

Abstract
Background
The Tec family kinases are implicated in signaling from lymphocyte antigen receptors and are activated following phosphorylation by Src kinases. For most Tec kinases, this activation requires an interaction between their pleckstrin homology (PH) domains and the products of phosphoinositide 3-Kinase, which localizes Tec kinases to membrane RAFTs. Rlk/Txk is a Tec related kinase expressed in T cells that lacks a pleckstrin homology domain, having instead a palmitoylated cysteine-string motif. To evaluate Rlk's function in T cell receptor signaling cascades, we examined the requirements for Rlk localization and activation by Src family kinases.
Results
We demonstrate that Rlk is also associated with RAFTs, despite its lack of a pleckstrin homology domain. Rlk RAFT association requires the cysteine-string motif and is independent of PI3 Kinase activity. We further demonstrate that Rlk can be phosphorylated and activated by Src kinases, leading to a decrease in its half-life. A specific tyrosine in the activation loop of Rlk, Y420, is required for phosphorylation and activation, as well as for decreased stability, but is not required for lipid RAFT association. Mutation of this tyrosine also prevents increased tyrosine phosphorylation of Rlk after stimulation of the T cell receptor, suggesting that Rlk is phosphorylated by Src family kinases in response to T cell receptor engagement.
Conclusions
Like the other related Tec kinases, Rlk is associated with lipid RAFTs and can be phosphorylated and activated by Src family kinases, supporting a role for Rlk in signaling downstream of Src kinases in T cell activation.
Background
Engagement of antigen receptors on lymphocytes leads to the rapid sequential activation of non-receptor tyrosine kinases, including well-studied members of the Src and the ZAP-70/Syk families [1,2]. Recently, the Tec kinases have also been implicated as important components of signaling from antigen and other lymphocyte cell surface receptors (reviewed in [3,4,5].) This subfamily includes six members: Btk, Tec, Itk/Tsk/Emt, Bmx/Emt, Dsrc29, and Rlk/Txk [3,6,7]. The importance of the Tec kinases in antigen receptor signaling was first demonstrated by the observation that mutations in BTK result in the human disease X-linked agammaglobulinemia (XLA) and the murine counterpart X-linked immunodeficiency (xid). These severe B cell immunodeficiencies are characterized by diminished numbers of mature B cells and reduced immunoglobulin levels associated with impaired PLC-γ activation and Ca++ mobilization in response to surface IgM stimulation (sIgM) [8,9,10,11]. Btk kinase activity and tyrosine phosphorylation increase upon stimulation of sIgM as well as the IL-5 and IL6 receptors in B cells, and upon crosslinking of the high affinity IgE receptor (FcεRII) in mast cells [12,13,14,15]. It is now clear that the other Tec kinases participate in similar signaling cascades in multiple cell-types. Itk, a Tec family kinase that is predominantly expressed in T cells, is tyrosine phosphorylated and activated after stimulation of either the T cell receptor (TCR) or CD28 in T cell lines, and FcεRII in mast cells [12,16,17,18]. Recently, we have found that Rlk/Txk can also be phosphorylated after TCR engagement [19]. Furthermore, we have found that combined mutation of Rlk and Itk leads to profound defects in TCR signaling associated with impaired activation of PLC-γ [20]. Such data suggest that members of this family may function similarly in signaling pathways downstream from these cell surface antigen receptors.
Several lines of evidence indicate that, in these signaling cascades, Tec family members act downstream of Src kinases. Src kinases are activated within seconds of sIgM stimulation in B cells, while Btk is activated within minutes [14]. Moreover, Src family kinases have been found to phosphorylate and activate Btk, Itk, Tec and Rlk when co-expressed [16,19,21,22,23]. Phosphopeptide mapping and mutational analyses of Btk have identified Y551 in the activation loop of the kinase domain as the major Src-transphosphorylation site. Phosphorylation of Y551 and a putative autophosphorylation site Y223 in the SH3 domain of Btk [23,24] are observed following BCR activation, suggesting that these tyrosine phosphorylations are functionally important for antigen receptor signaling. Although physical binding of full length Btk to Src family kinases in vivo has not been reported, in vitro interactions have been demonstrated between proline-rich motifs in the amino-terminal sequence of Btk and the Src homology (SH) 3 domains of Src family kinases [25,26].
Recently, it has been found that the activation of Btk and Itk by Src family kinases is potentiated by and requires the activity of Phosphoinositide 3-kinase (PI3K), the products of which interact with the PH domains of Tec kinases [27,28]. This interaction helps localize Tec family kinases to the membrane where phosphorylation by Src kinases can occur. Data suggest that, for Itk, this membrane association specifically targets the protein to glycolipid enriched detergent-insoluble membrane microdomains known as "RAFTs" and that both this association and the activation of Itk requires the PH domain [29].
rlk/TXK encodes a kinase expressed in T cells and mast cells that closely resembles members of the Btk/Tec family of kinases [30,31,32]. However, in contrast to the other Tec family members, Rlk lacks a pleckstrin homology domain. Instead, we have found that the rlk gene encodes a distinctive palmitoylated cysteine-string motif near its amino-terminus. Furthermore, we have found that an internal translational start-site, downstream of the region encoding the cysteine-string motif, gives rise to a second isoform that lacks the palmitoylated cysteine string and that can be localized in the nucleus [19]. Although these distinctive features suggest that Rlk may have atypical functions, Rlk shares with Btk kinases the homologous proline-rich motif that binds the SH3 domains of Src kinases and the tyrosines at equivalent sites in the SH3 and kinase domains.
We have recently shown that Rlk can be phosphorylated and activated by the Src family kinase Fyn. However, consistent with the lack of a pleckstrin homology domain, activation of Rlk is independent of PI3K activity [19]. To further characterize the mechanism of activation of Rlk/Txk, we studied the nature of Rlk's localization and interaction with Src family kinases. We report here a physical association of Rlk with Fyn both in vitro and when co-expressed in 293T cells. Similar to Btk and Itk, we find that a conserved tyrosine in the activation loop of the Rlk kinase domain, Y420, mediates specificity for transphosphorylation of Rlk by Src family kinases. This tyrosine is also critical for in vivo activation of Rlk, since mutation of Y420 to phenylalanine (Y420F) reduces phosphorylation of Rlk upon co-expression with Fyn or after engagement of the TCR. Furthermore, we have observed that the phosphorylation of Rlk by Fyn increases Rlk turnover, consistent with the increased turnover observed with other TCR signaling molecules following activation. In contrast, localization of Rlk does not appear to rely on tyrosine phosphorylation or kinase activity of Rlk. Instead, we find a constitutive association of a fraction of the full-length Rlk with a detergent-insoluble RAFT fraction. Association with the RAFT compartment does not depend on tyrosine phosphorylation, nor on PI3K activation, but rather on the presence of the cysteine string motif. Thus, despite lacking a pleckstrin homology domain, Rlk is localized in RAFTs, is phosphorylated by Src family kinases, and may transduce signals downstream of Src family kinases during T cell activation.
Results
Rlk Interacts with Fyn
Rlk/Txk is a Tec related kinase that is predominantly expressed in cells of the T cell lineage [30,31,32]. We have recently demonstrated that Rlk is phosphorylated following stimulation of the Jurkat T cell line by crosslinking of the CD3ε chain of the TCR [19]. We have further observed that Rlk can be phosphorylated and activated by co-expression with the Src family kinase, Fyn, in a PI3K-independent fashion. In contrast, activation of Btk and Itk by Src family kinases is PI3K-dependent. To further characterize the mechanism of Rlk activation by Src family kinases, we examined potential interactions of Rlk with Src kinases both in vitro and in vivo.
