

Abstract
Cleavage plane orientation has been thought to govern the fate of neural stem cell progeny, but supporting evidence in the neocortex has been sparse. A new study shows that mouse Inscuteable-mediated control of cleavage plane orientation regulates the output of neural progenitor cells.
Evolution of the human neocortex is characterized by enormous increases in neuron number and an associated transformation of a smooth (lissencephalic) cortex, typical of rodents, to a highly folded (gyrencephalic) cortex, typical of primates (Lui et al., 2011). These phenotypes are rooted in proliferative events during embryonic development, when differences in the patterns of division in neural progenitor cells directly influence neuronal output across species. The molecular basis for how these different cell division patterns are established is a critical element in our understanding of neocortical evolution.
Studies over the last decade have defined two major subtypes of neuronal “stem” and progenitor cells in the developing neuroepithelium of the rodent neocortex (Noctor et al., 2004; Kriegstein and Alvarez-Buylla, 2009). Radial glial (RG) cells constitute the major population of neural stem cells and occupy the ventricular zone (VZ). During the peak phase of neurogenesis (around embryonic day 13 to 18 in mice), RG cells predominantly undergo asymmetric division to self-renew while simultaneously giving rise either directly to a neuron, or to an intermediate progenitor (IP) cell. These IP cells (also known as basal progenitors) occupy the subventricular zone (SVZ) and undergo symmetric divisions to amplify neuron number. How the two different modes of RG cell asymmetric division are controlled is not known.
The Drosophila central nervous system has served as a model system for understanding how the polarized distribution of cell fate determinants is coordinated with cleavage plane angle to define the symmetry of division (reviewed by Knoblich, 2008). Cell divisions with a cleavage plane parallel to the epithelium (horizontal) are often asymmetrical, since the polarized determinants are segregated unevenly, whereas those with a cleavage plane orthogonal to the epithelium (vertical) are generally symmetrical because the determinants are evenly partitioned into the daughter cells. A key player in the control of mitotic spindle orientation is Inscuteable (Insc), which segregates to the apical cortex of the dividing neuroblast. Without the presence of Inscuteable, both the position of the mitotic spindle and the distribution of cell fate determinants become randomized (Yu et al., 2006; Knoblich, 2008). It has long been thought that such molecular machinery could be evolutionarily conserved and also control symmetry of division in the neuroepithelium of the mammalian central nervous system (Fishell and Kriegstein, 2003). However, there has yet been no clear picture of the contribution of cleavage plane orientation to cell fate specification in rodents, largely because RG cell division is predominantly horizontal (vertical cleavage plane) during asymmetric division.
In this issue, the Knoblich group present convincing evidence that mouse Inscuteable (mINSC) regulates mitotic spindle orientation, which surprisingly controls the generation of IP cells vs. neurons in the developing mouse neocortex (Postiglione et al, 2011). The authors first demonstrate that mInsc is expressed in the neocortex during mid-neurogenesis and is enriched in the spindle midzone in anaphase progenitor cells. To assess whether or not mInsc is a functional homolog of Drosophila Insc, the authors took an elegant approach and generated transgenic flies expressing mInsc, observing similar localization of mInsc in the Drosophila neuroblast.
The authors next investigated the function of mInsc by generating conditional loss-of-function and gain-of-function mice. mInsc mediates the orientation of retina precursor division (Zigman et al., 2005), but whether this is also true in RG cells has not been clear. Through careful measurements of spindle orientation and the angle of division in RG cells, the authors showed that 63% of the mitotic spindles in control embryos were at angles between 0 and 30 (horizontal) while 33% were between 30 and 60 (oblique). Vertically orientated spindles (between 60 and 90) were rare, representing less than 3% of all the mitotic cells. The authors then evaluated mInsc conditional knockout mice (NesCre/+;mInscfl/fl) and found that the majority of mitotic spindles (95%) were between 0 and 30, with oblique and vertical spindles strongly reduced. Overexpression of mInsc in the conditional knock-in mouse (NesCre/+;R26ki/ki) yielded the opposite phenotype, where oblique and vertical spindles were significantly increased (63%). Therefore, loss of mInsc results in the enrichment of horizontal divisions, whereas overexpression of mInsc randomizes the cleavage plane.
What then are the consequences of changing the mitotic spindle angle of RG cells? Analysis of conditional mInsc knockout mice revealed a decrease in cortical thickness, while conditional mInsc overexpression led to an increase in cortical thickness. These phenotypes were attributed to major changes in the number of neurons, as histological analysis using layer-specific neuronal markers demonstrated a uniform decrease in neurons with mInsc deletion and an increase with mInsc overexpression across all cortical layers (Postiglione et al, 2011).
