

Abstract
Background
Single nucleotide polymorphism (SNP) based association studies aim at identifying SNPs associated with phenotypes, for example, complex diseases. The associated SNPs may influence the disease risk individually (main effects) or behave jointly (epistatic interactions). For the analysis of high throughput data, the main difficulty is that the number of SNPs far exceeds the number of samples. This difficulty is amplified when identifying interactions.
Results
In this paper, we propose an Adaptive Group Lasso (AGL) model for large-scale association studies. Our model enables us to analyze SNPs and their interactions simultaneously. We achieve this by introducing a sparsity constraint in our model based on the fact that only a small fraction of SNPs is disease-associated. In order to reduce the number of false positive findings, we develop an adaptive reweighting scheme to enhance sparsity. In addition, our method treats SNPs and their interactions as factors, and identifies them in a grouped manner. Thus, it is flexible to analyze various disease models, especially for interaction detection. However, due to the intensive computation when millions of interaction terms needs to be searched in the model fitting, our method needs to combined with some filtering methods when applied to genome-wide data for detecting interactions.
Conclusion
By using a wide range of simulated datasets and a real dataset from WTCCC, we demonstrate the advantages of our method.
Background
Rapid Improvements of high-throughput genotyping technologies enable us to detect genetic variations with much finer resolution than before. In genome-wide association (GWA) studies of complex diseases, a few thousands samples are collected and hundreds of thousands of single nucleotide polymorphisms (SNPs) have been genotyped for each sample [1].
Researchers have been investigating disease-associated gene mapping for decades and various approaches have been proposed. However, most of them have used a single-SNP based strategy, in which each SNP is analyzed individually (see [2] for a comprehensive review). Due to the sophisticated regulatory mechanism encoded in the human genome, it is widely agreed that complex traits are typically caused by multiple genetic variations. One type of genetic variation influences the traits individually. This is known as main effects. Another type of genetic variation is that SNPs may show little effect individually, but strong effects jointly. This is known as epistasis or multilocus interactions [3]. Therefore, multi-locus based approaches are believed to have higher power than single-locus based ones. Identifying epistatic interactions arises as an important problem in multi-locus based approaches [4].
Recently, an increasing number of research has reported the presence of epistatic interactions in complex diseases, such as type-2 diabetes [5]. In order to detect epistatic interaction, various computational and statistical methods have developed [4]. For example, Nelson et al. [6] proposed a combinatorial partitioning method (CPM) that enumerated multi-locus genotypes and evaluated them with phenotypes. Culverhouse et al. [7] proposed a restricted partitioning method (RPM) to improve the efficiency of CPM. Millstein et al. [8] developed a testing framework when epistasis is present. Ritchie et al. [9] proposed a multifactor-dimensionality reduction (MDR) method that identified interactions based on classification accuracy through exhaustive search. Zhang and Liu [10] proposed a Bayesian epistasis association mapping (BEAM) method to address the issue of epistasis mapping in genome-wide scale by using Markov Chain Monte Carlo (MCMC) method. In spite of their promising performance, most of these methods only show their successes in association studies on small-scale data sets.
From our view, detecting disease-associated SNPs and their interactions can be cast as the variable selection problem in the framework of regression analysis. Standard tools for variable assessment are the methods of multivariate regression. In traditional applications of multivariate regression, the number of variables is less than the number of samples. In the context of SNP-based disease association studies, however, the number of SNPs is far more than the number of samples, making it difficult or even impossible to directly apply standard multivariate regression methods.
It is widely agreed in GWA studies that only a small fraction of SNPs is disease-associated. In the multivariate regression framework, this implies that most regression coefficients should be zero. This motivates us to impose a sparsity constraint to the regression model. In addition, SNPs are bi-allelic markers (i.e., with allele A and a). Each SNP has only three genotypes: two homozygous genotypes (AA and aa) and one heterozygous genotype (Aa). Therefore, each SNP can be naturally treated as a three-level factor and be coded with three dummy variables. Similarly, the interaction between two SNPs can be treated as a nine-level factor. In order to encourage sparsity on factors (groups of variables) rather than a single dummy variable, we impose a group constraint on the set of dummy variables that represent a disease model (e.g., a single locus model or a two-locus model). Hence, we propose an Adaptive Group Lasso (AGL) method to identify main effects and epistatic interactions from large-scale SNP data. Since Lasso [11] is well known for imposing a sparsity constraint at the variable level, we employ Group Lasso [12,13] to impose the sparsity constraint at the factor level, and develop adaptive reweighting to enhance the sparsity and to reduce false positive finding.
Results and discussion
In this section, we evaluate the performance of our method using both simulated and real data. In simulation studies, we compare our method with some recent competitors under a wide range of epistatic models. For the real case-control study, we use the rheumatoid arthritis (RA) data set from the Wellcome Trust Case Control Consortium (WTCCC).
Simulation studies
In simulation studies, we mainly compare our method AGL with Lasso, BEAM and MDR.
We choose Lasso [14,15] for comparison due to the close relationship between our method and the two Lasso methods. For the identification of main effects, they presume additive, dominant or recessive effects when fitting the Lasso model. For the identification of interactions, Hoggart et. al [14] do not consider this issues and Wu et. al [15] only restrict themselves to the SNPs with strong main effects. Our method is different from their methods in the following sense:
(1) We do not presume particular types of main effects and interactions. Thus, our model is more flexible.
(2) We impose a sparsity constraint at the factor level instead of at the variable level.