The proline-rich region of Btk has been found to bind to SH3 domains of Src family kinses in vitro. To determine whether the amino-terminus of Rlk/Txk interacts with Src family SH3 domains in vitro, we performed in vitro filter binding analyses. GST fusions encoding the Rlk amino-terminal and SH3 domains or the amino-terminus alone (containing the cysteine string and proline-rich region) bound biotinylated GST fusions of the SH3 domains of Fyn, Hck, and Lyn, but not those of Src or Lck, despite the ability of these constructs to bind other proline rich sequences (Fig. 1B and data not shown) [25,33]. The Rlk fusion proteins also failed to efficiently bind SH3 domains from other kinases, including Abl, Csk and Btk. A weaker but detectable binding to the SH3 domain of Grb2, however, was observed. These data agree both with two-hybrid experiments and in vitro filter binding assays that have demonstrated specific binding of the proline-rich region near the amino-terminus of BTK and Tec to the SH3 domains of the Src family kinases Fyn, Lyn and Hck [22,25,26].
Figure 1.

Interactions of Rlk with Src Family kinases. A, Structure of endogenous Rlk protein (top) and Rlk-GST fusion constructs (bottom). Numbers of the constructs correspond to the lanes in part B. B,The amino-terminus of Rlk can bind the SH3 domains of Src family kinases, Fyn, Lyn and Lck. GST fusion proteins encoding the amino-terminus or the amino-terminus and SH3 domain were separated on a 10% SDS-PAGE, transferred to nitrocellulose, then probed with biotinylated GST-fusion proteins encoding the SH3 domains of various signaling proteins. Lane 1 GST-Rlk amino-terminus, Lane 2 GST-Rlk amino-termus-SH3 domain, Lane 3 GST control. SH3 domain fusion proteins used to probe blots are indicated above each panel. C, Co-immunopreciptation of Rlk and Fyn. 293T cells were transfected with expression constructs of Rlk or FynT alone or in combination, and lysed at 24 hours. Equal amounts of detergent-soluble cell lysates were immunoprecipitated with anti-Rlk; immune complexes were washed, resolved by SDS-PAGE and immunoblotted (IB) with anti-Fyn-3 (Santa Cruz) (top panel), then stripped and blotted with anti-Rlk (bottom panel). Fyn expression was assessed by immunoblotting equivalent amounts of cell lysate with anti-Fyn (middle panel). Lane 1: Mock transfection, Lane 2: Rlk WT, Lane 3: Fyn WT, Lane 4: Rlk +Fyn.
Because Fyn is expressed in T cells and Hck and Lyn are not, we pursued the connection between Rlk and the T cell-specific isoform p59FynT. Co-immunoprecipitation demonstrated an interaction between these proteins in extracts of cells in which both Rlk and Fyn were expressed (Fig. 1C). Anti-Rlk serum precipitated Fyn from lysates only when Rlk was expressed (Fig. 1C, lane 4). Likewise, anti-Fyn serum could precipitate Rlk only when both Rlk and Fyn were produced (data not shown). This interaction was not dependent on the activity of either kinase, as co-immunoprecipitation was observed with either wild-type (WT) or kinase-inactive versions of the two proteins. Furthermore, both isoforms of Rlk were also able to co-precipitate with Fyn (data not shown). Finally, we have not observed co-precipitation of full-length Rlk with several other proteins involved in TCR signaling (unpublished observations).
Rlk co-localizes with a portion of Fyn
To further examine potential interactions between Rlk and Fyn we compared the subcellular distribution of Rlk to that of p59FynT. For these and subsequent analyses, we utilized Green Fluorescent Protein (GFP)-tagged versions of Rlk. We have previously demonstrated that these fusion proteins are kinase-active and exhibit localization patterns similar to those observed by indirect immunofluorescence for wild type Rlk . Furthermore, we have found that, like WT Rlk, they co-precipitate with Fyn (data not shown). However, unlike WT Rlk, Rlk-GFP does not co-migrate with immunoglobulin heavy chain. Since these GFP fusions behaved similarly to wildtype Rlk in all known respects except for electrophoretic mobility, we used GFP fusions of mutant and wild type versions of Rlk to facilitate studies of Rlk localization and activation.
We co-transfected Fyn and Rlk-GFP expressing constructs into HeLa cells, and visualized Fyn with anti-Fyn serum and Texas-red conjugated secondary antibody (Fig. 2). Examination of cells expressing both kinases demonstrated that a portion of Fyn co-localized with a portion of the Rlk protein as indicated by the yellow staining in overlapping images (center panel). Thus, Fyn and Rlk can reside in similar compartments within the cell, further supporting a potential interaction between these two molecules.
Figure 2.

Rlk co-localizes with a portion of p59FynT. HeLa cells were transiently co-transfected with constructs expressing Rlk-GFP and wildtype FynT, fixed, stained with anti-Fyn (Fyn3, Santa Cruz) and Texas-Red anti-rabbit antiserum and visualized by fluorescent microscopy after counterstaining with DAPI. (A) Cells visualized for Rlk-GFP. (B) Cells visualized for both Rlk-GFP and Fyn (Texas Red). (C) Cells visualized for Fyn (Texas Red) only.
Fyn phosphorylates and activates Rlk: A Conserved Tyrosine (Y420) Provides Specificity For Phosphorylation of Rlk By Src Kinases
To further examine the consequences of the interaction between Rlk and Src Family kinases, we examined tyrosine phosphorylation of Rlk by Fyn using co-expression studies. As previously demonstrated, co-expression of kinase-active Fyn and Rlk-GFP in 293T cells led to a marked increase in the tyrosine phosphorylation and kinase activity of wild type Rlk-GFP (Fig. 3A, lane 1 and 2; 3B, lane 1 and 2). This was not observed in cells transfected with kinase-inactive Fyn, and the increase in kinase activity on enolase appeared to be due to Rlk rather than Fyn, since we barely detect phosphorylation of enolase when WT Fyn is co-expressed with kinase-inactive Rlk [19]. Thus, the phosphorylation of Rlk appears to cause an increase in Rlk kinase activity.
Figure 3.

Tyrosine 420 of Rlk provides specificity for Fyn transphosphorylation. (A) Wild type Rlk, Rlk-GFPY420F or Rlk-GFPY91F were expressed alone or with wild type Fyn; equal amounts of cell lysates were resolved by SDS-PAGE and subject to immunoblotting with antiphosphotyrosine (4G10, upper panel), or anti-Rlk (α-Rlk, lower panel). (B) In vitro kinase activity of Rlk-GFPY420F. In vitro autokinase activity (middle panel) and in vitro kinase activity on enolase (bottom panel) of anti-Rlk immune complexes; immunoprecipitates of cell lysates expressing the indicated wild type or mutant Rlk with or without WT Fyn were recovered, washed, subject to in vitro kinase assays with acid-denatured enolase as described, resolved by SDS-PAGE and visualized by autoradiography. Levels of Rlk expression are shown in the top panel. Rlk mutants include WT: wild type Rlk-GFP, KI: kinase inactive Rlk-GFP (K299R), and Rlk-GFPY420F. (C) In vitro kinase activity of Rlk-GFP and Rlk-GFPY91F (bottom panel). Levels of Rlk expression are shown in the top panel. (D) Wild type Rlk or Rlk-GFPY6F were expressed alone or with wild type Fyn and immunoblotted with anti-phosphotyrosine (upper panel) or anti-Rlk, (lower panel).
