

Abstract
Asthma, a chronic inflammatory disease is typically characterized by bronchoconstriction and airway hyper-reactivity. A wealth of studies applying chemistry, molecular and cell biology to animal model systems and human asthma over the last decade have revealed that asthma is associated with increased synthesis of the gaseous molecule nitric oxide (NO). The high NO levels in the oxidative environment of the asthmatic airway lead to greater formation of reactive nitrogen species (RNS) and subsequent oxidation and nitration of proteins, which adversely affect protein functions that are biologically relevant to chronic inflammation. In contrast to the high levels of NO and nitrated products, there are lower levels of beneficial S-nitrosothiols (RSNO), which mediate bronchodilation, due to greater enzymatic catabolism of RSNO in the asthmatic airways. This review discusses the rapidly accruing data linking metabolic products of NO as critical determinants in the chronic inflammation and airway reactivity of asthma.
Introduction
Asthma is a chronic disease of airway inflammation, typically characterized by bronchial hyper-responsiveness, airflow limitation and mucus hyper-secretion, leading to the symptoms of cough, wheezing and shortness of breath [1–3]. Although many pathologic processes participate in asthma pathogenesis, a large body of evidence supports an important role for airway-derived nitric oxide (NO) in the mechanism underlying asthma. NO has effects as a bronchodialator via relaxation of airway smooth muscle [4]. NO binds to soluble Guanylate Cyclase (sGC); this leads to activation of the protein, followed by enhanced production of intracellular messenger cGMP, which is responsible for initiating downstream signaling for vasodilation [5]. Studies have established that sGC expression and activity are reduced in experimental asthma, and that this contributes to the observed airway hyperreactivity [6]. The sGC mRNA expression is transcrptionally regulated in culured cells by inflammatory stimuli, e.g. lipopolysaccharide, IL-1β or cytokines mixture. It may be that the high levels of inflammatory cytokines in asthmatic airways contribute to the lower level of sGC expression [6–8]. Furthermore, studies also identify an NO/cysteine interaction, in addition to the classical heme iron-NO activation, that is important in mediating activation of sGC; formation of a putative thiol-NO adduct apparently enables maximal sGC activation. However under oxidative stress, the thiol is oxidized, which prevents NO binding and retards sGC activation [9]. Thus, the oxidative stress in the asthmatic airway may contribute to deficient sGC-mediated bronchodilation through both the loss of sGC expression as well as cysteine oxidation that consequently abrogates maximal sGC activation via NO-cysteine interactions [9]. Considerable evidence supports detrimental effects of NO in airway inflammation through formation of reactive nitrogen species (RNS) that mediate inflammation and injury [1, 4]. Increased generation of RNS and reactive oxygen species (ROS) are well documented in asthma [10–12]. The asthmatic airways are infiltrated with eosinophils and neutrophils, which contain peroxidases, eg. eosinophil-peroxidase (EPO) and myeloperoxidase (MPO) [13, 14]. The peroxidases use chloride and/or bromide present in tissues to form hypohalous acids HOCl and HOBr [15–17]. Hypohalous acids are potent oxidants and chlorinate and/or brominate protein tyrosines to form 3-chlorotyrosine (Cl-Y) and 3-bromotyrosine [17] markers of MPO and EPO activity, respectively. MPO also uses the oxidation of nitrite (an oxidation product of NO) to generate the nitrogen dioxide radical [14, 15, 18] to nitrate protein tyrosine and form 3-nitrotyrosine [16, 18–20]. The detection of 3-nitrotyrosine in the lung of asthmatic patients provides definitive evidence for numerous biochemical targets of NO that are likely to have functional consequences in the asthmatic airway [13, 14, 18, 21]. This review will discuss the biochemical reactions of NO and its metabolic products in the pathophysiology of asthma.
Asthma and Oxidative Stress
Reactive oxygen species (ROS) are highly reactive molecules with unpaired electrons that can quickly react with other chemical compounds, potentially altering their structure and function [22–24]. Superoxide (O2•−), hydrogen peroxide (H2O2) and hydroxyl radicals (•OH) are potent ROS in biological systems (Fig 1). O2•− is unstable and reacts rapidly with proteins that contain transition-metal prosthetic groups, such as heme moieties or iron–sulfur clusters [25, 26]. Under normal condition, a very fine balance is maintained between ROS production and protection from oxidative injury by antioxidants, which include non-enzymatic (eg. vitamins and thiols) and enzymatic (eg. catalase, superoxide dismutases and glutathione peroxidase) mechanisms. During diseased conditions, an increased ROS production and/or reduced antioxidant defenses create an imbalance, resulting in oxidative stress, a final common pathway that generates and maintains inflammation and injury via proinflammatory cytokine release and altered enzymatic function [24]. Thus, oxidative stress, a condition representing an imbalance between oxidants and antioxidants in favor of the oxidants, is found to be involved in aging and various diseases, such as cancer, stroke, cardiovascular diseases and diabetes [27]. Measure of stable byproducts of oxidation has documented that oxidative stress is present in asthma. In a study by Ercan et al., children with asthma had higher plasma levels of malondialdehyde (MDA) and 8-isoprostanes (lipid peroxidation end product) compared with healthy controls and the MDA levels were higher in children with more severe asthma [28]. Acute asthma exacerbations worsen the degree of oxidative burden. Patients hospitalized for an acute asthma exacerbation, have higher levels of urinary oxidative metabolites and total plasma antioxidant capacity is also reduced during these episodes [29–31]. Furthermore, asthmatic subjects with more obstructed airways also have a higher degree of oxidative stress [32].
Fig. 1. Production of reactive oxygen species (ROS).

Superoxide (O2•−) reacts rapidly with itself, or is catalytically converted to form hydrogen peroxide (H2O2) by superoxide dismutase. Under pathologic conditon, extremely toxic reactions of superoxide and hydrogen peroxide which form hydroxyl radical occur via the Haber–Weiss and Fenton chemistry reactions in the presence of metal ions. Hydrogen peroxide is converted by myeloperoxide (MPO) or eosinophil peroxidase (EPO) to highly reactive halogenating acids, such as hypobromous acid (HOX, X=Br/Cl) or hypochlorous acid, which can further react with superoxide to produce halides and hydroxyl radical.
Over the past two decades, a multitude of studies have revealed that in addition to oxidative stress in asthma, there is nitrative stress related to an increase of nitric oxide (NO) and its reaction products. NO, a relatively stable free radical, is increased in exhaled air of asthmatic individuals as compared to healthy individuals [33]. Individuals with asthma have 3-fold higher than normal NO concentrations in the lower airway and in the exhaled breath [14, 34–36]. Exhaled NO levels are inversely correlated with airflow parameters in asthmatic patients [37–41]. Exhaled NO in asthmatics increases during the late asthmatic response after experimental Ag challenge, a method used to mimic asthma attacks and study the mechanisms of episodic asthma in human subjects [34, 42].
Nitric Oxide Synthases in Asthma
While antioxidants remove superoxide (O=O•), there is no known nitric oxide (N=O•) removal system. NO produced in the lungs is an important physiologic regulator of airway events, including airway tone, pulmonary vascular tone, mucin secretion, mucociliary clearance through effects on ciliary beat frequency and immune surveillance including tumoricidal and bactericidal effects [43–46].
Nitric oxide is formed when one of the chemically equivalent guanido groups of the essential amino acid L-arginine (L-arg) is oxidized by five electrons, forming NO• and L-citrulline [47]. The reaction is catalyzed by enzymes nitric oxide synthases (NOS) with the help of several co-factors, including flavones (FAD, FMN), tetrahydrobiopterin (BH4) and NADPH. The NOS exist in several isoforms (Table 1); all isoforms are stereospecific and active as homodimers [48]. NOS1 and NOS3, originally identified in neuronal and endothelial cells, respectively, depend on increases in calcium to bind calmodulin, leading to enzyme activation and picomolar levels of NO production [44]. NOS2 is inducible in diverse cell types by cytokines and contains calmodulin as a subunit, allowing the production of nanomolar levels of NO at resting levels of intracellular calcium [44]. All isoforms are present in the lung [49, 50].
Table 1.
Nitric Oxide Synthases.
| Isoform | Chromosomal localization | Expression | Activity |
|---|---|---|---|
| nNOS/NOS1 | 12q24. | Constitutive | Ca2+ dependent |
| iNOS/NOS2 | 17cen-q12 | Inducible | Ca2+ independent |
| eNOS/NOS3 | 7q35-36 | Constitutive | Ca2+ dependent |
To investigate the role of NO in asthma, mice with targeted deletions of the three known isoforms of NOS (NOS1, 2, and 3) were studied using an established model of allergic asthma, the ovalbumin (OVA)-sensitized and challenged (OVA/OVA) mouse model. NOS2 isoform was significantly upregulated in the lungs of OVA/OVA wild-type (WT) mice and was undetectable in similarly treated NOS2-deficient mice. However airway hyperresponsiveness between these two groups were found to be similar, perhaps due to limitations of deletion models when there is genetic redundancy. Airway responsiveness in NOS1-deficient and NOS1/3-deficient mice was significantly less than that observed in WT mice. Overall, these studies support that NOS genotypes are associated with asthma [51]. In humans, the higher than normal NO concentrations in exhaled breath of is closely associated with increased transcriptional activation of the NOS2 gene [35, 52].
Airway epithelium expresses NOS2 in the healthy lung [34, 53, 54], where the high levels of NO are involved in innate host defense [46]. The NOS2 protein and activity are greatly increased in asthmatic airways [35, 55, 56] and inhibition of NOS2-derived NO attenuates antigen-induced airway constriction, inflammatory, and remodeling processes, reducing both collagen and elastic fiber deposition in a guinea pig model of allergic asthma [57, 58]. NOS2 gene expression is substantially regulated at the level of transcription [45, 46, 59–62]. The expression of NOS2 is driven by a combination of the interferon-γ (IFN) activation of Janus kinase (JAK)/signal transducer and activator of transcription 1 (STAT-1) and activation of nuclear factor NF- κβ and AP-1 [44, 60–62]. IFNγ signal transduction is the primary molecular event required for the expression of NOS2 in airway human epithelia [54, 59, 63, 64].
L-Arginine Metabolism By Arginase Regulates NO Synthesis
Model systems of allergic airway inflammation and studies in human asthma reveal that L-arg metabolism is altered in asthma [65–75]. L-arg the substrate for NO synthases (NOS) is also the substrate for arginases [68, 76, 77]. The activity of NOS and arginases are coordinately regulated by L-arg availability (Fig. 2) [74].
Fig. 2. Molecular consequences of nitrative stress.
