

I. HIV-1 Drug Resistance
A. Introduction
HIV-1 RT and protease sequencing and drug susceptibility testing have been done in research settings for more than ten years to elucidate the genetic mechanisms of resistance to antiretroviral drugs. Retrospective studies have shown that the presence of drug resistance before starting a new drug regimen is an independent predictor of virologic response to that regimen (DeGruttola et al., 2000; Hanna and D’Aquila, 2001; Haubrich and Demeter, 2001). Prospective studies have shown that patients whose physicians have access to drug resistance data, particularly genotypic resistance data, respond better to therapy than control patients whose physicians do not have access to the same data (Baxter et al., 2000; Cohen et al., 2000; De Luca et al., 2001; Durant et al., 1999; Melnick et al., 2000; Meynard et al., 2000; Tural et al., 2000). The accumulation of retrospective and prospective data has led three expert panels to recommend the use of resistance testing in the treatment of HIV-infected patients (EuroGuidelines Group for HIV Resistance, 2001; Hirsch et al., 2000; US Department of Health and Human Services Panel on Clinical Practices for Treatment of HIV Infection, 2000) (Table 1).
Table 1.
Expert Panel Recommendations on HIV Drug Resistance Testing
| Recommendations (EuroGuidelines Group for HIV Resistance, 2001; Hirsch et al., 2000; US Department of Health and Human Services Panel on Clinical Practices for Treatment of HIV Infection, 2000) | |
|---|---|
| Primary HIV-1 infection | The DHHS and IAS-USA state that resistance testing should be considered. The more recent EuroGuidelines state that testing should be strongly considered, reflecting the increasing rates of primary HIV-1 drug resistance. |
| Established HIV-1 infection | Not generally recommended by any of the guidelines. Detection of drug resistance is inversely proportional to the time since infection because rates of resistance were lower in the past and because resistant strains tend to be overgrown by susceptible strains that were either co-transmitted at the time of infection or that resulted from back mutations. |
| First regimen failure | Resistance testing is recommended by all three panels. Patients with virologic failure while receiving drug combinations have virus isolates that are not necessarily resistant to all of the drugs in the combination and because HIV-1 may develop drug resistance by more than one mechanism and each mechanism may have different consequences for cross-resistance. |
| Suboptimal viral suppression after initiation of HAART | Recommended by DHHS. Not specifically recommended by the IAS-USA and EuroGuidelines committees but probably falls under the “First regimen failure” category. |
| Multiple regimen failures | Recommend testing to optimize the number of active drugs in the next regimen; exclude drugs to which response is unlikely. |
| Pregnancy | Recommend testing to optimize maternal treatment and prophylaxis for neonate. |
| Post-exposure prophylaxis | Addressed by EuroGuidelines which recommends testing but cautions that treatment should not be delayed while waiting for the test result. Rather, the results of the test should be used to modify the treatment. |
There have been several recent reviews on methods for assessing HIV-1 drug resistance (Demeter and Haubrich, 2001; Hanna and D’Aquila, 2001; Richman, 2000) and on the mutations associated with drug resistance (Deeks, 2001; Hammond et al., 1999; Loveday, 2001; Miller, 2001; Shafer et al., 2000b). This review will detail the use of HIV-1 genotypic resistance testing in research settings where it is used to learn about the mechanisms and clinical significance of drug resistance and in clinical settings where it is used to help guide anti-HIV treatment.
B. Evolution of HIV-1 drug resistance
The evolution of HIV-1 drug resistance within an individual depends on the generation of genetic variation in the virus and on the selection of drug-resistant variants during therapy. HIV-1 genetic variability is a result of the inability of HIV-1 RT to proofread nucleotide sequences during replication (Mansky, 1998). It is exacerbated by the high rate of HIV-1 replication in vivo, the accumulation of proviral variants during the course of HIV-1 infection, and genetic recombination when viruses with different sequences infect the same cell. As a result, innumerable genetically distinct variants (quasispecies) evolve in individuals in the months following primary infection (Coffin, 1995).
The HIV-1 quasispecies in an individual undergoes continuous genetic variation, competition, and selection. Development of drug resistance depends on the size and heterogeneity of the HIV-1 population within an individual, the extent to which virus replication continues during drug therapy, the ease of acquisition of a particular mutation (or set of mutations), and the effect of drug-resistance mutations on drug susceptibility and virus fitness. Some mutations selected during drug therapy confer measurable phenotypic resistance by themselves, whereas other mutations increase resistance when present with other mutations or compensate for the diminished replicative activity that can be associated with drug resistance.
It has been estimated that every possible single point mutation occurs between 104 and 105 times per day in an untreated HIV-1-infected individual and that double mutants also occur commonly (Coffin, 1995). It is not known, however, whether multidrug-resistant viruses already exist at low frequencies in untreated persons or if they are generated by residual viral replication during therapy (Ribeiro and Bonhoeffer, 2000). Answers to this question depend on the effective population number of HIV-1 in vivo. Some authors have argued in favor of a high effective population number and a deterministic model of HIV-1 evolution (Rouzine and Coffin, 1999); others have argued in favor of a lower effective population number and a stochastic model of HIV-1 evolution (Brown, 1997; Brown and Richman, 1997; Frost et al., 2000).
Although HIV-1 drug resistance is usually acquired during anti-HIV drug therapy, drug resistance can also be transmitted between individuals. In the United States and Europe about 10% of new infections are with HIV-1 isolates harboring resistance to at least one of three classes of anti-HIV drugs (Balotta et al., 2000; Boden et al., 1999; Briones et al., 2001; Duwe et al., 2001; Grant et al., 1999; Harzic et al., 1999; Little et al., 1999; Salomon et al., 2000; Simon et al., 2001; Tamalet et al., 2000; Yerly et al., 1999). Recent studies suggest that transmitted HIV-1 drug resistance is gradually increasing (Little, 2000; UK Collaborative Group on Monitoring the Transmission of HIV Drug Resistance, 2001).
C. HIV-1 protease
The HIV-1 protease enzyme is responsible for the post-translational processing of the viral Gag and Gag-Pol polyproteins to yield the structural proteins and enzymes of the virus. The enzyme is an aspartic protease composed of two non-covalently associated structurally identical monomers 99 amino acids in length (Figure 1). Its active site resembles that of other aspartic proteases and contains the conserved triad, Asp-Thr-Gly, at positions 25–27. The hydrophobic substrate cleft recognizes and cleaves 9 different sequences to produce the matrix, capsid, nucleocapsid, and p6 proteins from the Gag polyprotein and the protease, RT, and integrase proteins from the Gag-Pol polyprotein (Erickson et al., 1999; Miller 2001). The enzyme contains a flexible flap region that closes down on the active site upon substrate binding.
Figure 1.

Structural model of HIV-1 protease homodimer labeled with protease inhibitor resistance mutations. The polypeptide backbone of both protease subunits (positions 1–99) is shown. The active site (positions 25–27) is displayed in ball and stick mode. The protease was co-crystallized with a protease inhibitor, which is displayed in space-fill mode.
There are six FDA-approved protease inhibitors (PIs): amprenavir, indinavir, lopinavir (manufactured in combination with ritonavir), nelfinavir, ritonavir, and saquinavir. Mutations associated with PI resistance are found at more than 20 different residues of the enzyme (Table 2) (Figure 1). Resistance is mediated by structural changes that reduce binding affinity between the inhibitor and the mutant protease molecule. The effects of non-active site mutations are less obvious and appear to involve other mechanisms: alterations in enzyme catalysis, effects on dimer stability, alterations in inhibitor binding kinetics, or active site re-shaping through long-range structural perturbations (Erickson et al., 1999; Miller 2001). The three-dimensional structures of wildtype HIV-1 protease and of several drug-resistant mutant forms bound to various inhibitors have been determined by crystallography (Baldwin et al., 1995; Chen et al., 1995; Mahalingam et al., 1999).
Table 2.
HIV-1 Protease Inhibitor (PI) Drug Resistance Mutations
| Codon | Mechanism | Effect on resistance | References |
|---|---|---|---|
| 8 | Substrate cleft | R8Q/K confers high-level resistance to some of the earliest PIs. It occurs extremely rarely and its effect on current PIs is not known. | Gulnik et al., 1995; Ho et al., 1994 |
| 10 | Accessory (polymorphic) | L10I/F/V/R are associated with resistance to all PIs when present with other mutations. | Hertogs et al., 2000b; Para et al., 2000; Shafer et al., 1999b; Zolopa et al., 1999b |
| 20 | Accessory (polymorphic) | K20R/M/I are associated with resistance to IDV, RTV, LPV, and possibly other PIs when present with other mutations. Variants at this position occur commonly in several non-B subtypes | Hertogs et al., 2000b; Shafer et al., 1999b, Condra et al. 1996, Molla et al., 1996 |
| 24 | Accessory | L24I is associated with IDV and LPV resistance when present with other mutations. | Condra et al., 1996; Kempf et al., 2000 |
| 30 | Substrate cleft | D30N causes resistance to NFV. D30N is perhaps the only protease mutations that does not confer cross-resistance to multiple PIs. | Patick et al., 1998; Patick et al., 1996; Zolopa et al., 1999b |
| 32 | Substrate cleft | V32I is a substrate cleft mutation that confers resistance to IDV, RTV, and APV. Although it is in the substrate cleft, this mutation has a minimal effect on drug resistance. | Condra et al., 1996; Snowden et al., 2000 |
| 36 | Accessory (polymorphic) | M36I/V are associated with resistance to all PIs when present with other mutations. Variants at this position occur commonly in non-B subtypes. | Hertogs et al., 2000b; Shafer et al., 1999b |
| 46 | Enzyme flap | M46I increases resistance to IDV, RTV, NFV, APV, and LPV when present with other mutations. | Condra et al., 1996; Partaledis et al., 1995; Patick et al., 1996; Zolopa et al., 1999b |
| 47 | Enzyme flap | I47V increases resistance to APV when present with I50V. Its effect on other PIs has not been well-characterized. | Partaledis et al., 1995 |
| 48 | Substrate cleft | G48V causes resistance to SQV. G48V also confers limited cross-resistance to NFV, IDV, and RTV. Its effect on APV and LPV is not known. |
Jacobsen et al., 1995 Winters et al., 1998a |
| 50 | Substrate cleft | I50V causes resistance to APV. I50V also contributes resistance to RTV and LPV. Occurs primarily in patients receiving APV as their first PI. | Partaledis et al., 1995; Snowden et al., 2000; Xu et al., 2001; Parkin et al., 2001; Prado et al., 2001 |
| 53 | Enzyme flap | F53L is a substrate cleft mutation that occurs only in isolates from treated patients. It is associated with resistance to IDV, RTV, LPV, SQV, and possibly NFV and APV. | Kempf et al., 2000; Shafer, et al., 1999b; Gulnik et al., 1995 |
| 54 | Enzyme flap | I54V/L/T increase resistance to each of the PIs when present with other mutations. I54M occurs in patients receiving APV. | Hertogs et al., 2000b; Kempf et al., 2000; Snowden et al., 2000; Zolopa et al., 1999b; Condra et al., 1996 |
| 63 | Accessory (polymorphic) | L63P/A/Q/S/H/C/T/I occur commonly in untreated persons but are also associated with resistance to PIs when present with other mutations. |
Hertogs et al., 2000b; Shafer et al., 1999b, Condra et al. 1996 Yahiet al. 1999 |
| 71 | Accessory (polymorphic) | A71V/T is associated with resistance to IDV, RTV, SQV, NFV, LPV and probably APV when present with other mutations. | Hertogs et al., 2000b; Shafer et al., 1999b; Zolopa et al., 1999b, Condra et al., 1996, Molla et al., 1996 |
| 73 | Accessory | G73S/T/C are associated with resistance to IDV, SQV, NFV and possibly the remaining PIs when present with other mutations. | Hertogs et al., 2000b; Shafer et al., 1999b, Dulioust et al., 1998, Zolopa et al., 2001 |
| 77 | Accessory (polymorphic) | V77I is a common polymorphism that is associated with drug resistance in isolates containing other mutations. | Hertogs et al., 2000b; Shafer et al., 1999b, Patick et al., 1998 |
| 82 | Substrate cleft | V82A/T/F/S cause resistance to IDV, RTV, and LPV. When present with other mutations, these mutations contribute resistance to NFV, APV, SQV. V82I is a polymorphism that does not appear to be associated with drug resistance. | Condra et al., 1996; Falloon et al., 2000; Kempf et al., 2000; King et al., 1995; Molla et al., 1996; Shafer et al., 1998; Sham et al., 1998 |
| 84 | Substrate cleft | I84V contributes resistance to each of the PIs. | Condra et al., 1996; Hertogs et al., 2000b; Kempf et al., 2000; Molla et al., 1996; Patick et al., 1996; Snowden et al., 2000 |
| 88 | Accessory | N88D/S/T increase NFV resistance particularly when present with D30N or M46I. These mutations may cause low-level cross-resistance to IDV and RTV. N88S causes hyper-susceptibility to APV. | Patick et al., 1998; Ziermann et al., 2000 |
| 90 | Impacts on substrate cleft | L90M causes resistance to SQV and NFV. When present with other mutations it contributes resistance to IDV, RTV, APV, and LPV. | Condra et al., 1996; Jacobsen et al., 1995; Molla et al., 1996; Para et al., 2000 |
| 93 | Accessory (polymorphic) | I93L is a common polymorphism that is associated with drug resistance in isolates containing other mutations. | Shafer et al., 1999b, Molla et al., 1996, Patick et al., 1998 |
APV: amprenavir
IDV: indinavir
LPV: lopinavir
NFV: nelfinavir
RTV: ritonavir
SQV: saquinavir.