Like other Btk family members, Rlk contains a highly conserved tyrosine (Y420) within a consensus sequence for phosphorylation recognized by Src family tyrosine kinases [23]. This site corresponds to the location of the autophosphorylation site in the activation loop of Src-related kinases [34]. To determine whether this tyrosine is required for Rlk activation by Src family kinases, we mutated it to phenylalanine (Y420F). In contrast to WT Rlk-GFP, tyrosine phosphorylation and kinase activity of the RlkY420F mutant was not significantly increased by co-expression with Fyn (Fig. 3A, B and data not shown). These results suggest that Y420 is a major site of phosphorylation by Fyn and that phosphorylation at this site leads to activation of the Rlk kinase. Furthermore, since baseline tyrosine phosphorylation and kinase activity of the Y420F mutant is also lower than WT (Fig. 3B and data not shown), it is possible that phosphorylation by endogenous Src family kinases may contribute to the activity of Rlk in 293T cells, although we cannot exclude the possibility that this mutant lowers kinase activity for other reasons.
The major autophosphorylation site in Btk maps to Y223 within the SH3 domain [24]; an analogous tyrosine is conserved in Rlk (Y91). Mutation of this tyrosine (Y91F) reduced tyrosine phosphorylation of Rlk approximately two-fold when the Y91F mutant was expressed alone or with Fyn (Fig. 3A). However, Fyn still significantly enhanced the phosphorylation of Rlk (Y91F), consistent with use of an intact transphosphorylation site (Y420) (Fig. 3A). The in vitro kinase activity of Rlk-Y91F was also lower (about half that of wild type); however, the kinase activity was still augmented by the presence of Fyn (Fig. 3C). These results suggest that tyrosine 91 may be required for optimal phosphorylation and kinase activity of Rlk, but is distinct from the major site phosphorylated by Src family kinases.
Finally, since the larger isoform of Rlk consistently exhibits higher levels of tyrosine phosphorylation (see Fig. 3A), we mutated the single tyrosine (Y6) not found on the shorter isoform. Mutation of Y6 did not greatly reduce the tyrosine phosphorylation of Rlk, either in the presence or absence of co-expressed Fyn (Fig. 3D). Significantly, the larger isoform still exhibited a higher level of tyrosine phosphorylation than the shorter form of Rlk. Thus, the increased tyrosine phosphorylation of the larger isoform cannot be attributed solely to a phosphorylation site present on the larger protein species.
Rlk localizes in RAFTs
Since the relative increase in tyrosine phosphorylation of the larger form of Rlk did not result solely from an additional site for tyrosine phosphorylation, it was possible that this increased phosphorylation of the longer isoform of Rlk may result from preferential localization close to Src family kinases. Like other proteins modified by palmitoylation, the Src family kinase Fyn is found localized in RAFTs, a membrane microdomain fraction enriched in signaling molecules that can be isolated by sucrose gradient fractionation. To examine whether Rlk is also associated with RAFTs, we performed sucrose gradiant fractionation of lysates from Jurkat lymphoma cell lines transfected with Rlk-GFP. Upon such fractionation, we observe the Src family kinase Fyn constitutively associated with the RAFT fraction (Fig. 4A). In contrast, PLC-γ was not found in the RAFT fraction constitutively, but a portion became inducibly associated with RAFTs upon activation. Similar to Fyn, a portion of Rlk was also found constitutively associated with RAFTs. Notably, this association consisted primarily of the larger form of Rlk which contains the cysteine-string and is palmitoylated. Furthermore, the association of Rlk with RAFTs only marginally increased upon stimulation through the TCR.
Figure 4.
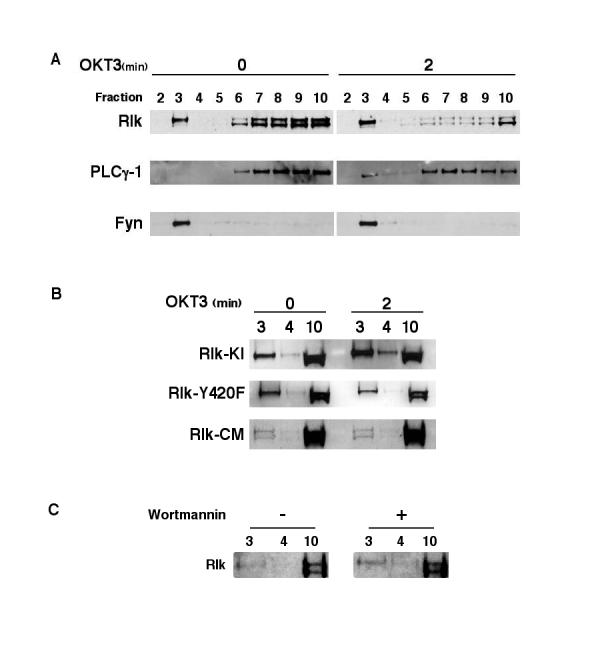
Rlk is associated with Lipid RAFTs: RAFT association does not require tyrosine phosphorylation, but does require the Cysteine-string. (A) Jurkat-Tag cells were electroporated with Rlk-GFP and 24 hours later were either left unstimulated or stimulated with anti-CD3e. Cells were lysed in 1% Triton X-100 and lysates fractionated over a 40%-30%-5% sucrose step gradient for 16 hr at 200,000 g. Collected fractions were separated by SDS-PAGE and analyzed for presence of Fyn, Rlk and PLC-γ1. RAFTs are included in fraction 3 or fractions 3-4. (B) Jurkat-Tag cells were electroporated with either Rlk-GFPKI, Rlk-GFPY420F or Rlk-GFPCM and treated as in (A). Only fractions 3,4 and 10 are shown. For Rlk-GFPCM, a longer exposure is shown to demonstrate the residual RAFT association. (C) Jurkat-Tag cells were electroporated with Rlk-GFP, treated with Wortmannin or carrier (DMSO) and fractionated as above.
Upon TCR signaling, many molecules have been found to show markedly increased association with RAFTs. Since we have found that Rlk can be phosphorylated by Src family kinases leading to activation of its kinase, we evaluated whether phosphorylation could influence RAFT association. Neither mutation of a conserved lysine in the kinase domain, which rendered Rlk kinase inactive, nor mutation of the Src family kinase phosphorylation site, tyrosine 420, prevented association with the RAFT fraction. Thus, phosphorylation and activation of Rlk does not appear to contribute to its association with RAFTs (Fig. 4B).
A key mechanism of targeting proteins to RAFTs is lipid modification. Since the long form of Rlk is palmitoylated on its cysteine string motif and palmitoylated proteins can be associated with RAFTs, we examined the localization of a mutant version of RlkCM in which the cysteine string is mutated, thereby preventing palmitoylation of the long form of Rlk. The cysteine-string mutant demonstrated marked reduction in RAFT association (Fig. 4B). Indeed, while a small amount of the Rlk cysteine-string mutant was still found associated with the RAFT fraction, the amount of the long isoform of RlkCM in the RAFT fraction was equivalent to that of the short isoform. This observation suggests that the cysteine-string may be a major mechanism of targeting full-length Rlk to lipid RAFTs. However, the localization of a small portion of both the short and long isoforms of the cysteine string mutant to RAFTs suggests that other targeting mechanisms may also contribute to the localization of this kinase in RAFTs.
Recent data suggests that both Itk and Btk can also be associated with lipid RAFTs [29,35]. For Itk, this association requires the PH domain and products of PI3K. For example, in the Jurkat lymphoma cell line, which is deficient in the inositol phosphatase PTEN, Itk is constitutively associated with the membrane [36]. Since our RAFT analyses of RLK were performed in Jurkat cells, we also examined the effects of PI3K inhibition on Rlk localization. Treatment of Jurkat cells with Wortmannin had no effect on Rlk association with RAFTs (Fig. 4C). Thus, RAFT localization of Rlk, like its activation by Src family kinases, appears to be independent of the activity of PI3K.