To link the alterations of neuron production to the progenitor cell subtypes responsible, the authors examined the M phase index and the cell cycle exit index (Q fraction).
Surprisingly, the average cell cycle length and exit rates of neural progenitors did not change in the NesCre/+;mInscfl/fl or the NesCre/+;R26ki/ki mice, indicating that mInsc has little to no general role in regulating the cell cycle. Finally, the authors carefully examined the composition of progenitor cells in the mutants that would lead to the observed changes in neuron number. Remarkably, the number of progenitor cells present outside the ventricular zone, a feature usually associated with IP cells, is increased in mInsc overexpression animals. By performing in utero electroporation of RFP+ plasmids and observing the products of cell division after 24 hours, the authors found that 23% of RFP+ cells in wild type, 10% in mInsc mutants, and more than 50% in mInsc overexpression animals, were Tbr2+ IP cells. This suggested that the changes in orientation of the mitotic spindle caused RG cells to preferentially make IP cells instead of neurons, thereby increasing the transit-amplifying population and neuron number.
This study raises intriguing new questions about neocortical development and permits alternative interpretations for the phenotype of the reported mInsc mutant mice. Although the observed increase in non-ventricular progenitor cells in mInsc overexpression animals is most obviously due to increases in IP cell number, aberrant non-ventricular progenitors also included those that express Pax6, a feature usually associated with RG cells. Future studies characterizing the morphology and behavior of these non-ventricular progenitor cells will help delineate whether these Pax6 and Tbr2 expressing cells are the same or different cell types. This question will be important to resolve, as the abundance of non-ventricular Pax6+ progenitor cells has recently been shown to be predictive of neocortical size across species and suggested to be important in neocortical evolution (Lui et al., 2011). Analyses of the developing neocortex in humans, ferrets, and mice (Hansen et al., 2010; Fietz et al., 2010; Reillo et al., 2010; Shitamukai et al., 2011; Wang et al., 2011) have defined a new class of neural stem cells known as oRG cells, which function as a non-ventricular counterpart to RG cells and serve to further expand neuron number. Furthermore, an elegant study by Shitamukai et al. showed that removal of LGN in the mouse, which induces oblique cleavage planes in RG cells, results in the generation of non-ventricular progenitors resembling oRG cells. Because oRG cells are also thought to generate IP cells and neurons, we suggest the intriguing possibility that randomization of cleavage plane in mInsc overexpression mutants could also have the same effect, where an oblique or horizontal division results in an oRG cell, which further proliferates to generate IP cells away from the ventricle (Figure 1).
Figure 1.
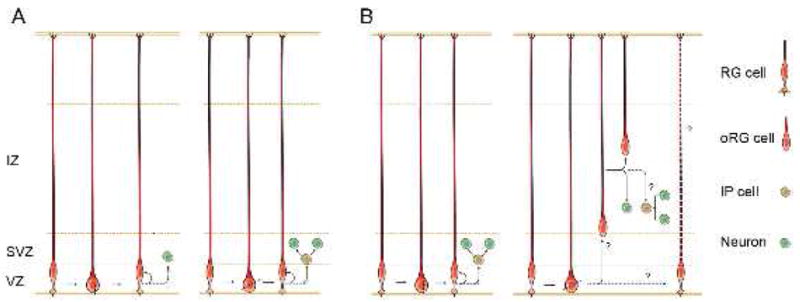
Radial glial cell divisions mediated by different cleavage plane orientations.
(A) Postiglione et al propose that radial glial cells (RG) that divide with a vertical cleavage plane generate a neuron and self-renew (left panel). Overexpression of Inscuteable randomizes the cleavage plane angle, which induces a greater proportion of oblique RG cell divisions. The authors observe that the number of IP cells is increased under this mode of division, and suggest that oblique cleavages preferentially produce IP cells (right panel).
(B) An alternative interpretation is that vertical divisions produce IP cells (left panel), and oblique divisions produce oRG cells (right panel)(Konno et al., 2008; Shitamukai et al., 2011). The oRG cells may function as non-ventricular stem cells that also produce IP cells or neurons (Hansen et al., 2010; Wang et al., 2011).