(3) Our model includes all possible interactions and is able to identify interactions with weak main effects.
We also compare with BEAM [10] which arises as a powerful epistasis mapping method. Both methods share the concept of three SNP classes: unassociated SNPs, SNPs with main effects and SNPs with interactions. BEAM builds a Bayesian partition model based on these three classes. It is worth mentioning that there is only a single group of interacting SNPs in the BEAM model. To identify multiple interacting groups, BEAM implicitly makes use of MCMC to visit possible interactions. We explicitly allow multiple groups of interacting SNPs and impose additive effects between those groups. Comprehensive comparison studies between BEAM and other related methods have been carried out in [10].
Due to the limited space, the comparison with MDR is given in the supplementary. We further conduct null simulation to estimate the type I error rate of our method.
In the following experiments, we use five-fold cross-validation in model fitting process. We use Bonferroni correction to adjust our p-value and set the significance threshold as 0.3 in simulation studies. In the released version of BEAM, the threshold is set as 0.3. Thus, we choose the same threshold.
Comparison with Lasso
We conduct experiments under two scenarios.
• Scenario 1: Identification of main effects.
To illustrate our point, we consider two disease models M1-1 and M1-2, as given in Table 1. M1-1 is a multiplicative model used in both [16] and [10]. M1-2 is proposed in [17] to exhibit the interference effect. We choose these two models with different minor allele frequencies (MAF) to illustrate the influence of model specification when identifying main effects. Under each model setting, we generate 100 data sets which contains 1000 SNPs.
Table 1.
Two epistasis models (left: M1-1; right: M1-2).
| Model 1-1 | AA | Aa | aa | Model 1-2 | AA | Aa | aa |
|---|---|---|---|---|---|---|---|
| BB | α | α | α | BB | α | α | α |
| Bb | α | α(1 + θ)2 | α(1 + θ)3 | Bb | α | α(1 + θ) | α |
| bb | α | α(1 + θ)3 | α(1 + θ)4 | bb | α | α | α(1 + θ) |
The performance of Lasso and AGL is summarized in Fig. 1. The power of each method is calculated as the ratio between the number of successful identifications of disease loci and the number of data sets. Lasso performs slightly better than our method for model M1-1. But it performs much worse than our method for model M1-2. Here are the reasons: Firstly, we impose additive effects of SNPs in Lasso model fitting and then perform statistical tests with df = 1. Secondly, for AGL we do not assume additive effects of SNPs but use a more general model structure (see our model (2) in Method) and perform statistical tests with df = 2. Therefore, Lasso performs better under M1-1 since the imposed additive structure in Lasso agrees well with the structure in model M1-1. M1-2 exhibits interference effect which can not be well approximated by additive, dominant or recessive effect. Lasso performs much worse than Adaptive Group Lasso due to the model mismatch.
Figure 1.

The performance comparison between Lasso and AGL on identification of main effects. We generate 100 replicates under each setting. Both 1000 samples and 2000 samples with balanced design are simulated. For Model 1-1, the main effect can be well approximated by the additive effect. Thus, Lasso outperforms AGL slightly. For Model 1-2, the main effect can not be approximated by additive, dominant or recessive effect. Thus, AGL outperforms Lasso significantly.
• Scenario 2: Identification of interaction effects.
The model mismatch problem of Lasso is more serious when identifying interactions. Here we consider four epistatic models M1-3 ~ M1-6 used in [18], as given in Table 2. Here we report the performance of Lasso and AGL under these four models. The comparison results of other epistatic models in [18] are similar. For Lasso method, we take different main effects and their interactions into consideration during Lasso model fitting. Here we use our interaction model (see model (7) in Method). We simulate two associated SNPs based on the disease models, and gradually increase the number of noise SNPs. Fig. 2 summarizes the experimental results. The power is calculated as the proportion of the 100 data sets in which interactions of the disease associated SNPs are detected. Lasso with presumed main effects (additive, dominant or recessive) loses its power rapidly as the number of noise SNPs increases, while AGL keeps its power when more noise SNPs are involved in model fitting.
Table 2.
Four pure epistatisis models used in [18].
| Model 1-3 | h2= 0.3, pa = 0.4, qb = 0.4 | Model 1-4 | h2= 0.2, pa= 0.4, qb= 0.4 | ||||
|---|---|---|---|---|---|---|---|
| AA | Aa | aa | AA | Aa | aa | ||
| BB | 0.077 | 0.689 | 0.417 | BB | 0.086 | 0.536 | 0.641 |
| Bb | 0.763 | 0.150 | 0.491 | Bb | 0.677 | 0.275 | 0.096 |
| bb | 0.196 | 0.657 | 0.247 | bb | 0.219 | 0.413 | 0.712 |
| Model 1-5 | h2 = 0.1, pa = 0.4, qb = 0.4 | Model 1-6 | h2 = 0.05, pa = 0.4, qb = 0.4 | ||||
| AA | Aa | aa | AA | Aa | aa | ||
| BB | 0.068 | 0.299 | 0.017 | BB | 0.005 | 0.179 | 0.251 |
| Bb | 0.289 | 0.044 | 0.285 | Bb | 0.211 | 0.100 | 0.026 |
| bb | 0.048 | 0.262 | 0.174 | bb | 0.156 | 0.098 | 0.156 |
Here we only provide four pure epistatisis models used in comparison with Lasso. The complete model list used in comparison with BEAM is provided in our supplementary document (please see Additional File 1).
