

Abstract
BACKGROUND AND PURPOSE
Cannabis extracts and several cannabinoids have been shown to exert broad anti-inflammatory activities in experimental models of inflammatory CNS degenerative diseases. Clinical use of many cannabinoids is limited by their psychotropic effects. However, phytocannabinoids like cannabidiol (CBD), devoid of psychoactive activity, are, potentially, safe and effective alternatives for alleviating neuroinflammation and neurodegeneration.
EXPERIMENTAL APPROACH
We used experimental autoimmune encephalomyelitis (EAE) induced by myelin oligodendrocyte glycoprotein (MOG) in C57BL/6 mice, as a model of multiple sclerosis. Using immunocytochemistry and cell proliferation assays we evaluated the effects of CBD on microglial activation in MOG-immunized animals and on MOG-specific T-cell proliferation.
KEY RESULTS
Treatment with CBD during disease onset ameliorated the severity of the clinical signs of EAE. This effect of CBD was accompanied by diminished axonal damage and inflammation as well as microglial activation and T-cell recruitment in the spinal cord of MOG-injected mice. Moreover, CBD inhibited MOG-induced T-cell proliferation in vitro at both low and high concentrations of the myelin antigen. This effect was not mediated via the known cannabinoid CB1 and CB2 receptors.
CONCLUSIONS AND IMPLICATIONS
CBD, a non-psychoactive cannabinoid, ameliorates clinical signs of EAE in mice, immunized against MOG. Suppression of microglial activity and T-cell proliferation by CBD appeared to contribute to these beneficial effects.
LINKED ARTICLES
This article is part of a themed issue on Cannabinoids in Biology and Medicine. To view the other articles in this issue visit http://dx.doi.org/10.1111/bph.2011.163.issue-7
Keywords: cannabinoids, cannabidiol, microglia, T cells, EAE, MOG
Introduction
The anti-inflammatory properties of cannabinoids, constituents of the Cannabis sativa plant, have been appreciated since ancient times and have been supported experimentally, in models of inflammation in vitro and in vivo (Tanasescu and Constantinescu, 2010). Indeed, immune cells express the elements of the cannabinoid system including endocannabinoid ligands, endocannabinoid enzymatic machinery and cannabinoid CB receptors, mostly of the CB2 type and many fewer of the CB1 type (Mackie, 2005; nomenclature follows Alexander et al. (2009). Moreover, expression of CB1 and CB2 receptors and the activity of the endocannabinoid system are regulated in response to inflammation, a finding that further confirms the involvement of the cannabinoid system in immune processes (Croxford and Yamamura, 2005; Maresz et al., 2005; Pietr et al., 2009).
The clear and promising therapeutic potential of cannabinoids (Klein and Newton, 2007) is limited due to central, CB1-mediated, psychotropic effects of many of these materials. A good example is Δ9-tetrahydrocannabinol (THC) as the most studied psychoactive cannabinoid, activating both CB1 and CB2 receptors (Bayewitch et al., 1995). The preferential CB2 receptor expression on immune cells offers an attractive opportunity to regulate the function of the immune system with CB2 receptor ligands. Studies on CB1/CB2 receptor knock-down mice revealed the existence of other possible, receptor and non-receptor, cannabinoid targets (Járai et al., 1999), confirmed to be immunomodulatory (Kaplan et al., 2003; 2008;). Thus, cannabinoid compounds not acting on CB1 or CB2 receptors seem to offer new tools to manipulate inflammation. Cannabidiol (CBD) is one of these compounds and is the major Cannabis-derived non-CB1/CB2 receptor ligand (Showalter et al., 1996). This compound is not only devoid of psychotropic effects but also is able to inhibit many central effects of CB1 receptor ligands, for example, the anxiogenic and psychotogenic activities of THC (Zuardi, 2008). Interestingly, despite different pharmacological and behavioural effects, CBD shares with ‘classic’ psychocannabinoids many beneficial effects, including its capacity to act as an immunomodulator.
Indeed, CBD exerts a wide range of anti-inflammatory properties and regulates cell cycle and function of various immune cells. These effects include suppression of humoral responses, such as release of cytokines, chemokines, growth factors, as well as suppression of immune cell proliferation, activation, maturation, migration and antigen presentation (Mechoulam et al., 2007). In an earlier publication, we showed that CBD inhibited production of the cytokine IL-6 and the chemokine CCL-2 by activated microglial cells and in parallel activated intracellular anti-inflammatory pathways (Kozela et al., 2010).
Among the many types of neurodegenerative diseases in which inflammation is involved, multiple sclerosis (MS) is one of those clearly induced and driven by dysfunctional immune system activity. In MS, myelin autoreactive peripheral T cells migrate into the CNS and initiate cytotoxic, degenerative processes that include demyelination, oligodendrocyte cell death and axonal degeneration. These effects lead to neurological deficits such as visual and sensory disturbances, motor weakness, tremor, ataxia and progressive disability as the main clinical symptoms (Compston and Coles, 2008). Infiltrating T cells are constantly reactivated within the CNS parenchyma by microglia. Microglial cells via chemokine and cytokine release and constant antigen presentation potentiate T-cell recruitment to the CNS and facilitate their polarization into cytotoxic phenotypes (Th1 or Th17). Moreover, via released chemokines such as CCL-2, microglial cells recruit other immune cells of myeloid origin, specialized in epitope spreading and phagocytosis of myelin including monocytes, macrophages, B cells and dendritic cells (Jack et al., 2005; Koning et al., 2009). Depletion of microglia or impairment of their function can attenuate disease progression in experimental animal models of MS supporting their role in initiation and development of this disease (Huitinga et al., 1990; Heppner et al., 2005). Thus, suppression of microglia will potentially reduce inflammatory lesions and limit demyelination within the CNS.
One of the best described and commonly used animal models of MS is experimental autoimmune encephalomyelitis (EAE) which is induced when the animals are immunized with myelin components, for example, myelin basic protein, proteolipid protein or myelin oligodendrocyte glycoprotein (MOG) or by passive transfer of autoreactive myelin specific T cells to produce demyelination and MS-like neurological and clinical signs (Shevach, 1999). Using the MOG-induced EAE mouse model, we investigated if systemically given CBD at the time of symptomatic disease onset could affect the progression of the disease. We observed that CBD ameliorated the severity of the EAE in MOG-injected mice. Moreover, CBD attenuated microglia activation in MOG-immunized animals and inhibited MOG-induced proliferation of encephalitogenic T cells.
Methods
Animals and experimental design
All animal care and experimental procedures complied with and were approved under the guidance and regulations of the Weizmann Institute of Science and Tel Aviv University. EAE was induced in 30 8-week-old female C57BL/6 mice (Harlan Laboratories, Rehovot, Israel) by two subcutaneous injections of MOG35-55 fragment (encompassing amino acids 35–55 of MOG) on days 1 and 8, injected in the left and right flanks respectively. Each injection contained 300 µg MOG in 200 µL of complete Freund's adjuvant (CFA) containing Mycobacterium tuberculosis at 200 µg (Sigma, Israel). Control mice received only CFA without MOG (Ctrl, n = 10).
CBD (5 mg·kg−1; CBD + EAE group) or its vehicle (cremophor, ethanol and PBS, v : v : v 1:1:18; EAE group) was injected (i.p.) into 15 MOG-immunized mice immediately at the onset of disease signs for 3 consecutive days (on days 19, 20 and 21). Clinical disease scores were recorded daily until 30 days after first immunization. At this time point, spinal cords were collected for further pathological and immunological studies. The signs of EAE were scored as follows: 0, no clinical signs; 1, loss of tail tonicity; 2, partial hind limb paralysis; 3, complete hind limb paralysis; 4, partial frontal limb paralysis; 5, total paralysis; 6, death (Lev et al., 2004). In parallel, we used control mice which received only CFA (Ctrl) and were also injected with CBD on days 19, 20 and 21 (Ctrl + CBD group, each group n = 10). The days of EAE onset and respective CBD injections were chosen based on our previous experience with this model (Lev et al., 2004).
CBD (National Institute on Drug Abuse, Rockville, MD, USA) solution was prepared freshly before each of the treatments. The dose of CBD was chosen based on previous studies with systemic administration of the drug in which CBD at 5 mg·kg−1 exhibited maximal efficiency at relieving rheumatoid signs in collagen-induced arthritis (Malfait et al., 2000). MOG synthesis was carried out by the Weizmann Institute Synthesis Unit, using a solid-phase technique on a peptide synthesizer (Applied Biosystems Inc., Foster City, CA, USA).