Sequence analysis of drug resistance clones has shown that mutations at several of the protease cleavage sites also contribute to drug resistance (Cote et al., 2001; Doyon et al., 1996; Mammano et al., 1998; Zhang et al., 1997) (Table 3). Growth kinetic studies have shown that the cleavage site mutations in some circumstances improve the kinetics of protease enzymes containing drug-resistance mutations and that these mutations appear to be compensatory rather than primary. There have been no reports that changes at cleavage sites alone can cause PI resistance.
Table 3.
HIV-1 Protease Cleavage Sites
| Site | AA | Position | References |
|---|---|---|---|
| gag | |||
| MA/CA | SQNY/PIV | 1187–1188 | Mammano et al., 1998, Cote et al., 2001 |
| CA/p2 | ARVL/AEA | 1880–1881 | Mammano et al., 1998, Cote et al., 2001 |
| p2/NC | ATIM/MQR | 1920–1921 | Mammano et al., 1998), Cote et al., 2001 |
| NC/p1 | RQAN/FLG | 2085–2086 | Mammano et al., 1998), Cote et al., 2001, Zhang et al., 1997, Bally et al., 2000 |
| p1/p6 | PGNF/LQS | 2136–2137 | Mammano et al., 1998, Cote et al., 2001, Bally et al., 2000, Doyon et al., 1996 |
| pol | |||
| TF/PRSF | SV/PQI | 2257–2258 | Cote et al., 2001 |
| PR/RT | TLNF/PIS | 2552–2553 | Cote et al., 2001 |
| RT (p51/p66) | AETF/YVD | 3869–3870 | Cote et al., 2001 |
| RT/IN | RKVL/FLD | 4232–4233 | Cote et al., 2001 |
MA - Matrix
CA - Capsid
NC - Nucleocapsid
TF - Transframe
PR- Protease
RT- Reverse Transcriptase
IN - Integrase.
Scissile bonds are indicated by the slashes in the amino acid sequence.
Nucleic acid positions in relation to HXB2 (GenBank accession No. K03455) are indicated in the Position column.
D. HIV-1 reverse transcriptase (RT)
The RT enzyme is responsible for RNA-dependent DNA polymerization and DNA-dependent DNA polymerization. RT is a heterodimer consisting of p66 and p51 subunits (Figure 2). The p51 subunit is composed of the first 450 amino acids of the RT gene. The p66 subunit is composed of all 560 amino acids encoded by the RT gene. Although the p51 and p66 subunits share 450 amino acids, their relative arrangements are significantly different. The p66 subunit contains the DNA-binding groove and the active site; the p51 subunit displays no enzymatic activity and functions as a scaffold for the enzymatically active p66 subunit. The p66 subunit has five subdomains including the “fingers”, “palm”, and “thumb” subdomains which participate in polymerization, and the “connection” and “RNase H” subdomains (Huang et al., 1998; Kohlstaedt et al., 1992).
Figure 2.
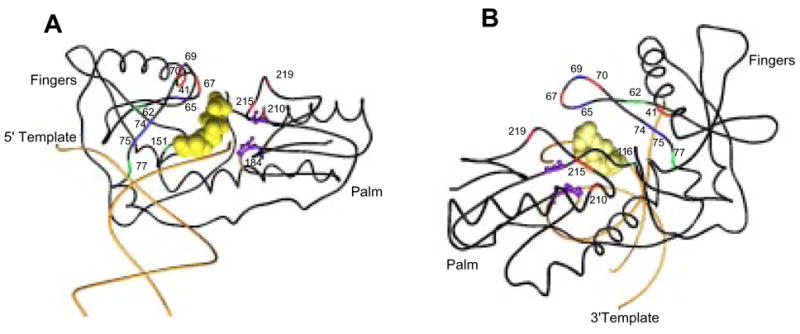
Structural model of HIV-1 reverse transcriptase (RT) labeled with nucleoside RT inhibitor (NRTI) resistance mutations. The polypeptide backbone of the fingers and palm domain (positions 1–235), and DNA primer and template strands are shown. The active site positions (110, 185, 186) are displayed in ball and stick mode. The incoming nucleotide is displayed in space-fill mode. These drawings are based on the structure published by Huang, et al., 1998 and are shown in “front” (a) and “back” (b) views.
The nucleoside RT inhibitors (NRTIs) are prodrugs that are triphosphorylated by host cellular enzymes. The triphosphorylated NRTIs then compete with natural deoxynucleoside triphosphates (dNTPs) for incorporation into the newly synthesized DNA chains where they cause chain termination. There are two biochemical mechanisms of NRTI drug resistance. The first mechanism is mediated by mutations that allow the RT enzyme to discriminate against NRTI during synthesis, thereby preventing their addition to the primer DNA chain (Larder and Stammers, 1999; Sarafianos et al., 1999a; Sarafianos et al., 1999b; Huang et al., 1998). The second mechanism is mediated by mutations in RT that increase the rate of hydrolytic removal of the chain terminating NRTI and enable continued DNA synthesis (Arion et al., 1998; Arion et al., 2000; Boyer et al., 2001; Meyer et al., 1999; Meyer et al., 1998).
The non-nucleoside RT inhibitors (NNRTIs) bind to a hydrophobic pocket in the RT enzyme close to, but not contiguous with, the active site. These compounds inhibit HIV-1 replication allosterically by displacing the catalytic aspartate residues relative to the polymerase binding site (Esnouf et al., 1995; Kohlstaedt and Steitz, 1992; Spence et al., 1995). The mutations responsible for NNRTI resistance are in the hydrophobic pocket which bind the inhibitors. A single mutation in this pocket may result in high-level resistance to one or more NNRTIs. Resistance usually emerges rapidly when NNRTI are administered as monotherapy or in the presence of incomplete virus suppression suggesting that resistance is caused by the selection of a pre-existing population of mutant viruses within an individual (Havlir et al., 1996; Jackson et al., 2000; Wei et al., 1995).
Most NRTI and NNRTI drug resistance mutations are in the 5′ polymerase coding regions, particularly in the “fingers” and “palm” subdomains (Figures 2 and 3) (Tables 4 and 5) (Larder and Stammers, 1999; Sarafianos et al., 1999b). The earliest three-dimensional structures of HIV-1 RT showed the enzyme bound to an NNRTI (Kohlstaed et al., 1992) and bound to a double stranded DNA molecule (Ding et al., 1998; Jacobo-Molina et al., 1993). In 1998, a new structure showed the interaction between the catalytic complex and the incoming dNTP (Huang et al., 1998). Structures of mutant enzymes have also been determined (Ren et al., 1998; Sarafianos et al., 1999a, Stammers et al., 2001).
Figure 3.

Structural model of HIV-1 reverse transcriptase (RT) labeled with non-nucleoside RT inhibitor (NNRTI) resistance mutations. The polypeptide backbone of the complete p66 subunit (positions 1–560), and DNA primer and template strands are shown. This drawing is based on the structure provided by Kohlstaedt and Steitz, 1992 in which the RT is co-crystallized with nevirapine, which is displayed in space-fill mode. The positions associated with NNRTI resistance are shown surrounding the hydrophobic pocket to which nevirapine and other NNRTIs bind.
Table 4.
HIV-1 Nucleoside RT Inhibitor (NRTI) Drug-Resistance Mutations
| Codon | Mechanism | Effect on Resistance1 | References |
|---|---|---|---|
| 41 | Classical AZT2 | M41L increases AZT resistance when present with T215Y/F. | Kellam et al., 1992; Larder et al., 1994 |
| 44 | Accessory5 | E44A/D occurs with increased frequency in patients receiving multiple NRTI. It has recently been shown to cause low-level 3TC resistance when present with V118I. | Delaugerre et al., 2001; Hertogs et al., 2000a |
| 62 | MNR4 | A62V is associated with multinucleoside resistance caused by Q151M. | Iversen et al., 1996; Shirasaka et al., 1995b |
| 65 | β2-β3 loop region3 | K65R causes high-level resistance to DDC and low-to-intermediate levels of resistance to ddI, ABC, and 3TC. | Gu et al., 1994a; Gu et al., 1994b; Tisdale et al., 1997; Zhang et al., 1994 |
| 67 | Classical AZT,2 β2-β3 loop region 3 | D67N contributes to AZT resistance usually with mutations at codons 70 or 215. D67E/G occurs in heavily treated patients. | Larder and Kemp, 1989 |
| 69 | β2-β3 loop region3 | T69D/N/A cause ddC and ddI resistance and may cause low-level D4T resistance particularly when present in isolates with classical AZT resistance mutations. Insertions at this codon are by themselves associated with low level resistance to each of the NRTI. Together with AZT resistance mutations, insertions are associated with moderate-to-high levels of resistance to AZT, ddI, ddC, d4T, and 3TC. | Bloor et al., 1998; Fitzgibbon et al., 1992; Hertogs et al., 1998; Winters et al., 1998; Tamalet et al., 1998; Winters et al., 2001 |
| 70 | Classical AZT,2 β2-β3 loop region3 | K70R causes AZT resistance. | Larder and Kemp, 1989; de Jong et al., 1996, Shulman et al., 2001 |
| 74 | β2-β3 loop region3 | L74V causes ddI, ddC, and ABC resistance. L74V partially suppresses T215Y-mediated AZT resistance. | St. Clair et al., 1991; Tisdale et al., 1997; Kozal et al., 1994a |
| 75 | MNR4 | V75T/M/A causes d4T resistance and may cause low-level ddI and ddC resistance. V75I increases multinucleoside resistance caused by Q151M when present with F77L and F116Y. |
Lacey and Larder, 1994 Bloor et al., 1998; Iversen, et al., 1996; Shirasaka et al., 1995b |
| 77 | MNR4 | F77L increases multinucleoside resistance caused by Q151M when present with V75I or F116Y. | Iversen et al., 1996; Shirasaka et al., 1995b |
| 115 | Accessory5 | Y115F causes low-level resistance to ABC. | Tisdale et al., 1997 |
| 116 | MNR4 | F116Y increases multinucleoside resistance caused by Q151M when present with F77L or V75I. | Iversen et al., 1996; Shirasaka et al., 1995b |
| 118 | Accessory5 | V118I occurs with increased frequency in patients receiving multiple NRTI. It has recently been shown to cause intermediate 3TC resistance when present with E44A/D. | Delaugerre et al., 2001; Hertogs et al., 2000a |
| 151 | MNR4 | Q151M causes intermediate levels of resistance to AZT, ddI, ddC, d4T, and ABC. Q151M, together with its associated changes at codons 62, 75, 77, and 116 causes high-level resistance to these NRTI and low-level resistance to 3TC. | Iversen et al., 1996; Schmit et al., 1998; Shafer et al., 1994; Shirasaka et al., 1995b; Van Laethem et al., 2000 |
| 184 | Close to active site3 | M184V/I cause high-level 3TC resistance and low-level ddI, ddC, and ABC resistance. M184V/I partially suppresses T215Y-mediated AZT resistance. | Boucher et al., 1993; Gu et al., 1992; Larder et al., 1995; Schuurman et al., 1995; Tisdale et al., 1997; Tisdale et al., 1993 |
| 210 | Classical AZT2 | L210W increases AZT resistance when present with mutations at position 215. | Harrigan et al., 1996; Hooker et al., 1996 |
| 215 | Classical AZT2 | T215Y/F causes AZT resistance and also limits the effectiveness of d4T, ABC, ddI, and ddC. T215S/C/D represent transitions between T and Y or F. | Kozal et al., 1993; Larder, et al., 1991; Larder and Kemp, 1989; Rey et al., 1998; Yerly et al., 1998; Japour et al., 1998; Lanier et al., 1998; Izopet et al., 1998 |
| 219 | Classical AZT2 | K219Q/E increase AZT resistance when present with K70R or T215Y/F. K219N/R occur commonly in heavily NRTI-treated patients. | Larder and Kemp, 1989, Larder et al., 1991 |
3TC: lamivudine, ABC: abacavir, AZT: zidovudine, d4T: stavudine, ddI: didanosine, ddC: zalcitibine.
Classical AZT resistance mutations: Various combinations of these mutations have been shown to mediate ATP and pyrophosphate (PP)-dependent hydrolytic removal (pyrophosphorolysis) of zidovudine monophosphate from a terminated cDNA chain (Arion et al., 1998; Boyer et al., 2001; Meyer et al., 1999; Meyer et al., 1998; Meyer et al., 2000b), to cause a compensatory increase in RT processivity (Arion et al., 1998; Arts et al., 1998; Caliendo et al., 1996), and to confer cross-resistance to d4T, ABC, and limit the effectiveness of ddI and ddC (Coakley et al., 2000; Harrigan et al., 2000; Holodniy et al., 1996; Izopet et al., 1999; Japour et al., 1995; Lanier et al., 1999; Mayers et al., 1999; Montaner et al., 2000; Pellegrin et al., 1999; Shulman et al., 2001).
Mutations at position 184 and mutations in the β2-β3 loop region cause resistance by decreasing affinity of RT for the nucleoside analog. Several of these mutations, including M184V and L74V, interfere with the activity of the classical AZT resistance mutations.
MNR or multinucleoside resistance mutations: Q151M is the primary mutation. Mutations at positions 62, 75, 77, and 116 are secondary. Q151 possibly interacts directly with the 3′-OH of the incoming ddNTP (Sarafianos et al., 1999b).
Accessory: The combination of mutations at positions 44 and 118 have been shown to confer low-level 3TC resistance (Hertogs et al., 2000a). But the increasing prevalence of these mutations in isolates from heavily treated patients that also have classical AZT resistance mutations, suggests a broader role (Delaugerre et al., 2001). G333E is a polymorphism that facilitates AZT resistance in isolates with M184V and multiple classical AZT resistance mutations.
Table 5.
