

Abstract
During pre-synaptic embryonic development, neuronal processes traverse short distances to reach their targets via growth cone. Over time, neuronal somata are separated from their axon terminals due to skeletal growth of the enlarging organism (Weiss 1941; Gray, Hukkanen et al. 1992). This mechanotransduction induces a secondary mode of neuronal growth capable of accommodating continual elongation of the axon (Bray 1984; Heidemann and Buxbaum 1994; Heidemann, Lamoureux et al. 1995; Pfister, Iwata et al. 2004).
Axon Stretch Growth (ASG) is conceivably a central factor in the maturation of short embryonic processes into the long nerves and white matter tracts characteristic of the adult nervous system. To study ASG in vitro, we engineered bioreactors to apply tension to the short axonal processes of neuronal cultures (Loverde, Ozoka et al. 2011). Here, we detail the methods we use to prepare bioreactors and conduct ASG. First, within each stretching lane of the bioreactor, neurons are plated upon a micro-manipulated towing substrate. Next, neurons regenerate their axonal processes, via growth cone extension, onto a stationary substrate. Finally, stretch growth is performed by towing the plated cell bodies away from the axon terminals adhered to the stationary substrate; recapitulating skeletal growth after growth cone extension.
Previous work has shown that ASG of embryonic rat dorsal root ganglia neurons are capable of unprecedented growth rates up to 10mm/day, reaching lengths of up to 10cm; while concurrently resulting in increased axonal diameters (Smith, Wolf et al. 2001; Pfister, Iwata et al. 2004; Pfister, Bonislawski et al. 2006; Pfister, Iwata et al. 2006; Smith 2009). This is in dramatic contrast to regenerative growth cone extension (in absence of mechanical stimuli) where growth rates average 1mm/day with successful regeneration limited to lengths of less than 3cm (Fu and Gordon 1997; Pfister, Gordon et al. 2011). Accordingly, further study of ASG may help to reveal dysregulated growth mechanisms that limit regeneration in the absence of mechanical stimuli.
Keywords: Bioengineering, Issue 54, tissue engineering, live imaging, axon stretch growth, nerve development, neuron, neuroscience
Protocol
1. Overview of The Axon Stretch Growth Bioreactor System
Two predominant approaches have been utilized to apply experimental forces to neurons. In the first approach, forces are applied to the entire neuron (Lu, Franze et al. 2006; Chetta, Kye et al. 2010; Lindqvist, Liu et al. 2010). In the second approach, forces are applied directly to the axon by pulling on the growth cone. Using glass needles, this latter approach has been used to model formation of new axon (Bernal, Pullarkat et al. 2007; O'Toole, Lamoureux et al. 2008; Lamoureux, Heidemann et al. 2010), to identify force thresholds for elongation and retraction (Dennerll, Lamoureux et al. 1989; Zheng, Lamoureux et al. 1991; Lamoureux, Zheng et al. 1992), and to analyze neurotransmitter clustering in axon terminals (Siechen, Yang et al. 2009). A unique tissue engineering extension of this method was developed to produce large nerve constructs using an automated Axon Stretch Growth (ASG) bioreactor system (Iwata, Browne et al. 2006; Pfister, Iwata et al. 2006). Recently, we developed a miniaturized version of the bioreactor system to study ASG microscopically in real time (Loverde, Ozoka et al. 2011). Here, we detail the 9-day protocol (table 1) we currently use to prepare bioreactors and perform ASG routinely.
Overall Operation: The ASG bioreactor system is comprised of three main components, 1) a bioreactor chamber with independent lanes (elongated wells) where neurons are cultured and stretched, 2) an automated linear motion table to apply stretching forces and 3) a step motor drive controller with software to control stretch growth (figure 1).
Briefly, fabrication of bioreactor prototypes was performed in a machine shop using a vertical milling machine (Bridgeport, Elmira, NY). For biocompatibility, ease of sterilization and durability, the internal bioreactor components were machined from 3/8" polyetheretherketone (PEEK). Transparent polycarbonate was used for lids to allow for light microscopy and viewing of cultures. Corrosion resistant 316 stainless steel screws and hardware complete the assembly (McMaster-Carr, Elmhurst, IL).