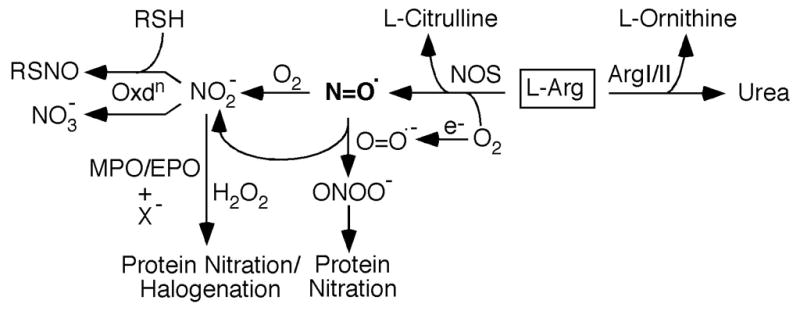
Nitric oxide synthase used L-Arg as substrate to produce nitric oxide (N=O•); L-Citrulline is propduced as the by-product of this reaction. Arginases I and II use L-Arg as substrate to generate urea and L-ornithine. Nitric oxide can rapidly oxidize to nitrite (NO2−), which can be further oxidized to nitrate (NO3−). Superoxide (O2•−) reacts rapidly with nitric oxide to produce peroxynitrite (ONOO−), which can readily nitrate proteins. In presence of hydrogen peroxide (H2O2), nitrite (NO2−) and/or halide (X−), myeloperoxidase/eosinophil peroxidase system can also promote protein nitration and/or halogenation. In presence of thiols (RSH), nitrite is also involved in nitrosylation reaction to produce S-nitrosothiols (RSNO).
Asthmatic patients with acute asthma exacerbation have higher serum arginase activity and lower levels of L-arg as compared with healthy control subjects [71]. When L-arg is subsequently metabolized to NO via NOS, L-citrulline is produced. L-citrulline is converted via endogenous arginine synthetic pathways to L-arg; this may be an important source of L-arg during prolonged NO synthesis by iNOS and enable the higher levels of exhaled NO found in asthmatic individuals [78]. In addition, increased levels of exhaled NO in asthma may also result from the nonenzymatic generation of NO from nitrite in the acid airway environment typical of asthma [79, 80]. Two isoforms of arginase enzymes may contribute to total arginase activity levels. Arginase I is highly expressed in the liver, where it serves a key role in the urea cycle, but is also present in cells that lack a complete urea cycle, which suggests other metabolic functions in nonhepatic tissues [81]. Arginase II is present in most tissues, including lung, and localized to the mitochondria within the cell [68, 71, 82, 83].
Greater lung expression of both the Arginase I and II gene transcripts is present in the murine model of allergic asthma as compared with control mice [74]. The guinea pig model of allergic asthma also confirms higher arginase activity during the early response to allergen challenge [70]. In the murine model of asthma, greater L-arg metabolism through arginases linked to airway hyperreactivity [73], suggesting that L-arg levels and its utilization by specific pathways may contribute to asthma pathophysiology. Kenyon et al have shown that treatment of mice with an arginase inhibitor increased the amount of NO produced in murine model of asthma [84]. Inhibition of arginase activity also increases protein S-nitrosylation and tyrosine nitration, and phenotypically enhances peribronchiolar and perivascular inflammation and mucus metaplasia in murine allergic asthma models [85].
Sapienza et al showed that inhalation of L-arg amplifies the inflammatory response in the asthmatic airways [86]. Genetic studies in humans reveal that arginase I and II single-nucleotide polymorphisms are associated with an increased relative risk for asthma and atopy; the effect of polymorphisms on arginase expression or functional activity in humans is still unknown [87]. Nevertheless, it is clear that L-arg/NO metabolism is a determinant of asthmatic airway inflammation.
NO in Oxidation Chemistry: Nitrative Stress
The functional role of NO in biological systems depends on its concentration and association with other biomolecules and proteins [88, 89]. Furthermore it is clear that byproducts of NO reactions can also have biological roles. NO reacts with oxygen or superoxide to form nitrite (NO2−), nitrate (NO3−) and reactive nitrogen species (RNS), such as peroxynitrite (Fig 2). In comparison to healthy controls, asthmatics have higher levels of NO, NO3−, and nitrotyrosine in the airways. Within minutes of an allergen-induced asthmatic response, NO3− increases markedly in asthmatics, while NO2− does not change, and NO decreases. Kinetic modeling suggests that NO reacts rapidly with O2•− produced by oxidative burst of leucocytes to produce ONOO−, which may decay to NO3− or cause the nitration of phenolic compounds [14, 90].
The half-life of peroxynitrite of 1 second at 37°C at pH 7.4 [57]. Peroxynitrite is in equilibrium with peroxynitrous acid (ONOOH), which can covalently modify protein tyrosyl residues through nitration, leading to structural change and potential alterations in biological function [91–94]. In acid environments, ONOO− is protonated to yield peroxynitrous acid (ONOOH) can also react with thiol residues to form S-nitrosothiols (SNO), which function in unique signal transduction pathways in the cell [95–98]. Consistent with the ability of ONOOH to participate in nitration and nitrosylation reactions, GSNO levels increase in parallel to nitrotyrosine in asthmatic airways after allergen challenge in direct proportion to formation of reactive nitrogen species [14].
NO2− is a substrate for hemeperoxidases such as MPO and EPO. Peroxidase-catalyzed oxidation of NO2− results in the formation of nitrogen dioxide radical (NO2•) or related molecules [3, 15, 21, 99–101]. Final products of hemeperoxidases include 3-nitrotyrosine and dityrosine (Fig 3). In vitro studies have demonstrated that tyrosine nitration can cause either a gain or loss of protein function [102–106]. On average, proteins are composed of 4% tyrosine residues; but, chemical nitration of isolated proteins modifies only a subset of tyrosine residues, and the basis for this selectivity is not fully understood. This suggests that an innate property of the target protein or its location may predispose it toward nitration [107]. Nitration is likely not reversible and represents a pathophysiological modification, yet some reports have suggested that nitrated proteins can be repaired by a denitrase enzymic activity that is substrate dependent [108, 109]. However, denitrase enzyme needs to be purified and further characterized in relevance to its repair activity. Biological nitration of protein tyrosine is associated with over 50 diseases including transplant rejection, lung infection, central nervous system and ocular inflammation, shock, cancer, neurological disorders (e.g., amylotrophic lateral sclerosis, Alzheimer’s disease, Parkinson’s disease, and stroke), and asthma [35, 48, 110–114].
Fig. 3. Tyrosine containing amino acid oxidation and cross-linked products.
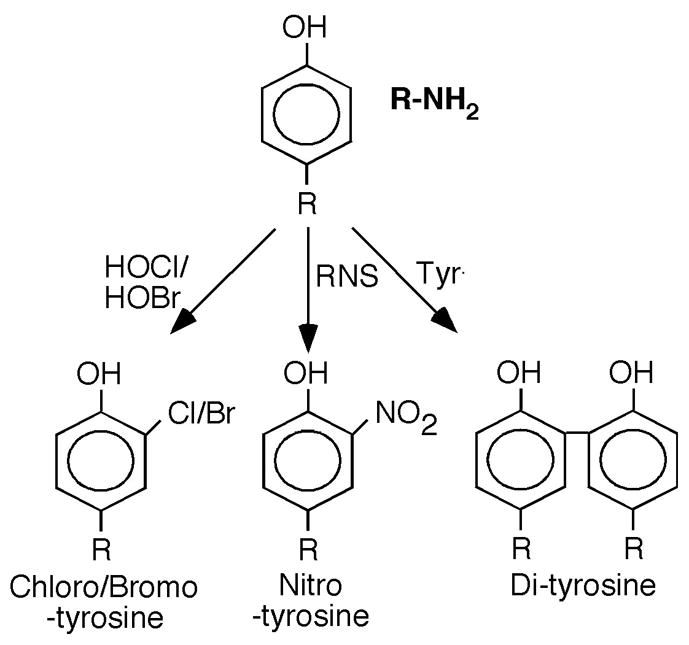
Protein oxidative damage mediated by peroxidase mediated reactive brominating species (HOBr), MPO-generated reactive chlorinating species (HOCl), reactive nitrating species (RNS), tyrosyl radical (Tyr·), may be identified by stable products formed by each pathway.
Previous studies have shown that ONOO− induces airway hyper-responsiveness in guinea pigs in vitro and in vivo [115]. Immunohistochemical studies, using antibodies to nitrotyrosine, have been used to identify modified proteins and suggest a possible involvement of NO-derived oxidants in asthma [48, 114]. Enhanced nitrotyrosine staining is present in airways of asthmatic lung [18, 48, 114]. Subsequent immunolocalization studies revealed that both airway epithelium and eosinophils are major cellular sources for nitrotyrosine formation in asthmatic airways [14]. Protein modification through nitration in asthma has been conclusively shown by mass spectrometry-based quantification, which demonstrates 10-fold increases in nitrotyrosine content in proteins recovered from airways of subjects with asthma compared to non-asthmatic controls [18]. Using NOS2-deficient mice in a model of allergen-induced asthmatic response, nitrotyrosine was absent in airway epithelial cells, but eosinophil recruitment and hyper-responsiveness were still present, suggesting that asthmatic response can occur independent of reactive nitrogen species production [116].
In vitro studies have shown that reactive oxygen and nitrogen species lead to oxidative and nitrative modification of tyrosine and inactivation of superoxide dismutases (SOD), MnSOD and ECSOD, while another major superoxide dismutase CuZnSOD can be inactivated by ROS and RNS through targeting of critical histidine residues and formation of histidinyl radicals [105, 117, 118]. As SOD scavenges superoxide, inactivation of this particular type of enzyme may contribute to cell injury through increased free radical damage [105].
The ovalbumin sensitized and challenged mouse model of asthma has allowed for careful dissection of NO biochemical events [11]. In this model, mice were immunized with ovulbumin and two weeks later challenged with ovulbumin aerosol daily [11]. The extent and range of protein nitration was determined by two-dimensional gel electrophoresis followed by proteomic analysis. Nitrated proteins are detected within days of allergen challenge of sensitized mice (Fig. 4). Nitrotyrosine containing proteins, identified by antibody recognition, were subjected to tryptic digestion and the resulting peptides used to identify the protein by database searching (Table 2). Similar to previous reports, many of the modified proteins are key enzymes for energy production, such as pyruvate kinase, lactate dehydrogenase, malate dehydrogenase, aldolase A and glyceraldehyde phosphate dehydrogenase. This indicates that cells divert from glycosis to other metabolic pathways for energy within cells during the inflammatory response. Dihydropyriminidase related protein, which is involved in axonal growth and has been identified as a target of protein nitration in Alzheimer’s disease [119], was also identified as a target for nitration during the asthmatic response. Annexin II and III, members of the annexin superfamily of calcium and phospholipid binding proteins [120], and annexin I, an important mediator of glucocorticoid action [15] are all nitrated in the experimental model of asthma.
Fig. 4. Increased nitrotyrosine in ova sensitized and challenged mouse lung.

[A] Western blot analysis of nitrotyrosine in tissue lysate of asthmatic mouse lung at days 0, 2, 4 and 6 (lanes 2, 4, 6 and 8) and corresponding controls at respective days (lanes 1, 3, 5 and 7) revealed more nitration in asthmatic lungs compared to controls. Lower panel is β-actin Western analyses for loading control. [B] Densitometric analysis of the Western blots shows that total nitrotyrosine band intensity compared to β-actin is significantly increased in asthmatic lung compared to control. (*) indicates p<0.05. [C] 2D patterns of anti-nitrotyrosine immunopositive protein in asthmatic mouse lung compared to control at day 6. Lung tissue samples from control and ova/ova mice were subjected to proteomic analysis. Coomassie blue stained polyacrylamide gels of control [I] and ova/ova mouse lung tissue [II] are shown with the corresponding Western blots (lower panel; C and D respectively). Although the control blot [III] shows some degree of nitration, the blot [IV] representing the profile of ova/ova mouse lung shows more intense nitration. The protein spots, corresponding to the immunoreactive proteins observed in western blot [IV] on coomassie stained gel [II] were identified by tandem mass spectrometry. Reproduced with permission from The Journal of Immunology (Copyright 2006. The American Association of Immunologists, Inc.).