HIV-1 Non-Nucleoside RT Inhibitor Drug-Resistance Mutations
| Codon | Effect on Resistance | References |
|---|---|---|
| 98 | A98G is associated with low-level resistance to each of the available NNRTIs. A98S is a polymorphism that is not associated with NNRTI resistance. | Petropoulos et al., 2000; Bacheler et al., 2001 |
| 100 | L100I causes high-level resistance to EFV and NVP. Its effect on DLV susceptibility is not known. L100I suppresses T215Y-mediated AZT resistance. | Bacheler et al., 2001; Byrnes et al., 1994; Fujiwara et al., 1998; Petropoulos et al., 2000 |
| 101 | K101E is associated with intermediate resistance to the NNRTIs. K101R/Q occasionally occur in patients who have not received NNRTI and have not been associated with resistance to the current NNRTI. | Bacheler et al., 2001; Hanna et al., 2000a; Petropoulos, et al., 2000 |
| 103 | K103N causes resistance to NVP, DLV, and EFV. K103R occurs in about 1%-5% of persons receiving NRTI and probably does not cause NNRTI resistance. | Young et al., 1995, Bacheler et al., 2001; Demeter et al., 2000; Hanna et al., 2000a; Petropoulos et al., 2000; Shulman et al., 2000a |
| 106 | V106A causes high-level resistance to NVP, intermediate resistance to DLV, and low-level resistance to EFV. V106I occurs occasionally in patients not receiving NNRTIs and causes little if any phenotypic NNRTI resistance. | Bacheler et al., 2001; Byrnes et al., 1993; Larder et al., 1993a; Young et al., 1995 |
| 108 | V108I causes low-level resistance to each of the available NNRTI. | Petropoulos et al., 2000; Bacheler et al., 2001 |
| 179 | V179D/E is associated with resistance to the NNRTIs. V179I is probably a polymorphism that is not associated with NNRTI resistance. | Byrnes et al., 1993; Kleim et al., 1996; Winslow et al., 1996 |
| 181 | Y181C/I causes resistance to NVP and DLV. It causes low-level resistance to EFV. Y181C partially reverses T215Y-mediated AZT resistance. | Bacheler et al., 2001; Byrnes, et al., 1994; Byrnes et al., 1993; Larder, 1992; Petropoulos, et al., 2000 |
| 188 | Y188L causes high-level resistance to NVP and EFV and intermediate resistance to DLV. Y188C causes high-level resistance to NVP and low-level resistance to NVP and DLV. Y188H causes low-level resistance to each of the NNRTIs. | Byrnes et al., 1993 Bacheler, et al., 2001; Petropoulos et al., 2000 |
| 190 | G190A/S causes resistance to NVP and EFV but not to DLV. G190E and other mutations at this position may cause resistance to each of the NNRTIs, although these mutations have been associated with decreased HIV-1 replication. | Bacheler et al., 2001; Hanna, et al., 2000a; Huang et al., 2000b; Kleim et al., 1994 |
| 225 | P225H is associated with EFV resistance when present with other NNRTI mutations. It confers hyper-susceptibility to DLV. | Bacheler et al., 2001; Pelemans et al., 1998 |
| 227 | F227L is a recently described mutation that is associated with resistance to NVP, DLV, and EFV when present with other NNRTI mutations. | Bacheler et al., 2001; Balzarini et al., 1998 |
| 230 | M230L is a recently described mutation that causes high-level resistance to each of the currently available NNRTI. | Huang et al., 2000c |
| 234 | L234I confers resistance to an experimental NNRTI, AG-1549. Its effect on current NNRTIs is not known. | Fujiwara et al., 1998 |
| 236 | P236L causes DLV resistance. It confers hyper-susceptibility to NVP. | Dueweke et al., 1993 |
DLV: delavirdine, EFV: efavirenz, NVP: nevirapine
II. Approaches to HIV-1 Drug Resistance Testing
A. Source of HIV-1
Plasma is the main source of virus used for testing HIV-1 drug resistance in clinical settings. The sequence of plasma virus represents the quasispecies most recently selected for by antiretroviral drug therapy because plasma contains only actively replicating virus (Perelson et al., 1996). The evolution of HIV-1 sequences in peripheral blood mononuclear cells (PBMC) lags behind that in plasma (Koch et al., 1999; Kozal et al., 1993; Simmonds et al., 1991; Smith et al., 1993; Wei et al., 1995), and in patients failing therapy, mutations observed in plasma-isolated virus may not become the dominant quasispecies in PBMC until several weeks later.
Other sources of virus for HIV-1 drug resistance studies include resting T-lymphocytes and macrophages, lymph nodes, cerebrospinal fluid (CSF), and genital secretions. Reservoirs of virus harbored in long-lived cells such as resting T lymphocytes and macrophages pose the ultimate obstacle to HIV-1 eradication and have been an area of intense study (Pantaleo et al., 1998) (Finzi et al., 1997; Furtado et al., 1999; Wong et al., 1997b; Zhang et al., 2000). In patients with complete virus suppression during therapy (plasma HIV-1 RNA <50 copies/ml) there is generally little or no evidence of virus evolution in proviral DNA in long-lived cells (Ramratnam et al., 2000; Zhang et al., 2000). However, in patients with lapses in therapy, the population of virus within long-lived cells becomes replenished and may evolve drug resistant forms (Finzi et al., 1999; Ramratnam et al., 2000; Zhang et al., 1999).
The concentration of HIV-1 in lymph nodes is usually two to three orders of magnitude greater than the concentration of HIV-1 in plasma (Cavert et al., 1997; Chun et al., 1997; Embretson et al., 1993; Wang et al., 2000; Wong et al., 1997a). In patients receiving HAART, the concentration of HIV-1 in lymph nodes decreases in parallel with plasma viremia (Cavert et al., 1997; Notermans et al., 1998; Wong et al., 1997a). Several groups have sequenced protease and RT genes of virus recovered from proviral DNA in peripheral and visceral lymph nodes to determine the effect of antiviral therapy on virus in these heavily infected parts of the body (Haddad et al., 2000; Wong et al., 1997c). Most patients with undetectable levels of virus in the plasma still have detectable virus in lymph nodes (Wong et al., 1997a; Wong et al., 1997b), but there is no evidence that mutations develop in lymph nodes and yet remain absent from plasma (Dybul et al., 2000; Gunthard et al., 1998a; Schapiro et al., 1998; Erice et al., 2001).
It is not known to what extent HIV-1 in the central nervous system (CNS) reflects infection of the brain or simply mirrors what is present in the plasma (Daar, 1998). Some drugs, particularly the PIs, penetrate poorly into the CNS, and CSF virus has been sequenced to determine if the CNS is a sanctuary that permits virus replication during HAART therapy (Cunningham et al., 2000; Di Stefano et al., 1995; Gunthard et al., 2001; Venturi et al., 2000). But virologic failure in the CNS is rarely observed in the absence of virologic rebound in the plasma.
HIV-1 in the genital tract has been studied to determine whether specific variants are more likely to be transmitted either sexually or perinatally (Panther et al., 2000; Poss et al., 1995). In patients with ongoing virus replication, virus levels in genital secretions are usually proportional to virus levels in plasma. HIV-1 variants with genotypic resistance have been reported in genital secretions of both males and females receiving incompletely suppressive anti-HIV therapy (Di Stefano et al., 1999; Eron et al., 1998; Si-Mohamed et al., 2000). As in other latently infected cells, proviral HIV-1 DNA can occasionally be detected in nonspermatazoal mononuclear cells even in patients with plasma HIV-1 RNA levels <50 copies/ml (Gunthard et al., 2001; Zhang et al., 1998).
B. Phenotypic drug susceptibility testing
Phenotypic drug-susceptibility assays measure drug inhibition of HIV-1 in vitro. Two companies have developed standardized assays amenable to high-throughput performance (Virco, Mechelen, Belgium and ViroLogic, South San Francisco, CA, USA) (Hertogs et al., 1998; Petropoulos et al., 2000). Both assays amplify the entire protease, much of RT and some of gag from HIV-1 RNA extracted from patient plasma. The amplified material is incorporated into a pol-deleted recombinant virus construct using either ligation or homologous recombination. A standardized virus inoculum is then used to infect a cell line and virus replication is measured in the presence and absence of a range of concentrations of different antiretroviral drugs. Drug susceptibility is reported as the concentration of drug required to inhibit virus replication by 50% (IC50).
Recombinant virus susceptibility assays have several advantages over older non-recombinant assays. Recombinant virus assays can be done using plasma, whereas non-recombinant assays require the isolation of PBMCs. Recombinant virus assays use PCR to amplify protease and RT, dramatically decreasing the need for virus culture. Finally, recombinant virus assays can be performed under highly uniform conditions because the backbone of the virus construct, which remains constant, can be tailored for replication in the cells used for susceptibility testing.
The use of recombinant viruses for susceptibility testing, however, may not always be optimal. PI resistance is modulated by mutations at gag-pol cleavage sites. Four of the nine protease cleavage sites in the recombinant virus come from the patient virus sample, but five come from the laboratory virus construct. If a patient’s virus sample contained compensatory mutations at one of these five cleavage sites, the recombinant virus (lacking the compensatory mutations) might give inaccurate drug susceptibility results. The possibility that anomalous results might occur while testing either highly mutant viruses or viruses belonging to non-B subtypes has not yet been studied.
Recombinant and non-recombinant susceptibility tests suffer from the fact that the antiviral activities of NRTIs in vitro differ from the antiviral activities of these drugs in vivo. Abacavir and didanosine have one hundred times less antiviral activity than zidovudine in vitro. Yet abacavir and possibly also didanosine are more potent than zidovudine in vivo. Didanosine is weak in vitro because it is poorly triphosphorylated to its active form in the activated lymphocytes required for in vitro susceptibility testing (Gao et al., 1993; Shirasaka et al., 1995a). In vitro resistance to didanosine and stavudine are often impossible to detect even in patients experiencing virologic rebound while receiving these drugs. The poor triphosphorylation of didanosine may partly explain the difficulty in detecting didanosine resistance; but difficulty in detecting stavudine resistance is related to less well understood properties of the cells used for susceptibility testing (Meyer et al., 2000a; Lennerstrand et al., 2001).
C. HIV-1 genotypic testing
Genotypic tests are used more commonly in clinical settings because of their wider availability, lower cost, and shorter turnaround time. Genotypic tests provide more insight into the potential for resistance to emerge because they detect mutations present as mixtures, even if the mutation is present at a level too low to affect drug susceptibility in a phenotypic assay and they detect transitional mutations that do not cause resistance by themselves but indicate the presence of selective drug pressure. Genotypic testing has been shown to be clinically useful in four of five prospective randomized studies (Durant et al., 1999; Baxter et al., 2000; Tural et al., 2000, De Luca et al., 2000, Meynard et al., 2000); whereas phenotypic testing has been shown to be clinically useful in one of four prospective randomized studies (Cohen et al., 2000; Meynard et al., 2000; Melnick et al., 2000; Haubrich et al, 2001).
Figure 4 shows the distribution of known drug-resistance mutations within HIV-1 protease and RT. Mutations which may confer resistance to one or more drugs by themselves are represented by tall lines. Accessory mutations that confer resistance only when present with other mutations are represented by short lines. Sequencing for clinical purposes should encompass nearly all of the protease and positions 41–236 of the RT. Mutations at position 283 have been shown to cause low-level resistance to NNRTIs when present with other mutations (Brown et al., 2000); G333E is a naturally occurring polymorphism that facilitates zidovudine resistance in isolates from some patients receiving zidovudine and lamivudine that also have multiple zidovudine resistance mutations (Kemp et al., 1998) but dual resistance to these drugs usually emerges without this change (Masquelier et al., 1999; Shafer et al., 1998; Kuritzkes et al., 2000).
Figure 4.

Schematic diagram of showing the distribution of drug resistance mutations within the HIV-1 protease and RT genes. The nucleotide numbers relative to the HXB2 genome are shown (2258–3872). The protease is shown in green and the RT is shown in blue. Tall lines indicate the positions of mutations that confer resistance in the absence of other mutations. Short lines indicate the positions of accessory mutations that confer resistance only when present with other drug-resistance mutations. Protease inhibitor resistance mutations are shown in green; nucleoside RT inhibitor mutations are in blue; non-nucleoside RT inhibitor mutations are in yellow.
D. Clonal and population-based sequencing
The extent of genetic variation within an individual is lowest at the time of initial infection; and shortly after infection, usually only a single variant is detected (Burger et al., 1991; Diaz et al., 1997; Liu et al., 1997; Pang et al., 1992; Wolfs et al., 1992). Whether or not additional strains are transmitted but remain at levels too low to be detected is not known. The initial HIV-1 quasispecies, however, is more complex in those uncommon cases in which a person is initially infected with viruses from more than one source (Diaz et al., 1995; Long et al., 2000; Pieniazek et al., 1995; Zhu et al., 1995).
During the course of infection, virus sequence variation within an individual may range from about 1% to >5% in hypervariable regions of env (Brown, 1991; Wolfs et al., 1992). Several studies suggest that virus diversity is greater in patients mounting an anti-HIV-1 immune response (Liu et al., 1997; Lukashov et al., 1995; Shankarappa et al., 1999; Wolinsky et al., 1996). Genetic variability is usually lower in the plasma virus population compared with the cellular proviral DNA population. In the absence of drug therapy, genetic variability is usually lower in those genes coding for conserved proteins such as protease and RT than in envelope proteins (Imamichi et al., 2001; Quinones-Mateu et al., 1996).
Direct PCR, or population-based, sequencing is done in clinical settings because it is quicker and more affordable than sequencing multiple clones. Clonal sequencing is performed in research settings to answer questions about the evolution of HIV-1 drug resistance. For both population-based and clonal sequencing, the ability to detect minor variants is inversely related to the proportion of the minor variants within the whole virus population. In direct PCR sequencing, electrophoretic double peaks indicating a nucleotide mixture occur only when the second nucleotide is present in at least 20 percent of the total virus population. (D’Aquila, 2000; Gunthard et al., 1998b; Larder et al., 1993b; Schuurman et al., 1999b; Shafer et al., 2000c). With either method, unequal amplification of viral variants present as a mixture may occur because one or more species may be preferentially amplified during PCR due to differences in primer binding (Becker-Pergola et al., 2000b).