Figure 2.

The performance comparison between Lasso and AGL on pure epistasis model. Different main effects (additive, dominant and recessive) and their interactions are taken into consideration when fitting the Lasso model. The number in the x-axis is the number of SNPs simulated in the experiments. Lasso with a presumed model performs poorly with increasing number of noise SNPs, while AGL is robust under all settings.
Generally speaking, if the underlying interaction could be well characterized by Lasso with a presumed model structure, e.g., additive model, then the statistical power of Lasso would be higher than that of AGL because Lasso uses less degree of freedom. However, since the underlying interaction is generally unknown and its possible pattern may cover a wide range of spectrum [19], AGL can serves as a valuable tool for discovering interactions in larger model space. Hence, AGL and Lasso may be complementary to each other in GWA studies.
We show the effect of adaptive reweighting in Fig. 3. The first reweighting greatly reduces the number of selected dummy variable groups and the reweighting process converges in a few iterations (typically less than 5 iterations). The adaptive reweighting process reduces the number of unassociated groups and leads to more accurate p-value calculation in the statistical testing.
Figure 3.

The reweighting effect of Adaptive Group Lasso. The reweighting greatly reduces the number of selected SNP pairs after the first iteration. This effect is more obvious when identifying interactions in M1-3, M1-4, M1-5, M1-6.
On the other hand, however, it can also be seen that unassociated groups may enter the final model even after adaptive reweighting. Hence, the selected groups in the final model may not be associated with the phenotype. In this regard, the significance assessment is critically needed.
Comparison with BEAM
We shall not compare our method with BEAM when genetic heterogeneities are present since BEAM is not developed to handle these cases (the authors of BEAM had made it clear in [10]). We shall compare with BEAM from two perspectives:
1. The ability of detecting epistatic interactions when the main effect is weak or even absent.
2. The ability of detecting multiple interactions.
Detecting epistatic interactions with weak main effect
A wide range of interaction models without marginal effects has been discussed in [19]. Here we consider the 40 pure epistatic models in [18] to compare the performance between AGL and BEAM. The details of these models are available in the supplementary document. The heritability h2(see definition in [19]) of these 40 models ranges from 0.05 to 0.2 and the
MAF ranges from 0.2 to 0.4. We use 100 data sets for each disease model. There are 200 cases, 200 controls, and 1000 SNPs in each data set.
The comparison between our method and BEAM in Fig. 4 shows that AGL is superior to BEAM for detecting epistatic interactions without main effects. For the models with MAF = 0.2, 0.4 and h2 ≥ 0.1, the power of our method is above 95%, while that of BEAM is roughly 20%. The performances of the two methods degrade as the heritability h2 decreases: the power of BEAM is lower than 5% for the models with MAF = 0.2 and h2 ≤ 0.1, while the power of our method still remains at about 75% for some of these models and is even higher for the models with MAF = 0.4.
Figure 4.

The performance comparison between AGL and BEAM (B) based on pure epistatic models. We generate 100 datasets for each model. Each dataset contains 400 sample (Nu = 200, Nd = 200) and 1000 SNPs. BEAM runs 5 × 106 Markov Chain Monte Carlo iterations which are 10 times of pairwise exhaustive search. The comparison shows that our method outperforms BEAM for these pure epistatic models.
Our model includes all possible interactions (1000 × 999/2 interactions) in the model fitting process, so there is no chance to miss interesting interactions. The good performance of our model is due to the group-sparsity constraint: It identifies interactions in a grouped manner. This is very helpful to weaken the influence of noise SNPs.
The poor performance of BEAM is not due to the statistical testing power of the B-statistics [10], but the sampling efficiency. We carefully examined the interactions in the disease models with h2 ≥ 0.1: Those interactions are very significant even after the Bonferroni correction of B-statistics. Notice that BEAM runs 5 × 106 MCMC iterations which are 10 times of pairwise exhaustive search in the simulation study. Thus, our conjecture is that MCMC might converge too slowly to find the ground truth (We provide some evidence in the supplementary to support our conjecture).
Detecting multiple interactions
Another disadvantage of BEAM is that it only allows a single interacting group in its Bayesian partition model. To compensate for this limitation, BEAM uses MCMC sampling strategy to visit possible interactions during model optimization. In contrast, our approach allows multiple interacting groups and imposes additive effects for these interactions. This flexibility enables us to have a higher power to identify multiple interactions. Due to the limited space, we show our comparison result of detecting multiple interactions in the supplementary.
Null simulation study
To validate the use of our p-value and to estimate the type-I error, we conduct null simulation studies in two cases:
• Case 1: We generate 100 null datasets. Each dataset contains 10 K SNPs and 1000 samples. All the SNPs are generated independently with MAF uniformly distributed in [0.05, 0.5]. In this case, the nominal type-I error rates should be 10, 20, 30 per one million SNPs for significance thresholds at 0.1, 0.2, 0.3.
• Case 2: We use genomeSIMLA [20] to simulate the SNP data based on the marker information on the Affymetrix 500 K chip from human chromosome 1. Linkage disequilibrium (LD) exists among SNPs. We also generate 100 null datasets, each of which contains 38836 SNPs and 1000 samples. Due to LD pattern, the error rate should lower than the nominal error rate.
We summarize the type-I error of our model (2) in Table 3. For Case 1, Our results are reasonable for the three nominal levels. For Case 2, our type-I error rates show that our method is conservative when LD exists.