Table 2.
Identification of nitrated proteins in ova sensitized and challenged mice.
| No.* | Proteins | pI | Mol Wt. (kDa) | Accession No | Coverage | Number of peptides matched | Function |
|---|---|---|---|---|---|---|---|
| 1 | Annexin III | 5.6 | 36 | CAA04887 | 62% | 54 | Associated with cytoplasmic granules in neutrophils and monocytes and translocates to the plasma membrane in activated cells |
| 2 | HBβ chain | 8.5 | 14 | XP_489729 | 38% | 9 | Subunit of hemoglobin |
| Profilin | 8.5 | 14 | CAB87382 | 79% | 39 | Regulates actin polymerization in response to extracellular signals | |
| 3 | GTP binding protein | 7.0 | 24 | BAA01555 | 23% | 20 | Involved in signal transduction |
| 4 | MnSOD | 8.6 | 24 | CAA59335 | 24% | 31 | Antioxidant enzyme |
| 5 | Glutathione S-transferase μ2 | 8.1 | 25 | P10649 | 92% | 75 | Catalyze the conjugation of glutathione to numerous potentially genotoxic compounds |
| 6 | Glutathione S-transferase μ2 | 7.3 | 25 | P15626 | 91% | 81 | |
| 7 | Carbonyl reductase | 9.1 | 26 | NP_031647 | 95% | 94 | Catalyzes the NADPH-dependent reduction of ketones on steroids and prostaglandins |
| 8 | GTP binding protein β chain | 7.6 | 35 | NP_034440 | 50% | 41 | Involved in signal transduction |
| 9 | Annexin II | 7.6 | 38 | AAN86740 | 80% | 90 | Ca2+-dependent phopholipid binding protein, mediates corticosteroid activity and play role in secretion |
| 9 | Lactate dehydrogenase | 7.6 | 36 | CAA26360 | 24% | 19 | Metabolic enzyme, converts lactate to pyruvate |
| 10 | Glyceraldehyde 3-phosphate dehydrogenase | 8.4 | 36 | AAH83149 | 69% | 67 | Enzyme in the glycolysis and gluconeogenesis pathways |
| Malate Dehydrogenase | 8.9 | 36 | CAA30274 | 48% | 20 | Metabolic enzyme of TCA cycle, converts malate to oxaloacetate | |
| 11 | Aldolase A | 8.3 | 39 | AAA37210 | 65% | 70 | Metabolic enzyme of glycolysis pathway |
| 12 | Guanine nucleotide binding protein | 7.6 | 35 | NP_034444 | 29% | 39 | Mediates the activation of cytosolic messengers in signal transduction pathway |
| 13 | Transketolase | 7.2 | 68 | AAH55336 | 44% | 73 | Reversible link between glycolysis and the pentose phosphate pathway |
| 14 | Methylmalonate semialdehyde dehydrogenase | 8.5 | 57 | AAG44988 | 42% | 29 | Responsible for the oxidative decarboxylation of malonate- and methylmalonate semialdehydes to acetyl- and propionyl-CoA. |
| 15 | Pyruvate Kinase | 7.1 | 58 | NP_598428 | 69% | 82 | Metabolic enzyme, converts phosphoenolpyruvate to pyruvate |
| 16 | Catalase | 7.7 | 59 | AAA66054 | 58% | 54 | Catalyses the conversion of toxic hydrogen peroxide into water and oxygen |
| 17 | Aldehyde Dehydrogenase | 7.9 | 54 | NP_036051 | 52% | 67 | Acts in detoxifying a wide variety of organic compounds, toxins and pollutants. |
| 18 | Thioether S-methyl transferase | 6.0 | 30 | S52102 | 52% | 46 | Metabolic enzyme |
| 19 | Antioxidant protein 2 | 5.7 | 24 | AC53277 | 83% | 48 | A member of a family of thiol-specific antioxidants |
| 20 | Carbonic Anhydrase II | 6.5 | 29 | AAA37356 | 23% | 26 | Facilitates the transport of carbon dioxide |
| 21 | Selenium binding protein | 5.8 | 53 | Q91X87 | 56% | 52 | Participates in intra-golgi protein transport |
| 22 | Mouse secretory protein YM-1 | 5.6 | 44 | AAB62394 | 45% | 43 | Secretory protein |
| 23 | Serum Albumin | 5.7 | 70 | NP_033784 | 35% | 36 | Transporter of many small molecules in the blood |
| 24 | Annexin I | 7.0 | 39 | NP_034860 | 77% | 76 | Carries anti-inflammatory role and regulate Ca2+-phospholipid activity |
| 25 | Dihydropyriminidase related protein | 6.0 | 62 | 1351260 | 43% | 36 | Involved in axonal growth |
| 26 | ER60 protease | 5.8 | 57 | JC2385 | 49% | 70 | Cysteine protease of the endoplasmic reticulum |
| 27 | Protein disulfide isomerase | 6.0 | 56 | AAA39906 | 32% | 57 | Resident foldase of the endoplasmic recticulum, catalyses the formation and isomerisation of disulphide bonds during protein folding |
| 28 | Aldehyde Reductase | 6.7 | 36 | AAO72144 | 37% | 30 | Catalyzes reversibly the oxidation of an aldose to an alditol. |
Nitrated proteins found on 2D gel of ova sensitized and challenged mice lung were identified by peptide mass mapping using product-ion (MS/MS) spectra. Proteins are shown with pI, molecular weight, accession number, percentage coverage, number of peptides matched and a brief description of the function of the protein. Reproduced with permission from The Journal of Immunology (Copyright 2006. The American Association of Immunologists, Inc.).
Notably, catalase, MnSOD, glutathione S-transferase, antioxidant protein 2 and carbonic anhydrase II are nitrated in the murine model of asthma, although these enzymes are present in different compartments of the cell. Studies identify loss of activity of catalase and SOD in asthma [121]. MnSOD is nitrated in the asthmatic airways, but the dominant oxidant modifications are related to hydroxyl radical like oxidants presumably via Fenton or Haber-Weiss chemical events [122] Nitration of MnSOD at tyr (Tyr34), which is in the active site of the enzyme, leads to loss of enzyme function, and tissue injury [123, 124].
Other studies have shown that catalase and glutathione-S-tranferase activity are lost by exposure to reactive nitrogen species in vitro [125, 126]. Catalase activity is 50% lower in bronchoalveolar lavage of asthmatic lungs, as compared to healthy controls [11]. Catalase isolated from asthmatic airway epithelial cells has increased protein oxidation markers, including nitrotyrosine and chlorination and oxidation of sulfhydryls, linking oxidative modification to the reduced activity in vivo. Tyrosine oxidant modifications of catalase occur in asthma: chlorination of tyrosine by peroxidase-catalyzed halogenation, and oxidative cross-linking of tyrosine as monitored by dityrosine, a product of tyrosyl radical [11]. Interestingly, catalase contains a putative chlorination site (KXHY) at Tyrosine 236, which may influence the susceptibility of the enzyme to peroxidase activity [127]. Tyrosine modification itself is not likely the complete cause of the loss of catalase activity. Other oxidative modifications, specifically oxidation of the cysteine 377 to cysteic acid, contribute to activity loss of the enzyme [11]. Altogether, the studies provide a picture of an abundance of reactive oxygen and nitrogen biochemical reactions in the asthmatic airway, and the potential downstream pathologic consequences.
NO in the S-Nitrosation Reaction in Asthma
The covalent addition of NO to certain protein moieties, such as thiol residues of cysteines, is termed nitrosation and can promote altered expression or function of enzymes, structural and signaling proteins [128, 129]. Not all cysteine residues are equally susceptible to S-nitrosylation. In fact, the majority of proteins are modified at a single critical cysteine residue [130]. The sources of nitrosylating groups might include nitrogen oxide species that originate exogenously or endogenously. For example, NOS activity can lead to SNO formation; each NOS isoform is capable of producing SNOs in multiple tissues, cells and subcellular compartments. SNO formation appears to depend on the microenvironment of the protein itself [131–135]. On the other hand, NO also may be exchanged between transition metal ion and thiols in proteins [136, 137]. In addition, binding of NOS to adaptor proteins may localize nitrosylation reactions, but there are many more nitrosylated substrates than NOS binding partners. Recent work by Kornberg et al suggests that the S-nitrosylated GAPDH functions as a nuclear nitrosylase via a redox signal transduction cascade mechanism [138, 139].
Nitrosothiols (RSNO) are important reservoirs of bioactive NO, able to take up and store NO for later release as needed. There are several mechanisms for the decomposition of RSNO to produce NO, including nonenzymatic catalysis with copper or reaction with ascorbate [140–143], as well as through enzymatic pathways that act on R-SNO, such as thioredoxin reductase [144], xanthine oxidase [145], Cu/Zn superoxide dismutase [146]. GSNO reductase [GSNOR; alcohol dehydrogenase 5 (EC 1.1.1.1)] catalyses the denitrosylation of S-nitrosoglutathione (GSNO), but not other protein SNO [128, 147]. The decomposition of GSNO leads to formation of NH3 and oxidized glutathione (GSSG).
RSNO are found in the human airway lining fluid, with the most common derivative GSNO at levels of 0.2 to 0.5 μM [148, 149]. GSNO has clearly defined physiologic roles in the airways, including regulation of ciliary beat frequency, immune response and bronchodilation. The bronchodilatory effects of GSNO are largely cGMP independent [148, 150] and result from S-nitrosylation of ion channels, receptor systems, and other myocyte proteins [150–153]. SNOs also have pulmonary vascular smooth muscle relaxant effects [152, 154, 155], and thus serve as a mechanism to couple perfusion via vasodilation to ventilation via bronchodilation [155, 156]. GSNO also contributes to airway homeostasis by antimicrobial and anti-inflammatory properties [157].
Many studies now definitively link GSNO deficiency to asthma pathophysiology [149, 158, 159]. RSNO are present at lower than normal levels in the tracheal aspirates of children with acute asthma exacerbations [149, 160] as well as in the bronchoalveolar lavage fluid of well-controlled asthmatic individuals [14]. Accelerated catabolism of GSNO in asthma results in the overall lower levels of GSNO and contributes to the consequent higher release of free NO as measured in exhaled breath of asthmatics [149]. Most accumulating evidence points to greater enzymatic catabolism of GSNO via GSNOR in asthma [161]. GSNOR-deficient mice, which are unable to break down GSNO and thus have high airway levels, are protected from airway hyper-reactivity in the murine ovulbumin asthma model [158]. Recently, Que et al have also shown that the low level of GSNO in asthmatic bronchoalveolar lavage fluid is inversely correlated with GSNOR expression in lung lavage cell lysates [159]. Furthermore, the GSNOR activity in bronchoalveolar lavage samples was higher in asthma as compared to healthy control subjects and was inversely related to the degree of airway hyperresponsiveness [159, 161]. Altogether, studies suggest that GSNO/NO metabolism is a determinant of airway reactivity in asthma.