By sequencing multiple clones, one can measure the frequency of distinct variants within the HIV-1 quasispecies and assess the contribution of individual clones to the evolution of drug resistance. Clonal sequencing may detect dual infection and in vivo recombination (Long et al., 2000; Pieniazek et al., 1995), and can assess the co-linearity of mutations within a viral genome (D’Aquila, 2000; Zhang et al., 1997). Sequencing multiple clones from two different tissue samples or from the same tissue at two times enables statistical comparisons between the two virus populations (Imamichi et al., 2001; Wong et al., 1997c; Zhang et al., 2000). Finally, the composition of the HIV-1 quasispecies sheds light on the rates of mutation fixation, the fitness of mutant variants, and the effective size of the virus population in vivo (Brown and Cleland, 1996; Brown and Richman, 1997; Goudsmit et al., 1997; Goudsmit et al., 1996; Gunthard et al., 1999; Najera et al., 1995; Rodrigo, 1999; Rouzine and Coffin, 1999).
Cloning PCR-amplified genetic material into a plasmid or bacteriophage does not guarantee that each of the recovered clones will contain the sequence of a different virus. If the DNA used for cloning was amplified from a small number of cDNA or proviral DNA molecules, there is a risk that some clones may have been derived from the same initial template and thus be “PCR siblings” rather than DNA from different viruses. To guarantee the recovery of unique clones, some groups perform a limiting dilution of unamplified nucleic acid (viral RNA, cDNA, or proviral DNA) to ensure that only a single target sequence is amplified in each reaction (Brown and Simmonds, 1995); other groups dilute the starting material (but not to a limiting dilution) and then create one molecular clone from each dilution (Bacheler et al., 2000; Condra et al., 1996).
PCR may introduce errors and may also cause recombination due to template switching (Brown and Simmonds, 1995; Learn et al., 1996; Meyerhans et al., 1989). Taq polymerase is a low fidelity DNA polymerase which lacks proof-reading activity. The error rate for Taq is 20–100 × 106 (Sambrook and Russell, 2001). A 2 kb product amplified by nested PCR (e.g. 30 cycles × 2) could contain as many as 12 nucleotide changes, some of which will alter the encoded amino acid sequence (Learn et al., 1996). Use of a high fidelity thermostable DNA polymerase such as Pfu will minimize this problem (Cline et al., 1996). Limiting dilution PCR is required to prevent PCR recombination (Brown and Simmonds, 1995).
E. Sequencing by hybridization
The Affymetrix GeneChip® is designed to determine the complete sequence of HIV-1 protease and the first 1200 nucleotides of HIV-1 RT. Affymetrix uses photolithography and light-directed combinatorial chemistry to create precisely positioned and densely packed arrays of oligonucleotide probes on a glass wafer. The wafers are packaged in plastic cartridges that serve as hybridization chambers. Fluorescein-labeled RNA is transcribed from a cDNA sample and hybridized to the probes bound on a glass surface. Binding of the RNA to complementary probes is detected using a laser scanner. The GeneChip® is divided into several thousand segments each containing millions of similar probes designed to interrogate each of the nucleotide positions in a test DNA or RNA molecule. Every nucleotide in the test molecule requires at least four sets of oligonucleotide probes to determine whether that nucleotide is an A, C, G, or T.
The design or tiling of Affymetrix gene chips requires prior knowledge of the most commonly expected polymorphisms in a gene. Because of this requirement, this method of sequencing is also referred to as “re-sequencing”. The genetic variability of HIV-1 poses the ultimate challenge to sequencing by hybridization. Several studies comparing GeneChip® and dideoxynucleotide sequencing and have found that dideoxynulceotide sequencing is more reliable at detecting HIV-1 protease and RT mutations (Gunthard et al., 1998b; Hanna et al., 2000b; Vahey et al., 1999; Wilson et al., 2000) (Table 6). The current GeneChip® is not capable of detecting insertion or deletions and is unreliable at sequencing viral subtypes other than the subtype B on which the chip tiling has been based. In addition, genomic regions containing clusters of adjacent mutations can interfere with probe hybridization and result in errors. Improved microarrays for sequencing isolates belonging to subtypes A–F and for detecting insertions are under development (Myers et al., 2000).
Table 6.
Reproducibility of HIV-1 Protease and Reverse Transcriptase Sequencing
| Reference | Comparison | Main Findings |
|---|---|---|
| (Demeter et al., 1998) | Dideoxynucleotide cycle sequencing of cultured cell pellets in 11 laboratories | 99.7% nucleotide concordance. The homogeneity of cultured virus and the fact that RNA extraction and reverse transcription were not required probably contributed to the high concordance. |
| (Gunthard et al., 1998b) | Comparison of dideoxynucleotide and GeneChip sequencing of PCR-amplified DNA | 98.8% nucleotide concordance. |
| (Schuurman, et al., 1999b) | Dideoxy terminator and GeneChip sequencing of mixtures of plasmid clones in 23 laboratories. Mutations at three positions were studied. | 20/23 laboratories reliably detected 100% wildtype and 100% mutant codons. Most laboratories detected the mutants present as 50% mixtures. Laboratories were inconsistent at detecting mutations present as 25% mixtures. |
| (Schuurman, et al., 1999a) | Dideoxynucleotide and GeneChip sequencing of plasma samples seeded with different proportions of plasmid clones. | Detailed results pending. The number of correctly identified results did not vary among the three most widely used methods (ABI Kit, homebrew assay with ABI sequencer, VGI kit and sequencer). |
| (Vahey et al., 1999) | Comparison of dideoxynucleotide and GeneChip sequencing | GeneChip sequencing inaccurately tested non-subtype B sequences. |
| (Shafer et al., 2000c) | Dideoxynucleotide sequencing of replicate plasma samples in the same laboratory | 99.4% amino acid concordance. Most discordances were partial and were caused by the distribution of genetic variants within the plasma samples. |
| (Hanna et al., 2000b) | Comparison of dideoxynucleotide and GeneChip sequencing of cultured virus stock supernatant. | 97.4% concordance. Dideoxyterminator sequencing was more sensitive at detecting drug-resistance mutations. |
| (Wilson et al., 2000) | Comparison of dideoxynucleotide and GeneChip sequencing and Line Probe Assay | 96.6% concordance between dideoxynucleotide and GeneChip sequencing. |
| (Shafer et al., 2001) | Comparison of dideoxynucleotide sequencing of 46 plasma samples in two laboratories | 99.1% concordance. 90% of discordances were partial defined as one laboratory detecting a mixture and the second laboratory detecting just one of the components of the mixture. |
Point mutation assays are inexpensive and have the potential to be highly sensitive for mutations present in only a small proportion of circulating viruses (Servais et al., 2001a; Van Laethem et al., 1999). The INNO-LiPA® HIV-1 RT Assays (Innogenetics, Ghent, Belgium) is designed to detect NRTI-associated mutations at codons 41, 69, 70, 74, 75, 184, and 215 in the RT gene (Clarke et al., 2000; Stuyver et al., 1997). The INNO-LiPA® HIV-1 protease assay is designed to detect mutations at codons 30, 46, 48, 50, 54, 82, 84, and 90 in the protease gene (De Smet et al., 2000; Servais et al., 2001a; Servais et al., 2001b). The INNO-LiPA® assays have probes for wildtype and mutant alleles of each codon attached to a nitrocellulose strip. Biotin-labeled RT-PCR product from the patient sample is hybridized to the strip. An avidin-enzyme complex and the enzyme substrate produce a color change on the paper strip where the PCR product has hybridized with a probe. This assay is currently limited because it can only detect a subset of drug resistance mutations and has a 10% rate of uninterpretable results due to poor hybridization, which is particularly likely to occur when uncommon mutations occur at key codons (Puchhammer-Stockl et al., 1999; Servais et al., 2001a). Other point-mutation assays have been developed but have not been used in clinical settings (Eastman et al., 1998; Shafer et al., 1996).
III. Dideoxynucleotide cycle sequencing
A. General considerations
The template for HIV-1 protease and RT sequencing is prepared from plasma by RNA isolation, reverse transcription, and PCR amplification. These same procedures are required whether one uses a dedicated kit or an assembly of reagents obtained from separate vendors. Two commercial HIV-1 RT and protease genotyping kits described below are available and are under consideration by the FDA for use in clinical settings (Dileanis et al., 2000a; Ruiz et al., 2000). These kits have stronger quality control and validation profiles than home brew methods but may be more expensive and less versatile.
B. Preventing sample contamination
There are three major issues related to sample contamination in HIV-1 genotyping: contamination with RNases, cross-contamination of one sample with another, and contamination of PCR reactions with exogenous DNA and PCR products from previous reactions. RNases are present on hair and skin, and can easily contaminate solutions, plasticware, and equipment. Degradation of HIV-1 RNA by RNases can be minimized by wearing gloves, opening and handling samples and solutions behind protective shielding, and adding an RNase inhibitor to reverse transcription reactions (Sambrook and Russell, 2001).
Standard approaches have been adopted to minimize sample cross-contamination and contamination with PCR products from previous reactions (Kwok and Higuchi, 1989). Pre-and post-PCR steps should be performed in separate rooms and there should be no transfer of supplies, reagents or equipment from the post-PCR area to the pre-PCR area. Disposable gowns and gloves should be worn and stored in the pre-PCR area. A separate workstation - laminar flow hood with UV lights and dedicated pipettes - in the pre-PCR room or in another “clean room” should be used for all reagent preparation. Positive displacement pipettors and aerosol-resistant pipette tips should be used for all procedures and pipette tips should be changed between each addition. Reagents should be added to tubes before adding plasma RNA or DNA.
Each set of PCR reactions should include a negative control to be analyzed by agarose gel or sequencing to monitor for contamination. A uracil N-glycosylase (UNG) system can also be used to minimize contamination of PCR reactions with products generated in previous amplifications. In this system, PCR reactions are performed with deoxyuracil triphosphate (dUTP) in place of dTTP. Any contaminating dU-containing PCR products are degraded by UNG prior to PCR during a brief incubation (e.g. 50°C for 10 minutes). UNG is destroyed at the start of PCR by incubation at a higher temperature (e.g. 93°C).
C. RNA extraction
HIV-1 RNA extraction typically involves virus concentration, virus disruption, RNA recovery, and RNA purification. Typically, 0.2 ml – 1.0 ml of plasma is ultracentrifuged at ≥ 20,000 g for 30 to 60 minutes at 4°C. The resulting virus pellet, which is invisible, can be used directly or resuspended in nuclease-free water.
Most HIV-1 RNA extraction methods utilize a lysis solution that contains a chaotropic guanidine salt as its active ingredient. Guanidine thiocyanate releases RNA from the virus particle and protects it from degradation by RNases. Lysis solutions may also include phenol, urea, glycogen, carrier RNA, an RNase inhibitor, or dithiothreitol to promote efficient lysis and limit degradation of released RNA. The lysis solution is added to plasma or the concentrated virus pellet. Although the lysed virus is no longer infectious, guanidine thiocyanate is toxic and should be handled appropriately.
The released RNA can then be recovered using alcohol precipitation or by attachment to silica particles or columns. Alcohol precipitation can be performed either directly from the lysis solution (Erali and Hillyard, 1999) or after phenol-chloroform purification steps (Chomczynski and Sacchi, 1987). There are several commercial kits designed to facilitate RNA extraction. The QIAamp Viral RNA Mini Kit (Qiagen, Valencia, CA) contains a proprietary lysis reagent for virus disruption and a column containing a silica-based resin for RNA recovery and purification (Fischer et al., 1999; Fransen et al., 1998). The NucliSens Isolation Kit (Organon-Teknika, Durham, NC) is based on the Boom method (Boom et al., 1990) that uses a guanidinium thiocyanate-containing lysis reagent for virus disruption and silica particles for RNA recovery (Niubo et al., 2000; Verhofstede et al., 1996). The AMPLICOR HIV-1 MONITOR™ Test (Roche Diagnostics, Branchburg, NJ) performs HIV-1 quantitation using a 142-bp gag gene segment (Erali and Hillyard, 1999; Fransen et al., 1998; Mulder et al., 1997). Several laboratories have found that the RNA isolated using this kit can also be used for protease and RT sequencing (Shafer et al., 2000c). However, the kit is expensive, making its use as an extraction method practical only for laboratories performing both HIV-1 quantitation and sequencing.
D. Reverse transcription and PCR
Extracted viral RNA must be reverse transcribed to cDNA prior to PCR amplification. Reverse transcription involves incubating the RNA with dNTPs, a commercial RT enzyme, and a DNA primer. RT enzymes such as MMuLV (Moloney murine leukemia virus) and AMV (avian myeloblastosis virus) are most commonly used. RT enzymes that lack RNase H activity (e.g. Superscript, GIBCO BRL, Gaithersburg, MD), have increased thermostability, enabling reverse transcription to be performed at higher temperatures. This may increase yield by disrupting secondary RNA structure. Reverse transcription can be performed with random hexamer primers or, more commonly, a single HIV-1-specific primer. Reverse transcription and PCR can be carried out in one-step or two-step formats.
Following reverse transcription, PCR amplification is necessary to obtain enough target DNA for sequencing. PCR primers should bind to regions of the virus that are conserved and unaffected by drug treatment. Taq polymerase is adequate for population based sequencing, because Taq-induced errors may not be detected if many molecules were present in the starting nucleic acid preparation. However, sequence analyses of PCR-generated cloned DNA should be performed using an enzyme with proof reading activity, such as, Pfu which has an error rate of 1.6 × 106, compared to 20–100 × 106 for Taq polymerase (Cline et al., 1996; Sambrook and Russell, 2001).
In highly optimized system, samples with plasma HIV-1 RNA levels >1,000 copies/mL often have enough genetic material for sequencing after only one round of 30–40 PCR cycles. While nested PCR increases DNA yield and the sensitivity and specificity of the amplification, it also increases the effort and cost required for amplification, and the risk of sample cross-contamination.