Table 3.
Null simulation: empirical type-I error from 100 simulated data sets.
| Cases | Error rates per one million SNPs of different thresholds after Bonferroni correction | ||
| threshold = 0.1 | threshold = 0.2 | threshold = 0.3 | |
| Case 1 | 11 | 18 | 28 |
| Case 2 | 3.08 | 4.89 | 7.47 |
The nominal type-I error rates should be 10, 20, and 30 per one million SNPs for significance thresholds at 0.1, 0.2, and 0.3, respectively.
Analysis of WTCCC data
Main effects
Current version of BEAM software can not handle WTCCC data on the genome-wide scale. To compare with BEAM on the real data, we apply our main effect model (2) and BEAM to analyze WTCCC Rheumatoid Arthritis (RA) data in the chromosome-wise manner. For BEAM, we run 108 MCMC and set the significant threshold as 0.3 after Bonferroni correction. BEAM does not report any interactions. The identified main effects of the two methods agree with each other. Some of them are given in Table 4 and the details are given in the supplementary document. The significant result from WTCCC [1] could be reproduced.
Table 4.
Some significant SNPs identified by our method on WTCCC RA data. These SNPs covers the regions which are moderate associated with RA as reported in [1].
| SNPs | Location | Related Genes | P-value |
|---|---|---|---|
| rs7539166 | 1p36 | TMEM51 | < 10-30 |
| rs384843 | 1p36 | NBPF3 | 5.2 × 10-9 |
| rs7516721 | 1p36 | GPR3 | 7.7 × 10-14 |
| rs4959053 | 6p21 | PSORS1C1 | < 10-30 |
| rs6457617 | 6p21 | MHC region | 1.3 × 10-15 |
| rs6973565 | 7q32 | PLXNA4 | 1.2 × 10-9 |
| rs10250029 | 7q32 | CHCHD3 | < 10-30 |
| rs10751815 | 10p15 | ADARB2 | < 10-30 |
| rs4266996 | 10p15 | unknown | < 10-30 |
| rs17147777 | 10p15 | unknown | < 10-30 |
| rs8129909 | 21q22 | IGSF5 | < 10-30 |
| rs16999716 | 21q22 | DSCAM | 9.2 × 10-9 |
| rs4542939 | 21q22 | DSCAM | 8.3 × 10-9 |
| rs13047947 | 21q22 | PDE9A | < 10-30 |
| rs140344 | 22q12 | unkown | < 10-30 |
| rs5749509 | 22q12 | SYN3 | < 10-30 |
| rs6518796 | 22q12 | SYN3 | < 10-30 |
• WTCCC reports that SNP rs6457617 located at 6p21 shows a very strong association. Our experiment verifies this result.
• We do not identify SNP rs6679677 located at 1p13 reported by WTCCC. Instead, we identify two SNPs rs7551793 and rs948620 near SNP rs6679677. The signals of these two SNPs are much stronger than rs6679677.
• We also identify SNPs in the moderate association regions reported in [1]. We summarize the result in Table 4.
Interactions
Detecting interactions in genome-wide scale is very challenging and multi-stage strategies are often explored. For example, MDR [9] usually is combined with TuRF [21] which serves as a filter to remove those noise SNPs. Currently, our method AGL can not be directly applied to genome-wide scale SNP data since it is too computationally intensive to exhaustively search for all SNP pairs. As suggested in simulation study, our method keeps its statistical power when about 500,000 SNP pairs are considered in our model. Thus, the main difficulty is the computation burden of searching for all SNP pairs. Thus, a filtering method is necessary for our method.
For identification of epistatic interactions, we focus on two candidate regions: 6p21 and 7p21. These two regions are reported in our previous work named SNPHarvester [22] which is a filtering method. Here we apply our interaction model (7) and report some SNP interactions in Table 5.
Table 5.
Some significant SNP groups identified by our method in the candidate regions on WTCCC RA data.
| SNP Groups | Location | Related Genes | P-value |
|---|---|---|---|
| (rs4988822, rs3135392) | 6p21.3 | (HLA-DRA, HLA-DRA) | 3.33 × 10-15 |
| (rs17429127, rs2157082) | 6p21.3 | (HLA-DRA, HLA-DRA) | < 10-30 |
| (rs1358169, rs6460831) | 7p21.3 | (THSD7A, THSD7A) | < 10-30 |
• For the region 6p21, we select a segment covering the SNPs reported in [22]. This segment contains 250 SNPs from rs3135366 to rs461338. We enumerate all possible interactions and include them in our model. Our method reports two interacting pairs: (rs4988822, rs3135392) and (rs17429127, rs2157082). These SNPs are related to gene HLA-DRA. The result in [23] reports that there is a strong association between RA and HLA gene family. Notice that the two SNPs rs4988822 and rs3135392 cannot be identified by univariate analysis due to their weak main effects (Their p-value is at the level of 10-3 given by univariate analysis). However, they do show a strong interaction. We also run BEAM on the selected segment (5 × 106 MCMC). BEAM reports (rs9469220, rs3957146) and (rs3129872, rs9272723) as the two most significant interactions based on the B-statistics. We carefully check these two pairs based on the logistic regression model. We find that the interaction of the SNP pair (rs9469220, rs3957146) is very weak using the standard χ2 test based on logistic regression models with df = 4, while the interaction of the SNP pair (rs3129872, rs9272723) is strong. We further explore the reason why our method does not report the interacting pair (rs3129872, rs9272723). We observe that SNP rs9268645, which is highly correlated (strong LD) with SNP rs3129872, enters our model as a main effect term. Consequently, the pair (rs3129872, rs9272723) does not enter the model as a interaction term. This shows that the SNP pair (rs3129872, rs9272723) should not be reported as an interacting pair.