In addition to effects on bronchial smooth muscle and airway reactivity, nitrosylation also impacts pro-inflammatory signal transduction in asthma. NO attenuates NF-κB p50–p65 heterodimer DNA binding and inhibits pro-inflammatory gene transcription through S-nitrosylation of the p50 subunit [162]. The p65 binding partner of p50 is also found to be S-nitrosylated in cytokine stimulated cells [163]. S-nitrosylation of p65 inhibits NF-κB-dependent gene transcription, and nuclear levels of S-nitrosylated p65 correlate with decreased DNA binding of the p50–p65 heterodimer [163]. Very recently Olson et al have tested the therapeutic benefit of S-nitrosothiols in the murine model of ovalbumin induced allergic inflammation [164]. They found that instillation of GSNO suppressed NF-B activation, but did not significantly affect overall markers of inflammation or mucus metaplasia, which may suggest its bronchodilatory effects outweigh its anti-inflammatory properties.
Summary
There is definitive evidence for alterations in NO synthetic and catabolic pathways in asthma. There are greater levels of toxic RNS, and equally important, lower levels of beneficial RSNO in asthmatic airways. In contrast to stable nitration products, the labile nature of RSNO has presented challenges to comprehensive identification of in vivo targets in asthma. However, mass spectrometry and derivatization techniques are providing exciting insight into the wide array of events controlled by nitrosylation chemistry in the cell. In parallel, physiologic studies have identified that Arginine/NO metabolic processes regulate airway hyper-reactivity and inflammation, key components that define asthma. Detailed understanding of the biochemical pathways that control NO metabolism and its physiologic functions will enable future rational design of strategies to optimize levels of beneficial NO products, while limiting the formation of more damaging species, in the treatment of asthma.
Research Highlights.
Asthma is a disease characterized by unique alterations in nitric oxide (NO) metabolism.
Greater overall synthesis by the NO-synthase type 2 enzyme is accompanied by a shift towards more nitration of proteins in the airway, and less nitrosylation of critical thiol residues in proteins.
The greater nitration of specific proteins has been associated with adverse effects on protein functions, while loss of S-nitrosothiol (SNO) products leads to loss of bronchodilator effects.
Thus, the abnormal NO metabolism contributes to the defining features of asthma, including chronic airway inflammation, injury and bronchoconstriction.
Acknowledgments
The authors acknowledge grant support from the NIH [RO1 HL69170, PO1 HL081064, RC1 HL099303, UL1 RR024989 and KL2 RR024990]. SCE is a Senior Fellow of the American Asthma Foundation Sandler Program for Asthma Research.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Elias JA, Zhu Z, Chupp G, Homer RJ. Airway remodeling in asthma. J Clin Invest. 1999;104:1001–1006. doi: 10.1172/JCI8124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Frieri M. Asthma concepts in the new millennium: update in asthma pathophysiology. Allergy Asthma Proc. 2005;26:83–88. [PubMed] [Google Scholar]
- 3.Iijima H, Duguet A, Eum SY, Hamid Q, Eidelman DH. Nitric oxide and protein nitration are eosinophil dependent in allergen-challenged mice. American journal of respiratory and critical care medicine. 2001;163:1233–1240. doi: 10.1164/ajrccm.163.5.2003145. [DOI] [PubMed] [Google Scholar]
- 4.Ricciardolo FL, Geppetti P, Mistretta A, Nadel JA, Sapienza MA, Bellofiore S, Di Maria GU. Randomised double-blind placebo-controlled study of the effect of inhibition of nitric oxide synthesis in bradykinin-induced asthma. Lancet. 1996;348:374–377. doi: 10.1016/s0140-6736(96)04450-9. [DOI] [PubMed] [Google Scholar]
- 5.Munzel T, Feil R, Mulsch A, Lohmann SM, Hofmann F, Walter U. Physiology and pathophysiology of vascular signaling controlled by guanosine 3′,5′-cyclic monophosphate-dependent protein kinase [corrected] Circulation. 2003;108:2172–2183. doi: 10.1161/01.CIR.0000094403.78467.C3. [DOI] [PubMed] [Google Scholar]
- 6.Papapetropoulos A, Simoes DC, Xanthou G, Roussos C, Gratziou C. Soluble guanylyl cyclase expression is reduced in allergic asthma. American journal of physiology. 2006;290:L179–184. doi: 10.1152/ajplung.00330.2005. [DOI] [PubMed] [Google Scholar]
- 7.Bousquet J, Jeffery PK, Busse WW, Johnson M, Vignola AM. Asthma. From bronchoconstriction to airways inflammation and remodeling. American journal of respiratory and critical care medicine. 2000;161:1720–1745. doi: 10.1164/ajrccm.161.5.9903102. [DOI] [PubMed] [Google Scholar]
- 8.Takata M, Filippov G, Liu H, Ichinose F, Janssens S, Bloch DB, Bloch KD. Cytokines decrease sGC in pulmonary artery smooth muscle cells via NO-dependent and NO-independent mechanisms. American journal of physiology. 2001;280:L272–278. doi: 10.1152/ajplung.2001.280.2.L272. [DOI] [PubMed] [Google Scholar]
- 9.Fernhoff NB, Derbyshire ER, Marletta MA. A nitric oxide/cysteine interaction mediates the activation of soluble guanylate cyclase. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:21602–21607. doi: 10.1073/pnas.0911083106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gerard C. Biomedicine. Asthmatics breathe easier when it’s SNO-ing. Science (New York, NY. 2005;308:1560–1561. doi: 10.1126/science.1114163. [DOI] [PubMed] [Google Scholar]
- 11.Ghosh S, Janocha AJ, Aronica MA, Swaidani S, Comhair SA, Xu W, Zheng L, Kaveti S, Kinter M, Hazen SL, Erzurum SC. Nitrotyrosine proteome survey in asthma identifies oxidative mechanism of catalase inactivation. J Immunol. 2006;176:5587–5597. doi: 10.4049/jimmunol.176.9.5587. [DOI] [PubMed] [Google Scholar]
- 12.Lane C, Knight D, Burgess S, Franklin P, Horak F, Legg J, Moeller A, Stick S. Epithelial inducible nitric oxide synthase activity is the major determinant of nitric oxide concentration in exhaled breath. Thorax. 2004;59:757–760. doi: 10.1136/thx.2003.014894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Andreadis AA, Hazen SL, Comhair SA, Erzurum SC. Oxidative and nitrosative events in asthma. Free Radic Biol Med. 2003;35:213–225. doi: 10.1016/s0891-5849(03)00278-8. [DOI] [PubMed] [Google Scholar]
- 14.Dweik RA, Comhair SA, Gaston B, Thunnissen FB, Farver C, Thomassen MJ, Kavuru M, Hammel J, Abu-Soud HM, Erzurum SC. NO chemical events in the human airway during the immediate and late antigen-induced asthmatic response. Proceedings of the National Academy of Sciences of the United States of America. 2001;98:2622–2627. doi: 10.1073/pnas.051629498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Brennan ML, Wu W, Fu X, Shen Z, Song W, Frost H, Vadseth C, Narine L, Lenkiewicz E, Borchers MT, Lusis AJ, Lee JJ, Lee NA, Abu-Soud HM, Ischiropoulos H, Hazen SL. A tale of two controversies: defining both the role of peroxidases in nitrotyrosine formation in vivo using eosinophil peroxidase and myeloperoxidase-deficient mice, and the nature of peroxidase-generated reactive nitrogen species. The Journal of biological chemistry. 2002;277:17415–17427. doi: 10.1074/jbc.M112400200. [DOI] [PubMed] [Google Scholar]
- 16.Eiserich JP, Hristova M, Cross CE, Jones AD, Freeman BA, Halliwell B, van der Vliet A. Formation of nitric oxide-derived inflammatory oxidants by myeloperoxidase in neutrophils. Nature. 1998;391:393–397. doi: 10.1038/34923. [DOI] [PubMed] [Google Scholar]
- 17.Hazen SL, Heinecke JW. 3-Chlorotyrosine, a specific marker of myeloperoxidase-catalyzed oxidation, is markedly elevated in low density lipoprotein isolated from human atherosclerotic intima. J Clin Invest. 1997;99:2075–2081. doi: 10.1172/JCI119379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.MacPherson JC, Comhair SA, Erzurum SC, Klein DF, Lipscomb MF, Kavuru MS, Samoszuk MK, Hazen SL. Eosinophils are a major source of nitric oxide-derived oxidants in severe asthma: characterization of pathways available to eosinophils for generating reactive nitrogen species. J Immunol. 2001;166:5763–5772. doi: 10.4049/jimmunol.166.9.5763. [DOI] [PubMed] [Google Scholar]
- 19.van der Vliet A, Eiserich JP, Halliwell B, Cross CE. Formation of reactive nitrogen species during peroxidase-catalyzed oxidation of nitrite. A potential additional mechanism of nitric oxide-dependent toxicity. The Journal of biological chemistry. 1997;272:7617–7625. doi: 10.1074/jbc.272.12.7617. [DOI] [PubMed] [Google Scholar]
- 20.Wu W, Chen Y, Hazen SL. Eosinophil peroxidase nitrates protein tyrosyl residues. Implications for oxidative damage by nitrating intermediates in eosinophilic inflammatory disorders. The Journal of biological chemistry. 1999;274:25933–25944. doi: 10.1074/jbc.274.36.25933. [DOI] [PubMed] [Google Scholar]
- 21.Duguet A, Iijima H, Eum SY, Hamid Q, Eidelman DH. Eosinophil peroxidase mediates protein nitration in allergic airway inflammation in mice. American journal of respiratory and critical care medicine. 2001;164:1119–1126. doi: 10.1164/ajrccm.164.7.2010085. [DOI] [PubMed] [Google Scholar]
- 22.Holguin F, Fitzpatrick A. Obesity, asthma, and oxidative stress. J Appl Physiol. 108:754–759. doi: 10.1152/japplphysiol.00702.2009. [DOI] [PubMed] [Google Scholar]
- 23.Stephens JW, Khanolkar MP, Bain SC. The biological relevance and measurement of plasma markers of oxidative stress in diabetes and cardiovascular disease. Atherosclerosis. 2009;202:321–329. doi: 10.1016/j.atherosclerosis.2008.06.006. [DOI] [PubMed] [Google Scholar]