Histology and immunocytochemistry
Histological analysis of spinal cord sections was used to define severity of inflammation and demyelination and the effect of CBD treatment on these parameters. Spinal cords were dissected 30 days after first immunization of mice, fixed in 10% buffered formalin and embedded in paraffin. The presence of axonal pathology was supported by immunohistochemistry with anti-non-phosphorylated neurofilament H (SMI-32, Sternberg Antibodies, Emeryville, CA, USA) on 8 µm thick paraffin sections. Subsequently, 4 µm thick sections were stained with haematoxylin and eosin (H&E) and assessed for inflamed areas. The assessment of immune cell infiltration was performed using immunocytochemistry on 4 µm spinal cord sections and included staining for T cells (CD3+, Serotec, Kiddlington, Oxford, UK), microglia/macrophage number and their activation state (Iba-1, Wako, Richmond, VA, USA; and Mac-2/Galectin 3, Cedarlane, Burlington, Ontario, Canada).
It should be noted that the proteins, ionizing calcium-binding adaptor molecule 1 (Iba-1) and Mac-2/Galectin-3, do not discriminate between microglia, resident macrophages within the CNS, and invasive macrophage-like cells from peripheral sites that crossed the blood-brain barrier. Thus, in further description of the stainings we refer to microglia/macrophage populations. The Iba-1 protein is specifically expressed in microglia/macrophages and becomes up-regulated during the activation of these cells. Mac-2/Galectin-3 is another microglial/macrophage marker whose expression reflects specifically the activation state of these cells. For example, Mac-2/Galectin-3 expression is known to be increased in these cells following phagocytosis of damaged myelin and cell particles (Reichert and Rotshenker, 1999).
The immunostainings were visualized with respective secondary antibodies conjugated to Cy2 or Cy3 fluorochromes. Spinal cords from control, healthy mice that received 3 daily injections of either CBD or its vehicle were dissected and stained in parallel. Assessments of intensity and differences in immunofluorescence staining for Iba-1 and Mac-2/Galectin-3 were performed using the Image Pro Plus analysis software (Media cybernetics, Bethesda, MD, USA) and collected as arbitrary units representing optical density per area. The statistics was carried out on per cent values calculated from optical density values and is given in the appropriate figure legend. In the case of T cells, CD3+ cells were counted (three areas per mouse, three to four mice per treatment) and the cell numbers were subjected to statistical analysis.
Encephalitogenic T-cell line
The MOG35-55-specific T-cell line was established from lymph node cells of C57BL/6 female mice that had been primed 10 days earlier with MOG35-55 emulsified in CFA as previously described (Kaushansky et al., 2006). The T-cell line was maintained in vitro in RPMI-1640 (Biological Industries, Kibbutz Beit HaEmek, Israel) containing 5% fetal calf serum and supplemented with 10 U·mL−1 IL-2 (Peprotech Inc, Rocky Hill, NJ, USA), L-glutamine, 100 µg·mL−1 of streptomycin, 100 U·mL−1 of penicillin, 50 µM β-mercaptoethanol, 1 mM non-essential amino acids and 1 mM sodium pyruvate with alternate stimulation with MOG every 14 days as previously described (Ben-Nun and Lando, 1983; Kaushansky et al., 2006).
Encephalitogenic T-cell proliferation
The MOG35-55-reactive line of T cells raised from primed mice was cultured in 96-well plates (1.5 × 104 cells·per well) together with irradiated (25Gy) splenic antigen presenting cells (APCs; 5 × 105 cells per well). APCs were isolated from spleens of naïve C57BL/6 mice. Lysis of erythrocytes was performed using ACK solution (150 mM NH4Cl, 10 mM KHCO3 and 0.1 mM EDTA). The cells were cultured in 0.2 mL RPMI-1640 containing 2.5% fetal calf serum and supplemented with L-glutamine, 50 µM β-mercaptoethanol, and in the presence of MOG35-55 peptide (1 or 2.5 µg·mL−1). CBD at final concentrations of 1, 5 or 10 µM or its vehicle (0.1% ethanol in RPMI-1640) was added to the cells together with the MOG. The cells were then incubated for 72 h at 37°C in 5% CO2 humidified air (Ben-Nun and Lando, 1983; Kaushansky et al., 2006). Cell proliferation was measured by pulsing the cells with [3H]Thymidine (0.5 µCi·well−1) for the last 16 h of the incubation period and the cells were then harvested and counted using a Matrix 96 Direct β counter (Packard Instr., Meriden, CT, USA). To evaluate if CB1 or CB2 receptors were involved in the effects of CBD on T-cell proliferation, SR141716 (CB1 receptor antagonist; RTI International, NC, USA) and SR144528 (CB2 receptor antagonist; RTI International, NC, USA) at 1 µM concentration were applied 30 min before the application of CBD (5 or 10 µM) and MOG at 1 µg·mL−1. The proliferative responses of the cell microcultures were converted to per cent values with MOG effect expressed as 100%. Per cent values were calculated from Stimulation Index (SI) values which are the fold changes in mean counts per minute (cpm) of MOG cultures over mean cpm of cultures without MOG (spontaneous proliferation). Statistical analysis was performed on per cent values. Each experiment was carried out in triplicates and repeated three or four times.
Statistical analysis
EAE clinical scores were reported as average ± SD. Repeated measure analysis of variance (anova) was used for statistical analysis of these data. The fluorescence intensity and T-cell proliferation data were expressed as the mean ± SEM and analysed for statistical significance using one-way anova, followed by Bonferroni post hoc tests. P < 0.05 was considered significant. Graph Pad Prism program (La Jolla, CA, USA) was used for statistical analysis of the data.
Results
CBD ameliorates clinical signs of MOG-induced EAE and disease progression
C57BL/6 mice immunized with MOG35-55 developed severe EAE disease with complete hind limb paralysis [EAE mice, average clinical score 2.52 ± 0.54; Figure 1; with 13 out of 15 mice exhibiting clinical signs of EAE (EAE ratio 13/15)]. As shown in Figure 1, three injections (i.p.) of CBD (5 mg·kg−1, one a day) during the onset of clinical disease resulted in amelioration of the disease signs during the days of injections as well as markedly delaying disease progression. On day 21, MOG-induced EAE mice exhibited limp tail and partial paresis of the hind limbs (clinical score 1.03 ± 0.29; EAE ratio 9/15). However, MOG-induced EAE mice that received CBD during days 19–21 exhibited minimal clinical signs of EAE (average clinical score 0.13 ± 0.09, P < 0.05; Figure 1; EAE ratio 1/15). Interestingly, 2 days after the CBD injections (day 23), the clinical score of CBD-treated mice increased slowly, reaching mild disease severity at the end of the study (average clinical score 1 and EAE ratio 6/15 vs. average clinical score of 2.54 ± 0.54 in untreated EAE mice, P < 0.05; EAE ratio 13/15).
Figure 1.

Cannabidiol (CBD) ameliorates the clinical signs and disease progression of EAE induced by myelin oligodendrocyte glycoprotein (MOG). EAE was induced in C57BL/6 mice by flank s.c. immunization with MOG35-55. Clinical disease scores were recorded daily until day 30 after induction. CBD (CBD + EAE) or its vehicle (EAE group), was injected i.p. during the onset of the disease for 3 consecutive days (days 19–21; indicated by arrows). The mean clinical scores ± SD are shown. Each group consisted of 15 mice. Repeated measure anova showed significant differences between the EAE and CBD + EAE groups (P < 0.001).
CBD effect on spinal cord of healthy, non-EAE mice
Neither mice that received CFA alone nor those that received CFA with 5 mg·kg−1 of CBD exhibited any clinical signs of EAE (data not shown). Next, we evaluated if CBD injections in these healthy mice affected the spinal cords. As shown on Figure 2A, CBD injections did not affect the pattern of H&E histological staining, compared with the group of healthy mice, treated only with CFA. Moreover, Iba-1 microglial expression in Ctrl + CBD group treated with CBD and CFA did not differ from that in the mice given only CFA (Figure 2B). CD3+ T cells were absent in spinal cords of mice given CFA, with or without CBD (Figure 2C). The amount of staining is presented in Figures 5C and 6C.
Figure 2.