• For the region 7p21, we select a segment which covers the SNPs reported in [22]. The segment contains 250 SNPs from SNP rs1076224 to SNP rs1548882. We analyze this region and report one interacting pairs (rs1358169, rs6460831). The two SNPs rs1358169 and rs2526100 are related to gene THSD7A on chromosome 7, which has been reported to be associated with bone mineral density [24]. This shows plausible biological relevance. We also run BEAM on the segment. But it does not report any interaction.
• Neither BEAM nor our method finds significant interactions in the region 6q23.
The definition of interactions is not consistent in literatures. For example, the interaction effect of two SNPs is define via logistic regression models of genotypes and their combinations [3], while it is also defined via models of haplotypes [25]. The interactions reported above are based on the definition of interaction of a SNP pair in [3]. We extend this definition such that we can simultaneously handle interactions of multiple SNP pairs. The details are given in the Method section. We realize that the interacting SNPs reported above are close to each other on the physical map. This type of interaction effects may be caused by the haplotype effect. Detecting interactions of genes in different genome regions by analyzing genome-wide SNP data is still under investigation [4].
Conclusion
In this paper, we proposed an Adaptive Group Lasso method for large-scale SNP data analysis. The novelty of our method is that it analyzes SNPs and their interactions simultaneously. It imposes a sparsity constraint at the group level and enables us to identify associated SNPs (especially for interacting SNPs) from large-scale SNP data robustly. We show that our method outperforms its recent competitors in both simulation studies and real application.
The limitation of our method is that the interaction model can not be directly applied to genome-wide scale SNP data analysis. The main difficulty comes from the computation burden of searching for all SNP pairs. There are two possible solutions to solve this issue. One solution is to make use of some filtering method to reduce the number of SNPs to a manageable size, for example, [22]. Another solution is incorporating biological information. Pathway information [26] provides a biological clue to narrow down the search range for interaction detection. We shall investigate this in our future work.
Methods
SNPs are high-density bi-allelic markers. We use capital letters (e.g., A and B) and lowercase letters (e.g., a and b) to denote major and minor alleles, respectively. We also use G1 to denote the collection of all three genotypes of one SNP and use G2 to denote the collection of all nine combinations of two SNPs:
| (1) |
Our target is to identify disease-associated SNPs and their interactions. Researchers may have different understandings of epistatic interaction. To be clear, Our definition of interaction refers to the deviation from the additive model of multiple SNPs. In other words, the interaction effects refers to the phenotype variation that can be explained by joint effects of multiple SNPs but not by their main effects. This is consistent with the definition given in [3]. We achieve our goal through addressing the following key issues:
• Model: How to model the relationship between genotype and phenotype?
• Optimization: How to optimize the model structure?
• Significance assessment: How to test the significance of the detected association?
In the following, we first present our model for identification of main effects and epistatic interactions. Then, we give an optimization algorithm for large-scale SNP data analysis. Finally, we describe a method to assess the significance of the SNPs identified by our model.
The adaptive group Lasso model for identification of main effects
Suppose N samples with Nd cases and Nu controls have been genotyped at L loci for an association study. Now we have a design matrix X collecting these N samples and a response variable y ∈ ℝN indicating a sample from case or control. Since A SNP has only three genotypes, we treat it as a factor and code it with three dummy variables. We use [1 0 0] to code genotype AA, [0 1 0] to code genotype Aa and [0 0 1] to code genotype aa. Then the design matrix X becomes a N × (L × 3) matrix. We shall use Xj, a submatrix of size N × 3, to denote the columns of X corresponding to the j-th SNP. Similarly, Xij, a submatrix of size 1 × 3, corresponds to the i-th sample and the j-th SNP. We propose an Adaptive Group Lasso logistic regression model (AGL) for main effect identification:
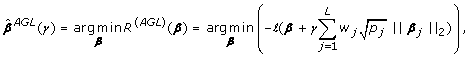 |
(2) |
where γ is a real value parameter controlling the trade-off between the likelihood and the constraint, pj = 3 is the number of dummy variables used to coding the j-th SNP for j = 1, ⋯, L, βj = [βj, AA, βj, Aa, βj, aa], 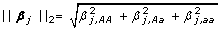 , and l(β) is the log-likelihood of logistic regression:
, and l(β) is the log-likelihood of logistic regression:
 |
(3) |
For convenience, we define an active set 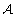 = {j|βj ≠ 0}. We use an iterative algorithm to adaptively assign weights wj in (2) as given in Algorithm 1.
= {j|βj ≠ 0}. We use an iterative algorithm to adaptively assign weights wj in (2) as given in Algorithm 1.
From the iterative algorithm, the penalty weight is adjusted according to its previous estimation. If the current estimation 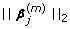 is small, i.e., SNPj is less likely to be associated with disease, then the penalty weight should increase to prevent SNPj from entering the model in the next iteration, and vice versa. The theoretical justification is given in the supplementary document.
is small, i.e., SNPj is less likely to be associated with disease, then the penalty weight should increase to prevent SNPj from entering the model in the next iteration, and vice versa. The theoretical justification is given in the supplementary document.