Fyn alters the half life of Rlk in the detergent soluble fraction of transfected cells
In experiments with Rlk, we often observed a change in the relative ratio of the two isoforms of Rlk when co-expressed with Fyn. Furthermore, activation of Jurkats appeared to be associated with a loss of Rlk from non-RAFT fractions (see Fig. 4A). Since phosphorylation of Rlk did not seem to affect association with RAFTs, we examined whether Fyn altered other properties of the Rlk protein. To start to address this issue, we measured the effect of Fyn on the protein stability of Rlk. Rlk was expressed in 293T cells, either in the absence or the presence of Fyn; the cells were labeled for 15 minutes with 35S methionine and cysteine, then incubated for increasing times with unlabelled methionine and cysteine. When expressed alone, labeled Rlk appeared stable in the detergent-soluble fraction for 90 minutes (Fig. 5A, top panel). When co-expressed with Fyn, however, Rlk disappeared from the soluble fraction at a significantly faster rate (Fig. 5A, lower panel, half-life between 30 and 60 minutes). The change in half-life was particularly noticeable for the larger form of Rlk. In the presence of kinase-inactive Fyn, the half-life of Rlk was not altered, suggesting that the phosphorylation of Rlk by Fyn is necessary for the decrease in half-life of Rlk (Fig. 5B). Similarly, co-transfection of wild type Fyn with RlkY420F did not alter the half-life of RlkY420F in the soluble fraction (data not shown), again arguing that phosphorylation of Rlk by Fyn is required for this increased turnover in the soluble fraction.
Figure 5.

Effect of Fyn on the half life of Rlk in the detergent soluble fraction. (A) 293T cells expressing WT Rlk with empty expression vector (top) or with Fyn WT (bottom) were pulse-labeled with Trans-35S-Label for fifteen minutes and chased with cold methionine and cysteine (20 mM each) for the indicated times. Cells were lysed at each time point, and the lysates were immunoprecipitated with anti-Rlk, washed, resolved with SDS-PAGE, and subject to fluorography. (B) Pulse-chase labeling of WT Rlk without (top) or with (bottom) kinase inactive Fyn.
Since this analysis included only Rlk protein in the soluble fractions of detergent lysates, these results may represent changes in protein stability, or subcellular localization and detergent solubility, factors that all can contribute to changes in intracellular signaling. In particular, since Rlk can be associated with detergent-insoluble RAFTs, it is possible that a change in subcellular localization or associations may contribute to the decrease in Rlk protein. To address these possibilities, we performed similar analyses after lysing cells under harsh detergent conditions (using 1% SDS), then diluting the samples before performing immunoprecipitations. Consistent with our observation that the phosphorylation status of Rlk does not significantly contribute to its association with RAFTs, we still observed a decreased half-life of Rlk co-expressed with Fyn when lysed under these harsh conditions (data not shown), arguing that phosphorylation by Fyn specifically accelerates Rlk turnover.
Rlk phosphorylation in response to TCR engagement requires Tyrosine 420
The ability of Src family kinases to activate Rlk and to promote Rlk protein turnover suggests that Rlk may function downstream of Src family kinases in T cell receptor (TCR) signaling. To determine the requirements for Rlk activation downstream from the TCR, we introduced constructs expressing Rlk-GFP or Rlk-GFPY420F into the Jurkat-Tag cell line. Stimulation of the cells with an antibody directed against CD3, OKT3, increased tyrosine phosphorylation of Rlk (Fig. 6). However, phosphorylation of the Y420F mutant was markedly reduced relative to WT Rlk-GFP. These results suggest that Rlk can be phosphorylated on Y420 in response to TCR stimulation, supporting a role for Rlk downstream of Src kinases in signaling from the TCR, analogous to Btk's role in signaling from sIgM.
Figure 6.

Y420 is required for Rlk tyrosine phosphorylation after TCR engagement. Jurkat-Tag cells were electroporated with either WT Rlk-GFP or Rlk-GFPY420F and twenty-four hours later, cells were stimulated with anti-CD3 antibody OKT3 for the indicated times. Cells were lysed and immunoprecipitated Rlk was immunoblotted with anti-phosphotyrosine (4G10) (top panel), or anti-Rlk (bottom panel).
Discussion
By several criteria, Rlk appears to be a member of the Tec family of tyrosine kinases, yet significant differences exist between Rlk and the other kinases in this family. To elucidate functional similarities and differences between Rlk and other Tec kinases in signaling from antigen receptors, we investigated the interactions of Rlk with Src family kinases. Our data suggest that such interactions occur and that the phosphorylation events leading to the activation of Rlk are analogous to those required for the other Tec family kinases. However, our data also suggest that the regulation of Rlk has distinct properties.
First, we have observed an in vitro interaction of the amino-terminus of Rlk with the SH3 domains of Src family kinases. Second, we have demonstrated interactions between these molecules in cells co-expressing both Rlk and the Src family kinase, Fyn. The two kinases can be immunoprecipitated when co-expressed. Moreover, co-expression with Fyn, as well as other Src family kinases (data not shown), leads to increased phosphorylation of Rlk and activation of the Rlk kinase, in agreement with observations made with other Tec family members.
It has been proposed that Src kinases activate Btk through phosphorylation of a tyrosine, Y551 in the activation loop of Btk, leading to increased kinase activity and autophosphorylation of a tyrosine in the SH3 domain. These patterns of Btk phosphorylation have been observed upon cross-linking of IgM, arguing that this pathway is functional in the signal transduction from antigen receptors [23,37,38]. Our mutational analysis of Rlk supports a similar mechanism for activation of Rlk by Src family kinases downstream of the TCR. Rlk kinase activity is increased by Src family kinase-mediated phosphorylation of a conserved tyrosine, Y420, equivalent to Y551 in the activation loop of the kinase domain of Btk. Mutation of this site causes a reduction in tyrosine phosphorylation and kinase activity of Rlk, which is not substantially enhanced upon co-expression with Fyn or upon TCR stimulation. Thus, this Src kinase phosphorylation site appears to be important for activation after TCR stimulation. Mutation of a second tyrosine, equivalent to the putative autophosphorylation site of Btk, slightly reduces tyrosine phosphorylation and kinase activity of Rlk. However, phosphorylation and kinase activity of this mutant still can be enhanced after co-expression with Fyn. Thus, despite its unique amino terminus, Rlk utilizes a transphosphorylation activation mechanism similar to other Btk family members.
Although our original SH3 domain binding experiments suggested a specific interaction between Tec kinases and the Src kinases Fyn, Lyn and Hck, we have been able to observe both activation by and co-precipitation with other Src family kinases including Lck and Src in 293T cells (data not shown). Thus, in vivo, interactions with Rlk may not be restricted to a particular subset of Src family kinases. Indeed, we have found that mutation of the proline-rich region did not abolish Rlk co-precipitation with either Fyn or Lck (data not shown), suggesting that interactions between these kinases may not occur solely through an SH3 mediated interaction, at least in the 293T expression system. Nonetheless, whether there is a specific connection between Rlk and the Src family kinase Fyn in T cells remains an interesting question. Recent data suggests that phosphorylation of the T cell adaptor molecule SLP-76 occurs after stimulation of T cells with altered peptide ligands in a Fyn-dependent manner that is independent of the adaptor molecule LAT [39,40]. Notably, Rlk has also been found to be capable of phosphorylating SLP-76 [41]. It will be of interest to determine whether Rlk participates in this novel activation pathway.
While Rlk is phosphorylated similarly to Btk and Itk, there are marked differences between these two kinases. Perhaps the most obvious of these differences is the lack of the pleckstrin homology domain in Rlk. Data suggest that membrane association of Itk and Btk requires the interaction of their PH domains with products of PI3K, thereby facilitating phosphorylation by Src family kinases [27,28]. In contrast, we have recently demonstrated that PI3K inhibitors do not alter activation of Rlk by Fyn, suggesting that the requirements for activation of Rlk by Src family kinases are distinct from the other Tec kinases. It is also of note that we can reproducibly co-immunoprecipitate Rlk and Fyn, perhaps due to their association with RAFTs, whereas we have been unable to co-precipitate Itk with Src family kinases when co-expressed in 293T cells (data not shown), again demonstrating distinct properties of the Rlk protein compared to certain other members of the Btk family. The combination of similar and distinct features is further supported by the observation that TXK can rescue phospholipase C-γ activation in BTK deficient DT40 cells, but fails to rescue a defect in apoptosis [42].