E. Sequencing
Cycle sequencing using a highly processive, heat stable DNA polymerase allows sequencing reactions to be performed with only 1.0 μg of double-stranded template. Automated sequencers detect fluorescence from one or more dyes to identify A, C, G, and T termination reactions. Fluorescent dye labels can be incorporated into DNA extension products using either 5′-or 3′-dye labeled dideoxynucleotide triphosphates (ddNTPs). With dye terminator labeling, each of the four ddNTPs is tagged with a different fluorescent dye. With dye primer labeling, extension products are identified using primers tagged with four different fluorescent dyes in four separate base-specific reactions. Dye primers produce more consistent peak heights making them better for detecting and quantifying mixtures of bases at a given position. But dye primer sequencing requires four separate sequencing reactions for each primer. In contrast, dye terminators require only one reaction and are also more versatile because unlabeled primers can be used.
The quality of the template DNA (e.g. PCR product) is the most important factor for obtaining high quality sequence data. PCR products should be purified away from excess primers and ddNTPs prior to sequencing, although under some conditions, PCR products can be sequenced without purification if the yield of the desired product is high and the product is specific (produces a single band when analyzed by gel electrophoresis). Cycle sequencing reaction products should be purified to remove unbound fluorescent labels prior to electrophoretic separation of the reaction products. Electrophoretic peaks should be sharp, well-defined, and scaled high in the first several hundred nucleotides sequenced. Noisy data is most commonly caused by a “dirty” DNA template containing more than one priming site or an impurity that inhibits the sequencing reaction. Sequence quality can also be affected by gel-, instrument, and software-related problems.
F. Applied Biosystems ViroSeq™ HIV-1 Genotyping System
The Applied Biosystems ViroSeq™ HIV-1 Genotyping System includes reagents and protocols for every step of genotyping from RNA extraction to generation of a genotyping report. Specific features of this system are described in Table 7. RNA is extracted from plasma by a modification of the extraction technique in the AMPLICOR HIV-1 MONITOR™ UltraSensitive Method (Roche Diagnostics). RNA is reverse-transcribed with MMuLV RT and a 1.8 kb product is amplified using AmpliTaq Gold DNA polymerase and AmpErase dUTP/UNG, which minimizes the risk of sample cross-contamination. PCR products are purified prior to sequencing. BigDye™ dideoxyterminator sequencing reactions are prepared for six kit-specific primers (an alternate 7th primer is provided). Sequencing reactions are purified using isopropanol or columns and then run on a an ABI Prism® automated sequencer (310, 377, or 3100).
Table 7.
HIV-1 Protease and RT Sequencing Kits
| Applied Biosystems ViroSeq™ HIV-1 Genotyping System | Visible Genetics TRUGENE™ HIV-1 Genotyping Kit | |
|---|---|---|
| Sample requirements | ||
| Plasma volume | 500 ul | Depends on method used for extraction |
| Viral load | >2,000 RNA copies/ml is recommended | >1,000 RNA copies/ml is recommended |
| RNA isolation | ||
| Lysis and centrifugation using a modification of the MONITOR Ultrasensitive Method (Roche Diagnostics). | Separate kits can be purchased for RNA extraction. | |
| RT/PCR | ||
| RT | Single primer, MMuLV RT, RNase inhibitor | Single primer, MMuLV RT, RNase inhibitor |
| PCR | Non-nested, 40 cycles with Ampli- Taq Gold DNA polymerase. 1.8 kb product | Non-nested, 27 cycles with proprietary DNA polymerase. 1.3 kb product |
| Controls | Positive and negative controls included beginning with the RT/PCR step. dUTP/UNG Amperase | Positive and negative controls included beginning with the RT/PCR step. |
| Sample clean up | Microcon columns | None |
| Analysis | Agarose gel | None |
| Equipment needed | 9600 or 9700 thermal cycler | Laboratory’s preferred thermal cycler |
| Sequencing | ||
| Method | BigDye™ dideoxyterminators | CLIP bi-directional |
| Number of primers | 6–7 primers, one per reaction | 6 reactions, each containing a primer mix |
| Equipment | 9600 or 9700 thermal cycler, ABI 310, 377, 3100 | Customized equipment for gel preparation and electrophoresis |
| Throughput | 16 samples per 7 hour run on ABI 377 (96-well). 32 samples per run over 24 hours on ABI 3100 | 1 sample per 50 minute run. With 4 Long Read Towers, 28 samples in 8 hours |
| Data analysis system | ||
| Region analyzed | Protease 1-99, RT 1-320 | Protease 1-99, RT 40-247 |
| Computer platform | MacIntosh for ABI 377/ABI 310, PC or MacIntosh for ABI3100 | PC |
| Software features | Primer assembly, auto trimming, full view of electropherograms, translation and alignment to reference sequence, quick tab to positions of interest for editing. | Primer assembly, auto trimming, full view of electropherograms, translation and alignment to reference sequence, quicktab to positions of interest for editing. Includes “fingerprinting” comparison to prior runs. |
| Mutations reported | Resistance mutations, novel variants, insertions/deletions | Resistance mutations, known polymorphisms, novel variants, insertions/deletions |
| Sequence format | FASTA | FASTA (Ns may need to be deleted for data analysis) |
| Other reported information | Information about sequence quality and editing | Drug resistance interpretation |
HIV-1-specific analysis software is used to assemble data from each sequencing primer, compare the assembly to a reference sequence, and generate a report of results. Sequences can be saved in the FASTA format. This assay has been successful at sequencing RNA from a majority of patients with plasma HIV-1 RNA levels ≥ 1,000 copies/ml and subtypes A–G (Cunningham et al., 2001; Dileanis et al., 2000b; Marlowe et al., 2000).
G. Visible Genetics TRUGENE™ HIV-1 Genotyping Kit
The Visible Genetics (VGI) TRUGENE™ HIV-1 Genotyping Kit includes reagents for use with dedicated VGI equipment and is specifically designed for sequencing HIV-1 protease and RT. Specific features of this system are described in Table 7. Reagents for RNA extraction are not included, but are available from VGI as a separate kit, TRUPREP™. Extraction using other commercial systems is also validated. The kit employs a combined RT-PCR amplification followed by 4 dye-primer sequencing reactions called CLIP reactions, each with multiple primers.
The 1.3 kb RT/PCR product includes sequences for the entire protease gene and of codons 1–247 of the RT gene. The CLIP sequencing reactions provide sequences for the entire protease gene and codons 40–247 of the RT gene. Sequencing reactions are run using VGI-specific gel preparation and electrophoresis equipment. Each gel provides sequence information on one isolate. Sequencing reactions use two DNA primers, each labeled with a different fluorescent dye, for bidirectional sequencing. The CLIP reactions perform semi-logarithmic amplification of the product while determining its sequence, improving the sensitivity of sequencing samples with low copy numbers (Lee et al., 2001). Generic VGI sequence analysis software with HIV-specific modules is used to assemble the sequencing data, compare the results to a reference sequence, and generate a report of the results.
H. Sequence reproducibility
Inter-laboratory comparisons of sequence results obtained using cultured PBMCs, uncultured plasma, and prepared mixtures of HIV-1 plasmid clones indicate that dideoxynucleoside sequencing is highly reproducible in experienced laboratories (Table 6). In one study, the sequence concordance among 13 research laboratories performing dideoxynucleotide sequencing on cultured cell pellets was 99.7% at all nucleotide positions and 97% at positions associated with zidovudine resistance (Demeter et al., 1998). Sequencing cultured cell pellets is simpler than sequencing clinical samples such as plasma because RNA extraction and reverse transcription are not necessary and because cultured virus is more homogeneous and contains fewer mixtures than uncultured virus (Delassus et al., 1991; Kusumi et al., 1992). Nonetheless, the high inter-laboratory concordance in this study supports the intrinsic reliability of the dideoxy method for HIV-1 analysis.
Two large multicenter comparisons of sequence results obtained from samples containing mixtures of plasmid clones (ENVA-1) and spiked plasma samples (ENVA-2) have also been performed (Schuurman et al., 1999a; Schuurman et al., 1999b). These studies found that the ability of the participating laboratories to detect mutations was directly proportional to the percent of mutant plasmid clones within each mixture. Only a minority of laboratories detected mutations in mixtures in which the mutant clones made up less than 25% of the total.
Two clinical laboratories assessed the reproducibility of HIV-1 RT and protease sequencing using plasma aliquots obtained from 46 heavily treated HIV-1 infected individuals. Although both laboratories used sequencing reagents from Applied Biosystems, each used a different in-house protocol for plasma HIV-1 RNA extraction, reverse transcription, PCR, and sequencing. Overall sequence concordance between the two laboratories was 99.0%. But about 90% of the discordances were partial, defined as one laboratory detecting a mixture while the second laboratory detected only one of the mixture’s components (Figure 5). Complete discordances were significantly more likely to occur in plasma samples with lower plasma HIV-1 RNA levels. Nucleotide mixtures were detected at approximately 1% of the nucleotide positions, and, in every case in which one laboratory detected a mixture, the second laboratory detected either the same mixture or one of the mixture’s components. The high concordance in detecting mixtures and the fact that most discordance between the two laboratories was partial suggest that most discordance was due to variation in sampling the HIV-1 quasispecies rather than to technical artifact.
Figure 5.

Matrices showing the exact numbers of nucleotide concordances and discordances between two laboratories (A - vertical, left and B - horizontal, top) performing dideoxynucleotide sequencing on cryopreserved plasma aliquots from 46 heavily treated HIV-1 infected patients. Exact matches are shown along the diagonal. The numbers of partial discordances are written in black on a grey background and the numbers of complete discordances are written in red on a white background. R (A/G) and Y (C/T) represent transitions. M (A/C), W (A/T), K (G/T), and S (C/G) represent transversions. Data from the protease is shown at top and data from the reverse transcriptase (RT) is shown beneath. One RT sequence had a B and another had an H (not shown). There were no Ns or other highly ambiguous nucleotides. Adapted from (Shafer et al., 2001).
IV Sequence analysis
A. Sequence editing
Current automated sequencing platforms provide programs for assembling sequences of overlapping primers and for viewing multiply-aligned, equally-spaced electropherograms. Aligned electropherograms should be inspected at positions demonstrating ambiguous base calling (indicating possible nucleotide mixtures), positions associated with drug resistance, and positions with amino acid differences from the consensus subtype B reference sequence. The Applied Biosystems and Visible Genetics HIV-1 genotyping systems provide additional programs with HIV-1-specific features.
Because HIV-1 from any infected individual is genetically heterogeneous, the sequence of most clinical samples will contain positions with more than one electrophoretic peak indicating a mixture of nucleotides. In heavily treated patients, about 1% of positions show evidence of a nucleotide mixture but mixtures of more than two nucleotides at a single position are extremely rare (Shafer et al., 2001). Base-calling is affected by the background signal generated in each sequencing reaction. Reagents that reduce background signal will lead to an increased ability to detect the presence of minor variants within a mixture (Zakeri et al., 1998). There is a trade-off between calling too many mixtures, some of which may be false positives, and calling too few mixtures. Each laboratory should establish a consistent method for calling mixtures. A mixture rate higher than 2%–3% and the presence of more than two nucleotides at a single position suggests a high degree of sequencing noise.
B. Sequence formats
After a sequence has been edited it should be stored in a plain text format (e.g., FASTA, GenBank, GCG). Although less informative than other formats, the FASTA format offers the simplest way of dealing with sequence data in a human- and computer-readable fashion. In this format, the sequence is preceded by a line which begins with the “>“ character and which may contain one or more identifiers separated by the “|” character. The FASTA format is compatible with GenBank searches and most sequence analysis programs and is available as an export option in the software for ABI sequencers. Sequences generated with the TRUGENE™ HIV-1 Genotyping System contain “Ns” at the 5′ end and in the region between the non-contiguous protease and RT sequences, which may need to be manually deleted prior to analysis of sequence data with other software systems. There are several other commonly used sequence formats, as well as freely available tools that convert sequence files between different formats (e.g. readseq, ClustalW) (Ouellette, 1998).
C. Data management
A typical sequencing project typically generates a vast amount of raw data. Intelligent management of these data can save many hours during subsequent sequence analyses. All raw data, including sequencing run folders, should be backed up prior to review or manipulation. Electropherograms for each sample, FASTA files, and other reports can be organized into folders. Standardized worksheets can be generated to track dates and details of sample extraction, reverse transcription and PCR, product analysis, and sequencing reactions.
Sequence files should be stored both individually and as part of a sequence alignment in which each sequence is aligned to a consensus reference sequence. Creating a multiple sequence alignment for HIV-1 protease and RT sequences is straightforward because insertions and deletions are rare. The Los Alamos Laboratory HIV Sequence Database website has a tutorial which contains links to free and inexpensive programs for creating and editing multiple sequence alignments (e.g. ClustalW, BioEdit, GeneDoc). Joe Felsenstein’s site at the University of Washington and the IUBIO archive each provide a comprehensive set of links to free sequence analysis tools (see Appendix for URLs).
Appendix.