The bioreactor chamber is comprised of a stretching frame that forms 3 lanes, an adjustable towing block that manipulates cells across the lanes, and protruding towing rods for external manipulation (figure 2). Neuronal cultures are plated onto disposable Aclar towing culture substrates supported by the towing block (figures 2 & 3). A disposable coverslip stationary substrate attaches to the bottom of the stretching frame and spans all 3 lanes. Prior to stretching, plated neurons must extend axonal processes from the towing substrate onto the stationary substrate via growth cone extension. The population of axons that bridge the substrates will subsequently undergo stretch by controlling displacement of the towing block.
The automated linear motion table consists of a stepper motor (HT23-397, Applied Motion Products, Watsonville, CA) and linear motion table (MIPS-2-10-1.0mm, Servo Systems, Monteville, NJ) mounted to a delrin alignment table. The bioreactor chamber is seated within the table parallel to the linear motion table. The towing rods extending from the bioreactor chamber are fastened to the linear motion table using an adaptor.
The step motor drive controller (Si 2035, Applied Motion Products) is programmed using the included Si programmer software to control manipulation of the towing substrates. Axonal stretch is applied in a stepwise fashion by taking a series of small displacement steps spaced by dwell times (Pfister, Iwata et al. 2004; Pfister, Iwata et al. 2006). This computer-controlled system provides the ability to program customized profiles for ASG, and is essential for continuous experimentation over several days to weeks.
2. Preparation of Bioreactor Chamber
- Prepare the disposable towing and stationary culture substrates for neuronal culture:
- Towing culture substrates are cut from 8.5" x 11" sheets of Aclar film (33C 2.0 mil, Structure Probe, West Chester, PA). Using a sharp blade cut the substrates to approximately 0.5 x 2.5cm, or slightly shorter than the width of the lanes to allow at least 1-2mm clearance on both sides.
- Lightly sand the lower 1/3rd of the towing culture substrates on both sides using fine 1200-grit sandpaper (McMaster Carr). Sanding of the towing substrates facilitates growth of axons from the towing substrates onto the coverslip stationary substrate.
- Cut a 5 x 7cm piece of Aclar or use a No. 1 glass coverslip to serve as the stationary substrate (#4865-1, Brain Research Labs, Newton, MA).
- Clean the culture substrates with a dilute Alconox solution and rinse thoroughly with purified dH2O.
- Sterilize the culture substrates by immersion in 70% ethanol for 30 minutes. Allow the substrates to air dry within a sterile tissue culture hood.
Clean the bioreactor chamber with dilute Alconox and autoclave sterilize within an autoclave container. Immediately after autoclaving, transfer to a sterile hood and allow to air dry.
- Continuing within the sterile hood, glue the culture substrates to the bioreactor using Silicon RTV (Dow Corning #732, McMaster Carr) and sterile cotton tipped swabs (McMaster Carr):
- With the towing block legs in their full upright position, glue the towing culture substrates to the towing block legs at the non-sanded portion. Avoid contact with the sanded culture surface.
- Glue the stationary culture substrate to the bottom of the bioreactor chamber.
- Remove excess glue and air pockets by gently depressing a dry swab against the glued substrates.
- Since Silicone RTV leaches acetic acid that is toxic to neurons, the bioreactor is left to dry under UV light within the hood for 2 full days prior to introducing neuronal cultures.
Lower the towing block legs to achieve a 2-3mm overlap between the tips of the sanded towing substrates and the stationary substrate. Crucial attention must be paid to the size of the overlap, if it is too large, the tips of the towing substrates may deflect off of the stationary substrate; reducing the number of axons that span the overlap (figure 3).
Position the towing block at the starting position, with the towing rods retracted inside the bioreactor. Tighten the immobilization screws to prevent movement of the towing rods prior to stretch growth.
3. Neuronal Cultures
Pool 1mL of 10 μg/mL high molecular weight poly-d-lysine (cat# 354210, BD, Bedford, MA) in serum-free media at the substrate interface area of each lane. Allow the solution to adhere undisturbed for 1 hour at room temperature. Rinse gently 3x with dH2O, followed by a final rinse with culture media. Pipetting should be performed at the back of the lanes, farthest the substrate interface.