Algorithm 1 Adaptive Reweighing Algorithm of Group Lasso
1. Set the iteration count m = 0. Initially set  = 1, j = 1, ⋯, L.
= 1, j = 1, ⋯, L.
2. Solve problem (2) to obtain β( ) and the active set
) and the active set 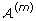 , where is determined by cross-validation.
, where is determined by cross-validation.
3. (1) Update the weight:
| (4) |
(2) Remove SNPj, when j ∉ .
4. If the active set does not change, stop; otherwise increment m and go to step 2.
The proposed model has following characteristics:
• Flexibility: Due to the dummy variable representation, our model is flexible to analyze different main effects (additive, dominant, recessive, interference [17]) in a unified way without presuming one particular type of effect. This flexibility will be pronounced when identifying epistatic interactions.
• Sparsity: The sparsity constraint 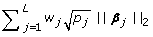 comes from the fact that most SNPs are unassociated. Notice that imposes a constraint to select a group of dummy variables rather than a single one. The weight wj is assigned adaptively in Algorithm 1 to enhances the sparsity at the factor level. If without reweighting, too many noise SNPs would enter the model in GWA studies.
comes from the fact that most SNPs are unassociated. Notice that imposes a constraint to select a group of dummy variables rather than a single one. The weight wj is assigned adaptively in Algorithm 1 to enhances the sparsity at the factor level. If without reweighting, too many noise SNPs would enter the model in GWA studies.
The adaptive group Lasso model for identification of epistatic interactions
Definition of interactions
Interactions between SNP1 and SNP2 are often defined via logistic regression models as in [3]. Let 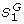 and
and 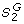 be the dummy variable coding for genotype G of SNP1 and SNP2, respectively. The main effect logistic regression model of SNP1 and SNP2 is:
be the dummy variable coding for genotype G of SNP1 and SNP2, respectively. The main effect logistic regression model of SNP1 and SNP2 is:
| (5) |
The full logistic regression model of SNP1 and SNP2 is:
| (6) |
Let LM and LF be the log likelihood of the main effect model and the full model, respectively. According to the likelihood ratio test, interaction effects are defined via the difference of the log likelihood of these two models, i.e., LF - LM. Hence, interaction effects can be interpreted as departure from linear models naturally [4].
Modelling interactions
The model for identification of epistatic interactions is an extension of model (2): Interaction terms are further included in a grouped manner. We treat the combination of two SNPs as a 9-level factor and use 9 dummy variables to code them. Let J1 be the index set of all SNPs:
J1 ≜ {1, 2,⋯, L}, and J2 be the index set of all pairwise interactions of L SNPs:
J1 ≜ {(1, 2), (1, 3)⋯, (L - 1, L)}. The design matrix X becomes a N × (L × 3 + L × (L - 1)/2) matrix, i.e., 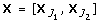 , where
, where 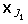 and
and  are the sub design matrices collecting main effect groups and interaction groups, respectively. Similarly, We use
are the sub design matrices collecting main effect groups and interaction groups, respectively. Similarly, We use  with j1 ∈ J1 to denote the i-th sample and the j1-th group in and
with j1 ∈ J1 to denote the i-th sample and the j1-th group in and 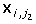 with j2 ∈ J2 to denote the j2-th group in . We propose the following model to identify epistatic interactions:
with j2 ∈ J2 to denote the j2-th group in . We propose the following model to identify epistatic interactions:
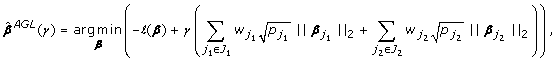 |
(7) |
where 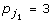 for j1 ∈ J1,
for j1 ∈ J1, 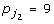 for j2 ∈ J2,
for j2 ∈ J2,  (definitions of G1 and G2 are given in Eqs. (1)), and ℓ(β) is the log-likelihood of logistic regression:
(definitions of G1 and G2 are given in Eqs. (1)), and ℓ(β) is the log-likelihood of logistic regression:
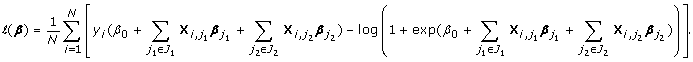 |
(8) |
The proposed model structure has the following characteristics:
• Our interaction model integrates the analysis of SNPs with main effects and interacting SNPs, and unassociated SNPs are prevented from entering the model by the sparsity constraint. Recall that interaction refers to the phenotype variation that can be explained by joint effects of multiple SNPs but not by their main effects, model (7) includes main effects to discourage the interaction terms to enter the model, when their effects can be mostly explained by the main effects. However, this may not be able to completely prevent spurious interactions from entering the model. The reason is that the interaction terms may explain more variances than their corresponding main effects. Hence, they are more likely to enter the final model even when they are penalized more heavily ( for j1 ∈ J1, for j2 ∈ J2). To overcome this difficulty, we resort to the statistical test of interaction effects.
• By using dummy variables, our model is flexible to model various interactions, including all epistatic models described in [17]. Both main effect terms and interaction terms enter the model in a grouped manner. Thus, our model is insensitive to the noise occurring at one level of the genotype combinations G1 and G2.
• Our model imposes additive effects for these interactions. Simultaneous analysis of all interactions achieves under this model structure.