Web Resources for HIV Drug Resistance Testing in Clinical and Research Settings
| Web address | Features | |
|---|---|---|
| Drug resistance data | ||
| Los Alamos National Laboratory HIV Sequence Database | http://hiv-web.lanl.gov | Searchable HIV-related data bases including a database of drug-resistance mutations. |
| Stanford HIV RT and Protease Sequence Database | http://hivdb.stanford.edu/hiv | A comprehensive database of HIV RT and protease sequences linked to treatment data and phenotypic drug susceptibility data |
| International AIDS Society-USA | http://hivinsite.ucsf.edu/ | Contains the most recently published guide lines (Hirsch et al., 2000) along with simplified diagrams of key drug resistance mutations. |
| Sequence analysis programs | ||
| PHYLIP | http://evolution.genetics.washington.edu/phylip.html | PHYLIP is a free, comprehensive package of programs for phylogenetic analysis of sequence data. This site also a comprehensive set of links to other sequence analysis tools. |
| IUBIO archive | http://iubio.bio.indiana.edu/soft/molbio/ | A comprehensive set of links to sequence analysis programs. |
| Los Alamos National Laboratory HIV Sequence Database | http://hiv-web.lanl.gov | Programs for making alignments and phylogenetic trees, estimating synonymous/nonsynonymous substitutions, detecting signature mutations and inter-subtype recombination. |
| Stanford HIV RT and Protease Sequence Database | http://hivdb.stanford.edu/hiv (link to http://hiv-4.stanford.edu/cgi-bin/hivseq-web.pl and http://hiv-4.stanford.edu/cgi-test/hivtest-web.pl) | Two programs including HIV-SEQ and a drug resistance interpretation program. The release notes for HIV-SEQ contain the consensus B protease and RT sequences. |
| BLAST | http://ncbi.nlm.nih.gov | Nucleotide and amino acid sequence similarity search |
| TreeView | http://taxonomy.zoology.gla.ac.uk/rod/treeview.html | Viewing and manipulating tree files |
| Suppliers of systems for HIV sequence analysis | ||
| Applied Biosystems | http://www.appliedbiosy-stems.com | ViroSeqTM HIV-1 Genotyping System, reagents and equipment for home-brew assays |
| Visible Genetics | http://www.visgen.com | TRUGENE TM HIV-1 Genotyping Kit, TRUPREPTM RNA isolation kit |
| Innogenetics | http://www.innogenetics.com | INNO-LiPA(R) HIV-1 line probe assays |
| Affymetrix | http://www.affymetrix.com | Affymetrix GeneChip(R) Systems |
| Companies that provide phenotypic assays | ||
| Virco, Inc. | http://www.vircolab.com | Services include the AntivirogramTM phenotyping assay, VirtualPhenotypeTM, and GENChecTM genotyping assay |
| ViroLogic | http://www.virologic.com | Services include the PhenoSenseTM phenotyping assay |
There are also several comprehensive sequence analysis packages, mostly commercial, that allow users to manipulate, analyze, and annotate sequence data (e.g. GCG, DNASTAR, MacVector, the Staden Package, GDE). Some of these packages are expensive and most of their analysis tools are designed to help predict the function of previously uncharacterized sequence data. Although these packages can be useful in the management of HIV-1 protease and RT sequences, many of the programs are not necessary because the structure and function of HIV-1 protease and RT are already known.
In contrast, most sequence laboratories require a laboratory information management system (LIMS) to help meet the challenge of keeping track of many sequence variants, each of which is linked to a variable amount of external data (i.e. the treatment regimen of the person from whom the virus isolate was obtained, the drug susceptibility of the sequenced isolate). Although there are several expensive commercial LIMS under development, most investigators have developed their own LIMS in which the information associated with a sequence is stored in a spreadsheet or database.
D. Sequence analysis for quality control
Laboratory cross-contamination and sample mix-ups are common. Learn et al (1996) described two studies in which laboratory contamination or sample mix-up resulted in the publication of erroneous data. Frenkel et al (1998) also used similar sequence analyses to demonstrate that reports of transient HIV-1 viremia in perinatally exposed infants were also laboratory artifacts. All new sequences should therefore be analyzed for the presence of laboratory artifacts. Each sequence should be compared to the sequences of other HIV-1 isolates amplified at the same time and the distribution of pairwise genetic distances between these sequences should be examined (Learn et al., 1996). Sequences from epidemiologically unrelated individuals should be examined to determine whether the sequences are more closely related to one another than expected. Sequences from the same individual should be examined to determine whether the sequences diverge more than expected.
Phylogenetic trees containing each of the sequences obtained in a laboratory can also help identify closely related sequences. For this purpose, trees are generally created using rapid distance-based methods (e.g. neighbor joining algorithm) rather than by slower character-based methods (e.g. maximum parsimony and maximum likelihood algorithms). Programs for creating neighbor-joining trees can be found in phylogenetic software packages such as PHYLIP (Felsenstein, 1993) and PAUP (Swofford, 1998), as well as, in less comprehensive packages such as TreeCon (Van de Peer and De Wachter, 1997). TreeView is a useful free program for viewing and manipulating tree files on PCs and Macs (Page, 1996).
The Los Alamos Laboratory HIV Sequence Database web site contains tutorials on sequence quality control and on making phylogenetic trees (Kuiken et al., 1999). It also contains the following useful pages: (i) GenBank BLAST Search which compares a new sequence to published isolates with highly similar sequences to help exclude contamination with commonly used laboratory HIV-1 isolates, (ii) Neighbor TreeMaker which accepts submitted sequence alignments and creates a phylogenetic tree using the PHYLIP neighbor joining tree program, (iii) SeqPublish which creates a formatted version of a sequence alignment which makes it easier to spot potential problems, and (iv) Vespa which detects signature sequence patterns that should be shared among sequences obtained from the same patient or from the same group of clustered sequences (Korber and Myers, 1992).
Sequence analyses for quality control purposes require an understanding of the expected degree of intra- and inter-patient sequence variability and the use of programs that compute the nucleotide distance between sequence pairs. Among untreated individuals in the United States with subtype B virus, the mean inter-individual nucleotide divergence in protease and RT sequences is about 3%–4% (Figure 6). Among treated individuals harboring viruses with drug-resistance mutations, the mean inter-individual nucleotide divergence is even higher. Among untreated individuals a nucleotide distance of <1% in the protease or <2% in the RT is highly unusual and should prompt closer examination of the sequences for the possibility of laboratory contamination. Among treated individuals harboring viruses with drug-resistance mutations, a nucleotide distance of <2% in the protease or <3% in the RT should prompt closer examination of the sequences.
Figure 6.

Distribution of pairwise nucleotide distances between subtype B isolates from untreated persons. The figure with protease sequences contains approximately 180 sequences obtained between 1985–1998 and has been adapted from Figure 2 in (Shafer et al., 1999b). The figure with RT sequences contains approximately 250 sequences obtained between 1985–2000 and has been taken from data in the HIV RT and Protease Sequence Database through April 2001 (Kantor et al., 2001).
In geographic regions where HIV-1 has recently been introduced, the mean nucleotide divergence of protease and RT sequences from different individuals may be low. In cases such as these it may be necessary to use additional approaches to exclude cross-contamination. These additional methods include using Vespa to detect signature mutations that segregate sequences into clusters (Korber and Myers, 1992) and the creation of trees using synonymous nucleotide distances. Synonymous nucleotide distances are likely to diverge at a higher rate than nonsynonymous nucleotides. Because mixtures are so common in sequences of isolates from treated persons, it is essential to know how ambiguous nucleotides are handled by programs used for distance analyses.
Analysis of synonymous and nonsynonymous nucleotide changes provides insight into the extent to which sequence changes are neutral or caused by evolutionary selection. The two most useful programs for estimating synonymous and nonsynonymous nucleotide distances include the SNAP (Los Alamos Sequence Database, (Kuiken et al., 1999)) and MEGA (Nei and Gojobori, 1986).
In the Stanford University Hospital Diagnostic Virology Laboratory, a sequence management program compares new sequences to each of the sequences generated within the preceding two months and to all of the previous sequences of viruses from the same person. Each report is accompanied by an internal report containing a histogram showing the nucleotide distances between the new sequence and recent sequences. This allows laboratory contamination to be excluded each time a new sequence is completed. To detect sample mix-up, sequences from the same individual that differ by more than 3% are flagged and followed by a position by position summary of the differences between current and previous sequences showing which differences are at drug resistance positions and which differences are partial sequence differences caused by mixtures.
E. Sequence analysis of non-B subtypes
It might be expected that the functional constraints on the protease and RT enzymes would prevent their genes from displaying much inter-subtype variation. But several groups have shown that the protease and RT genes of global isolates are sufficiently different from one another to allow subtype grouping (Becker-Pergola et al., 2000a; Gonzales et al., 2001; Pieniazek et al., 2000; Vergne et al., 2000; Yahi et al., 2001). Although most inter-subtype variation is caused by synonymous nucleotide substitutions, there are subtype-specific amino acid patterns (Becker-Pergola et al., 2000a; Cornelissen et al., 1997; Gonzales et al., 2001; Pieniazek et al., 2000; Vergne et al., 2000).
In North America and Europe, most HIV-1 isolates belong to subtype B. Subtype B, however, accounts for only a small proportion of HIV-1 isolates worldwide and non-B isolates have been identified with increased frequency, particularly in Europe. The available anti-HIV drugs have been developed by drug screening and susceptibility testing using subtype B isolates, and drug resistance mutations have been identified and characterized almost entirely in subtype B isolates.
A few studies have tested the in vitro susceptibility of non-B HIV-1 isolates to antiretroviral drugs. Although Group O isolates often demonstrate intrinsic resistance to the NNRTIs (Descamps et al., 1997; Quinones-Mateu et al., 1997), most studies have shown that non-B Group M isolates are as susceptible as subtype B isolates to each of the three anti-HIV drug classes (Palmer et al., 1998; Shafer et al., 1999a; Shafer et al., 1997; Tanuri et al., 1999). Nonetheless, the number of isolates tested in these studies are too few for these studies to be conclusive. Moreover, subtype G isolates with reduced susceptibility to protease inhibitors (Descamps et al., 1998b) and subtype F isolates with reduced susceptibility to some NNRTIs have been described (Apetrei et al., 1998).
Protease inter-subtype variation is more likely than RT inter-subtype variation to occur at positions associated with drug resistance. A protease substrate cleft mutation, V82I occurs commonly in subtype G isolates. A few isolates with V82I have been studied in vitro and found to have low-level (<5-fold) protease inhibitor resistance (Brown et al., 2001; Descamps et al., 1998b; King et al., 1995) but there is no evidence that V82I has the same clinical significance as other mutations at this position such as V82A/T/F/S (Table 2). Among HIV-1 isolates from untreated persons, resistance-associated variants at positions 10, 20, and 36 occur more commonly in non-B isolates, whereas resistance-associated variants at positions 63, 77, and 93 occur more commonly in subtype B isolates (Gonzales et al., 2001).
There is no evidence for novel drug resistance mutations in non-B HIV-1 isolates and it is generally assumed that drug-resistance mutations described in the context of subtype B isolates will exert the same phenotypic effects in all HIV-1 subtypes. In a few reports, mutations characterized in subtype B HIV-1 were also observed in non-B HIV-1 following exposure to antiretroviral drugs (Caride et al., 2000; Eshleman et al., 2001; Yahi et al., 2001; Weidle et al., 2001; Adje et al., 2001). A study comparing the genotypes and phenotypes of >30 non-subtype B samples found no evidence for natural resistance to current anti-HIV drugs for subtypes A-H (de Bethune et al., 1999). Furthermore, in that study, resistance mutations detected in samples from patients with prior antiretroviral treatment were consistent with treatment history and did not differ among subtypes (de Bethune et al., 1999).
Inter-subtype genetic variability may complicate HIV-1 genotyping because primers used for reverse transcription, PCR, and sequencing may have a lower rate of annealing to non-B compared with subtype B templates. But the extent to which this occurs has not been studied. Several studies analyzing non-B HIV-1 are listed in Table 8. Primers from these studies may be more useful for sequencing non-B isolates than the primers typically used for sequencing subtype B isolates. Both the Applied Biosystems ViroSeq™ HIV-1 Genotyping System (Dileanis et al., 2000a; Eshleman et al., 2001; Fontane et al., 2000; Marlowe et al., 2000) and the Visible Genetics TRUGENE™ HIV-1 Genotyping System (Fontane et al., 2000; Lambert et al., 2000) have been used for the analysis of some non-B subtypes but the primers used in both commercial systems are proprietary.
Table 8.
Non-Subtype B HIV-1 Protease and Reverse Transcriptase Sequencing
| Type of study | Reference | Subtypes | Genotyping system |
|---|---|---|---|
| Sequences from untreated individuals | Lambert et al., 2000 | A-G | TRUGENE™ |
| Pieniazek et al., 2000 | A-D, F, H | Home brew | |
| Becker-Pergola et al., 2000a | A, D | PE Biosystems HIV-1 Genotyping System (version 1) | |
| Cornelissen et al., 1997 | A-E | Home brew | |
| Vergne et al., 2000 | A-D, F, G, J, K, group O | Home brew | |
| Drug susceptibilities of isolates from untreated individuals | Shafer et al., 1999a; Shafer et al., 1997 | C | Home brew |
| Palmer et al., 1998 | A-E | No genotyping | |
| Descamps et al., 1998a | A-H | Not stated | |
| Tanuri et al., 1999 | F | Home brew | |
| Apetrei et al., 1998 | F | Home brew | |
| de Bethune et al., 1999 | A-H | Virco | |
| Descamps et al., 1997 | Group O | Home brew | |
| Sequences from treated individuals | Descamps et al., 1997 | Group O | No genotyping |
| de Bethune et al., 1999 | A-H | Virco | |
| Eshleman et al., 2001 | A, D | ViroSeq™ | |
| Ushiro-Lamb et al., 2000 | A-D, F | TRUGENE™ | |
| Caride et al., 2000 | A, F | Home brew | |
| Quinones-Mateu et al., 1998 | Group O | Home brew | |
| Yahi et al., 2001 | A, A-G, C, D, F | ViroSeq™ | |
| Weidle et al., 2001 | A, C, D | Home brew | |
| Assay performance | Fontane et al., 2000 | A-J | TRUGENE™, ViroSeq™, Home brew |
| Marlowe et al., 2000 | A-G | ViroSeq™ | |
| Vahey et al., 1999 | A-F | Affymetrix GeneChip™ | |
| HIV PRT 440 |
V Drug resistance interpretation
A. General approach
The first step in drug resistance interpretation is to generate a list of amino acid differences between the sample sequence and a wildtype reference sequence (usually, the subtype B consensus sequence). The second step is to use the list of mutations to infer drug susceptibility and likelihood of response to specific anti-HIV drugs. Genotypic interpretation is independent of the process of genotyping. Therefore, laboratories doing HIV-1 genotyping can provide physicians with the option of receiving a file with the sequence data because such data can then be analyzed by interpretation systems other than those used by the sequencing laboratory.