Isolate Dorsal Root Ganglia explants (DRGs) from an E16 rat pup. Using a stereomicroscope, dilute the explants into drops using petri dishes. Collect 3-4 explants using a 100 μL pipette and plate onto the edge of the sanded towing culture substrate. Care must be taken to plate the explants at the edge of the towing substrate using a small puddle of media. Up to 1mL of culture media per lane is sufficient to prevent evaporation for several hours while limiting movement of the explants. Media formulation: Neurobasal with B-27 + 0.5mM L-Glutamine (Invitrogen, Carlsbad, CA), 1% FBS-HI (Hyclone, Waltham, MA), 2.5g/L D-Glucose (G-7528, Sigma, St. Louis, MO), 20ng/mL NGF (13290-010, Invitrogen) and 20 μM FdU + 20 μM Uridine mitotic inhibitors (F-0503, U-3003, Sigma).
Attach the lid and transfer the bioreactor to an incubator for 1 hour or until cells adhere.
Fill the bioreactor with culture media at the point farthest the explants to avoid dislodgment. Incubate the bioreactor for a minimum of 5 days while neurons extend axonal processes onto the stationary substrate.
4. Axon Stretch Growth
- The bioreactor undergoes final preparation procedures inside a sterile hood:
- Fill reservoirs within the bioreactor with phosphate buffered saline to humidify the chamber and limit evaporation of culture media. In our system, the hollowed enclosure walls serve as reservoirs. Alternatively, small petri dish lids may be placed on either side of the towing block atop the culture lanes.
- Replace the culture media and fill the lanes to capacity. Pipetting should be performed at the back of the lanes, farthest the substrate interface.
Seat the bioreactor within the automated linear motion table. If the experiment is to be run within an incubator, it must not be humidified due to potential corrosion of the automated linear motion table.
Fasten the towing rod adaptor to the towing rods.
Using Si Programmer software jog the stage of the linear motion table to align with the towing rod adaptor and fasten it to the stage. For convenience, prepare Si Programmer sequences in advance in order to manipulate the stage in specific increments.
Loosen the immobilization screws on the bioreactor chamber to allow free movement of the towing block.
Stretch is applied in a stepwise fashion by initiating a series of small displacement steps spaced by dwell times (Pfister, Iwata et al. 2006). Our paradigm starts by taking 2 μm steps every 172 seconds, resulting in a net 1mm stretch over 24 hours. After one day, the stretch rate can be ramped by increasing the displacement or decreasing the dwell time (table 2).
Once ASG is initiated, media changes are typically not required. Over time, however, culture media generally turns acidic and is evident by a yellowish change in color. If further experimentation is required, old media is never fully drained, but instead only partially changed in order to minimize trauma to floating axons.
5. Representative Results:
Axonal processes can undergo remarkably rapid and robust stretch growth. Initially, the process begins with a period of slow stretching (≤ 1mm/day) that consists of small, infrequent displacements. Within the first 24 hours of stretching, mechanotransduction of neuronal growth pathways occurs, whereby neurons begin addition to the axon cylinder. Within 24 hours of continuous ASG, axons show an increasing tolerance to greater and more frequent displacements. In general, axons can withstand an increase in the stretch rate of 1mm/day every 12-24 hours (Pfister, Iwata et al. 2004; Pfister, Bonislawski et al. 2006; Pfister, Iwata et al. 2006). Increasing the stretch rate too soon, however, may lead to more rapid growth of select axons but will also lead to pathological occlusion that causes disconnection.
Stretch-growing axons have the tendency to form bundles, resembling the architecture of fascicles. Utilizing current protocols, the central, stretch grown portion of axon bundles have no adhesions to the culture substrate. Only the initially adhered, proximal and distal segments of stretch-growing axons remain attached to the culture substrates. Accordingly, the central portion of stretch grown axons float freely, and are sensitive to disruption due to handling.