- 24.Vincent HK, Innes KE, Vincent KR. Oxidative stress and potential interventions to reduce oxidative stress in overweight and obesity. Diabetes Obes Metab. 2007;9:813–839. doi: 10.1111/j.1463-1326.2007.00692.x. [DOI] [PubMed] [Google Scholar]
- 25.Gardner PR, Fridovich I. Superoxide sensitivity of the Escherichia coli aconitase. The Journal of biological chemistry. 1991;266:19328–19333. [PubMed] [Google Scholar]
- 26.Zheng L, Nukuna B, Brennan ML, Sun M, Goormastic M, Settle M, Schmitt D, Fu X, Thomson L, Fox PL, Ischiropoulos H, Smith JD, Kinter M, Hazen SL. Apolipoprotein A-I is a selective target for myeloperoxidase-catalyzed oxidation and functional impairment in subjects with cardiovascular disease. J Clin Invest. 2004;114:529–541. doi: 10.1172/JCI21109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tanuma N, Miyata R, Hayashi M, Uchiyama A, Kurata K. Oxidative stress as a biomarker of respiratory disturbance in patients with severe motor and intellectual disabilities. Brain Dev. 2008;30:402–409. doi: 10.1016/j.braindev.2007.12.001. [DOI] [PubMed] [Google Scholar]
- 28.Ercan H, Birben E, Dizdar EA, Keskin O, Karaaslan C, Soyer OU, Dut R, Sackesen C, Besler T, Kalayci O. Oxidative stress and genetic and epidemiologic determinants of oxidant injury in childhood asthma. J Allergy Clin Immunol. 2006;118:1097–1104. doi: 10.1016/j.jaci.2006.08.012. [DOI] [PubMed] [Google Scholar]
- 29.Nadeem A, Raj HG, Chhabra SK. Increased oxidative stress in acute exacerbations of asthma. J Asthma. 2005;42:45–50. doi: 10.1081/jas-200044774. [DOI] [PubMed] [Google Scholar]
- 30.Wedes SH, Khatri SB, Zhang R, Wu W, Comhair SA, Wenzel S, Teague WG, Israel E, Erzurum SC, Hazen SL. Noninvasive markers of airway inflammation in asthma. Clin Transl Sci. 2009;2:112–117. doi: 10.1111/j.1752-8062.2009.00095.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wu P, Roberts LJ, 2nd, Shintani AK, Sheller JR, Minton PA, Higgins SB, Hartert TV. Changes in urinary dinor dihydro F(2)-isoprostane metabolite concentrations, a marker of oxidative stress, during and following asthma exacerbations. Free Radic Res. 2007;41:956–962. doi: 10.1080/10715760701444600. [DOI] [PubMed] [Google Scholar]
- 32.Ochs-Balcom HM, Grant BJ, Muti P, Sempos CT, Freudenheim JL, Browne RW, McCann SE, Trevisan M, Cassano PA, Iacoviello L, Schunemann HJ. Antioxidants, oxidative stress, and pulmonary function in individuals diagnosed with asthma or COPD. Eur J Clin Nutr. 2006;60:991–999. doi: 10.1038/sj.ejcn.1602410. [DOI] [PubMed] [Google Scholar]
- 33.Comhair SA, Bhathena PR, Dweik RA, Kavuru M, Erzurum SC. Rapid loss of superoxide dismutase activity during antigen-induced asthmatic response. Lancet. 2000;355:624. doi: 10.1016/S0140-6736(99)04736-4. [DOI] [PubMed] [Google Scholar]
- 34.Dweik RA, Laskowski D, Abu-Soud HM, Kaneko F, Hutte R, Stuehr DJ, Erzurum SC. Nitric oxide synthesis in the lung. Regulation by oxygen through a kinetic mechanism. J Clin Invest. 1998;101:660–666. doi: 10.1172/JCI1378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Guo FH, Comhair SA, Zheng S, Dweik RA, Eissa NT, Thomassen MJ, Calhoun W, Erzurum SC. Molecular mechanisms of increased nitric oxide (NO) in asthma: evidence for transcriptional and post-translational regulation of NO synthesis. J Immunol. 2000;164:5970–5980. doi: 10.4049/jimmunol.164.11.5970. [DOI] [PubMed] [Google Scholar]
- 36.Persson MG, Zetterstrom O, Agrenius V, Ihre E, Gustafsson LE. Single-breath nitric oxide measurements in asthmatic patients and smokers. Lancet. 1994;343:146–147. doi: 10.1016/s0140-6736(94)90935-0. [DOI] [PubMed] [Google Scholar]
- 37.Kharitonov SA, O’Connor BJ, Evans DJ, Barnes PJ. Allergen-induced late asthmatic reactions are associated with elevation of exhaled nitric oxide. American journal of respiratory and critical care medicine. 1995;151:1894–1899. doi: 10.1164/ajrccm.151.6.7767537. [DOI] [PubMed] [Google Scholar]
- 38.Khatri SB, Ozkan M, McCarthy K, Laskowski D, Hammel J, Dweik RA, Erzurum SC. Alterations in exhaled gas profile during allergen-induced asthmatic response. American journal of respiratory and critical care medicine. 2001;164:1844–1848. doi: 10.1164/ajrccm.164.10.2106119. [DOI] [PubMed] [Google Scholar]
- 39.Massaro AF, Mehta S, Lilly CM, Kobzik L, Reilly JJ, Drazen JM. Elevated nitric oxide concentrations in isolated lower airway gas of asthmatic subjects. American journal of respiratory and critical care medicine. 1996;153:1510–1514. doi: 10.1164/ajrccm.153.5.8630594. [DOI] [PubMed] [Google Scholar]
- 40.Mehta S, Lilly CM, Rollenhagen JE, Haley KJ, Asano K, Drazen JM. Acute and chronic effects of allergic airway inflammation on pulmonary nitric oxide production. Am J Physiol. 1997;272:L124–131. doi: 10.1152/ajplung.1997.272.1.L124. [DOI] [PubMed] [Google Scholar]
- 41.Silkoff PE, Sylvester JT, Zamel N, Permutt S. Airway nitric oxide diffusion in asthma: Role in pulmonary function and bronchial responsiveness. American journal of respiratory and critical care medicine. 2000;161:1218–1228. doi: 10.1164/ajrccm.161.4.9903111. [DOI] [PubMed] [Google Scholar]
- 42.Barnes PJ. Nitric oxide and asthma. Res Immunol. 1995;146:698–702. doi: 10.1016/0923-2494(96)84921-2. [DOI] [PubMed] [Google Scholar]
- 43.Nathan C, Xie QW. Nitric oxide synthases: roles, tolls, and controls. Cell. 1994;78:915–918. doi: 10.1016/0092-8674(94)90266-6. [DOI] [PubMed] [Google Scholar]
- 44.Stuehr DJ. Mammalian nitric oxide synthases. Biochim Biophys Acta. 1999;1411:217–230. doi: 10.1016/s0005-2728(99)00016-x. [DOI] [PubMed] [Google Scholar]
- 45.Uetani K, Der SD, Zamanian-Daryoush M, de La Motte C, Lieberman BY, Williams BR, Erzurum SC. Central role of double-stranded RNA-activated protein kinase in microbial induction of nitric oxide synthase. J Immunol. 2000;165:988–996. doi: 10.4049/jimmunol.165.2.988. [DOI] [PubMed] [Google Scholar]
- 46.Zheng S, De BP, Choudhary S, Comhair SA, Goggans T, Slee R, Williams BR, Pilewski J, Haque SJ, Erzurum SC. Impaired innate host defense causes susceptibility to respiratory virus infections in cystic fibrosis. Immunity. 2003;18:619–630. doi: 10.1016/s1074-7613(03)00114-6. [DOI] [PubMed] [Google Scholar]
- 47.Moncada S, Higgs A. The L-arginine-nitric oxide pathway. N Engl J Med. 1993;329:2002–2012. doi: 10.1056/NEJM199312303292706. [DOI] [PubMed] [Google Scholar]
- 48.Saleh D, Ernst P, Lim S, Barnes PJ, Giaid A. Increased formation of the potent oxidant peroxynitrite in the airways of asthmatic patients is associated with induction of nitric oxide synthase: effect of inhaled glucocorticoid. Faseb J. 1998;12:929–937. [PubMed] [Google Scholar]
- 49.Kobzik L, Bredt DS, Lowenstein CJ, Drazen J, Gaston B, Sugarbaker D, Stamler JS. Nitric oxide synthase in human and rat lung: immunocytochemical and histochemical localization. Am J Respir Cell Mol Biol. 1993;9:371–377. doi: 10.1165/ajrcmb/9.4.371. [DOI] [PubMed] [Google Scholar]
- 50.Sanders SP. Nitric oxide in asthma. Pathogenic, therapeutic, or diagnostic? Am J Respir Cell Mol Biol. 1999;21:147–149. doi: 10.1165/ajrcmb.21.2.f158. [DOI] [PubMed] [Google Scholar]
- 51.De Sanctis GT, MacLean JA, Hamada K, Mehta S, Scott JA, Jiao A, Yandava CN, Kobzik L, Wolyniec WW, Fabian AJ, Venugopal CS, Grasemann H, Huang PL, Drazen JM. Contribution of nitric oxide synthases 1, 2, and 3 to airway hyperresponsiveness and inflammation in a murine model of asthma. J Exp Med. 1999;189:1621–1630. doi: 10.1084/jem.189.10.1621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Grasemann H, Yandava CN, Storm van’s Gravesande K, Deykin A, Pillari A, Ma J, Sonna LA, Lilly C, Stampfer MJ, Israel E, Silverman EK, Drazen JM. A neuronal NO synthase (NOS1) gene polymorphism is associated with asthma. Biochem Biophys Res Commun. 2000;272:391–394. doi: 10.1006/bbrc.2000.2794. [DOI] [PubMed] [Google Scholar]
- 53.Guo FH, De Raeve HR, Rice TW, Stuehr DJ, Thunnissen FB, Erzurum SC. Continuous nitric oxide synthesis by inducible nitric oxide synthase in normal human airway epithelium in vivo. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:7809–7813. doi: 10.1073/pnas.92.17.7809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Guo FH, Uetani K, Haque SJ, Williams BR, Dweik RA, Thunnissen FB, Calhoun W, Erzurum SC. Interferon gamma and interleukin 4 stimulate prolonged expression of inducible nitric oxide synthase in human airway epithelium through synthesis of soluble mediators. J Clin Invest. 1997;100:829–838. doi: 10.1172/JCI119598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Donnelly LE, Barnes PJ. Expression and regulation of inducible nitric oxide synthase from human primary airway epithelial cells. Am J Respir Cell Mol Biol. 2002;26:144–151. doi: 10.1165/ajrcmb.26.1.4477. [DOI] [PubMed] [Google Scholar]
- 56.Ricciardolo FL, Sterk PJ, Gaston B, Folkerts G. Nitric oxide in health and disease of the respiratory system. Physiol Rev. 2004;84:731–765. doi: 10.1152/physrev.00034.2003. [DOI] [PubMed] [Google Scholar]
- 57.Fischer A, Folkerts G, Geppetti P, Groneberg DA. Mediators of asthma: nitric oxide. Pulm Pharmacol Ther. 2002;15:73–81. doi: 10.1006/pupt.2001.0332. [DOI] [PubMed] [Google Scholar]
- 58.Prado CM, Leick-Maldonado EA, Yano L, Leme AS, Capelozzi VL, Martins MA, Tiberio IF. Effects of nitric oxide synthases in chronic allergic airway inflammation and remodeling. Am J Respir Cell Mol Biol. 2006;35:457–465. doi: 10.1165/rcmb.2005-0391OC. [DOI] [PubMed] [Google Scholar]