Optimization algorithm
In our Adaptive Reweighing Algorithm of Group Lasso (Algorithm 1), we need an algorithm to solve optimization problem (2) efficiently. We make use of the coordinate descent algorithm [13,27]. The advantage of the coordinate descent algorithm is that it has a closed-form solution of the least square problem when updating one group at a time. Therefore, it is suitable for large-scale data analysis. For the log-likelihood of logistic regression, the iteratively reweighed least square algorithm (IRLS) is efficient: A quadratic approximation is formed to the log-likelihood based on current estimation and the least square problem is solved by the coordinate descent algorithm. This process is repeated until its convergence which is guaranteed [13]. The detail of the algorithm is given in the supplementary.
Statistical testing
The statistical testing is to determine whether the identified SNPs (i.e., the SNPs in the active set) are significantly associated with the disease
Significance tests of main effects
Let 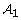 be the SNP set identified under model (2) and s1 denote the number of identified SNPs in . We use the following strategy to assess the significance of identified SNPs:
be the SNP set identified under model (2) and s1 denote the number of identified SNPs in . We use the following strategy to assess the significance of identified SNPs:
1. Re-fit the logistic regression model for the identified SNPs in and obtain the log-likelihood ℓfull.
2. Leaving SNPj ∈ (j = 1, ..., s1) out, fit the logistic regression models and obtain the log-likelihood ℓ\j.
3. Obtain p-value of SNPj using χ2 test based on the value 2(ℓfull - ℓ\j) with the degree of freedom (df) discussed below.
There are several key issues in the above procedure:
1. Since we use three dummy variables to code each SNP, collinearity exists when fitting logistic regression models. To overcome this difficulty, we fit the logistic regression model with a L2 regularization term
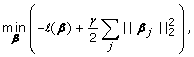 |
(9) |
where γ is a small number to avoid singularity. Here we set γ = 10-4.
2. For model (9), the effective degree of freedom is given by
| (10) |
where Γ is a (s1 + 1) × (s1 + 1) diagonal matrix with diagonal elements [0, γ, ..., γ], U is a diagonal matrix with diagonal elements pi(1 - pi). Here 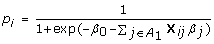 is evaluated after convergence of the logistic regression model (9) fitting.
is evaluated after convergence of the logistic regression model (9) fitting.
3. The degree of freedom of the χ2 test for SNPj is 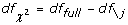 , where dffull and df\j are the effective degrees of freedom of the logistic regression models with all s1 SNPs and without SNPj, respectively.
, where dffull and df\j are the effective degrees of freedom of the logistic regression models with all s1 SNPs and without SNPj, respectively.
It is worth mentioning that our p-value is obtained after the selecting process in AGL fitting. The selecting process may affect the precision of p-value estimation. We justify our p-value by conducting null simulation later.
Significance tests of epistatic interactions
Our Tests of interactions is built upon the definition of interactions. The key point is that the main effects of the two SNPs should not be taken into account when testing their interaction effect. We conduct significance tests of epistatic interactions in the following way:
Let  be the set of the groups identified under model (7) and s2 is the number of the groups in .
be the set of the groups identified under model (7) and s2 is the number of the groups in .
1. Re-fit the logistic regression model for the identified groups in and obtain the log-likelihood ℓfull.
2. For each interaction term in with index l (l = 1, ..., s2), fit the logistic regression model with the main effect term replacing the interaction term and obtain the log-likelihood ℓ\l.
3. Obtain p-value of interaction term l using χ2 tests based on the value 2(ℓfull-ℓ\l) with df = dffull - df\l.
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
CY performed the implementations and drafted the manuscript. XW participated in the experimental design. QY, HX and WY finalized the manuscript. All authors read and approved the final manuscript.
Supplementary Material
The supplementary document provide some backgrounds of our method. It also provides details of our optimization algorithm, and comprehensive experimental results.
Contributor Information
Can Yang, Email: eeyang@ust.hk.
Xiang Wan, Email: eexiangw@ust.hk.
Qiang Yang, Email: qyang@ust.hk.
Hong Xue, Email: hxue@ust.hk.
Weichuan Yu, Email: eeyu@ust.hk.
Acknowledgements
This work was partially supported with the Grant GRF621707 from the Hong Kong Research Grant Council, grant RPC06/07.EG09, RPC07/08.EG25, and a postdoctoral fellowship award from the Hong Kong University of Science and Technology.
This article has been published as part of BMC Bioinformatics Volume 11 Supplement 1, 2010: Selected articles from the Eighth Asia-Pacific Bioinformatics Conference (APBC 2010). The full contents of the supplement are available online at http://www.biomedcentral.com/1471-2105/11?issue=S1.