For a variety of reasons, some axons cannot grow at the applied stretch rate. For example, a DRG neuron with two axonal processes, both of which are undergoing stretch, may not be able to translate sufficient protein and grow at the applied stretch rate. Axons that cannot accommodate the applied stretch will thin following Poisson's effect. Subsequent stretch will lead to occlusion of the axons, inhibiting assembly, leading to pathological disconnection. The majority of axons, however, are able to undergo ASG successfully and only a small percentage of axons undergo this pruning-like process.
 Figure 1. Axon Stretch Growth Bioreactor System. (A) Bioreactor culture chamber & Automated linear motion table, (B) Step motor drive controller and Si Programming software.
Figure 1. Axon Stretch Growth Bioreactor System. (A) Bioreactor culture chamber & Automated linear motion table, (B) Step motor drive controller and Si Programming software.
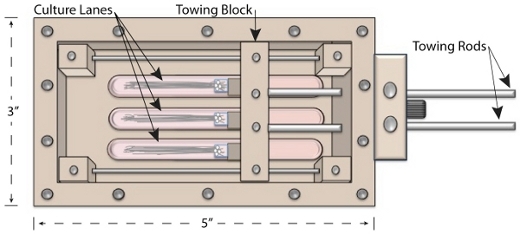 Figure 2. Bioreactor Culture Chamber. This cartoon depicts the bioreactor chamber from the top with the lid removed. The position of the towing block reflects the end stage of axon stretch growth. Stretch grown axons can be seen in bundles within the culture lanes.
Figure 2. Bioreactor Culture Chamber. This cartoon depicts the bioreactor chamber from the top with the lid removed. The position of the towing block reflects the end stage of axon stretch growth. Stretch grown axons can be seen in bundles within the culture lanes.
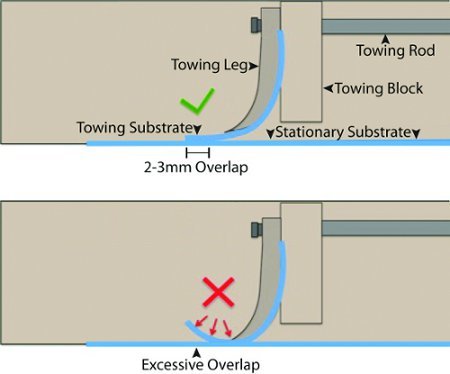 Figure 3. Culture Substrate Plating Interface. This cartoon depicts the components of the towing mechanism within each lane of the bioreactor from side view. (Top) Correct overlap of the towing and stationary culture substrates. (Bottom) Excessive overlap of the towing and stationary substrates causes the tip of the towing substrate to curl.
Figure 3. Culture Substrate Plating Interface. This cartoon depicts the components of the towing mechanism within each lane of the bioreactor from side view. (Top) Correct overlap of the towing and stationary culture substrates. (Bottom) Excessive overlap of the towing and stationary substrates causes the tip of the towing substrate to curl.
Movie 1. Attachment of Culture Substrates to Bioreactor Chamber. Click here to watch video
Movie 2. Plating of DRG Explants onto Towing Substrates. Click here to watch video
Movie 3. SiProgrammer Usage. Click here to watch video
Movie 4. Growth Cone Extension onto Stationary Substrate. Click here to watch video
Movie 5. Axon Stretch Growth. Click here to watch video
| Day | Step |
| 1 | Sterilization & Drying |
| 2 | Gluing & Assembly |
| 4 | Coatings & Neuronal Culture Plating |
| 9 | Stretch Growth Start |
Table 1. Experiment Schedule.
| Time [hr] | Stretch Rate [mm/day] | Dwell Time [s] | Total Length [mm] | Total Stretch Time [days] | |
| Pretension | 24 | 1 | 172.8 | 0 | 1 |
| Stretch | 24 | 1 | 172.8 | 1 | 2 |
| Stretch | 24 | 2 | 86.4 | 3 | 3 |
| Stretch | 24 | 3 | 57.6 | 6 | 4 |
| Stretch | 24 | 4 | 43.2 | 10 | 5 |
| Stretch | 24 | 5 | 34.6 | 15 | 6 |
Table 2. Stretch Rate Schedule. All stretch steps are 2 μm in displacement (10 step motor steps = 2 μm stretch).