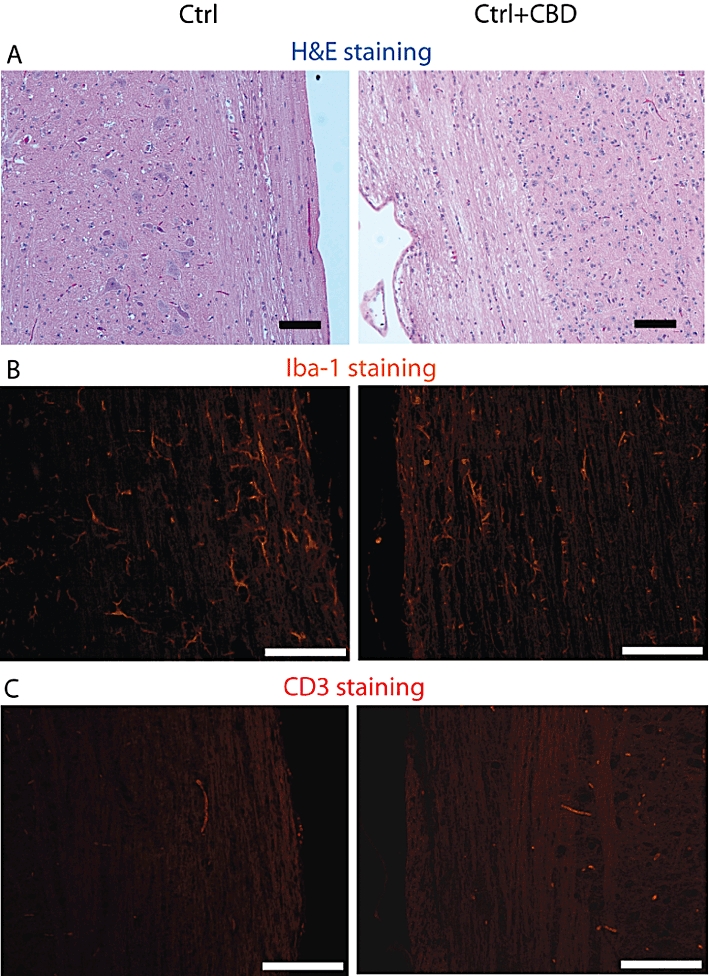
Cannabidiol (CBD) treatment of control, naïve C57BL/6 mice does not affect spinal cord integrity, basal microglial and T-cell number. The Figure shows representative stainings of spinal cord sections prepared from control healthy mice receiving complete Freund adjuvant either without CBD (Ctrl) or with CBD (Ctrl + CBD) treatment. (A) Haematoxylin and eosin (H&E) staining; (B) Iba-1 staining for the presence of microglial/macrophages; and (C) CD3+ staining for presence of T-cells. Scale bars represent 100 µm. Statistical analysis of the CD3+ and Iba-1 expression is presented in the bar graphs of Figures 5C (CD3+) and 6C (Iba-1).
Figure 5.
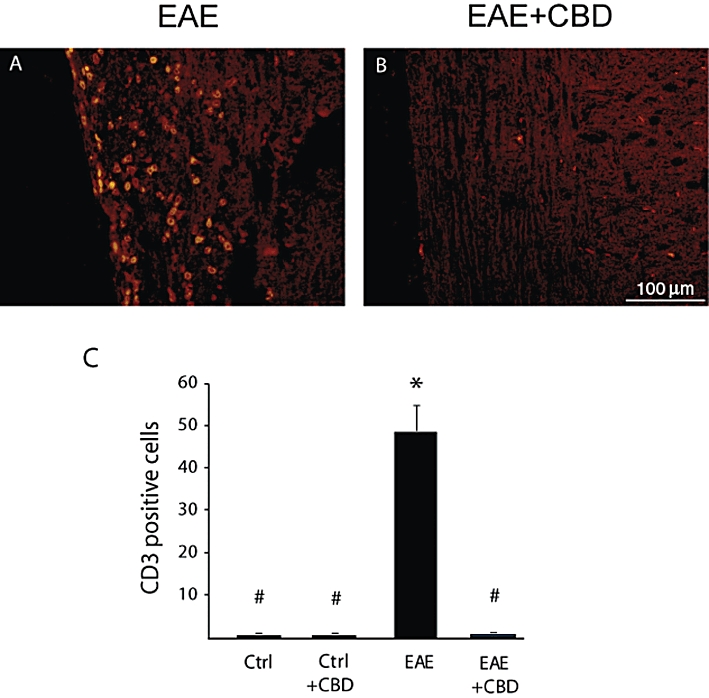
Cannabidiol (CBD) treatment attenuates EAE-induced T-cell infiltration into spinal cord of mice. CD3+ staining reveals that CBD treatment reduces the infiltration of T cells into the white matter of the spinal cords of EAE mice. (A) EAE mice. (B) EAE mice treated with CBD. Scale bar represents 100 µm. The results are presented as mean ± SEM of CD3+ cells from 9–12 spinal fragments from three to four mice per each experimental group. One-way anovaF(3,43) = 45.3, P < 0.0001, followed by Bonferroni post hoc test. *P < 0.001 versus Ctrl. #P < 0.001 versus EAE mice.
Figure 6.
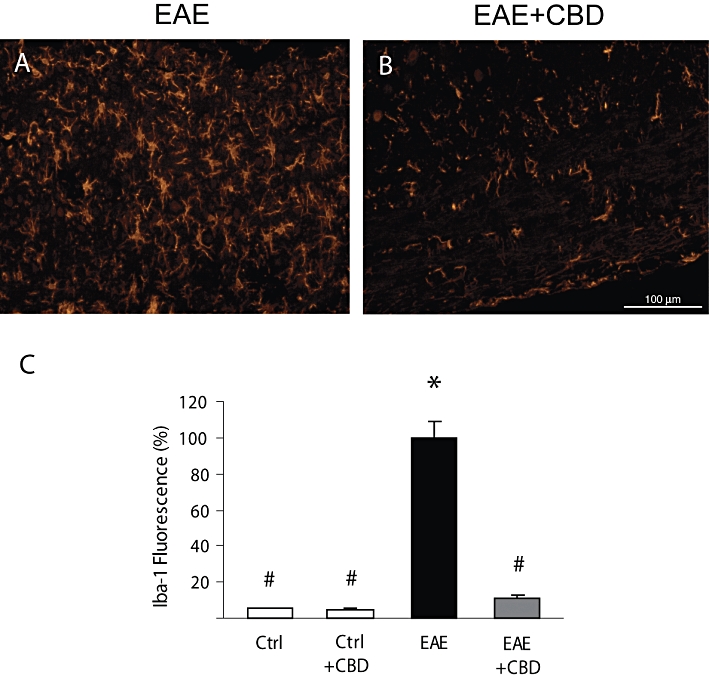
Cannabidiol (CBD) reduces the MOG-up-regulated Iba-1 expression in spinal cords of mice. Representative pictures showing that (A) MOG injections increase Iba-1 staining in spinal cord, and (B) that CBD injections significantly decrease the level of Iba-1 staining. (C) Quantification of Iba-1 fluorescence in sections of control (Ctrl) and of EAE mice with and without CBD treatment. The results are presented as mean ± SEM of total fluorescence from 8–11 spinal fragments from three to four mice per each experimental group. One-way anovaF(3,37) = 30.23, P < 0.0001, followed by Bonferroni post hoc test. *P < 0.001 versus Ctrl. #P < 0.001 versus EAE mice. Scale bar represents 100 µm.
CBD slows down MOG-induced axonal damage and inflammation in spinal cord of EAE mice
Mice were killed 30 days after EAE induction and spinal cord sections were analysed for the degree of damage to the white matter tissue, using the SMI-32 antibody which stains damaged axons. SMI-32 staining revealed extensive axonal degeneration in the MOG-immunized EAE mice. In contrast, CBD-treated EAE mice showed much lesser axonal damage (Figure 3). Next, we performed H&E histological staining to visualize the inflamed areas in the spinal cord. Results presented in Figure 4 show markedly weaker inflammation in sections of spinal cord derived from CBD-treated EAE mice, compared with untreated EAE mice. These results were further confirmed by immunocytochemical staining for CD3+ T lymphocytes and markers of macrophages/microglial cells. Spinal cords derived from the EAE mice were rich in CD3+ T-cell infiltrates (Figure 5A,C), while treatment with CBD profoundly and significantly decreased CD3+ infiltrates into the white matter (Figure 5B,C).