- 59.Marks-Konczalik J, Chu SC, Moss J. Cytokine-mediated transcriptional induction of the human inducible nitric oxide synthase gene requires both activator protein 1 and nuclear factor kappaB-binding sites. The Journal of biological chemistry. 1998;273:22201–22208. doi: 10.1074/jbc.273.35.22201. [DOI] [PubMed] [Google Scholar]
- 60.Taylor BS, de Vera ME, Ganster RW, Wang Q, Shapiro RA, Morris SM, Jr, Billiar TR, Geller DA. Multiple NF-kappaB enhancer elements regulate cytokine induction of the human inducible nitric oxide synthase gene. The Journal of biological chemistry. 1998;273:15148–15156. doi: 10.1074/jbc.273.24.15148. [DOI] [PubMed] [Google Scholar]
- 61.Xie QW, Whisnant R, Nathan C. Promoter of the mouse gene encoding calcium-independent nitric oxide synthase confers inducibility by interferon gamma and bacterial lipopolysaccharide. J Exp Med. 1993;177:1779–1784. doi: 10.1084/jem.177.6.1779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Xu W, Comhair SA, Zheng S, Chu SC, Marks-Konczalik J, Moss J, Haque SJ, Erzurum SC. STAT-1 and c-Fos interaction in nitric oxide synthase-2 gene activation. American journal of physiology. 2003;285:L137–148. doi: 10.1152/ajplung.00441.2002. [DOI] [PubMed] [Google Scholar]
- 63.Chu SC, Marks-Konczalik J, Wu HP, Banks TC, Moss J. Analysis of the cytokine-stimulated human inducible nitric oxide synthase (iNOS) gene: characterization of differences between human and mouse iNOS promoters. Biochem Biophys Res Commun. 1998;248:871–878. doi: 10.1006/bbrc.1998.9062. [DOI] [PubMed] [Google Scholar]
- 64.Crystal RG. Research opportunities and advances in lung disease. Jama. 2001;285:612–618. doi: 10.1001/jama.285.5.612. [DOI] [PubMed] [Google Scholar]
- 65.Ceylan E, Aksoy N, Gencer M, Vural H, Keles H, Selek S. Evaluation of oxidative-antioxidative status and the L-arginine-nitric oxide pathway in asthmatic patients. Respir Med. 2005;99:871–876. doi: 10.1016/j.rmed.2004.12.001. [DOI] [PubMed] [Google Scholar]
- 66.Gotoh T, Mori M. Arginase II downregulates nitric oxide (NO) production and prevents NO-mediated apoptosis in murine macrophage-derived RAW 264.7 cells. J Cell Biol. 1999;144:427–434. doi: 10.1083/jcb.144.3.427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Greene AL, Rutherford MS, Regal RR, Flickinger GH, Hendrickson JA, Giulivi C, Mohrman ME, Fraser DG, Regal JF. Arginase activity differs with allergen in the effector phase of ovalbumin- versus trimellitic anhydride-induced asthma. Toxicol Sci. 2005;88:420–433. doi: 10.1093/toxsci/kfi311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.King NE, Rothenberg ME, Zimmermann N. Arginine in asthma and lung inflammation. J Nutr. 2004;134:2830S–2836S. doi: 10.1093/jn/134.10.2830S. discussion 2853S. [DOI] [PubMed] [Google Scholar]
- 69.Lara AR, Erzurum SC. A urinary test for pulmonary arterial hypertension? American journal of respiratory and critical care medicine. 2005;172:262–263. doi: 10.1164/rccm.2505009. [DOI] [PubMed] [Google Scholar]
- 70.Maarsingh H, Leusink J, Bos IS, Zaagsma J, Meurs H. Arginase strongly impairs neuronal nitric oxide-mediated airway smooth muscle relaxation in allergic asthma. Respir Res. 2006;7:6. doi: 10.1186/1465-9921-7-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Morris CR, Poljakovic M, Lavrisha L, Machado L, Kuypers FA, Morris SM., Jr Decreased arginine bioavailability and increased serum arginase activity in asthma. American journal of respiratory and critical care medicine. 2004;170:148–153. doi: 10.1164/rccm.200309-1304OC. [DOI] [PubMed] [Google Scholar]
- 72.Vercelli D. Arginase: marker, effector, or candidate gene for asthma? J Clin Invest. 2003;111:1815–1817. doi: 10.1172/JCI18908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Yang M, Rangasamy D, Matthaei KI, Frew AJ, Zimmmermann N, Mahalingam S, Webb DC, Tremethick DJ, Thompson PJ, Hogan SP, Rothenberg ME, Cowden WB, Foster PS. Inhibition of arginase I activity by RNA interference attenuates IL-13-induced airways hyperresponsiveness. J Immunol. 2006;177:5595–5603. doi: 10.4049/jimmunol.177.8.5595. [DOI] [PubMed] [Google Scholar]
- 74.Zimmermann N, King NE, Laporte J, Yang M, Mishra A, Pope SM, Muntel EE, Witte DP, Pegg AA, Foster PS, Hamid Q, Rothenberg ME. Dissection of experimental asthma with DNA microarray analysis identifies arginase in asthma pathogenesis. J Clin Invest. 2003;111:1863–1874. doi: 10.1172/JCI17912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Zimmermann N, Rothenberg ME. The arginine-arginase balance in asthma and lung inflammation. Eur J Pharmacol. 2006;533:253–262. doi: 10.1016/j.ejphar.2005.12.047. [DOI] [PubMed] [Google Scholar]
- 76.Meurs H, Maarsingh H, Zaagsma J. Arginase and asthma: novel insights into nitric oxide homeostasis and airway hyperresponsiveness. Trends Pharmacol Sci. 2003;24:450–455. doi: 10.1016/S0165-6147(03)00227-X. [DOI] [PubMed] [Google Scholar]
- 77.Meurs H, McKay S, Maarsingh H, Hamer MA, Macic L, Molendijk N, Zaagsma J. Increased arginase activity underlies allergen-induced deficiency of cNOS-derived nitric oxide and airway hyperresponsiveness. Br J Pharmacol. 2002;136:391–398. doi: 10.1038/sj.bjp.0704725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kurt RWM. L-Arginine Metabolic Pathways. The Open Nitric Oxide Journal. 2010;2:9–19. [Google Scholar]
- 79.Hunt JF, Erwin E, Palmer L, Vaughan J, Malhotra N, Platts-Mills TA, Gaston B. Expression and activity of pH-regulatory glutaminase in the human airway epithelium. American journal of respiratory and critical care medicine. 2002;165:101–107. doi: 10.1164/ajrccm.165.1.2104131. [DOI] [PubMed] [Google Scholar]
- 80.Hunt JF, Fang K, Malik R, Snyder A, Malhotra N, Platts-Mills TA, Gaston B. Endogenous airway acidification. Implications for asthma pathophysiology. American journal of respiratory and critical care medicine. 2000;161:694–699. doi: 10.1164/ajrccm.161.3.9911005. [DOI] [PubMed] [Google Scholar]
- 81.Yu H, Yoo PK, Aguirre CC, Tsoa RW, Kern RM, Grody WW, Cederbaum SD, Iyer RK. Widespread expression of arginase I in mouse tissues. Biochemical and physiological implications. J Histochem Cytochem. 2003;51:1151–1160. doi: 10.1177/002215540305100905. [DOI] [PubMed] [Google Scholar]
- 82.Colleluori DM, Morris SM, Jr, Ash DE. Expression, purification, and characterization of human type II arginase. Arch Biochem Biophys. 2001;389:135–143. doi: 10.1006/abbi.2001.2324. [DOI] [PubMed] [Google Scholar]
- 83.Salimuddin, Nagasaki A, Gotoh T, Isobe H, Mori M. Regulation of the genes for arginase isoforms and related enzymes in mouse macrophages by lipopolysaccharide. Am J Physiol. 1999;277:E110–117. doi: 10.1152/ajpendo.1999.277.1.E110. [DOI] [PubMed] [Google Scholar]
- 84.Kenyon NJ, Bratt JM, Linderholm AL, Last MS, Last JA. Arginases I and II in lungs of ovalbumin-sensitized mice exposed to ovalbumin: sources and consequences. Toxicol Appl Pharmacol. 2008;230:269–275. doi: 10.1016/j.taap.2008.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Ckless K, Lampert A, Reiss J, Kasahara D, Poynter ME, Irvin CG, Lundblad LK, Norton R, van der Vliet A, Janssen-Heininger YM. Inhibition of arginase activity enhances inflammation in mice with allergic airway disease, in association with increases in protein S-nitrosylation and tyrosine nitration. J Immunol. 2008;181:4255–4264. doi: 10.4049/jimmunol.181.6.4255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Sapienza MA, Kharitonov SA, Horvath I, Chung KF, Barnes PJ. Effect of inhaled L-arginine on exhaled nitric oxide in normal and asthmatic subjects. Thorax. 1998;53:172–175. doi: 10.1136/thx.53.3.172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Li H, Romieu I, Sienra-Monge JJ, Ramirez-Aguilar M, Estela Del Rio-Navarro B, Kistner EO, Gjessing HK, Lara-Sanchez Idel C, Chiu GY, London SJ. Genetic polymorphisms in arginase I and II and childhood asthma and atopy. J Allergy Clin Immunol. 2006;117:119–126. doi: 10.1016/j.jaci.2005.09.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Lancaster JR., Jr Nitroxidative, nitrosative, and nitrative stress: kinetic predictions of reactive nitrogen species chemistry under biological conditions. Chem Res Toxicol. 2006;19:1160–1174. doi: 10.1021/tx060061w. [DOI] [PubMed] [Google Scholar]
- 89.Ridnour LA, Thomas DD, Mancardi D, Espey MG, Miranda KM, Paolocci N, Feelisch M, Fukuto J, Wink DA. The chemistry of nitrosative stress induced by nitric oxide and reactive nitrogen oxide species. Putting perspective on stressful biological situations. Biol Chem. 2004;385:1–10. doi: 10.1515/BC.2004.001. [DOI] [PubMed] [Google Scholar]
- 90.Comhair SA, Erzurum SC. Redox control of asthma: molecular mechanisms and therapeutic opportunities. Antioxid Redox Signal. 12:93–124. doi: 10.1089/ars.2008.2425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Crow JP, Beckman JS. Reactions between nitric oxide, superoxide, and peroxynitrite: footprints of peroxynitrite in vivo. Adv Pharmacol. 1995;34:17–43. doi: 10.1016/s1054-3589(08)61079-0. [DOI] [PubMed] [Google Scholar]
- 92.Freeman BA, White CR, Gutierrez H, Paler-Martinez A, Tarpey MM, Rubbo H. Oxygen radical-nitric oxide reactions in vascular diseases. Adv Pharmacol. 1995;34:45–69. doi: 10.1016/s1054-3589(08)61080-7. [DOI] [PubMed] [Google Scholar]
- 93.Radi R, Denicola A, Freeman BA. Peroxynitrite reactions with carbon dioxide-bicarbonate. Methods Enzymol. 1999;301:353–367. doi: 10.1016/s0076-6879(99)01099-x. [DOI] [PubMed] [Google Scholar]
- 94.Rochelle LG, Fischer BM, Adler KB. Concurrent production of reactive oxygen and nitrogen species by airway epithelial cells in vitro. Free Radic Biol Med. 1998;24:863–868. doi: 10.1016/s0891-5849(97)00375-4. [DOI] [PubMed] [Google Scholar]