HIV-1 drug resistance is rarely an all-or-none phenomenon and clinicians usually need the answers to the following two questions: (i) Does the genotype suggest that the patient will respond to a drug in a manner comparable to a patient with a wildtype isolate? (ii) Does the genotype suggest that the patient will obtain any antiviral benefit from the drug? The second question distinguishes HIV-1 susceptibility testing from anti-bacterial susceptibility testing. In the case of bacteria, it is usually possible to avoid using any drug with reduced susceptibility against a pathogen. It is also this second question that makes it necessary to include at least two and preferably three levels of resistance (e.g. low-level, intermediate, high-level).
Using HIV-1 sequences to infer drug susceptibility and likelihood of response to therapy is ideally performed by a computerized expert system because few clinicians can possibly remember all of what is known about each drug-resistance mutation. Table 9 contains several of the most commonly used systems for HIV-1 genotypic interpretation. During the next year, these algorithms will evolve and most likely converge through an ongoing process of inter-algorithm comparison and validation using clinical data sets. It is unlikely that algorithms will remain proprietary because there is no precedent for basing important medical decisions on proprietary unpublished data.
Table 9.
Algorithms for Interpreting HIV-1 Protease and RT Sequences
| Algorithm | Availability | Description |
|---|---|---|
| Resistance Collaborative Group (RCG) (DeGruttola et al., 2000) | Public | Table of rules developed for a standardized re-analysis of eight published studies linking drug resistance mutations and clinical outcome. Not intended for clinical use. |
| HIV RT and Protease Sequence Database (Shafer et al., 2000a) | Public | Method used at Stanford University Hospital Diagnostic Virology Laboratory. Mutations are assigned drug penalties. Drug penalties are added and drugs are assigned an inferred level of resistance. Drug penalties are hyperlinked to primary data linking mutation and drug. Program can be found at http://hivdb.stanford.edu. |
| French National Agency for AIDS Research (ANRS) (Rousseau et al., 2001) | Public | Table of rules listing mutations conferring genotypic resistance or possible genotypic resistance to anti-HIV drugs. |
| Retrogram™ (Virology Networks) | Proprietary | Drug-based rules. Updated regularly by an expert panel |
| GuideLines™ (Visible Genetics) | Proprietary | Drug-based rules. Updated regularly by an expert panel. |
| VirtualPhenotype™ (Virco; Mechelin, Belgium) (Verbiest, et al., 2000) | Proprietary | Pattern matching algorithm that uses a large genotype-phenotype correlative database to infer phenotypic properties based on sequence data.. |
An expert system performs reasoning over representations of human knowledge. It consists of a knowledge base and an inference engine. A computer-readable knowledge base will benefit patients, clinicians, and researchers because it is the first step towards creating an expert system and because it will identify gaps in what is known about drug-resistance mutations.
B. Knowledge base for drug resistance interpretation
How do we know what we know about drug-resistance mutations? The initial study of HIV-1 drug resistance typically involves culturing a wildtype HIV-1 isolate in the presence of increasing concentrations of the drug being studied, identifying mutations that allow the virus to continue to replicate, and confirming the phenotypic effect of these mutations in site-directed mutagenesis experiments (Table 10, Source A). While this process is the most rigorous means of demonstrating that a particular mutation confers drug resistance, it has two limitations. First, these studies identify mutations that are sufficient to cause resistance to the drug under evaluation without proving that these mutations are necessary for the development of drug resistance. Second, data from these studies are generally derived from isolates with only one or two drug-resistance mutations and rarely reflect the more complicated patterns of mutations observed in clinical isolates from patients receiving drug combinations.
Table 10.
Sources of Data on HIV Drug Resistance Mutations
| Source A. Correlation between genotype and phenotype based on laboratory isolates |
| The pre-clinical evaluation of a new drug often involves culturing a wildtype laboratory HIV-1 isolate in the presence of increasing drug concentrations, and identifying mutations that allow the virus to continue to replicate. Site-directed mutagenesis experiments are done to confirm that the mutations arising during virus passage in the presence of the drug confer drug resistance when introduced into a wildtype virus. Drug resistance mutations identified by this process acquire widespread acceptance, are referred to as “canonical” resistance mutations, and are often considered the predominant mutations responsible for resistance to the drug under evaluation. |
| Source B. Correlation between genotype and phenotype based on clinical isolates |
| Laboratory isolates often contain only one or two drug-resistance mutations and rarely reflect the more complicated patterns of mutations observed in clinical isolates from patients receiving combination drug therapy. The complexity of sequences obtained on clinical isolates often precludes site-directed mutagenesis. Instead statistical associations between drug resistance mutations and in vitro resistance are required to elucidate the role of specific mutations or mutation patterns in causing drug resistance. |
| Source C. Correlation between genotype and treatment history |
| Sequences of HIV-1 isolates from patients failing antiretroviral therapy are crucial observations of HIV evolution that show which virus mutations are most significant in vivo. Such data are also essential for elucidating the genetic mechanisms of resistance to drugs that are difficult to test in vitro. |
| Source D. Correlation between genotype and clinical outcome |
| Data correlating genotype and clinical response to subsequent antiretroviral therapy are the most clinically relevant and the most useful for clinicians who must select anti-HIV drugs for their patients. However, data of this type generally lags several years behind data from sources A–C. |
Three additional types of data must be taken into account before HIV genotyping can be used to the best advantage of patients. These data include correlations between genotype and phenotype of clinical HIV-1 isolates (Table 10, Source B), correlations between genotype and treatment history (Table 10, Source C), and correlations between genotype and clinical outcome (Table 10, Source D).
Data correlating genotype and phenotype of clinical isolates indicate the phenotypic effect of mutations in the genetic context in which they usually occur. Data correlating genotype and treatment history directly identify the mutations the virus used to escape anti-HIV drug suppression in vivo. Those data are particularly important for elucidating the mechanisms of resistance to drugs that are difficult to study in vitro. Data correlating genotype and the subsequent clinical outcome of patients who receive new treatment regimens identify which drugs are most likely to be effective in patients with specific resistance mutations.
Data correlating genotype with clinical outcome are the most relevant for clinical management of patients with HIV-1 infection. Such data, however, are available for a limited number of clinical scenarios, making it necessary for clinicians to rely on the other three types of available data (Table 10, Sources A–C) for clinical decision making.
C. Stanford University HIV RT and Protease Sequence Database
The HIV RT and Protease Sequence Database at Stanford University is an online database that contains published data on the first three types of data described above (Table 10, Sources A–C)). Efforts are underway to include data from studies that correlate genotype with clinical outcome (Kantor et al., 2001). As of May, 2001, the database contained 18,000 protease and RT sequences from about 3,500 individuals, as well as about 5,000 phenotypes on >1,000 HIV isolates. Users can retrieve sequence sets based on criteria such as previous drug treatment or presence of specific mutations. Users can also retrieve in vitro drug susceptibility results on isolates containing specific mutations.
The database website contains two sequence analysis program. The first program, HIV-SEQ, accepts user-submitted RT and protease sequences, compares them to a reference sequence, and uses the differences (mutations) as query parameters for interrogating the sequence database (Shafer et al., 2000a). HIV-SEQ allows users to examine new sequences in the context of previously published sequence data which provides two main advantages. Unusual sequence results can be detected allowing the person sequencing the gene to recheck the primary sequence output while it is still on the desktop. Unexpected associations between sequences or isolates can be discovered because the person analyzing a new sequence can immediately retrieve data on isolates sharing one or more mutations with the sequence.
The second program, a drug resistance interpretation program, accepts user-submitted protease and RT sequences and returns inferred levels of resistance to the 15 FDA-approved anti-HIV drugs. Each drug resistance mutation is assigned a drug penalty score; the total score for a drug is derived by adding the scores of each mutation associated with resistance to that drug. Using the total drug score, the program reports one of the following levels of inferred drug resistance: susceptible, potential low-level resistance, low-level resistance, intermediate resistance, high-level resistance. The genotypic interpretation is based on clinical, as well as, phenotypic data.
Mutation scores are derived from published literature linking mutations and anti-HIV drugs, including correlations between genotype and treatment history, genotype and phenotype, and genotype and clinical outcome (Table 10). The program usually assigns the same score to a mutation regardless of whether the mutation is present in pure form or as a mixture. Mutations that cause hypersusceptibility to a drug have a negative drug penalty score. The program generates a score sheet listing each of the drug penalty scores. A listing of all mutation/drug score pairs can be found with the program’s release notes (http://hiv-4.stanford.edu/cgi-test/hivtest-web.pl). Clicking on a drug penalty score provides a link to the primary data linking the mutation and drug. In addition to drug-penalty scores, the drug-resistance program generates a list of comments pertinent to the submitted sequence. Some of the comments are standard comments for specific mutations. Other comments are based on known interactions between combinations of mutations present in the sequence.
D. VirtualPhenotype™
Virco (Cambridge, UK and Mechelin, Belgium) uses a genotypic-phenotypic correlative data base to infer phenotypic properties based on sequence data. By performing both genotyping and phenotyping on >18,000 samples, they have generated a large genotype-phenotype database. The VirtualPhenotype™ is an interpretation system that uses the mutations within a sequence as parameters for searching the database for drug susceptibility data on isolates containing matching mutations (Verbiest et al., 2000). Over 60 unique amino acid positions are searched. The analysis includes a tabulation of the number of matches in the data base for each drug, and the distribution of phenotypes (fold increase in IC50) for the matching samples. The mean IC50 of the matching samples is interpreted using a biologically defined, drug-specific cut-off value, providing a quantitative prediction of drug resistance. The VirtualPhenotype™ has been shown to have a high correlation with results from Virco’s recombinant phenotypic assay, and to be an independent predictor of clinical response to antiretroviral therapy (Graham et al., 2001). The main limitations of the VirtualPhenotype™ are that it is based on a proprietary database and that it is entirely dependent on phenotypic data and cannot utilize data from clinical studies (Table 10, sources C and D).