Figure 3.
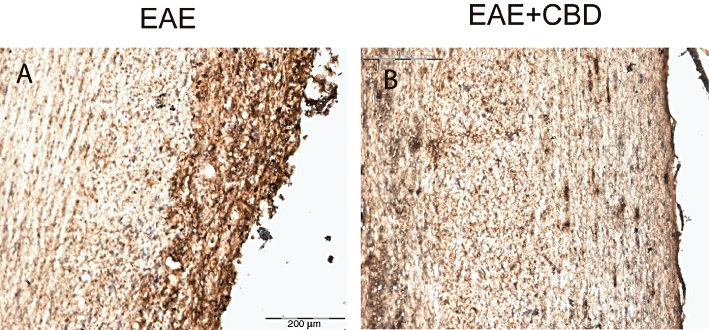
Cannabidiol (CBD) reduces the MOG-induced axonal damage in spinal cords of EAE mice. Mice were killed 30 days after EAE disease induction and sections of spinal cord were analysed for damaged axons using SMI-32 antibody. (A) MOG-induced EAE mice; (B) MOG-induced EAE mice treated with CBD (as described in Figure 1). Scale bar represents 200 µm.
Figure 4.

Cannabidiol (CBD) treatment attenuates EAE-induced immune cell infiltration into spinal cord of mice. Haematoxylin and eosin staining revealed that CBD treatment reduces immune cell infiltration into the white matter of the spinal cords of EAE mice. (A) EAE mice and (B) EAE mice treated with CBD. Scale bar represents 100 µm.
CBD decreases MOG-induced microglial/macrophage activation
As shown above, immunohistochemical staining for the presence of Iba-1 microglial marker showed that Iba-1 is expressed in the spinal cord of control mice at a very low level and that CBD injections to control naïve mice did not affect this basal expression (Figure 2B). Iba-1 expression was, however, dramatically increased on day 30 post MOG immunization (Figure 6A,C). In contrast, a profoundly reduced number of Iba-1 stained cells was present after CBD treatment of these mice (Figure 6B,C). The Iba-1 expression in CBD-treated MOG-induced EAE group remained slightly higher than that observed in naïve control animals showing that CBD treatment did not completely eliminate microglial activation. This result is in agreement with the mild but still present clinical signs as shown in Figure 1.
Mac-2/Galectin-3 is a microglial/macrophage marker whose expression reflects the activation state of these cells. Staining of spinal cord of healthy, naïve animals for Mac-2/Galectin-3 shows absence (or presence of very low number) of activated microglial cells (Figure 7C). CBD injections did not change Mac-2/Galectin-3 expression in these naïve control animals. However, staining of spinal cords derived from EAE mice resulted in dramatically increased number of Mac-2/Galectin-3 positive cells in the spinal cord (Figure 7A,C). This increase was significantly reduced by CBD injections during the onset of clinical EAE (Figure 7B,C).
Figure 7.
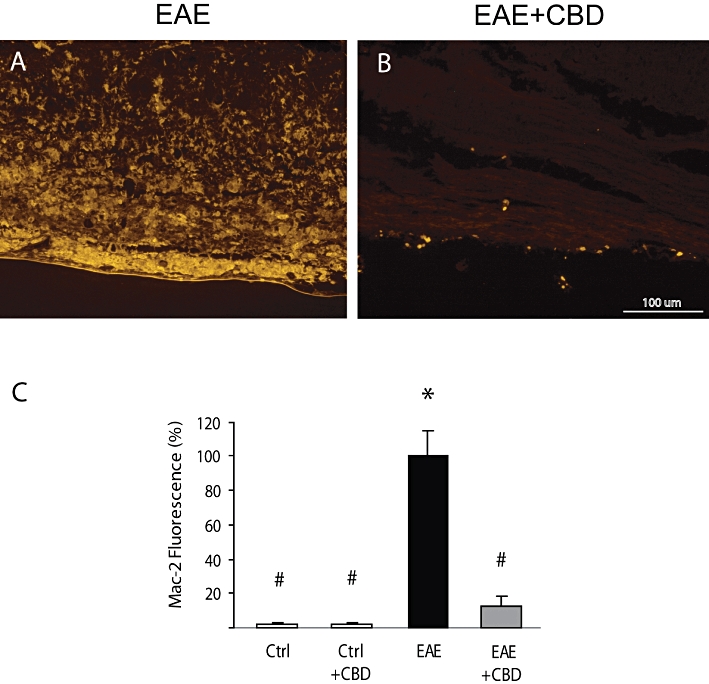
Cannabidiol (CBD) reduces the increased Mac-2/Galectin-3 expression in the spinal cords of EAE mice. Representative pictures showing that (A) MOG injections increase Mac-2 expression and (B) that CBD injections during the onset of EAE significantly decrease the level of Mac-2 staining. (C) Quantification of Mac-2 fluorescence in spinal cord sections of control and EAE mice with and without CBD treatment. The results are presented as mean ± SEM of total fluorescence from 8–11 spinal fragments from three to four mice per each experimental group. One-way anovaF(3,34) = 63.25, P < 0.0001, followed by Bonferroni post hoc test. *P < 0.001 versus Ctrl. #P < 0.001 versus EAE mice. Scale bar represents 100 µm.
CBD inhibits the MOG-induced proliferation of encephalitogenic T cells
Encephalitogenic MOG35-55 T cells were co-cultured with APCs in the presence of MOG at two concentrations (Ben-Nun and Lando, 1983; Kaushansky et al., 2006). We found that 1 µg·mL−1 of MOG induced 24 ± 11-fold increase in [3H]Thymidine incorporation and 2.5 µg·mL−1 produced a 42 ± 10-fold increase in T-cell proliferation. We assayed the effect of CBD on this MOG-induced proliferation. The results presented in Figure 8 show the inhibitory effect of CBD on the proliferation of these encephalitogenic MOG-specific T cells. We found that the proliferation of these cells stimulated with 1 µg·mL−1 (Figure 8A) or 2.5 µg·mL−1 (Figure 8B) of MOG35-55 was significantly down-regulated in the presence of 1–10 µM CBD. Neither SR141716 (a CB1 receptor antagonist) nor SR144528 (a CB2 receptor antagonist), both at 1 µM, affected the proliferative induction observed with MOG35-55 or the antiproliferative effect of CBD (Figure 8 Inset).
Figure 8.
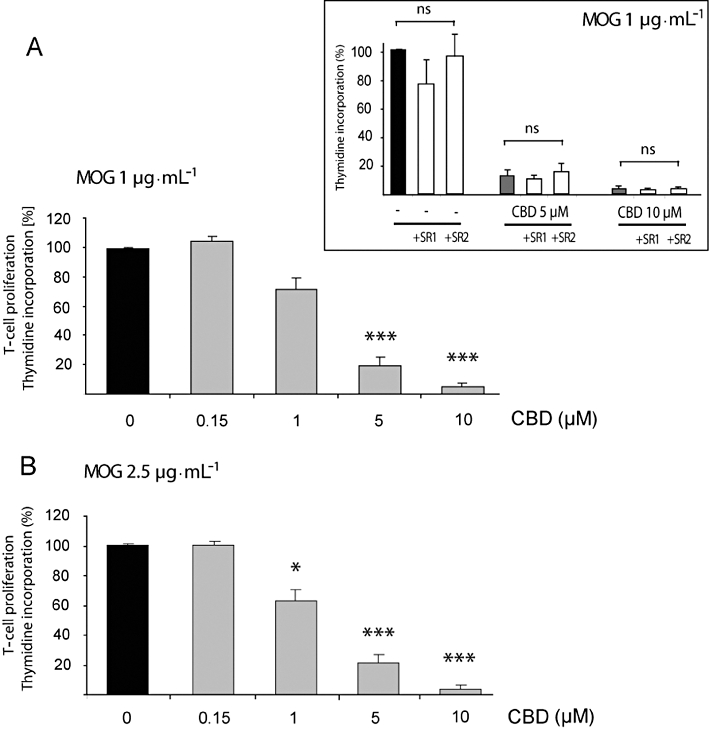
Cannabidiol (CBD) inhibits MOG-induced T-cell proliferation. MOG-induced T-cell proliferation was determined by [3H]thymidine incorporation. Encephalitogenic T cells were co-cultured with antigen presenting cells and exposed to MOG35-55 at 1 (A) and 2.5 µg·mL−1 (B). CBD was added 5 min before MOG. Assays were carried out each time in triplicate and mean per cent values ± SEM are shown based on four independent experiments. (A) One-way anovaF(4,19) = 124.7, P < 0.0001; (B) One-way anovaF(4,19) = 31.1, P < 0.0001, followed by Bonferroni post hoc test. *P < 0.05, ***P < 0.001 versus MOG alone at the respective concentration. Inset: neither the CB1 receptor antagonist (1 µM of SR141716; SR1) nor the CB2 receptor antagonist (1 µM of SR144528; SR2) applied 30 min before CBD and MOG affected MOG35-55 (1 µg·mL−1) – induced T-cell proliferation either in the absence or presence of CBD; one-way anovaF(8,26) = 26.8, P < 0.05; ns, non-significant.