- 95.Gaston BM, Carver J, Doctor A, Palmer LA. S-nitrosylation signaling in cell biology. Mol Interv. 2003;3:253–263. doi: 10.1124/mi.3.5.253. [DOI] [PubMed] [Google Scholar]
- 96.Lima B, Forrester MT, Hess DT, Stamler JS. S-nitrosylation in cardiovascular signaling. Circulation research. 106:633–646. doi: 10.1161/CIRCRESAHA.109.207381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Reynaert NL, Ckless K, Korn SH, Vos N, Guala AS, Wouters EF, van der Vliet A, Janssen-Heininger YM. Nitric oxide represses inhibitory kappaB kinase through S-nitrosylation. Proceedings of the National Academy of Sciences of the United States of America. 2004;101:8945–8950. doi: 10.1073/pnas.0400588101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Whalen EJ, Foster MW, Matsumoto A, Ozawa K, Violin JD, Que LG, Nelson CD, Benhar M, Keys JR, Rockman HA, Koch WJ, Daaka Y, Lefkowitz RJ, Stamler JS. Regulation of beta-adrenergic receptor signaling by S-nitrosylation of G-protein-coupled receptor kinase 2. Cell. 2007;129:511–522. doi: 10.1016/j.cell.2007.02.046. [DOI] [PubMed] [Google Scholar]
- 99.Abu-Soud HM, Hazen SL. Nitric oxide is a physiological substrate for mammalian peroxidases. The Journal of biological chemistry. 2000;275:37524–37532. doi: 10.1074/jbc.275.48.37524. [DOI] [PubMed] [Google Scholar]
- 100.Abu-Soud HM, Hazen SL. Nitric oxide modulates the catalytic activity of myeloperoxidase. The Journal of biological chemistry. 2000;275:5425–5430. doi: 10.1074/jbc.275.8.5425. [DOI] [PubMed] [Google Scholar]
- 101.Abu-Soud HM, Khassawneh MY, Sohn JT, Murray P, Haxhiu MA, Hazen SL. Peroxidases inhibit nitric oxide (NO) dependent bronchodilation: development of a model describing NO-peroxidase interactions. Biochemistry. 2001;40:11866–11875. doi: 10.1021/bi011206v. [DOI] [PubMed] [Google Scholar]
- 102.Aslan M, Ryan TM, Townes TM, Coward L, Kirk MC, Barnes S, Alexander CB, Rosenfeld SS, Freeman BA. Nitric oxide-dependent generation of reactive species in sickle cell disease. Actin tyrosine induces defective cytoskeletal polymerization. The Journal of biological chemistry. 2003;278:4194–4204. doi: 10.1074/jbc.M208916200. [DOI] [PubMed] [Google Scholar]
- 103.Balafanova Z, Bolli R, Zhang J, Zheng Y, Pass JM, Bhatnagar A, Tang XL, Wang O, Cardwell E, Ping P. Nitric oxide (NO) induces nitration of protein kinase Cepsilon (PKCepsilon ), facilitating PKCepsilon translocation via enhanced PKCepsilon -RACK2 interactions: a novel mechanism of no-triggered activation of PKCepsilon. The Journal of biological chemistry. 2002;277:15021–15027. doi: 10.1074/jbc.M112451200. [DOI] [PubMed] [Google Scholar]
- 104.Cassina AM, Hodara R, Souza JM, Thomson L, Castro L, Ischiropoulos H, Freeman BA, Radi R. Cytochrome c nitration by peroxynitrite. The Journal of biological chemistry. 2000;275:21409–21415. doi: 10.1074/jbc.M909978199. [DOI] [PubMed] [Google Scholar]
- 105.MacMillan-Crow LA, Crow JP, Kerby JD, Beckman JS, Thompson JA. Nitration and inactivation of manganese superoxide dismutase in chronic rejection of human renal allografts. Proceedings of the National Academy of Sciences of the United States of America. 1996;93:11853–11858. doi: 10.1073/pnas.93.21.11853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Vadseth C, Souza JM, Thomson L, Seagraves A, Nagaswami C, Scheiner T, Torbet J, Vilaire G, Bennett JS, Murciano JC, Muzykantov V, Penn MS, Hazen SL, Weisel JW, Ischiropoulos H. Pro-thrombotic state induced by post-translational modification of fibrinogen by reactive nitrogen species. The Journal of biological chemistry. 2004;279:8820–8826. doi: 10.1074/jbc.M306101200. [DOI] [PubMed] [Google Scholar]
- 107.Souza JM, Daikhin E, Yudkoff M, Raman CS, Ischiropoulos H. Factors determining the selectivity of protein tyrosine nitration. Arch Biochem Biophys. 1999;371:169–178. doi: 10.1006/abbi.1999.1480. [DOI] [PubMed] [Google Scholar]
- 108.Gow AJ, Duran D, Malcolm S, Ischiropoulos H. Effects of peroxynitrite-induced protein modifications on tyrosine phosphorylation and degradation. FEBS Lett. 1996;385:63–66. doi: 10.1016/0014-5793(96)00347-x. [DOI] [PubMed] [Google Scholar]
- 109.Kamisaki Y, Wada K, Bian K, Balabanli B, Davis K, Martin E, Behbod F, Lee YC, Murad F. An activity in rat tissues that modifies nitrotyrosine-containing proteins. Proceedings of the National Academy of Sciences of the United States of America. 1998;95:11584–11589. doi: 10.1073/pnas.95.20.11584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Beckman JS, Koppenol WH. Nitric oxide, superoxide, and peroxynitrite: the good, the bad, and ugly. Am J Physiol. 1996;271:C1424–1437. doi: 10.1152/ajpcell.1996.271.5.C1424. [DOI] [PubMed] [Google Scholar]
- 111.Comhair SA, Bhathena PR, Farver C, Thunnissen FB, Erzurum SC. Extracellular glutathione peroxidase induction in asthmatic lungs: evidence for redox regulation of expression in human airway epithelial cells. Faseb J. 2001;15:70–78. doi: 10.1096/fj.00-0085com. [DOI] [PubMed] [Google Scholar]
- 112.Halliwell B, Zhao K, Whiteman M. Nitric oxide and peroxynitrite. The ugly, the uglier and the not so good: a personal view of recent controversies. Free Radic Res. 1999;31:651–669. doi: 10.1080/10715769900301221. [DOI] [PubMed] [Google Scholar]
- 113.Ischiropoulos H. Biological tyrosine nitration: a pathophysiological function of nitric oxide and reactive oxygen species. Arch Biochem Biophys. 1998;356:1–11. doi: 10.1006/abbi.1998.0755. [DOI] [PubMed] [Google Scholar]
- 114.Kaminsky DA, Mitchell J, Carroll N, James A, Soultanakis R, Janssen Y. Nitrotyrosine formation in the airways and lung parenchyma of patients with asthma. J Allergy Clin Immunol. 1999;104:747–754. doi: 10.1016/s0091-6749(99)70283-6. [DOI] [PubMed] [Google Scholar]
- 115.Sadeghi-Hashjin G, Folkerts G, Henricks PA, Verheyen AK, van der Linde HJ, van Ark I, Coene A, Nijkamp FP. Peroxynitrite induces airway hyperresponsiveness in guinea pigs in vitro and in vivo. American journal of respiratory and critical care medicine. 1996;153:1697–1701. doi: 10.1164/ajrccm.153.5.8630623. [DOI] [PubMed] [Google Scholar]
- 116.Koarai A, Ichinose M, Sugiura H, Tomaki M, Watanabe M, Yamagata S, Komaki Y, Shirato K, Hattori T. iNOS depletion completely diminishes reactive nitrogen-species formation after an allergic response. Eur Respir J. 2002;20:609–616. doi: 10.1183/09031936.02.00274902. [DOI] [PubMed] [Google Scholar]
- 117.Alvarez B, Demicheli V, Duran R, Trujillo M, Cervenansky C, Freeman BA, Radi R. Inactivation of human Cu,Zn superoxide dismutase by peroxynitrite and formation of histidinyl radical. Free Radic Biol Med. 2004;37:813–822. doi: 10.1016/j.freeradbiomed.2004.06.006. [DOI] [PubMed] [Google Scholar]
- 118.MacMillan-Crow LA, Thompson JA. Tyrosine modifications and inactivation of active site manganese superoxide dismutase mutant (Y34F) by peroxynitrite. Arch Biochem Biophys. 1999;366:82–88. doi: 10.1006/abbi.1999.1202. [DOI] [PubMed] [Google Scholar]
- 119.Castegna A, Aksenov M, Thongboonkerd V, Klein JB, Pierce WM, Booze R, Markesbery WR, Butterfield DA. Proteomic identification of oxidatively modified proteins in Alzheimer’s disease brain. Part II: dihydropyrimidinase-related protein 2, alpha-enolase and heat shock cognate 71. J Neurochem. 2002;82:1524–1532. doi: 10.1046/j.1471-4159.2002.01103.x. [DOI] [PubMed] [Google Scholar]
- 120.Sopkova J, Raguenes-Nicol C, Vincent M, Chevalier A, Lewit-Bentley A, Russo-Marie F, Gallay J. Ca(2+) and membrane binding to annexin 3 modulate the structure and dynamics of its N terminus and domain III. Protein Sci. 2002;11:1613–1625. doi: 10.1110/ps.4230102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Novak Z, Nemeth I, Gyurkovits K, Varga SI, Matkovics B. Examination of the role of oxygen free radicals in bronchial asthma in childhood. Clin Chim Acta. 1991;201:247–251. doi: 10.1016/0009-8981(91)90375-m. [DOI] [PubMed] [Google Scholar]
- 122.Comhair SA, Xu W, Ghosh S, Thunnissen FB, Almasan A, Calhoun WJ, Janocha AJ, Zheng L, Hazen SL, Erzurum SC. Superoxide dismutase inactivation in pathophysiology of asthmatic airway remodeling and reactivity. Am J Pathol. 2005;166:663–674. doi: 10.1016/S0002-9440(10)62288-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.MacMillan-Crow LA, Crow JP, Thompson JA. Peroxynitrite-mediated inactivation of manganese superoxide dismutase involves nitration and oxidation of critical tyrosine residues. Biochemistry. 1998;37:1613–1622. doi: 10.1021/bi971894b. [DOI] [PubMed] [Google Scholar]
- 124.Macmillan-Crow LA, Cruthirds DL. Invited review: manganese superoxide dismutase in disease. Free Radic Res. 2001;34:325–336. doi: 10.1080/10715760100300281. [DOI] [PubMed] [Google Scholar]
- 125.Keng T, Privalle CT, Gilkeson GS, Weinberg JB. Peroxynitrite formation and decreased catalase activity in autoimmune MRL-lpr/lpr mice. Mol Med. 2000;6:779–792. [PMC free article] [PubMed] [Google Scholar]
- 126.Wong PS, Eiserich JP, Reddy S, Lopez CL, Cross CE, van der Vliet A. Inactivation of glutathione S-transferases by nitric oxide-derived oxidants: exploring a role for tyrosine nitration. Arch Biochem Biophys. 2001;394:216–228. doi: 10.1006/abbi.2001.2532. [DOI] [PubMed] [Google Scholar]
- 127.Bergt C, Fu X, Huq NP, Kao J, Heinecke JW. Lysine residues direct the chlorination of tyrosines in YXXK motifs of apolipoprotein A-I when hypochlorous acid oxidizes high density lipoprotein. The Journal of biological chemistry. 2004;279:7856–7866. doi: 10.1074/jbc.M309046200. [DOI] [PubMed] [Google Scholar]