Discussion
Two critical steps should be observed during preparation of bioreactors. First, an optimal overlap at the substrate interface is needed to ensure that axons can cross onto the stationary substrate. Aclar that is excessively curled or otherwise imperfect should not be used (figure 3). To optimize the overlap, confirm that the towing substrate is sanded evenly and contacts the stationary substrate uniformly over a 2-3mm long contact patch. The overlap should be optimized prior to each experiment by carefully adjusting the height of the towing legs.
Second, while providing for attachment of neurons, substrate coatings sustain considerable sheering forces caused by displacement of the bioreactor and contractile tension of the axons (Heidemann and Buxbaum 1990; Pfister, Iwata et al. 2004; Loverde, Ozoka et al. 2011). Substrates should be rinsed thoroughly with sterilized water both prior to and after lysine coating. Coatings should be applied from freshly thawed aliquots and spread as evenly as possible. Importantly, the substrates should not be moved or otherwise disturbed during the adhesion period. In subsequent steps, avoid contact with the substrates during plating, and pipet all solutions from the far end of the lanes away from the substrate interface.
Tolerances in the connections of each bioreactor component can manifest in the form of slack. During the initial period of ASG, slack in the system is evident as movement of the automated linear motion table occurs without movement of the towing block. Slack can vary per experiment, but is typically <1mm in our experience. For this reason, a "pre-tension" slack elimination phase of 1mm/day, for one day, precedes the ASG schedule during which the bioreactor parts engage and begin movement of the towing substrates.
Troubleshooting may be required if the displacement step of the automated linear motion table does not match the displacement of the towing block after the pre-tension phase is complete. Asynchronous, inaccurate displacements of the towing block are associated with 'stiction' or static friction within the towing hardware and flexing of the adaptor. To prevent these issues from occurring, movement of the towing block should be checked by hand, after assembly, for smooth near-effortless movement. If binding occurs, the towing assembly should be freed prior to experimentation. Sufficient stiffness of the adaptor is also necessary to assure accurate, synchronous displacements are not overcome by stiction of the towing hardware.
Disclosures
The ex-vivo mechanical elongation of neuronal cells is patented under US Patents #6264944 and #7429267.
Acknowledgments
This work was funded by NSF CAREER CBET-0747615. The authors would like to thank Drs. Douglas H. Smith and David F. Meaney for their mentorship and support.
References
- Bernal R, Pullarkat PA. Mechanical properties of axons. Phys Rev Lett. 2007;99:018301–018301. doi: 10.1103/PhysRevLett.99.018301. [DOI] [PubMed] [Google Scholar]
- Bray D. Axonal growth in response to experimentally applied mechanical tension. Dev Biol. 1984;102:379–389. doi: 10.1016/0012-1606(84)90202-1. [DOI] [PubMed] [Google Scholar]
- Chetta J, Kye C. Cytoskeletal dynamics in response to tensile loading of mammalian axons. Cytoskeleton (Hoboken) 2010;67:650–665. doi: 10.1002/cm.20478. [DOI] [PubMed] [Google Scholar]
- Dennerll TJ, Lamoureux P. The cytomechanics of axonal elongation and retraction. J Cell Biol. 1989;109:3073–3083. doi: 10.1083/jcb.109.6.3073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu SY, Gordon T. The cellular and molecular basis of peripheral nerve regeneration. Mol Neurobiol. 1997;14:1–2. doi: 10.1007/BF02740621. [DOI] [PubMed] [Google Scholar]
- Gray C, Hukkanen M. Rapid neural growth: calcitonin gene-related peptide and substance P- containing nerves attain exceptional growth rates in regenerating deer antler. Neuroscience. 1992;50:953–963. doi: 10.1016/0306-4522(92)90218-q. [DOI] [PubMed] [Google Scholar]