Interestingly, we also find that the full-length form of Rlk has a higher basal level of tyrosine phosphorylation and kinase activity than Itk when expressed in heterologous cells, consistent with potentially different localizations and upstream interactions (unpublished observations). The full-length isoform of Rlk also exhibits higher tyrosine phosphorylation and kinase activity than the short isoform. This differential phosphorylation does not appear to result solely from a distinct phosphorylation site on the full-length Rlk, since mutation of the only additional tyrosine found in the full-length Rlk (Y6) does not greatly reduce the phosphorylation or activity of Rlk when expressed either alone or in conjunction with Fyn. Instead, the increased phosphorylation may result in part from distinct properties of full-length Rlk. Indeed, we report here that the palmitoylated cysteine-string is responsible for the targeting of Rlk to RAFTs: detergent-insoluble, glycolipid-enriched membrane microdomains that contain many signaling molecules, including Src family kinases. Our data may provide the first demonstration that a cysteine-string motif can lead to protein association with RAFTs. Of note, Itk and Btk have been found to be associated with lipid RAFTs upon activation of antigen receptors [29,35]. However, the association of Itk with RAFTs requires its PH domain and is dependent on the products of PI3K. Indeed, in Jurkat cells, which are deficient in the inositol phophatase PTEN, Itk is constitutively associated with the membrane unless the cells are treated with inhibitors of PI3K or transfected with PTEN [36]. In contrast, we have found that Rlk association with RAFTs, like its activation by Src family kinases, is PI3K independent. Thus, although Rlk, like Itk and Btk, is activated by phosphorylation by Src family kinases and can be found in RAFTs, its requirements for this activation may be unique.
Co-expression of Rlk and Fyn also led to a rapid disappearance of Rlk from the detergent soluble fraction as measured by pulse-chase analysis (Fig. 5). This phenomenon could result from either a change in protein location to a detergent-insoluble fraction or increased turnover of Rlk. Activation of tyrosine kinases and other signal transduction molecules is associated both with changes in subcellular distribution and with changes in protein stability. For example, crosslinking of the T-cell receptor results in a transition of the receptor complex to a detergent-insoluble fraction [43,44,45], as well as to increased turnover of TCR components [46]. Nonetheless, the decreased half-life of Rlk in both detergent-soluble and insoluble fractions argues that activation of Rlk leads to increased degradation of the Rlk protein, similar to that observed for other components of the TCR complex, but not previously reported for Tec family kinases. The dependence of this increased turnover on phosphorylation of Rlk suggests that a phosphotyrosine-dependent association with another molecule may be required for this process, reminiscent of the increased degradation of the PDGF receptor and Syk caused by association with the ring-finger protein Cbl [47,48,49] or the Lck-dependent degradation of the TCR [50].
Recent evidence from gene-targeted mice suggests that Rlk may synergize with Itk in signaling from the T cell receptor to activate phospholipase C-γ [20]. The similarity of the phosphorylation of Rlk by Src family kinases to that which activates other Tec kinases suggests that Rlk also functions downstream of Src kinases in TCR signal transduction pathways. Rlk and Itk, may therefore function together in these TCR signaling events in a fashion similar to Btk in BCR signaling. Recent data argue that Itk interacts directly with Slp-76 and Grb2 to participate in a LAT-nucleated T cell signaling complex involved in PLC-γ activation [5,29]. It will be of interest to examine the interactions of Rlk with other components of this complex. Indeed, the potential in vitro binding of the amino-terminus of Rlk to the SH3 domain of Grb2 hints of similar protein interactions. Thus, although Rlk appears to be distinct among the Tec kinases in the lack of a PH domain, the presence of a palmitoylated cysteine string and the existence of a shorter isoform that can localize to the nucleus, Rlk may contribute to similar signaling pathways. Indeed, our data suggest that the cysteine string may serve as a surrogate to the PH domain for targeting Rlk to RAFTs. It is also notable that recent data suggest that Btk can translocate to the nucleus upon B cell activation, similar to the nuclear translocation we have reported for Rlk [19,51]. Nonetheless, the PI3K independence of the activation of Rlk activation and its association with RAFTs suggests that Rlk may also have distinct contributions to these signaling pathways.
Materials and Methods
Cell Culture and Reagents
293T and HeLa cells were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal calf serum, penicillin, streptomycin, and 10 mM glutamine. Jurkat cells expressing SV40 large T antigen (Jurkat Tag) were a generous gift from Dr. Gerald Crabtree (Stanford) and were maintained in RPMI 1640 supplemented with 10% fetal calf serum, penicillin, streptomycin, glutamine, and 50 μM 2-mercaptoethanol. Anti-murine Rlk was previously described. Anti-Fyn (FYN3) was purchased from Santa Cruz Biotechnology (Santa Cruz, CA); anti-phosphotyrosine (4G10) from Upstate Biotechnologies (Lake Placid, New York).
Transfections
Constructs were introduced into 293T cells by calcium phosphate transfection in medium containing 25 μM chloroquine (Sigma) [52]. 2-5 μg of each plasmid was used for each transfection (2 × 106 cells); the cells were harvested 24 to 36 hours later. Transfections into HeLa cells were performed identically, without the addition of chloroquine. Jurkat Tag cell lines (107 cells) were transfected by electroporation using a Biorad electroporator at 250 V and 960 μF or by square wave electroporation on a BTX 830 using settings of 300 V and 10 ms. 24 hours later, cells were stimulated with OKT3, lysed and Rlk analyzed as below.
Constructs and Mutagenesis
Full-length Rlk cDNA expression constructs pCIR, pcDNA3Rlk and Rlk-GFP have been described previously [19]. Point mutations were introduced into Rlk-GFP by a PCR based strategy [53] with oligonucleotides encoding the required mutations (Mutated residues are in bold): Y420F (GGAC GAT GAA TTC ATC AGT TCT TCT G), Y91F (GTC AAG GCT CTT TTT GAC TTC CTG CC). All mutations were confirmed by dideoxy sequencing. pLNCX-FynT was a generous gift from Dr. Cliff Lowell (USCF, San Francisco, CA). Glutathione S-Transferase (GST) fusion constructs were previously described [19].
In vitro filter binding assays
GST-Rlk fusion proteins were grown and induced as previously described [19]. Bacteria were sonicated in phosphate-buffered saline (PBS), Ph7.4 with 0.1% Triton X-100, insoluble material removed by centrifugation for 10 minutes at 14,000 rpm and soluble GST fusion proteins purified with GST-Sepharose beads (Pharmacia). Approximately 2 μg of GST fusion proteins were separated by SDS-PAGE, transferred to Nitrocellulose and probed with biotinylated SH3-GST fusion proteins as previously described [25].
Immunofluorescence
HeLa cells were transfected with Rlk and Fyn expression constructs, 20 hours later passed onto coverslips and 12-24 hours later fixed with 1% paraformaldehyde for 10 minutes at room temperature. Cells were stained with Fyn3 antiserum at 1: 500, washed with PBS and stained with Texas Red conjugated anti-Rabbit Goat serum (Cappel). Nuclei were counterstained with DAPI, cells were visualized on a Zeiss Axioplan microscope with a Charge Coupled Device (CCD) camera.