In parallel, we evaluated the effect of CBD on the spontaneous proliferation of T cells (without MOG activation). As described above, this spontaneous proliferation was very low compared with the MOG-induced proliferation. We found that neither 0.15 µM (SI = 1.1 ± 0.0), 5 µM (SI = 1.1 ± 0.0) nor 10 µM (SI = 0.9 ± 0.0) of CBD affected the basal level of proliferation of the T cells in the absence of MOG. Altogether, these results show that CBD selectively inhibited antigen-induced cell proliferation but did not affect basal (non-antigen-induced) proliferation.
Discussion
Our data showed that CBD, when administered systemically for 3 days during clinical EAE disease onset, ameliorated the EAE clinical signs and slowed its progression. The attenuation of EAE progression was accompanied by diminished axonal damage and immune cell infiltration (including T cells and microglial cells) and activation. Using a highly specific autoreactive T-cell line, we observed that CBD decreased the MOG-induced proliferation of these cells.
The effectiveness of cannabinoids in ameliorating the clinical signs of EAE in rodents had been previously shown using mostly mixed affinity CB1/CB2 receptor ligands (THC, WIN552122) in either myelin peptide or virus-induced models (Lyman et al., 1989; Arévalo-Martín et al., 2003; Croxford and Miller, 2003). It should be noted that the immunosuppressive effects of CB1 or mixed CB1/CB2 receptor ligands are mostly evident at doses inducing central effects like sedation and hypothermia (Lyman et al., 1989; Croxford et al., 2008). CBD and its derivatives are devoid of such psychoactive effects and offer a plethora of beneficial activities. Among them, the immunosuppressive, anti-oxidative and neuroprotective properties of CBD are the most relevant for the complex pathology of MS. CBD was reported to be well tolerated when chronically administered to humans for its anxiolytic and antipsychotic activities (Cunha et al., 1980; Consroe et al., 1991). Moreover, CBD (in a mixture with THC, as in Sativex) has been already approved for the treatment of pain and spasticity associated with MS (Zajicek et al., 2003). These features suggest that CBD and CBD derivatives are relatively safe and effective alternatives for the treatment of autoimmune and/or neurodegenerative diseases, such as MS.
Our finding that CBD treatment ameliorates EAE differs from the results by Maresz et al. (2007) which showed that CBD had no beneficial effect in their experimental system. The discrepancy between our results and those of Maresz may result from species differences (C57BL/6 vs. ABH mice) and the antigen used (MOG35-55 peptide vs. spinal cord homogenate) as it is well known that susceptibility, severity and pathology of antigen-induced EAE is highly species and antigen dependent (Shevach, 1999). Moreover, even THC, used in their experiments, had to be given at very high doses of 20 mg·kg−1 and had little or no effect at 5 and 2.5 mg·kg−1. CBD was, however, effective in alleviating signs of other autoimmune inflammatory diseases such as in collagen-induced arthritis (Malfait et al., 2000) and in autoimmune diabetes (Weiss et al., 2008), showing its potential in ameliorating this class of diseases.
Cannabinoids and immune effects
There are many ways in which the cannabinoid system affects immune processes. Cannabinoids were reported to affect the number and function of various immune cells, including T cells, B cells, natural killer cells, dendritic cells, microglia and macrophages in rodents and humans (Nahas et al., 1977; Croxford and Yamamura, 2005; Tanasescu and Constantinescu, 2010). Cannabinoids suppress the production of proinflammatory cytokines including TNFα, Il-1β, IL-2, IL-6, IL-12 and IFN-γ, as well as mitogen-induced cell proliferation, migration, antigen presentation and trafficking into inflamed tissues (Croxford and Yamamura, 2005; Rieder et al., 2010; Tanasescu and Constantinescu, 2010). Because the CB2 receptor is robustly expressed on immune cells (Mackie, 2005), many of these effects were assumed or have been shown to be mediated via the CB2 receptor (Buckley et al., 2000). CB1 receptor-dependent immunosuppressive effects were also observed, despite their moderate expression in immune cells (Cabral et al., 2001).
However, studies with CB1 and CB2 receptor knockout mice as well as the use of specific CB1 or CB2 receptor antagonists and of non-CB1/CB2 receptor ligands, revealed that many anti-inflammatory effects of cannabinoids do not involve the classic CB1/CB2 receptor pathways (Puffenbarger et al., 2000; Kaplan et al., 2003; 2008; Walter et al., 2003; Kozela et al., 2010). Indeed, it is known that non-CB1/CB2 receptor immunomodulatory effects exist and include control of cytokine release (Kaplan et al., 2003; Kozela et al., 2010) and migratory activity of microglial cells (Walter et al., 2003; McHugh et al., 2010). Interestingly, some of the effects of known CB1/CB2 receptor agonists also seem to involve non-CB1/CB2 targets and the effects of non-CB1/CB2 receptor ligands were slightly diminished in cells from CB1/CB2 receptor knockout mice (Kaplan et al., 2003; 2008;) suggesting that several types of cannabinoid targets may complement or interact with each other.
Consequently, CBD, a compound active on non-CB1 or CB2 receptors (Showalter et al., 1996), has been shown to possess a wide anti-inflammatory profile. CBD was shown to decrease TNFα, IL-2 and IFNγ release from activated splenocytes and macrophages (Malfait et al., 2000; Jan et al., 2007; Kaplan et al., 2008). It also suppresses concavalin A and collagen induced T-cell proliferation (Malfait et al., 2000; Jan et al., 2007), microglial migration (Walter et al., 2003) and cytokine release (Kozela et al., 2010). It suppresses antigen-specific antibody production in splenocytes (Jan et al., 2007), as well as attenuates endothelial inflammation and barrier disruption (Rajesh et al., 2007). These activities of CBD may contribute to its beneficial effects in EAE, as observed in our hands, because many of these immune processes were reported to be involved in EAE pathology at different stages of the disease model.
Role of microglial cells in EAE
It is well established that microglia become rapidly activated when the integrity of the CNS is disturbed as a consequence of lesions, neurotoxicity, infections and autoimmune diseases (Hanisch and Kettenmann, 2007). This activation includes the release of a variety of cytotoxic agents, for example, cytokines and chemokines, and of trophic factors that recruit other specialized cells from the periphery into the brain parenchyma. Microglial activation is accompanied by morphological changes and surface expression of various molecules that serve as adhesion molecules, enzymes, complement and immunoglobulin receptors and molecules related to T-cell activation (Hanisch and Kettenmann, 2007). For example, the Iba-1 protein, located within a segment of the major histocompatibility complex class III region, is specifically expressed in macrophages/microglia and its expression is up-regulated during the activation of these cells. Mac-2 is a member of the Galectin-3 family of galactose binding lectins, is another microglia differentiation and activation marker. In this regard, microglial cells expressing Mac-2 are capable of phagocytosis of myelin, while resting microglia are not (Reichert and Rotshenker, 1999).