References
- WTCCC. Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature. 2007;447:661–678. doi: 10.1038/nature05911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balding D. A tutorial on statistical methods for population association studies. Nature Reviews Genetics. 2006;7:781–791. doi: 10.1038/nrg1916. [DOI] [PubMed] [Google Scholar]
- Cordell H. Epistasis: what it means, what it doesn't mean, and statistical methods to detect it in humans. Human Molecular Genetics. 2002;11:2463–2468. doi: 10.1093/hmg/11.20.2463. [DOI] [PubMed] [Google Scholar]
- Cordell HJ. Detecting gene-gene interactions that underlie human diseases. Nat Rev Genet. 2009;10:392–404. doi: 10.1038/nrg2579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cho Y, Ritchie M, Moore J, Park J, Lee KU, Shin H, Lee H, Park K. Multifactor-dimensionality reduction shows a two-locus interaction associated with Type 2 diabetes mellitus. Diabetologia. 2004;47:549–554. doi: 10.1007/s00125-003-1321-3. [DOI] [PubMed] [Google Scholar]
- Nelson M, Kardia S, Ferrell R, Sing C. Combinatorial partitioning method to identify multilocus genotypic partitions that predict quantitative trait variation. Genome Research. 2001;11:458–470. doi: 10.1101/gr.172901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Culverhouse R, Klein T, Shannon W. Detecting epistatic interactions contributing to quantitative traits. Genetic Epidemiology. 2004;27:141–152. doi: 10.1002/gepi.20006. [DOI] [PubMed] [Google Scholar]
- Millstein J, Conti D, Gilliland F, Gauderman W. A testing framework for identifying susceptibility genes in the presence of epistasis. Am J Hum Genet. 2006;78:15–27. doi: 10.1086/498850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ritchie M, Hahn L, Roodi N, Bailey L, Dupont W, Parl F, Moore J. Multifactor-dimensionality reduction reveals high-order interactions among estrogenmetabolism genes in sporadic breast cancer. Am J Hum Genet. 2001;69:138–147. doi: 10.1086/321276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Liu J. Bayesian inference of epistatic interactions in case-control studies. Nature Genetics. 2007;39:1167–1173. doi: 10.1038/ng2110. [DOI] [PubMed] [Google Scholar]
- Tibshirani R. Regression shrinkage and selection via the Lasso. Journal of the Royal Statistical Society, series B. 1996;58:267–288. [Google Scholar]
- Yuan M, Lin Y. Model selection and estimation in regression with grouped variables. Journal of the Royal Statistical Society: Series B. 2006;68:49–67. doi: 10.1111/j.1467-9868.2005.00532.x. [DOI] [Google Scholar]
- Meier L, Geer S, Buhlmann P. The group lasso for logistic regression. Journal of the Royal Statistical Society: Series B. 2008;70:53–71. [Google Scholar]
- Hoggart C, Whittatker J, Iorio M, Balding D. Simultaneous Analysis of All SNPs in Genome-wide and Re-Sequencing Association Studies. PLoS Genetics. 2008;4(7):e1000130. doi: 10.1371/journal.pgen.1000130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu T, Chen Y, Hastie T, Sobel E, Lange K. Genomewide Association Analysis by Lasso Penalized Logistic Regression. Bioinformatics. 2009;25(6):714–721. doi: 10.1093/bioinformatics/btp041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marchini J, Donnelly P, Cardon LR. Genome-wide strategies for detecting multiple loci that influence complex diseases. Nature Genetics. 2005;37(4):413–417. doi: 10.1038/ng1537. [DOI] [PubMed] [Google Scholar]
- Li W, Reich J. A Complete Enumeration and Classification of Two-Locus Disease Models. Human Heredity. 2000;50:334–349. doi: 10.1159/000022939. [DOI] [PubMed] [Google Scholar]
- Velez D, White B, Motsinger A, Bush W, Ritchie M, Williams S, Moore J. A balanced accuracy function for epistasis modeling in imbalanced datasets using multifactor dimensionality reduction. Genetic Epidemiology. 2007;31:306–315. doi: 10.1002/gepi.20211. [DOI] [PubMed] [Google Scholar]
- Culverhouse R, Suarez B, Lin J, Reich T. A perspective on epistasis: limits of models displaying no main effect. Am J Hum Genet. 2002;70:461–471. doi: 10.1086/338759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dudek S, Motsinger A, Velez D, Williams S, Ritchie M. Data simulation software for whole-genome association and other studies in human genetics. Pacific Symposium on Biocomputing. 2006. [PubMed]
- Moore J, White B. Tuning ReliefF for genomewide genetic analysis. Lecture Notes Computer Science. 2007;4447:166–175. full_text. [Google Scholar]
- Yang C, He Z, Wan X, Yang Q, Xue H, Yu W. SNPHarvester: a filtering-based approach for detecting epistatic interactions in genome-wide association studies. Bioinformatics. 2009;25(4):504–511. doi: 10.1093/bioinformatics/btn652. [DOI] [PubMed] [Google Scholar]
- Gregersen PK, Silver J, Winchester RJ. The shared epitope hypothesis: An approach to understanding the molecular genetics of susceptibility to rheumatoid arthritis. Arthritis Rheum. 1987;30:1205–1213. doi: 10.1002/art.1780301102. [DOI] [PubMed] [Google Scholar]
- Mori S, Kou I, Sato H, Emi M, Ito H, Hosoi T, Ikegawa S. Association of genetic variations of genes encoding thrombospondin, type 1, domain-containing 4 and 7A with low bone mineral density in Japanese women with osteoporosis. Journal of Human Genetics. 2008;53(8):694–697. doi: 10.1007/s10038-008-0300-4. [DOI] [PubMed] [Google Scholar]
- Zhao J, Jin L, Xiong M. Test for Interaction between Two Unlinked Loci. Am J Hum Genet. 2006;79(5):831–845. doi: 10.1086/508571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang K, Li M, Bucan M. Pathway-Based Approaches for Analysis of Genomewide Association Studies. Am J Hum Genet. 2007;81:1278–1283. doi: 10.1086/522374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman J, Hastie T, Hofling H, Tibshirani R. Pathwise coordinate optimization. The Annals of Applied Statistics. 2007;1:302–332. doi: 10.1214/07-AOAS131. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The supplementary document provide some backgrounds of our method. It also provides details of our optimization algorithm, and comprehensive experimental results.