Western Blotting and Immunoprecipitation
24 hours following transfection cells were washed once with PBS and lysed in NP40 Lysis Buffer (0.5% Nonidet-P 40, 50 mM HEPES pH 7.4, 5 mM EDTA, 50 mM NaCl, 10 mM NaPO4, 50 mM NaF, 1 mM sodium orthovanadate, 1 mM AEBSF, 2 ug/ml aprotinin) for 10 minutes on ice. Total protein extracts were directly mixed with 2X SDS protein sample buffer (1X: 50 mM Tris pH 6.8, 2% SDS, 10% glycerol, 0.1% Bromphenol Blue plus 5% B-mercaptoethanol). Alternatively, lysates were clarified by centrifugation and detergent-soluble protein was analyzed. Equivalent amounts of protein were boiled in SDS sample buffer and separated by 10% SDS-PAGE and transferred onto nitrocellulose (Schleicher and Schuell). The membranes were blocked in TBST (10 mM Tris-HCl pH 8.0, 150 mM NaCl, 0.1% Tween-20) with 5% nonfat dry milk (w/v). The membranes were incubated with the appropriate primary antibody at 1/2000 (anti-Rlk and 4G10, anti-phosphotyrosine) or 1:1000 (anti-Fyn) dilution overnight at 4°C or 1-2 hours at room temperature. Membranes were washed in TBST, incubated with the appropriate secondary antibody at 1/10000 dilution (Boehringer Mannheim) for 1 hr and protein was visualized by enhanced chemiluminescence (Amersham).
For immunoprecipitation studies, equivalent amounts of detergent soluble protein were incubated with affinity purified anti-Rlk sera for 2-4 hours at 4°C. The complexes were recovered with Protein A-Sepharose (Sigma) and washed two times with ice cold NP40 wash buffer (1% NP40, PBS, 1 mM sodium orthovanadate) and analyzed by Western blotting using either anti-Rlk serum or anti-Fyn serum (1:1000).
In Vitro Kinase Assays
Cell extracts were prepared in NP40 Lysis Buffer and immunoprecipitated as described above. Complexes were recovered with Protein A Sepharose, washed twice with ice cold NP40 wash buffer, washed twice with ice cold Rlk kinase buffer (30 mM HEPES pH 7.4, 150 mM NaCl, 5 mM MgCl2, 5 mM MnCl2, 100 μM sodium orthovanadate). The kinase reaction was carried out in kinase buffer supplemented with 1 μg acid denatured enolase and 10 μCi of [γ-32P] ATP (Redivue, Amersham) for 5 minutes at room temperature. The reaction was quenched with SDS sample buffer, boiled and separated by 10% SDS-PAGE. Phosphorylated proteins were detected by autoradiography or quantified on a PhosphoImager. The level of Rlk protein was analyzed by immunoblotting.
Raft Isolation
Jurkat Tag cells were transfected by square wave electroporation using the parameters of 300 V and 10 ms. For each condition 8 cuvettes containing 107 cells in 0.5 ml were electroporated with 25 μg of the appropriate DNA. Cells were grown over night in RPMI containing 20% fetal calf serum. In some experiments, cells were pretreated with Wortmannin (100 nM) for 3 hrs prior to fractionation. For TCR stimulation, cells for each condition were concentrated into 0.5 ml, warmed to 37°C for 10 minutes and then treated with 10 μg OKT3. Ice cold PBS was added to the cells after 2 minutes OKT3 treatment and the cells were quickly pelleted and lysed in 0.5 ml TENV (10 mM Tris 7.5, 150 mM NaCl, 5 mM EDTA, 1 mM Na3VO4) containing 1% Triton X-100. The cells were homogenized with 12 strokes in 2 ml Dounce homogenizers after 30 minutes of incubation on ice. Homogenates were adjusted to 40% sucrose by addition of TENV containing 80% sucrose. The lysate was transferred to a TLS55 ultracentrifuge tube, over-layered sequentially with 30% and 5% sucrose in TENV. Sucrose gradients were centrifuged at 200,000 g for 16 hours and 200 μl fractions were removed from the top of the gradient for analysis by SDS-PAGE.
Metabolic Labeling and Pulse Chase
5 × 106 293T cells were transfected with 10 μg of the indicated Rlk construct and 10 μg of empty vector or the indicated Fyn construct. Twenty four hours after transfection, the cells were split equally among five to seven plates and grown in complete DMEM for another 24 hours. The cells were placed in serum-free Met(-), Cys(-) DMEM for 1 hour prior to pulse labeling. Cells were pulse labeled with 0.5 mCi of Trans- [35S]-Label (ICN) in 0.5 ml of Met(-)Cys(-) medium for 15 minutes at 37°C. After 15 minutes, the monolayers were chased with complete DMEM supplemented with 20 mM unlabeled methionine and cysteine (Sigma) at 37°C. At the indicated chase time, the cells were lysed with 1 ml cold NP40 Lysis Buffer and the lysates were clarified by centrifugation. Rlk was immunoprecipitated with affinity purified anti-Rlk antibody for 3 hours at 4°C and immune complexes were recovered with Protein A-Sepharose, washed as described above, and boiled in SDS-sample buffer. Labeled proteins were separated by 10% PAGE, and analyzed by fluorography.
Abbreviations
PH, pleckstrin homology; PI3K, phosphoinositide 3-Kinase; SH2, Src homology-2, SH3, Src homology-3; XLA, X-linked agammaglobulinemia; xid; x-linked immunodeficiency; TCR, T cell receptor; sIgM, surface immunoglobulin M; (GAP), GTPase-Activating Protein; GST, Glutathione S-Transferase; SDS, Sodium Dodecyl Sulfate; PBS, phosphate buffered saline; NP40, Nonidet P-40; PAGE, polyacrylamide gel electrophoresis.
Acknowledgments
Acknowledgements
Mario Chamorro, Michael J Czar and Jayanta Debnath are equal contributors to this article.
P.L.S. was a Special Fellow of the Leukemia Society of America and is presently a Searle Scholar. M.C. is presently in the Program in Cell Biology, Cornell Weill-Sloan Kettering Institute Graduate Program. The authors would like to thank D. W. McVicar, R. Wange and members of the Schwartzberg, Varmus and Lenardo laboratories for critical feedback and L. Matis for the original Rlk cDNA clone.
Contributor Information
Mario Chamorro, Email: chamorrm@mskcc.org.
Michael J Czar, Email: mczar@nhgri.nih.gov.
Jayanta Debnath, Email: jayanta_debnath@hms.harvard.edu.
Genhong Cheng, Email: genhongc@microbio.ucla.edu.
Michael J Lenardo, Email: MLENARDO@niaid.nih.gov.
Harold E Varmus, Email: varmus@mskcc.org.
Pamela L Schwartzberg, Email: pams@nhgri.nih.gov.