- Heidemann SR, Buxbaum RE. Tension as a regulator and integrator of axonal growth. Cell Motil Cytoskeleton. 1990;17:6–10. doi: 10.1002/cm.970170103. [DOI] [PubMed] [Google Scholar]
- Heidemann SR, Buxbaum RE. Mechanical tension as a regulator of axonal development. Neurotoxicology. 1994;15:95–107. [PubMed] [Google Scholar]
- Heidemann SR, Lamoureux P. Cytomechanics of axonal development. Cell Biochem Biophys. 1995;27:135–155. doi: 10.1007/BF02738107. [DOI] [PubMed] [Google Scholar]
- Iwata A, Browne KD. Long-term survival and outgrowth of mechanically engineered nervous tissue constructs implanted into spinal cord lesions. Tissue Eng. 2006;12:101–110. doi: 10.1089/ten.2006.12.101. [DOI] [PubMed] [Google Scholar]
- Lamoureux P, Heidemann SR. Growth and elongation within and along the axon. Dev Neurobiol. 2010;70:135–149. doi: 10.1002/dneu.20764. [DOI] [PubMed] [Google Scholar]
- Lamoureux P, Zheng J. A cytomechanical investigation of neurite growth on different culture surfaces. J Cell Biol. 1992;118:655–661. doi: 10.1083/jcb.118.3.655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindqvist N, Liu Q. Retinal glial (Muller) cells: sensing and responding to tissue stretch. Invest Ophthalmol Vis Sci. 2010;51:1683–1690. doi: 10.1167/iovs.09-4159. [DOI] [PubMed] [Google Scholar]
- Loverde JR, Ozoka VC. Live Imaging of Axon Stretch Growth in Embryonic and Adult Neurons. J. Neurotrauma. 2011. Forthcoming. [DOI] [PubMed]
- Lu YB, Franze K. Viscoelastic properties of individual glial cells and neurons in the CNS. Proc Natl Acad Sci U S A. 2006;103:17759–17764. doi: 10.1073/pnas.0606150103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Toole M, Lamoureux P. A physical model of axonal elongation: force, viscosity, and adhesions govern the mode of outgrowth. Biophys J. 2008;94:2610–2620. doi: 10.1529/biophysj.107.117424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfister BJ, Bonislawski DP. Stretch-grown axons retain the ability to transmit active electrical signals. FEBS Lett. 2006;580:3525–3531. doi: 10.1016/j.febslet.2006.05.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfister BJ, Gordon T. Biomedical Engineering Strategies for Peripheral Nerve Repair: Surgical Applications, State of the Art, and Future Challenges. Crit Rev Biomed Eng. 2011;39:81–124. doi: 10.1615/critrevbiomedeng.v39.i2.20. [DOI] [PubMed] [Google Scholar]
- Pfister BJ, Iwata A. Extreme stretch growth of integrated axons. J Neurosci. 2004;24:7978–7983. doi: 10.1523/JNEUROSCI.1974-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfister BJ, Iwata A. Development of transplantable nervous tissue constructs comprised of stretch-grown axons. J Neurosci Methods. 2006;153:95–103. doi: 10.1016/j.jneumeth.2005.10.012. [DOI] [PubMed] [Google Scholar]
- Siechen S, Yang S. Mechanical tension contributes to clustering of neurotransmitter vesicles at presynaptic terminals. Proc Natl Acad Sci U S A. 2009;106:12611–12616. doi: 10.1073/pnas.0901867106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith DH. Stretch growth of integrated axon tracts: extremes and exploitations. Prog Neurobiol. 2009;89:231–239. doi: 10.1016/j.pneurobio.2009.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith DH, Wolf JA. A new strategy to produce sustained growth of central nervous system axons: continuous mechanical tension. Tissue Eng. 2001;7:131–139. doi: 10.1089/107632701300062714. [DOI] [PubMed] [Google Scholar]
- Weiss P. Nerve patterns: The mechanics of nerve growth. Growth, Third Growth Symposium. 1941;5:163–203. [Google Scholar]
- Zheng J, Lamoureux P. Tensile regulation of axonal elongation and initiation. J Neurosci. 1991;11:1117–1125. doi: 10.1523/JNEUROSCI.11-04-01117.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]