Microglial cells are considered as key players in MS at various stages of the disease (Ponomarev et al., 2005; Koning et al., 2009). Depletion or blockade of microglia reduces inflammatory lesions, demyelination and EAE disease progression (Huitinga et al., 1990; Heppner et al., 2005). Thus, arrest of microglial function is postulated to be beneficial in EAE/MS treatment. Indeed, we showed here that CBD injections diminished the activation of microglia as measured by Iba-1 and Mac-2 expression in the spinal cord of MOG-injected mice. As mentioned above, although Mac-2 and particularly Iba-1 are generally accepted expression markers for microglia (Reichert and Rotshenker, 1999; Heppner et al., 2005), these proteins are also expressed on some perivascular macrophages and macrophages infiltrating the CNS during pathological conditions. Thus, the inhibitory activity of CBD may apply to microglia as well as to several other macrophage-like cells. Previously, we and others have shown that CBD in vitro decreases the production of TNFα, IL-1β, IL-6 and ROS species in LPS-activated microglia (El-Remessy et al., 2006; Liou et al., 2008; Kozela et al., 2010). Moreover, the microglial production of one of the most potent T-cell attracting chemokines, CCL-2, was also reduced following CBD treatment (Kozela et al., 2010; Juknat et al., 2011). CBD also reduces the migration of microglial cells (Walter et al., 2003). These data suggest that CBD is a potent inhibitor of various functions of microglial cells. Thus, by silencing microglial cells, CBD may be able to prevent further inflammatory processes within the CNS parenchyma, that is, myelin antigen exposure, T-cell infiltration, cytokine/chemokine release and recruitment of other immune cells.
Antiproliferative effects of CBD
We showed that CBD decreased MOG-induced encephalitogenic T-cell proliferation. This antiproliferative effect of CBD was not mediated via CB1 or CB2 receptors. High proliferation rate is a core response of immune cells (including T cells) following various stimuli, such as cytokines, mitogens and antigens presented by APCs. With regard to EAE or MS, it was shown that myelin-specific T-cell clones penetrate the blood-brain barrier and the CNS parenchyma initiating demyelination. We observed that CBD inhibited the MOG-induced proliferation of encephalitogenic T cells isolated from mice, previously immunized against MOG. This potent antiproliferative activity of CBD probably contributes significantly to the EAE-ameliorating effects of CBD and is in agreement with the lower number of T cells present in the spinal cords of CBD-treated EAE mice. Indeed, CBD has been reported to induce apoptosis in transformed T-cell lines (McKallip et al., 2006; Lee et al., 2008) as well as in primary splenic lymphocytes including CD4+, CD8+ and B220+ subsets (Lee et al., 2008; Wu et al., 2008). It had a similar apoptotic activity towards lymphocytes in peripheral blood (Ignatowska-Jankowska et al., 2009). Moreover, concavalin A-induced (Jan et al., 2007) and collagen-induced stimulation of T cells (Malfait et al., 2000) was inhibited by CBD. On the other hand, primary monocytes were observed to be resistant to the apoptotic effects of CBD (Gallily et al., 2003). This difference in apoptotic susceptibility of various immune cell types to CBD is of considerable significance but its mechanism is still unknown.
Anti-oxidant and neuroprotective properties of CBD
Several cannabinoids including THC and CBD exert anti-oxidative and neuroprotective properties (Mechoulam et al., 2007). Most of the current MS therapies are directed against various immune cells to achieve immunosuppressive effects. However, increasing evidence shows that immunosuppression alone was not sufficient for therapeutic effect especially in late, secondary progressive MS (Bennett and Stüve, 2009; Jones and Coles, 2010). In these cases, the neurodegenerative processes become resistant to immunomodulation. Indeed, neurodegeneration that consists of neuronal and axonal loss can result from oxidative stress and excitotoxicity and is driven by activated microglial cells and macrophagic/monocytic infiltrates (Hanisch and Kettenmann, 2007). It appears that the cannabinoid system could provide a rescue mechanism in such conditions. Accordingly, the ameliorating activity of THC-like cannabinoids combines CB2 receptor-mediated inhibition of autoreactive T cells and CB1 receptor-mediated neuroprotective activity on neurons (Maresz et al., 2007). Similarly, Croxford et al. (2008) pointed out that cannabinoid-mediated neuroprotection rather than immunosuppression, was relevant for the recovery process at the later, remissive stages of MS.
The main source of reactive oxygen and nitrogen species within the CNS under neurodegenerative conditions are reactive microglia and astrocytes (Hanisch and Kettenmann, 2007). Oxidative signalling negatively affects neuronal and axonal survival, especially of axons lacking a myelin sheath, and results in irreversible damage. Accordingly, deletion of nuclear factor-erythroid 2-related 2 (Nrf2), a redox-sensitive transcription factor that regulates expression of many protective antioxidant and detoxication enzymes and transporters (Kobayashi and Yamamoto, 2005), resulted in exacerbation of EAE (Johnson et al., 2010). CBD has been frequently described as a potent neuroprotective and anti-oxidant agent. CBD reduced glutamate excitotoxicity and hydroperoxide-induced oxidative neuronal damage (Hampson et al., 1998) and provided neuroprotection against 6-hydroxy-dopamine (Lastres-Becker et al., 2005), β-amyloid (Iuvone et al., 2004) and prion toxicities to neurons (Dirikoc et al., 2007) and against neuroinflammation (Esposito et al., 2007; Zuardi, 2008). In this regard, we have shown that exposure of microglial cells in culture to CBD up-regulates a number of anti-oxidative genes including genes that are involved in glutathione synthesis. Many of these genes are under the control of the Nrf2 transcription factor (Juknat et al., 2011). Thus, the anti-oxidative properties of CBD may significantly contribute to the alleviation of EAE pathology and accompany its anti-inflammatory beneficial properties.
In summary, we have shown that CBD administered to MOG-immunized C57BL/6 mice, at the onset of EAE disease, reduced the severity of the clinical signs of EAE. CBD treatment was accompanied by diminished axonal loss and inflammation (infiltration of T cells and microglial activation). Moreover, CBD prevented proliferation of myelin-specific T cells in vitro. These observations suggest that CBD may have potential for alleviating MS-like pathology.
Acknowledgments
This work was supported by the Dr Miriam and Sheldon G. Adelson Center for the Biology of Addictive Diseases and by the Dr Miriam and Sheldon G. Adelson Medical Research Foundation. A. J. and N. R. were supported by the Center for Absorption in Science in Israel.
Glossary
Abbreviations
- CBD
cannabidiol
- EAE
experimental autoimmune encephalomyelitis
- MOG
myelin oligodendrocyte glycoprotein
Conflict of interest
The authors state no conflict of interest.
Supporting Information
Teaching Materials; Figs 1–8 as PowerPoint slide.