- 128.Liu L, Hausladen A, Zeng M, Que L, Heitman J, Stamler JS. A metabolic enzyme for S-nitrosothiol conserved from bacteria to humans. Nature. 2001;410:490–494. doi: 10.1038/35068596. [DOI] [PubMed] [Google Scholar]
- 129.Molina y Vedia L, McDonald B, Reep B, Brune B, Di Silvio M, Billiar TR, Lapetina EG. Nitric oxide-induced S-nitrosylation of glyceraldehyde-3-phosphate dehydrogenase inhibits enzymatic activity and increases endogenous ADP-ribosylation. The Journal of biological chemistry. 1992;267:24929–24932. [PubMed] [Google Scholar]
- 130.Stamler JS, Toone EJ, Lipton SA, Sucher NJ. (S)NO signals: translocation, regulation, and a consensus motif. Neuron. 1997;18:691–696. doi: 10.1016/s0896-6273(00)80310-4. [DOI] [PubMed] [Google Scholar]
- 131.Ascenzi P, Colasanti M, Persichini T, Muolo M, Polticelli F, Venturini G, Bordo D, Bolognesi M. Re-evaluation of amino acid sequence and structural consensus rules for cysteine-nitric oxide reactivity. Biol Chem. 2000;381:623–627. doi: 10.1515/BC.2000.081. [DOI] [PubMed] [Google Scholar]
- 132.Ckless K, Reynaert NL, Taatjes DJ, Lounsbury KM, van der Vliet A, Janssen-Heininger Y. In situ detection and visualization of S-nitrosylated proteins following chemical derivatization: identification of Ran GTPase as a target for S-nitrosylation. Nitric Oxide. 2004;11:216–227. doi: 10.1016/j.niox.2004.06.002. [DOI] [PubMed] [Google Scholar]
- 133.Hess DT, Matsumoto A, Kim SO, Marshall HE, Stamler JS. Protein S-nitrosylation: purview and parameters. Nature reviews. 2005;6:150–166. doi: 10.1038/nrm1569. [DOI] [PubMed] [Google Scholar]
- 134.Kim SF, Huri DA, Snyder SH. Inducible nitric oxide synthase binds, S-nitrosylates, and activates cyclooxygenase-2. Science (New York, NY. 2005;310:1966–1970. doi: 10.1126/science.1119407. [DOI] [PubMed] [Google Scholar]
- 135.Mayer B, Pfeiffer S, Schrammel A, Koesling D, Schmidt K, Brunner F. A new pathway of nitric oxide/cyclic GMP signaling involving S-nitrosoglutathione. The Journal of biological chemistry. 1998;273:3264–3270. doi: 10.1074/jbc.273.6.3264. [DOI] [PubMed] [Google Scholar]
- 136.Romeo AA, Capobianco JA, English AM. Superoxide dismutase targets NO from GSNO to Cysbeta93 of oxyhemoglobin in concentrated but not dilute solutions of the protein. Journal of the American Chemical Society. 2003;125:14370–14378. doi: 10.1021/ja0289752. [DOI] [PubMed] [Google Scholar]
- 137.Stamler JS. S-nitrosothiols in the blood: roles, amounts, and methods of analysis. Circulation research. 2004;94:414–417. doi: 10.1161/01.RES.0000122071.55721.BC. [DOI] [PubMed] [Google Scholar]
- 138.Kornberg MD, Sen N, Hara MR, Juluri KR, Nguyen JV, Snowman AM, Law L, Hester LD, Snyder SH. GAPDH mediates nitrosylation of nuclear proteins. Nature cell biology. 12:1094–1100. doi: 10.1038/ncb2114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Stamler JS, Hess DT. Nascent nitrosylases. Nature cell biology. 12:1024–1026. doi: 10.1038/ncb1110-1024. [DOI] [PubMed] [Google Scholar]
- 140.Benhar M, Forrester MT, Stamler JS. Protein denitrosylation: enzymatic mechanisms and cellular functions. Nature reviews. 2009;10:721–732. doi: 10.1038/nrm2764. [DOI] [PubMed] [Google Scholar]
- 141.Kashiba-Iwatsuki M, Yamaguchi M, Inoue M. Role of ascorbic acid in the metabolism of S-nitroso-glutathione. FEBS Lett. 1996;389:149–152. doi: 10.1016/0014-5793(96)00560-1. [DOI] [PubMed] [Google Scholar]
- 142.Yates DH. Role of exhaled nitric oxide in asthma. Immunol Cell Biol. 2001;79:178–190. doi: 10.1046/j.1440-1711.2001.00990.x. [DOI] [PubMed] [Google Scholar]
- 143.Zeng H, Spencer NY, Hogg N. Metabolism of S-nitrosoglutathione by endothelial cells. Am J Physiol Heart Circ Physiol. 2001;281:H432–439. doi: 10.1152/ajpheart.2001.281.1.H432. [DOI] [PubMed] [Google Scholar]
- 144.Nikitovic D, Holmgren A. S-nitrosoglutathione is cleaved by the thioredoxin system with liberation of glutathione and redox regulating nitric oxide. The Journal of biological chemistry. 1996;271:19180–19185. doi: 10.1074/jbc.271.32.19180. [DOI] [PubMed] [Google Scholar]
- 145.Trujillo M, Alvarez MN, Peluffo G, Freeman BA, Radi R. Xanthine oxidase-mediated decomposition of S-nitrosothiols. The Journal of biological chemistry. 1998;273:7828–7834. doi: 10.1074/jbc.273.14.7828. [DOI] [PubMed] [Google Scholar]
- 146.Jourd’heuil D, Laroux FS, Miles AM, Wink DA, Grisham MB. Effect of superoxide dismutase on the stability of S-nitrosothiols. Arch Biochem Biophys. 1999;361:323–330. doi: 10.1006/abbi.1998.1010. [DOI] [PubMed] [Google Scholar]
- 147.Reynaert NL, Ckless K, Wouters EF, van der Vliet A, Janssen-Heininger YM. Nitric oxide and redox signaling in allergic airway inflammation. Antioxid Redox Signal. 2005;7:129–143. doi: 10.1089/ars.2005.7.129. [DOI] [PubMed] [Google Scholar]
- 148.Gaston B, Reilly J, Drazen JM, Fackler J, Ramdev P, Arnelle D, Mullins ME, Sugarbaker DJ, Chee C, Singel DJ, et al. Endogenous nitrogen oxides and bronchodilator S-nitrosothiols in human airways. Proceedings of the National Academy of Sciences of the United States of America. 1993;90:10957–10961. doi: 10.1073/pnas.90.23.10957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Gaston B, Sears S, Woods J, Hunt J, Ponaman M, McMahon T, Stamler JS. Bronchodilator S-nitrosothiol deficiency in asthmatic respiratory failure. Lancet. 1998;351:1317–1319. doi: 10.1016/S0140-6736(97)07485-0. [DOI] [PubMed] [Google Scholar]
- 150.Janssen LJ, Premji M, Lu-Chao H, Cox G, Keshavjee S. NO(+) but not NO radical relaxes airway smooth muscle via cGMP-independent release of internal Ca(2+) American journal of physiology. 2000;278:L899–905. doi: 10.1152/ajplung.2000.278.5.L899. [DOI] [PubMed] [Google Scholar]
- 151.Matalon S, Hardiman KM, Jain L, Eaton DC, Kotlikoff M, Eu JP, Sun J, Meissner G, Stamler JS. Regulation of ion channel structure and function by reactive oxygen-nitrogen species. American journal of physiology. 2003;285:L1184–1189. doi: 10.1152/ajplung.00281.2003. [DOI] [PubMed] [Google Scholar]
- 152.Nozik-Grayck E, Whalen EJ, Stamler JS, McMahon TJ, Chitano P, Piantadosi CA. S-nitrosoglutathione inhibits alpha1-adrenergic receptor-mediated vasoconstriction and ligand binding in pulmonary artery. American journal of physiology. 2006;290:L136–143. doi: 10.1152/ajplung.00230.2005. [DOI] [PubMed] [Google Scholar]
- 153.White TA, Walseth TF, Kannan MS. Nitric oxide inhibits ADP-ribosyl cyclase through a cGMP-independent pathway in airway smooth muscle. American journal of physiology. 2002;283:L1065–1071. doi: 10.1152/ajplung.00064.2002. [DOI] [PubMed] [Google Scholar]
- 154.Doctor A, Platt R, Sheram ML, Eischeid A, McMahon T, Maxey T, Doherty J, Axelrod M, Kline J, Gurka M, Gow A, Gaston B. Hemoglobin conformation couples erythrocyte S-nitrosothiol content to O2 gradients. Proceedings of the National Academy of Sciences of the United States of America. 2005;102:5709–5714. doi: 10.1073/pnas.0407490102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Moya MP, Gow AJ, Califf RM, Goldberg RN, Stamler JS. Inhaled ethyl nitrite gas for persistent pulmonary hypertension of the newborn. Lancet. 2002;360:141–143. doi: 10.1016/S0140-6736(02)09385-6. [DOI] [PubMed] [Google Scholar]
- 156.Snyder AH, McPherson ME, Hunt JF, Johnson M, Stamler JS, Gaston B. Acute effects of aerosolized S-nitrosoglutathione in cystic fibrosis. American journal of respiratory and critical care medicine. 2002;165:922–926. doi: 10.1164/ajrccm.165.7.2105032. [DOI] [PubMed] [Google Scholar]
- 157.Gaston B, Drazen JM, Jansen A, Sugarbaker DA, Loscalzo J, Richards W, Stamler JS. Relaxation of human bronchial smooth muscle by S-nitrosothiols in vitro. J Pharmacol Exp Ther. 1994;268:978–984. [PubMed] [Google Scholar]
- 158.Que LG, Liu L, Yan Y, Whitehead GS, Gavett SH, Schwartz DA, Stamler JS. Protection from experimental asthma by an endogenous bronchodilator. Science (New York, NY. 2005;308:1618–1621. doi: 10.1126/science.1108228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Que LG, Yang Z, Stamler JS, Lugogo NL, Kraft M. S-nitrosoglutathione reductase: an important regulator in human asthma. American journal of respiratory and critical care medicine. 2009;180:226–231. doi: 10.1164/rccm.200901-0158OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Gaston B, Drazen JM, Loscalzo J, Stamler JS. The biology of nitrogen oxides in the airways. American journal of respiratory and critical care medicine. 1994;149:538–551. doi: 10.1164/ajrccm.149.2.7508323. [DOI] [PubMed] [Google Scholar]
- 161.Greenwald R, Fitzpatrick AM, Gaston B, Marozkina NV, Erzurum S, Teague WG. Breath formate is a marker of airway S-nitrosothiol depletion in severe asthma. PLoS One. 5:e11919. doi: 10.1371/journal.pone.0011919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Marshall HE, Stamler JS. Inhibition of NF-kappa B by S-nitrosylation. Biochemistry. 2001;40:1688–1693. doi: 10.1021/bi002239y. [DOI] [PubMed] [Google Scholar]
- 163.Kelleher ZT, Matsumoto A, Stamler JS, Marshall HE. NOS2 regulation of NF-kappaB by S-nitrosylation of p65. The Journal of biological chemistry. 2007;282:30667–30672. doi: 10.1074/jbc.M705929200. [DOI] [PubMed] [Google Scholar]
- 164.Olson N, Kasahara DI, Hristova M, Bernstein R, Janssen-Heininger Y, van der Vliet A. Modulation of NF-{kappa}B and HIF-1 by S-Nitrosoglutathione Does Not Alter Allergic Airway Inflammation in Mice. Am J Respir Cell Mol Biol. doi: 10.1165/rcmb.2010-0035OC. [DOI] [PMC free article] [PubMed] [Google Scholar]