References
- Wange RL, Samelson LE. Complex complexes: signaling at the TCR. Immunity. 1996;5:197–205. doi: 10.1016/s1074-7613(00)80315-5. [DOI] [PubMed] [Google Scholar]
- Weiss A, Littman DR. Signal transduction by lymphocyte antigen receptors. Cell. 1994;76:263–274. doi: 10.1016/0092-8674(94)90334-4. [DOI] [PubMed] [Google Scholar]
- Rawlings DJ, Witte ON. The Btk subfamily of cytoplasmic tyrosine kinases: structure, regulation and function. Semin Immunol. 1995;7:237–246. doi: 10.1006/smim.1995.0028. [DOI] [PubMed] [Google Scholar]
- Satterthwaite AB, Li Z, Witte ON. Btk function in B cell development and response. Semin Immunol. 1998;10:309–316. doi: 10.1006/smim.1998.0123. [DOI] [PubMed] [Google Scholar]
- Schaeffer EM, Schwartzberg PL. Tec family kinases in lymphocyte signaling and function. Curr Opin Immunol. 2000;12:282–288. doi: 10.1016/S0952-7915(00)00088-1. [DOI] [PubMed] [Google Scholar]
- Mano H, Ishikawa F, Nishida J, Hirai H, Takaku F. A novel protein-tyrosine kinase, tec, is preferentially expressed in liver. Oncogene. 1990;5:1781–1786. [PubMed] [Google Scholar]
- Desiderio S, Siliciano JD. The Itk/Btk/Tec family of protein-tyrosine kinases. Chem Immunol. 1994;59:191–210. [PubMed] [Google Scholar]
- Rawlings DJ, Saffran DC, Tsukada S, Largaespada DA, Grimaldi JC, Cohen L, Mohr RN, Bazan JF, Howard M, Copeland NG, et al. Mutation of unique region of Bruton's tyrosine kinase in immunodeficient XID mice. Science. 1993;261:358–361. doi: 10.1126/science.8332901. [DOI] [PubMed] [Google Scholar]
- Thomas JD, Sideras P, Smith CI, Vorechovsky I, Chapman V, Paul WE. Colocalization of X-linked agammaglobulinemia and X-linked immunodeficiency genes. Science. 1993;261:355–358. doi: 10.1126/science.8332900. [DOI] [PubMed] [Google Scholar]
- Tsukada S, Saffran DC, Rawlings DJ, Parolini O, Allen RC, Klisak I, Sparkes RS, Kubagawa H, Mohandas T, Quan S, et al. Deficient expression of a B cell cytoplasmic tyrosine kinase in human X-linked agammaglobulinemia. Cell. 1993;72:279–290. doi: 10.1016/0092-8674(93)90667-f. [DOI] [PubMed] [Google Scholar]
- Vetrie D, Vorechovsky I, Sideras P, Holland J, Davies A, Flinter F, Hammarstrom L, Kinnon C, Levinsky R, Bobrow M, et al. The gene involved in X-linked agammaglobulinaemia is a member of the src family of protein-tyrosine kinases. Nature. 1993;361:226–233. doi: 10.1038/361226a0. [DOI] [PubMed] [Google Scholar]
- Kawakami Y, Yao L, Miura T, Tsukada S, Witte ON, Kawakami T. Tyrosine phosphorylation and activation of Bruton tyrosine kinase upon Fc epsilon RI cross-linking. Mol Cell Biol. 1994;14:5108–5113. doi: 10.1128/mcb.14.8.5108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsuda T, Takahashi-Tezuka M, Fukada T, Okuyama Y, Fujitani Y, Tsukada S, Mano H, Hirai H, Witte ON, Hirano T. Association and activation of Btk and Tec tyrosine kinases by gp130, a signal transducer of the interleukin-6 family of cytokines. Blood. 1995;85:627–633. [PubMed] [Google Scholar]
- Saouaf SJ, Mahajan S, Rowley RB, Kut SA, Fargnoli J, Burkhardt AL, Tsukada S, Witte ON, Bolen JB. Temporal differences in the activation of three classes of non-transmembrane protein tyrosine kinases following B-cell antigen receptor surface engagement. Proc Natl Acad Sci U S A. 1994;91:9524–9528. doi: 10.1073/pnas.91.20.9524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato S, Katagiri T, Takaki S, Kikuchi Y, Hitoshi Y, Yonehara S, Tsukada S, Kitamura D, Watanabe T, Witte O, et al. IL-5 receptor-mediated tyrosine phosphorylation of SH2/SH3-containing proteins and activation of Bruton's tyrosine and Janus 2 kinases. J Exp Med. 1994;180:2101–2111. doi: 10.1084/jem.180.6.2101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heyeck SD, Wilcox HM, Bunnell SC, Berg LJ. Lck phosphorylates the activation loop tyrosine of the Itk kinase domain and activates Itk kinase activity. J Biol Chem. 1997;272:25401–25408. doi: 10.1074/jbc.272.40.25401. [DOI] [PubMed] [Google Scholar]
- Gibson S, August A, Branch D, Dupont B, Mills GM. Functional LCK Is required for optimal CD28-mediated activation of the TEC family tyrosine kinase EMT/ITK. J Biol Chem. 1996;271:7079–7083. doi: 10.1074/jbc.271.28.16662. [DOI] [PubMed] [Google Scholar]
- August A, Gibson S, Kawakami Y, Kawakami T, Mills GB, Dupont B. CD28 is associated with and induces the immediate tyrosine phosphorylation and activation of the Tec family kinase ITK/EMT in the human Jurkat leukemic T-cell line. Proc Natl Acad Sci U S A. 1994;91:9347–9351. doi: 10.1073/pnas.91.20.9347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Debnath J, Chamorro M, Czar MJ, Schaeffer EM, Leonardo MJ, Varmus HE, Schwartzberg PL. Rlk/Txk encodes two forms of a novel cysteine-string tyrosine kinae activated by Src family kinases. Mol Cell Biol. 1999;19:1498–1507. doi: 10.1128/mcb.19.2.1498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaeffer EM, Debnath J, Yap G, McVicar D, Liao XC, Littman DR, Sher A, Varmus HE, Lenardo MJ, Schwartzberg PL. Requirement for Tec kinases Rlk and Itk in T cell receptor signaling and immunity. Science. 1999;284:638–641. doi: 10.1126/science.284.5414.638. [DOI] [PubMed] [Google Scholar]
- Mahajan S, Fargnoli J, Burkhardt AL, Kut SA, Saouaf SJ, Bolen JB. Src family protein tyrosine kinases induce autoactivation of Bruton's tyrosine kinase. Mol Cell Biol. 1995;15:5304–5311. doi: 10.1128/mcb.15.10.5304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mano H, Yamashita Y, Miyazato A, Miura Y, Ozawa K. Tec protein-tyrosine kinase is an effector molecule of Lyn protein-tyrosine kinase. Faseb J. 1996;10:637–642. doi: 10.1096/fasebj.10.5.8621063. [DOI] [PubMed] [Google Scholar]
- Rawlings DJ, Scharenberg AM, Park H, Wahl MI, Lin S, Kato RM, Fluckiger AC, Witte ON, Kinet JP. Activation of BTK by a phosphorylation mechanism initiated by SRC family kinases. Science. 1996;271:822–825. doi: 10.1126/science.271.5250.822. [DOI] [PubMed] [Google Scholar]
- Park H, Wahl MI, Afar DE, Turck CW, Rawlings DJ, Tam C, Scharenberg AM, Kinet JP, Witte ON. Regulation of Btk function by a major autophosphorylation site within the SH3 domain. Immunity. 1996;4:515–525. doi: 10.1016/s1074-7613(00)80417-3. [DOI] [PubMed] [Google Scholar]
- Cheng G, Ye ZS, Baltimore D. Binding of Bruton's tyrosine kinase to Fyn, Lyn, or Hck through a Src homology 3 domain-mediated interaction. Proc Natl Acad Sci U S A. 1994;91:8152–8155. doi: 10.1073/pnas.91.17.8152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang W, Malek SN, Desiderio S. An SH3-binding site conserved in Bruton's tyrosine kinase and related tyrosine kinases mediates specific protein interactions in vitro and in vivo. J Biol Chem. 1995;270:20832–20840. doi: 10.1074/jbc.270.35.20832. [DOI] [PubMed] [Google Scholar]