References
- Alexander SPH, Mathie A, Peters JA. Guide to Receptors and Channels (GRAC), 4th edn. Br J Pharmacol. 2009;158:S1–S254. doi: 10.1111/j.1476-5381.2009.00499.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arévalo-Martín A, Vela JM, Molina-Holgado E, Borrell J, Guaza C. Therapeutic action of cannabinoids in a murine model of multiple sclerosis. J Neurosci. 2003;23:2511–2516. doi: 10.1523/JNEUROSCI.23-07-02511.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayewitch M, Avidor-Reiss T, Levy R, Barg J, Mechoulam R, Vogel Z. The peripheral cannabinoid receptor: adenylate cyclase inhibition and G protein coupling. FEBS Lett. 1995;375:143–147. doi: 10.1016/0014-5793(95)01207-u. [DOI] [PubMed] [Google Scholar]
- Bennett JL, Stüve O. Update on inflammation, neurodegeneration, and immunoregulation in multiple sclerosis: therapeutic implications. Clin Neuropharmacol. 2009;32:121–132. doi: 10.1097/WNF.0b013e3181880359. [DOI] [PubMed] [Google Scholar]
- Ben-Nun A, Lando Z. Detection of autoimmune cells proliferating to myelin basic protein and selection of T cell lines that mediate experimental autoimmune encephalomyelitis (EAE) in mice. J Immunol. 1983;130:1205–1209. [PubMed] [Google Scholar]
- Buckley NE, McCoy KL, Mezey E, Bonner T, Zimmer A, Felder CC, et al. Immunomodulation by cannabinoids is absent in mice deficient for the cannabinoid CB(2) receptor. Eur J Pharmacol. 2000;396:141–149. doi: 10.1016/s0014-2999(00)00211-9. [DOI] [PubMed] [Google Scholar]
- Cabral GA, Harmon KN, Carlisle SJ. Cannabinoid-mediated inhibition of inducible nitric oxide production by rat microglial cells: evidence for CB1 receptor participation. Adv Exp Med Biol. 2001;493:207–214. doi: 10.1007/0-306-47611-8_24. [DOI] [PubMed] [Google Scholar]
- Compston A, Coles A. Multiple sclerosis. Lancet. 2008;372:1502–1517. doi: 10.1016/S0140-6736(08)61620-7. [DOI] [PubMed] [Google Scholar]
- Consroe P, Laguna J, Allender J, Snider S, Stern L, Sandyk R, et al. Controlled clinical trial of cannabidiol in Huntington's disease. Pharmacol Biochem Behav. 1991;40:701–708. doi: 10.1016/0091-3057(91)90386-g. [DOI] [PubMed] [Google Scholar]
- Croxford JL, Miller SD. Immunoregulation of a viral model of multiple sclerosis using the synthetic cannabinoid R+WIN55,212. J Clin Invest. 2003;111:1231–1240. doi: 10.1172/JCI17652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Croxford JL, Yamamura T. Cannabinoids and the immune system: potential for the treatment of inflammatory diseases? J Neuroimmunol. 2005;166:3–18. doi: 10.1016/j.jneuroim.2005.04.023. [DOI] [PubMed] [Google Scholar]
- Croxford JL, Pryce G, Jackson SJ, Ledent C, Giovannoni G, Pertwee RG, et al. Cannabinoid-mediated neuroprotection, not immunosuppression, may be more relevant to multiple sclerosis. J Neuroimmunol. 2008;193:120–129. doi: 10.1016/j.jneuroim.2007.10.024. [DOI] [PubMed] [Google Scholar]
- Cunha JM, Carlini EA, Pereira AE, Ramos OL, Pimentel C, Gagliardi R, et al. Chronic administration of cannabidiol to healthy volunteers and epileptic patients. Pharmacology. 1980;21:175–185. doi: 10.1159/000137430. [DOI] [PubMed] [Google Scholar]
- Dirikoc S, Priola SA, Marella M, Zsürger N, Chabry J. Nonpsychoactive cannabidiol prevents prion accumulation and protects neurons against prion toxicity. J Neurosci. 2007;27:9537–9544. doi: 10.1523/JNEUROSCI.1942-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Remessy AB, Al-Shabrawey M, Khalifa Y, Tsai NT, Caldwell RB, Liou GI. Neuroprotective and blood-retinal barrier-preserving effects of cannabidiol in experimental diabetes. Am J Pathol. 2006;168:235–244. doi: 10.2353/ajpath.2006.050500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esposito G, Scuderi C, Savani C, Steardo L, Jr, De Filippis D, Cottone P, et al. Cannabidiol in vivo blunts β-amyloid induced neuroinflammation by suppressing IL-1 β and iNOS expression. Br J Pharmacol. 2007;151:1272–1279. doi: 10.1038/sj.bjp.0707337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallily R, Even-Chena T, Katzavian G, Lehmann D, Dagan A, Mechoulam R. γ-irradiation enhances apoptosis induced by cannabidiol, a non-psychotropic cannabinoid, in cultured HL-60 myeloblastic leukemia cells. Leuk Lymphoma. 2003;44:1767–1773. doi: 10.1080/1042819031000103917. [DOI] [PubMed] [Google Scholar]
- Hampson AJ, Grimaldi M, Axelrod J, Wink D. Cannabidiol and (-)Delta9-tetrahydrocannabinol are neuroprotective antioxidants. Proc Natl Acad Sci U S A. 1998;95:8268–8273. doi: 10.1073/pnas.95.14.8268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanisch UK, Kettenmann H. Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nat Neurosci. 2007;10:1387–1394. doi: 10.1038/nn1997. [DOI] [PubMed] [Google Scholar]
- Heppner FL, Greter M, Marino D, Falsig J, Raivich G, Hovelmeyer N, et al. Experimental autoimmune encephalomyelitis repressed by microglial paralysis. Nat Med. 2005;11:146–152. doi: 10.1038/nm1177. [DOI] [PubMed] [Google Scholar]
- Huitinga I, Van Rooijen N, De Groot CJ, Uitdehaag BM, Dijkstra CD. Suppression of experimental allergic encephalomyelitis in Lewis rats after elimination of macrophages. J Exp Med. 1990;172:1025–1033. doi: 10.1084/jem.172.4.1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ignatowska-Jankowska B, Jankowski M, Glac W, Swiergel AH. Cannabidiol-induced lymphopenia does not involve NKT and NK cells. J Physiol Pharmacol. 2009;3(Suppl):99–103. [PubMed] [Google Scholar]
- Iuvone T, Esposito G, Esposito R, Santamaria R, Di Rosa M, Izzo AA. Neuroprotective effect of cannabidiol, a non-psychoactive component from Cannabis sativa, on β-amyloid-induced toxicity in PC12 cells. J Neurochem. 2004;89:134–141. doi: 10.1111/j.1471-4159.2003.02327.x. [DOI] [PubMed] [Google Scholar]
- Jack C, Ruffini F, Bar-Or A, Antel JP. Microglia and multiple sclerosis. J Neurosci Res. 2005;81:363–373. doi: 10.1002/jnr.20482. [DOI] [PubMed] [Google Scholar]
- Jan TR, Su ST, Wu HY, Liao MH. Suppressive effects of cannabidiol on antigen-specific antibody production and functional activity of splenocytes in ovalbumin-sensitized BALB/c mice. Int Immunopharmacol. 2007;7:773–780. doi: 10.1016/j.intimp.2007.01.015. [DOI] [PubMed] [Google Scholar]
- Járai Z, Wagner JA, Varga K, Lake KD, Compton DR, Martin BR, et al. Cannabinoid-induced mesenteric vasodilation through an endothelial site distinct from CB1 or CB2 receptors. Proc Natl Acad Sci U S A. 1999;96:14136–14141. doi: 10.1073/pnas.96.24.14136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson DA, Amirahmadi S, Ward C, Fabry Z, Johnson JA. The absence of the pro-antioxidant transcription factor Nrf2 exacerbates experimental autoimmune encephalomyelitis. Toxicol Sci. 2010;114:237–246. doi: 10.1093/toxsci/kfp274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones JL, Coles AJ. New treatment strategies in multiple sclerosis. Exp Neurol. 2010;225:34–39. doi: 10.1016/j.expneurol.2010.06.003. [DOI] [PubMed] [Google Scholar]
- Juknat A, Pietr M, Kozela E, Rimmerman N, Levy R, Coppola G, et al. Differential transcriptional profiles mediated by exposure to the cannabinoids cannabidiol and Δ9-tetrahydrocannabinol in BV-2 microglial cells. Br J Pharm. 2011 doi: 10.1111/j.1476-5381.2011.01461.x. DOI: 10.1111/j.1476-5381.2011.01461.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan BL, Rockwell CE, Kaminski NE. Evidence for cannabinoid receptor-dependent and -independent mechanisms of action in leukocytes. J Pharmacol Exp Ther. 2003;306:1077–1085. doi: 10.1124/jpet.103.051961. [DOI] [PubMed] [Google Scholar]
- Kaplan BL, Springs AE, Kaminski NE. The profile of immune modulation by cannabidiol (CBD) involves deregulation of nuclear factor of activated T cells (NFAT) Biochem Pharmacol. 2008;76:726–737. doi: 10.1016/j.bcp.2008.06.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaushansky N, Zhong MC, Kerlero de Rosbo N, Hoeftberger R, Lassmann H, Ben-Nun A. Epitope specificity of autoreactive T and B cells associated with experimental autoimmune encephalomyelitis and optic neuritis induced by oligodendrocyte-specific protein in SJL/J mice. J Immunol. 2006;177:7364–7376. doi: 10.4049/jimmunol.177.10.7364. [DOI] [PubMed] [Google Scholar]