Interestingly, although both LGN and mInsc control cleavage plane orientation, their mutant phenotypes are not the same. Loss of LGN induces oblique divisions and drives the formation of non-ventricular RG cells, but does not drastically affect the rates of neuronal production (Konno et al., 2008; Shitamukai et al., 2011). Overexpression of mInsc also induces oblique divisions and results in a non-ventricular progenitor population. However, neuronal production is massively increased in this case, suggesting that mInsc may also be involved in controlling proliferative capacity. These differential effects highlight the importance of understanding the molecular consequences of cell divisions with a given cleavage plane orientation. It is worth noting that so far there is little data in the developing mammalian cortex concerning the relationship between spindle orientation and the cytoplasmic distribution of cell fate determinants in dividing cells. Future studies will be needed to show how the orientation of cell divisions relates to the distribution of cell fate determinants, and whether these factors are related to cell cycle length and cell fate choice. We anticipate that further work in this field will continue to shed light on the intricate mechanisms of neural progenitor cell division.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Fietz SA, Kelava I, Vogt J, Wilsch-Brauninger M, Stenzel D, Fish JL, Corbeil D, Riehn A, Distler W, Nitsch R, et al. OSVZ progenitors of human and ferret neocortex are epithelial-like and expand by integrin signaling. Nat Neurosci. 2010;13:690–699. doi: 10.1038/nn.2553. [DOI] [PubMed] [Google Scholar]
- Fishell G, Kriegstein AR. Neurons from radial glia: the consequences of asymmetric inheritance. Curr Opin Neurobiol. 2003;13:34–41. doi: 10.1016/s0959-4388(03)00013-8. [DOI] [PubMed] [Google Scholar]
- Hansen DV, Lui JH, Parker PR, Kriegstein AR. Neurogenic radial glia in the outer subventricular zone of human neocortex. Nature. 2010;464:554–561. doi: 10.1038/nature08845. [DOI] [PubMed] [Google Scholar]
- Knoblich JA. Mechanisms of asymmetric stem cell division. Cell. 2008;132:583–597. doi: 10.1016/j.cell.2008.02.007. [DOI] [PubMed] [Google Scholar]
- Konno D, Shioi G, Shitamukai A, Mori A, Kiyonari H, Miyata T, Matsuzaki F. Neuroepithelial progenitors undergo LGN-dependent planar divisions to maintain self-renewability during mammalian neurogenesis. Nat Cell Biol. 2008;10:93–101. doi: 10.1038/ncb1673. [DOI] [PubMed] [Google Scholar]
- Kriegstein A, Alvarez-Buylla A. The glial nature of embryonic and adult neural stem cells. Annu Rev Neurosci. 2009;32:149–184. doi: 10.1146/annurev.neuro.051508.135600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lui JH, Hansen DV, Kriegstein AR. Development and evolution of the human neocortex. Cell. 2011;146:18–36. doi: 10.1016/j.cell.2011.06.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noctor SC, Martinez-Cerdeno V, Ivic L, Kriegstein AR. Cortical neurons arise in symmetric and asymmetric division zones and migrate through specific phases. Nat Neurosci. 2004;7:136–144. doi: 10.1038/nn1172. [DOI] [PubMed] [Google Scholar]
- Postiglione, Knoblich, et al. Neuron. 2011;(this issue) [Google Scholar]
- Reillo I, de Juan Romero C, Garcia-Cabezas MA, Borrell V. A Role for Intermediate Radial Glia in the Tangential Expansion of the Mammalian Cerebral Cortex. Cereb Cortex. 2010 doi: 10.1093/cercor/bhq238. [DOI] [PubMed] [Google Scholar]
- Shitamukai A, Konno D, Matsuzaki F. Oblique Radial Glial Divisions in the Developing Mouse Neocortex Induce Self-Renewing Progenitors outside the Germinal Zone That Resemble Primate Outer Subventricular Zone Progenitors. J Neurosci. 2011;31:3683–3695. doi: 10.1523/JNEUROSCI.4773-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X, Tsai JW, Lamonica B, Kriegstein AR. A new subtype of progenitor cell in the mouse embryonic neocortex. Nat Neurosci. 2011;14:555–561. doi: 10.1038/nn.2807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu F, Kuo CT, Jan YN. Drosophila neuroblast asymmetric cell division: recent advances and implications for stem cell biology. Neuron. 2006;51:13–20. doi: 10.1016/j.neuron.2006.06.016. [DOI] [PubMed] [Google Scholar]
- Zigman M, Cayouette M, Charalambous C, Schleiffer A, Hoeller O, Dunican D, McCudden CR, Firnberg N, Barres BA, Siderovski DP, et al. Mammalian inscuteable regulates spindle orientation and cell fate in the developing retina. Neuron. 2005;48:539–545. doi: 10.1016/j.neuron.2005.09.030. [DOI] [PubMed] [Google Scholar]