- August A, Sadra A, Dupont B, Hanafusa H. Src-induced activation of inducible T cell kinase (ITK) requires phosphatidylinositol 3-kinase activity and the Pleckstrin homology domain of inducible T cell kinase. Proc Natl Acad Sci U S A. 1997;94:11227–11232. doi: 10.1073/pnas.94.21.11227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Z, Wahl M, Eguino A, Stephens L, Hawkins P, Witte O. Phosphotidylinositol 3-kinase-γ activates Bruton's tyrosine kinase in concert with Src family kinases. Proc Natl Acad Sci, USA. 1997;94:13820–13825. doi: 10.1073/pnas.94.25.13820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bunnell S, Diehn M, Yaffe M, Findell P, Cantley L, Berg L. Biochemical Interactions Integrating Itk with the T cell Receptor-initiated Signaling Cascade. J Biol Chem. 2000;275:2219–2230. doi: 10.1074/jbc.275.3.2219. [DOI] [PubMed] [Google Scholar]
- Haire RN, Ohta Y, Lewis JE, Fu SM, Kroisel P, Litman GW. TXK, a novel human tyrosine kinase expressed in T cells shares sequence identity with Tec family kinases and maps to 4p12. Hum Mol Genet. 1994;3:897–901. doi: 10.1093/hmg/3.6.897. [DOI] [PubMed] [Google Scholar]
- Hu Q, Davidson D, Schwartzberg PL, Macchiarini F, Lenardo MJ, Bluestone JA, Matis LA. Identification of Rlk, a novel protein tyrosine kinase with predominant expression in the T cell lineage. J Biol Chem. 1995;270:1928–1934. doi: 10.1074/jbc.270.4.1928. [DOI] [PubMed] [Google Scholar]
- Sommers CL, Huang K, Shores EW, Grinberg A, Charlick DA, Kozak CA, Love PE. Murine txk: a protein tyrosine kinase gene regulated by T cell activation. Oncogene. 1995;11:245–251. [PubMed] [Google Scholar]
- Alexandropoulos K, Cheng G, Baltimore D. Proline-rich sequences that bind to Src homology 3 domains with individual specificities. Proc Natl Acad Sci U S A. 1995;92:3110–3114. doi: 10.1073/pnas.92.8.3110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanks SK, Quinn AM, Hunter T. The protein kinase family: conserved features and deduced phylogeny of the catalytic domains. Science. 1988;241:42–52. doi: 10.1126/science.3291115. [DOI] [PubMed] [Google Scholar]
- Guo B, Kato RM, Garcia-Lloret M, Wahl MI, Rawlings DJ. Engagement of the human pre-B cell receptor generates a lipid raft-dependent calcium signaling complex. Immunity. 2000;13:243–253. doi: 10.1016/s1074-7613(00)00024-8. [DOI] [PubMed] [Google Scholar]
- Shan X, Czar MJ, Bunnell SC, Liu P, Liu Y, Schwartzberg PL, Wange RL. Deficiency of PTEN in Jurkat T cells causes constitutive localization of Itk to the plasma membrane and hyperresponsiveness to CD3 stimulation. Mol Cell Biol. 2000;20:6945–6957. doi: 10.1128/MCB.20.18.6945-6957.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wahl MI, Fluckiger AC, Kato RM, Park H, Witte ON, Rawlings DJ. Phosphorylation of two regulatory tyrosine residues in the activation of Bruton's tyrosine kinase via alternative receptors. Proc Natl Acad Sci U S A. 1997;94:11526–11533. doi: 10.1073/pnas.94.21.11526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nisitani S, Kato RM, Rawlings DJ, Witte ON, Wahl MI. In situ detection of activated Bruton's tyrosine kinase in the Ig signaling complex by phosphopeptide-specific monoclonal antibodies. Proc Natl Acad Sci U S A. 1999;96:2221–2226. doi: 10.1073/pnas.96.5.2221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denny MF, Patai B, Straus DB. Differential T-cell antigen receptor signaling mediated by the Src family kinases Lck and Fyn. Mol Cell Biol. 2000;20:1426–1435. doi: 10.1128/MCB.20.4.1426-1435.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang J, Tilly D, Altman A, Sugie K, Grey HM. Inaugural article: T-cell receptor antagonists induce vav phosphorylation by selective activation of fyn kinase [In Process Citation]. Proc Natl Acad Sci U S A. 2000;97:10923–10929. doi: 10.1073/pnas.97.20.10923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider H, Guerette B, Guntermann C, Rudd CE. Resting lymphocyte kinase (Rlk/Txk) targets lymphoid adaptor SLP-76 in the cooperative activation of interleukin-2 transcription in T-cells. J Biol Chem. 2000;275:3835–3840. doi: 10.1074/jbc.275.6.3835. [DOI] [PubMed] [Google Scholar]
- Tomlinson MG, Kurosaki T, Berson AE, Fujii GH, Johnston JA, Bolen JB. Reconstitution of Btk signaling by the atypical tec family tyrosine kinases Bmx and Txk. J Biol Chem. 1999;274:13577–13585. doi: 10.1074/jbc.274.19.13577. [DOI] [PubMed] [Google Scholar]
- Rozdzial M, Malissen B, Finkel T. Tyrosine-phosphorylated T cell receptor zeta chain associates with the actin cytoskeleton upon activation of mature T lymphocytes. Immunity. 1995;3:623–633. doi: 10.1016/1074-7613(95)90133-7. [DOI] [PubMed] [Google Scholar]
- Montixi C, Langlet C, Bernard AM, Thimonier J, Dubois C, Wurbel MA, Chauvin JP, Pierres M, He HT. Engagement of T cell receptor triggers its recruitment to low-density detergent-insoluble membrane domains. Embo J. 1998;17:5334–5348. doi: 10.1093/emboj/17.18.5334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xavier R, Brennan T, Li Q, McCormack C, Seed B. Membrane compartmentation is required for efficient T cell activation. Immunity. 1998;8:723–732. doi: 10.1016/s1074-7613(00)80577-4. [DOI] [PubMed] [Google Scholar]
- Valitutti S, Muller S, Salio M, Lanzavecchia A. Degradation of T cell receptor (TCR)-CD3-zeta complexes after antigenic stimulation [see comments]. J Exp Med. 1997;185:1859–1864. doi: 10.1084/jem.185.10.1859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma YC, Huang XY. Identification of the binding site for Gqalpha on its effector Bruton's tyrosine kinase. Proc Natl Acad Sci U S A. 1998;95:12197–12201. doi: 10.1073/pnas.95.21.12197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyake S, Lupher ML, Jr, Druker B, Band H. The tyrosine kinase regulator Cbl enhances the ubiquitination and degradation of the platelet-derived growth factor receptor alpha. Proc Natl Acad Sci U S A. 1998;95:7927–7932. doi: 10.1073/pnas.95.14.7927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyake S, Mullane-Robinson KP, Lill NL, Douillard P, Band H. Cbl-mediated negative regulation of platelet-derived growth factor receptor-dependent cell proliferation. A critical role for Cbl tyrosine kinase-binding domain. J Biol Chem. 1999;274:16619–16628. doi: 10.1074/jbc.274.23.16619. [DOI] [PubMed] [Google Scholar]
- D'Oro U, Vacchio MS, Weissman AM, Ashwell JD. Activation of the Lck tyrosine kinase targets cell surface T cell antigen receptors for lysosomal degradation. Immunity. 1997;7:619–628. doi: 10.1016/s1074-7613(00)80383-0. [DOI] [PubMed] [Google Scholar]
- Webb CF, Yamashita Y, Ayers N, Evetts S, Paulin Y, Conley ME, Smith EA. The transcription factor bright associates with Bruton's tyrosine kinase, the defective protein in immunodeficiency disease. J Immunol. 2000;165:6956–6965. doi: 10.4049/jimmunol.165.12.6956. [DOI] [PubMed] [Google Scholar]
- Pear WS, Nolan GP, Scott ML, Baltimore D. Production of high-titer helper-free retroviruses by transient transfection. Proc Natl Acad Sci U S A. 1993;90:8392–8396. doi: 10.1073/pnas.90.18.8392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horton RM. In vitro recombination and mutagenesis of DNA. SOEing together tailor-made genes. Methods Mol Biol. 1997;67:141–149. doi: 10.1385/0-89603-483-6:141. [DOI] [PubMed] [Google Scholar]