- Klein TW, Newton CA. Therapeutic potential of cannabinoid-based drugs. Adv Exp Med Biol. 2007;601:395–413. doi: 10.1007/978-0-387-72005-0_43. [DOI] [PubMed] [Google Scholar]
- Kobayashi M, Yamamoto M. Molecular mechanisms activating the Nrf2-Keap1 pathway of antioxidant gene regulation. Antioxid Redox Signal. 2005;7:385–394. doi: 10.1089/ars.2005.7.385. [DOI] [PubMed] [Google Scholar]
- Koning N, Uitdehaag BM, Huitinga I, Hoek RM. Restoring immune suppression in the multiple sclerosis brain. Prog Neurobiol. 2009;89:359–368. doi: 10.1016/j.pneurobio.2009.09.005. [DOI] [PubMed] [Google Scholar]
- Kozela E, Pietr M, Juknat A, Rimmerman N, Levy R, Vogel Z. Cannabinoids Delta(9)-tetrahydrocannabinol and cannabidiol differentially inhibit the lipopolysaccharide-activated NF-kappaB and interferon- β/STAT proinflammatory pathways in BV-2 microglial cells. J Biol Chem. 2010;285:1616–1626. doi: 10.1074/jbc.M109.069294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lastres-Becker I, Molina-Holgado F, Ramos JA, Mechoulam R, Fernández-Ruiz J. Cannabinoids provide neuroprotection against 6-hydroxydopamine toxicity in vivo and in vitro: relevance to Parkinson's disease. Neurobiol Dis. 2005;19:96–107. doi: 10.1016/j.nbd.2004.11.009. [DOI] [PubMed] [Google Scholar]
- Lee CY, Wey SP, Liao MH, Hsu WL, Wu HY, Jan TR. A comparative study on cannabidiol-induced apoptosis in murine thymocytes and EL-4 thymoma cells. Int Immunopharmacol. 2008;8:732–740. doi: 10.1016/j.intimp.2008.01.018. [DOI] [PubMed] [Google Scholar]
- Lev N, Barhum Y, Melamed E, Offen D. Bax-ablation attenuates experimental autoimmune encephalomyelitis in mice. Neurosci Lett. 2004;359:139–142. doi: 10.1016/j.neulet.2004.01.076. [DOI] [PubMed] [Google Scholar]
- Liou GI, Auchampach JA, Hillard CJ, Zhu G, Yousufzai B, Mian S, et al. Mediation of cannabidiol anti-inflammation in the retina by equilibrative nucleoside transporter and A2A adenosine receptor. Invest Ophthalmol Vis Sci. 2008;49:5526–5531. doi: 10.1167/iovs.08-2196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyman WD, Sonett JR, Brosnan CF, Elkin R, Bornstein MB. Delta 9-tetrahydrocannabinol: a novel treatment for experimental autoimmune encephalomyelitis. J Neuroimmunol. 1989;23:73–81. doi: 10.1016/0165-5728(89)90075-1. [DOI] [PubMed] [Google Scholar]
- McHugh D, Hu SS, Rimmerman N, Juknat A, Vogel Z, Walker JM, et al. N-arachidonoyl glycine, an abundant endogenous lipid, potently drives directed cellular migration through GPR18, the putative abnormal cannabidiol receptor. BMC Neurosci. 2010;11:44. doi: 10.1186/1471-2202-11-44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKallip RJ, Jia W, Schlomer J, Warren JW, Nagarkatti PS, Nagarkatti M. Cannabidiol-induced apoptosis in human leukemia cells: a novel role of cannabidiol in the regulation of p22phox and nox4 expression. Mol Pharmacol. 2006;70:897–908. doi: 10.1124/mol.106.023937. [DOI] [PubMed] [Google Scholar]
- Mackie K. Distribution of cannabinoid receptors in the central and peripheral nervous system. Handb Exp Pharmacol. 2005;168:299–325. doi: 10.1007/3-540-26573-2_10. [DOI] [PubMed] [Google Scholar]
- Malfait AM, Gallily R, Sumariwalla PF, Malik AS, Andreakos E, Mechoulam R, et al. The nonpsychoactive cannabis constituent cannabidiol is an oral anti-arthritic therapeutic in murine collagen-induced arthritis. Proc Natl Acad Sci U S A. 2000;97:9561–9566. doi: 10.1073/pnas.160105897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maresz K, Carrier EJ, Ponomarev ED, Hillard CJ, Dittel BN. Modulation of the cannabinoid CB2 receptor in microglial cells in response to inflammatory stimuli. J Neurochem. 2005;95:437–445. doi: 10.1111/j.1471-4159.2005.03380.x. [DOI] [PubMed] [Google Scholar]
- Maresz K, Pryce G, Ponomarev ED, Marsicano G, Croxford JL, Shriver LP, et al. Direct suppression of CNS autoimmune inflammation via the cannabinoid receptor CB1 on neurons and CB2 on autoreactive T cells. Nat Med. 2007;13:492–497. doi: 10.1038/nm1561. [DOI] [PubMed] [Google Scholar]
- Mechoulam R, Peters M, Murillo-Rodriguez E, Hanus LO. Cannabidiol–recent advances. Chem Biodivers. 2007;4:1678–1692. doi: 10.1002/cbdv.200790147. [DOI] [PubMed] [Google Scholar]
- Nahas GG, Morishima A, Desoize B. Effects of cannabinoids on macromolecular synthesis and replication of cultured lymphocytes. Fed Proc. 1977;36:1748–1752. [PubMed] [Google Scholar]
- Pietr M, Kozela E, Levy R, Rimmerman N, Lin YH, Stella N, et al. Differential changes in GPR55 during microglial cell activation. FEBS Lett. 2009;583:2071–2076. doi: 10.1016/j.febslet.2009.05.028. [DOI] [PubMed] [Google Scholar]
- Ponomarev ED, Shriver LP, Maresz K, Dittel BN. Microglial cell activation and proliferation precedes the onset of CNS autoimmunity. J Neurosci Res. 2005;81:374–389. doi: 10.1002/jnr.20488. [DOI] [PubMed] [Google Scholar]
- Puffenbarger RA, Boothe AC, Cabral GA. Cannabinoids inhibit LPS-inducible cytokine mRNA expression in rat microglial cells. Glia. 2000;29:58–69. [PubMed] [Google Scholar]
- Rajesh M, Pan H, Mukhopadhyay P, Ba' tkai S, Osei-Hyiaman D, Hasko G, et al. Cannabidiol attenuates high glucose-induced endothelial cell inflammatory response and barrier disruption. J Leukoc Biol. 2007;82:1382–1389. [Google Scholar]
- Reichert F, Rotshenker S. Galectin-3/MAC-2 in experimental allergic encephalomyelitis. Exp Neurol. 1999;160:508–514. doi: 10.1006/exnr.1999.7229. [DOI] [PubMed] [Google Scholar]
- Rieder SA, Chauhan A, Singh U, Nagarkatti M, Nagarkatti P. Cannabinoid-induced apoptosis in immune cells as a pathway to immunosuppression. Immunobiology. 2010;215:598–560. doi: 10.1016/j.imbio.2009.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shevach EM. Animal models for autoimmune and inflammatory disease. In: Coligan JE, Kruisbeek AM, Margulies DH, Shevach EM, Strober W, editors. Current Protocols in Immunology. New York: John Wiley & Sons, Inc; 1999. pp. 15.0.1–15.0.5. [Google Scholar]
- Showalter VM, Compton DR, Martin BR, Abood ME. Evaluation of binding in a transfected cell line expressing a peripheral cannabinoid receptor (CB2): identification of cannabinoid receptor subtype selective ligands. J Pharmacol Exp Ther. 1996;278:989–999. [PubMed] [Google Scholar]
- Tanasescu R, Constantinescu CS. Cannabinoids and the immune system: an overview. Immunobiology. 2010;215:588–597. doi: 10.1016/j.imbio.2009.12.005. [DOI] [PubMed] [Google Scholar]
- Walter L, Franklin A, Witting A, Wade C, Xie Y, Kunos G, et al. Nonpsychotropic cannabinoid receptors regulate microglial cell migration. J Neurosci. 2003;23:1398–1405. doi: 10.1523/JNEUROSCI.23-04-01398.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiss L, Zeira M, Reich S, Slavin S, Raz I, Mechoulam R, et al. Cannabidiol arrests onset of autoimmune diabetes in NOD mice. Neuropharmacology. 2008;54:244–249. doi: 10.1016/j.neuropharm.2007.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu HY, Chu RM, Wang CC, Lee CY, Lin SH, Jan TR. Cannabidiol-induced apoptosis in primary lymphocytes is associated with oxidative stress-dependent activation of caspase-8. Toxicol Appl Pharmacol. 2008;226:260–270. doi: 10.1016/j.taap.2007.09.012. [DOI] [PubMed] [Google Scholar]
- Zajicek J, Fox P, Sanders H, Wright D, Vickery J, Nunn A, et al. Cannabinoids for treatment of spasticity and others symptoms related to multiple sclerosis (CAMS study): multicenter randomized placebo-controlled trial. Lancet. 2003;362:1517–1526. doi: 10.1016/S0140-6736(03)14738-1. [DOI] [PubMed] [Google Scholar]
- Zuardi AW. Cannabidiol: from an inactive cannabinoid to a drug with wide spectrum of action. Rev Bras Psiquiatr. 2008;30:271–280. doi: 10.1590/s1516-44462008000300015. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.