenesis, and cancer. 2011 doi: 10.1371/journal.pone.0019721. In Press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Schoenherr RM, Kelly-Spratt KS, Lin C, Whiteaker JR, Liu T, Holzman T, et al. Proteome and transcriptome profiles of a Her2/Neu-driven mouse model of breast cancer. Proteomics Clin Appl. 2011;5:179–88. doi: 10.1002/prca.201000037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Michlmayr A, Bachleitner-Hofmann T, Baumann S, Marchetti-Deschmann M, Rech-Weichselbraun I, Burghuber C, et al. Modulation of plasma complement by the initial dose of epirubicin/docetaxel therapy in breast cancer and its predictive value. Br J Cancer. 2010;103:1201–8. doi: 10.1038/sj.bjc.6605909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kessenbrock K, Plaks V, Werb Z. Matrix metalloproteinases: regulators of the tumor microenvironment. Cell. 2010;141:52–67. doi: 10.1016/j.cell.2010.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Qian BZ, Pollard JW. Macrophage diversity enhances tumor progression and metastasis. Cell. 2010;141:39–51. doi: 10.1016/j.cell.2010.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lazard D, Sastre X, Frid MG, Glukhova MA, Thiery JP, Koteliansky VE. Expression of smooth muscle-specific proteins in myoepithelium and stromal myofibroblasts of normal and malignant human breast tissue. Proc Natl Acad Sci U S A. 1993;90:999–1003. doi: 10.1073/pnas.90.3.999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Troester MA, Lee MH, Carter M, Fan C, Cowan DW, Perez ER, et al. Activation of host wound responses in breast cancer microenvironment. Clin Cancer Res. 2009;15:7020–8. doi: 10.1158/1078-0432.CCR-09-1126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Feng Y, Santoriello C, Mione M, Hurlstone A, Martin P. Live Imaging of Innate Immune Cell Sensing of Transformed Cells in Zebrafish Larvae: Parallels between Tumor Initiation and Wound Inflammation. PLoS Biol. 2010;8:e1000562. doi: 10.1371/journal.pbio.1000562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tayek JA. A review of cancer cachexia and abnormal glucose metabolism in humans with cancer. J Am Coll Nutr. 1992;11:445–56. doi: 10.1080/07315724.1992.10718249. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.