References
- 1.Apetrei C, Descamps D, Collin G, Loussert-Ajaka I, Damond F, Duca M, Simon F, Brun-Vezinet F. Human immunodeficiency virus type 1 subtype F reverse transcriptase sequence and drug susceptibility. J Virol. 1998;72:3534–3538. doi: 10.1128/jvi.72.5.3534-3538.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Arion D, Kaushik N, McCormick S, Borkow G, Parniak MA. Phenotypic mechanism of HIV-1 resistance to 3¢-azido-3¢-deoxythymidine (AZT): increased polymerization processivity and enhanced sensitivity to pyrophosphate of the mutant viral reverse transcriptase. Biochemistry. 1998;37:15908–15917. doi: 10.1021/bi981200e. [DOI] [PubMed] [Google Scholar]
- 3.Arion D, Sluis-Cremer N, Parniak MA. Mechanism by which phosphonoformic acid resistance mutations restore 3¢-azido-3¢-deoxythymidine (AZT) sensitivity to AZT-resistant HIV-1 reverse transcriptase. J Biol Chem. 2000;275:9251–9255. doi: 10.1074/jbc.275.13.9251. [DOI] [PubMed] [Google Scholar]
- 4.Arts EJ, Quinones-Mateu ME, Albright JL, Marois JP, Hough C, Gu Z, Wainberg MA. 3¢-Azido-3¢-deoxythymidine (AZT) mediates cross-resistance to nucleoside analogs in the case of AZT-resistant human immunodeficiency virus type 1 variants. J Virol. 1998;72:4858–4865. doi: 10.1128/jvi.72.6.4858-4865.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bacheler L, Jeffrey S, Hanna G, D’Aquila R, Wallace L, Logue K, Cordova B, Hertogs K, Larder B, Buckery R, Baker D, Gallagher K, Scarnati H, Tritch R, Rizzo C. Genotypic correlates of phenotypic resistance to efavirenz in virus isolates from patients failing nonnucleoside reverse transcriptase inhibitor therapy. J Virol. 2001;75:4999–5008. doi: 10.1128/JVI.75.11.4999-5008.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bacheler L, Ploughman L, Hertogs K, Larder B. Impact of baseline NNRTI resistance on the efficacy of efavirenz combination therapy in NNRTI therapy-naive pateints (study DMP 266-006) [abstract 88] Antiviral Therapy. 2000;5(Supplement 3):70. [Google Scholar]
- 7.Baldwin ET, Bhat TN, Gulnik S, Liu B, Topol IA, Kiso Y, Mimoto T, Mitsuya H, Erickson JW. Structure of HIV-1 protease with KNI-272, a tight-binding transition- state analog containing allophenylnorstatine. Structure. 1995;3:581–590. doi: 10.1016/s0969-2126(01)00192-7. [DOI] [PubMed] [Google Scholar]
- 8.Bally F, Martinez R, Peters S, Sudre P, Telenti A. Polymorphism of HIV type 1 gag p7/p1 and p1/p6 cleavage sites: clinical significance and implications for resistance to protease inhibitors. AIDS Res Hum Retroviruses. 2000;16:1209–13. doi: 10.1089/08892220050116970. [DOI] [PubMed] [Google Scholar]
- 9.Balotta C, Berlusconi A, Pan A, Violin M, Riva C, Colombo MC, Gori A, Papagno L, Corvasce S, Mazzucchelli R, Facchi G, Velleca R, Saporetti G, Galli M, Rusconi S, Moroni M. Prevalence of transmitted nucleoside analogue-resistant HIV-1 strains and pre-existing mutations in pol reverse transcriptase and protease region: outcome after treatment in recently infected individuals. Antiviral Therapy. 2000;5:7–14. [PubMed] [Google Scholar]
- 10.Balzarini J, Pelemans H, Esnouf R, De Clercq E. A novel mutation (F227L) arises in the reverse transcriptase of human immunodeficiency virus type 1 on dose-escalating treatment of HIV type 1-infected cell cultures with the nonnucleoside reverse transcriptase inhibitor thiocarboxanilide UC-781. AIDS Res Hum Retroviruses. 1998;14:255–60. doi: 10.1089/aid.1998.14.255. [DOI] [PubMed] [Google Scholar]
- 11.Baxter JD, Mayers DL, Wentworth DN, Neaton JD, Hoover ML, Winters MA, Mannheimer SB, Thompson MA, Abrams DI, Brizz BJ, Ioannidis JP, Merigan TC. A randomized study of antiretroviral management based on plasma genotypic antiretroviral resistance testing in patients failing therapy. CPCRA 046 Study Team for the Terry Beirn Community Programs for Clinical Research on AIDS. AIDS. 2000;14:F83–F93. doi: 10.1097/00002030-200006160-00001. [DOI] [PubMed] [Google Scholar]
- 12.Becker-Pergola G, Kataaha P, Johnston-Dow L, Fung S, Jackson JB, Eshleman SH. Analysis of HIV type 1 protease and reverse transcriptase in antiretroviral drug-naive Ugandan adults. AIDS Res Hum Retroviruses. 2000a;16:807–13. doi: 10.1089/088922200308800. [DOI] [PubMed] [Google Scholar]
- 13.Becker-Pergola G, Mellquist JL, Eshleman JR, Jackson JB, Eshleman SH. Improved detection of HIV-1 variants by analysis of replicate amplification reactions: Relevance to studies of HIV-1 vertical transmission. Molecular Diagnosis. 2000b;4:261–268. doi: 10.1016/s1084-8592(99)80001-0. [DOI] [PubMed] [Google Scholar]
- 14.Bloor S, Hertogs K, Larder B, Pauwels R, Larder BA. Virological basis for HIV-1 resistance to stavudine investigated by analysis of clinical samples [abstract 15] Antivir Ther. 1998;3 (Suppl 1):15–16. [Google Scholar]
- 15.Boden D, Hurley A, Zhang L, Cao Y, Guo Y, Jones E, Tsay J, Ip J, Farthing C, Limoli K, Parkin N, Markowitz M. HIV-1 drug resistance in newly infected individuals. JAMA. 1999;282:1135–41. doi: 10.1001/jama.282.12.1135. [DOI] [PubMed] [Google Scholar]
- 16.Boom R, Sol CJ, Salimans MM, Jansen CL, Wertheim-van Dillen PM, van der Noordaa J. Rapid and simple method for purification of nucleic acids. J Clin Microbiol. 1990;28:495–503. doi: 10.1128/jcm.28.3.495-503.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Boucher CA, Cammack N, Schipper P, Schuurman R, Rouse P, Wainberg MA, Cameron JM. High-level resistance to (−) enantiomeric 2¢-deoxy-3¢-thiacytidine in vitro is due to one amino acid substitution in the catalytic site of human immunodeficiency virus type 1 reverse transcriptase. Antimicrob Agents Chem other. 1993;37:2231–2234. doi: 10.1128/aac.37.10.2231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Boyer PL, Sarafianos SG, Arnold E, Hughes SH. Selective excision of aztmp by drug-resistant human immunodeficiency virus reverse transcriptase. J Virol. 2001;75:4832–42. doi: 10.1128/JVI.75.10.4832-4842.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Briones C, Perez-Olmeda M, Rodriguez C, del Romero J, Hertogs K, Soriano V. Primary genotypic and phenotypic HIV-1 drug resistance in recent seroconverters in Madrid. J Acquir Immune Defic Syndr. 2001;26:145–50. doi: 10.1097/00042560-200102010-00006. [DOI] [PubMed] [Google Scholar]
- 20.Brown AJ. Sequence variability in human immunodeficiency viruses: pattern and process in viral evolution. AIDS. 1991;5:S35–42. [PubMed] [Google Scholar]
- 21.Brown AJ. Analysis of HIV-1 env gene sequences reveals evidence for a low effective number in the viral population. Proc Natl Acad Sci U S A. 1997;94:1862–5. doi: 10.1073/pnas.94.5.1862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Brown AJ, Cleland A. Independent evolution of the env and pol genes of HIV-1 during zidovudine therapy. AIDS. 1996;10:1067–1073. [PubMed] [Google Scholar]
- 23.Brown AJ, Precious HM, Whitcomb J, Simon V, Daar ES, D’Aquila R, Keiser P, Connick E, Hellmann N, Petropoulos C, Markowitz M, Richman D, Little SJ. Reduced susceptibility of HIV-1 to protease inhibitors from patients with primary HIV infection by three distinct routes [abstract 424]. 8th Conference on Retroviruses and Opportunistic Infections.2001. [Google Scholar]
- 24.Brown AJ, Precious HM, Whitcomb JM, Wong JK, Quigg M, Huang W, Daar ES, D’Aquila RT, Keiser PH, Connick E, Hellmann NS, Petropoulos CJ, Richman DD, Little SJ. Reduced susceptibility of human immunodeficiency virus type 1 (HIV-1) from patients with primary HIV infection to nonnucleoside reverse transcriptase inhibitors is associated with variation at novel amino acid sites. J Virol. 2000;74:10269–10273. doi: 10.1128/jvi.74.22.10269-10273.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Brown AJ, Richman DD. HIV-1: gambling on the evolution of drug resistance? Nat Med. 1997;3:268–271. doi: 10.1038/nm0397-268. [DOI] [PubMed] [Google Scholar]
- 26.Brown AJL, Simmonds P. Sequence analysis of virus variability based on the polymerase chain reaction (PCR) In: Karn J, editor. HIV. A Practical Approach. Volume 1. Virology and Immunology. 1995. pp. 161–186. [Google Scholar]
- 27.Burger H, Weiser B, Flaherty K, Gulla J, Nguyen PN, Gibbs RA. Evolution of human immunodeficiency virus type 1 nucleotide sequence diversity among close contacts. Proc Natl Acad Sci U S A. 1991;88:11236–40. doi: 10.1073/pnas.88.24.11236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Byrnes VW, Emini EA, Schleif WA, Condra JH, Schneider CL, Long WJ, Wolfgang JA, Graham DJ, Gotlib L, Schlabach AJ. Susceptibilities of human immunodeficiency virus type 1 enzyme and viral variants expressing multiple resistance-engendering amino acid substitutions to reserve transcriptase inhibitors. Antimicrob Agents Chem other. 1994;38:1404–1407. doi: 10.1128/aac.38.6.1404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Byrnes VW, Sardana VV, Schleif WA, Condra JH, Waterbury JA, Wolfgang JA, Long WJ, Schneider CL, Schlabach AJ, Wolanski BS. Comprehensive mutant enzyme and viral variant assessment of human immunodeficiency virus type 1 reverse transcriptase resistance to nonnucleoside inhibitors. Antimicrob Agents Chem other. 1993;37:1576–1579. doi: 10.1128/aac.37.8.1576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Caliendo AM, Savara A, An D, DeVore K, Kaplan JC, D’Aquila RT. Effects of zidovudine-selected human immunodeficiency virus type 1 reverse transcriptase amino acid substitutions on processive DNA synthesis and viral replication. J Virol. 1996;70:2146–2153. doi: 10.1128/jvi.70.4.2146-2153.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Caride E, Brindeiro R, Hertogs K, Larder B, Dehertogh P, Machado E, de Sa CA, Eyer-Silva WA, Sion FS, Passioni LF, Menezes JA, Calazans AR, Tanuri A. Drug-resistant reverse transcriptase genotyping and phenotyping of B and non-B subtypes (F and A) of human immunodeficiency virus type I found in Brazilian patients failing HAART. Virology. 2000;275:107–15. doi: 10.1006/viro.2000.0487. [DOI] [PubMed] [Google Scholar]
- 32.Cavert W, Notermans DW, Staskus K, Wietgrefe SW, Zupancic M, Gebhard K, Henry K, Zhang ZQ, Mills R, McDade H, Schuwirth CM, Goudsmit J, Danner SA, Haase AT. Kinetics of response in lymphoid tissues to antiretroviral therapy of HIV-1 infection. Science. 1997;276:960–964. doi: 10.1126/science.276.5314.960. [DOI] [PubMed] [Google Scholar]
- 33.Chen Z, Li Y, Schock HB, Hall D, Chen E, Kuo LC. Three-dimensional structure of a mutant HIV-1 protease displaying cross- resistance to all protease inhibitors in clinical trials. J Biol Chem. 1995;270:21433–6. doi: 10.1074/jbc.270.37.21433. [DOI] [PubMed] [Google Scholar]
- 34.Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate- phenol-chloroform extraction. Anal Biochem. 1987;162:156–9. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- 35.Chun TW, Carruth L, Finzi D, Shen X, DiGiuseppe JA, Taylor H, Hermankova M, Chadwick K, Margolick J, Quinn TC, Kuo YH, Brookmeyer R, Zeiger MA, Barditch-Crovo P, Siliciano RF. Quantification of latent tissue reservoirs and total body viral load in HIV-1 infection. Nature. 1997;387:183–188. doi: 10.1038/387183a0. [DOI] [PubMed] [Google Scholar]
- 36.Clarke JR, Kaye S, Babiker AG, Hooker MH, Tedder R, Weber JN. Comparison of a point mutation assay with a line probe assay for the detection of the major mutations in the HIV-1 reverse transcriptase gene associated with reduced susceptibility to nucleoside analogues. J Virol Methods. 2000;88:117–24. doi: 10.1016/s0166-0934(00)00173-7. [DOI] [PubMed] [Google Scholar]
- 37.Cline J, Braman JC, Hogrefe HH. PCR fidelity of pfu DNA polymerase and other thermostable DNA polymerases. Nucleic Acids Res. 1996;24:3546–51. doi: 10.1093/nar/24.18.3546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Coakley EP, Gillis JM, Hammer SM. Phenotypic and genotypic resistance patterns of HIV-1 isolates derived from individuals treated with didanosine and stavudine. AIDS. 2000;14:F9–15. doi: 10.1097/00002030-200001280-00002. [DOI] [PubMed] [Google Scholar]
- 39.Coffin JM. HIV population dynamics in vivo: implications for genetic variation, pathogenesis, and therapy. Science. 1995;267:483–489. doi: 10.1126/science.7824947. [DOI] [PubMed] [Google Scholar]
- 40.Cohen C, Kessler H, Hunt S, Sension M, Farthing C, Conant M, Jacobson S, Nadler M, Verbiest W, Hertogs K, Ames M, Rinehart A, Graham N. Phenotypic resistance testing significantly improves response to therapy: final analysis of a randomized trial (VIRA 3001) [abstract 67] Antivir Ther. 2000;5 (Supplememt 3):67. [Google Scholar]
- 41.Condra JH, Holder DJ, Schleif WA, Blahy OM, Danovich RM, Gabryelski LJ, Graham DJ, Laird D, Quintero JC, Rhodes A, Robbins HL, Roth E, Shivaprakash M, Yang T, Chodakewitz JA, Deutsch PJ, Leavitt RY, Massari FE, Mellors JW, Squires KE, Steigbigel RT, Teppler H, Emini EA. Genetic correlates of in vivo viral resistance to indinavir, a human immunodeficiency virus type 1 protease inhibitor. J Virol. 1996;70:8270–8276. doi: 10.1128/jvi.70.12.8270-8276.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Cornelissen M, van dB, Zorgdrager F, Lukashov V, Goudsmit J. pol gene diversity of five human immunodeficiency virus type 1 subtypes: evidence for naturally occurring mutations that contribute to drug resistance, limited recombination patterns, and common ancestry for subtypes B and D. J Virol. 1997;71:6348–6358. doi: 10.1128/jvi.71.9.6348-6358.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Cote HC, Brumme ZL, Harrigan PR. Human immunodeficiency virus type 1 protease cleavage site mutations associated with protease inhibitor cross-resistance selected by indinavir, ritonavir, and/or saquinavir. J Virol. 2001;75:589–94. doi: 10.1128/JVI.75.2.589-594.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Cunningham PH, Smith DG, Satchell C, Cooper DA, Brew B. Evidence for independent development of resistance to HIV-1 reverse transcriptase inhibitors in the cerebrospinal fluid. Aids. 2000;14:1949–54. doi: 10.1097/00002030-200009080-00010. [DOI] [PubMed] [Google Scholar]
- 45.Cunningham S, Ank B, Lewis D, Lu W, Wantman M, Dileanis J, Jackson JB, Palumbo P, Krogstad P, Eshleman SH. Performance of the applied biosystems viroseq human immunodeficiency virus type 1 (hiv-1) genotyping system for sequence-based analysis of hiv-1 in pediatric plasma samples. J Clin Microbiol. 2001;39:1254–7. doi: 10.1128/JCM.39.4.1254-1257.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Daar ES. Does the human immunodeficiency virus (HIV) RNA concentration in cerebrospinal fluid help clinicians diagnose or understand HIV-induced neurological disease? [editorial; comment] Clin Infect Dis. 1998;26:1074–5. doi: 10.1086/520300. [DOI] [PubMed] [Google Scholar]
- 47.D’Aquila RT. Limits of resistance testing. Antivir Ther. 2000;5:71–6. [PubMed] [Google Scholar]
- 48.de Bethune MP, Hertogs K, Heyndrickx L, Vingerhoets J, Fransen K, Azijn H, Michiels L, Janssens W, Scholliers A, Larder B, Bloor S, Pauwels R, Van der Groen G. Does natural or acquired resistance to reverse transcriptase and protease inhibitors, observed in HIV-1 groups M (subtypes A-H) and O, differ from subtype B ? Antivir Ther. 1999;4(Suppl 1):33. Abstract 49. [Google Scholar]
- 49.de Jong MD, Veenstra J, Stilianakis NI, Schuurman R, Lange JM, de Boer RJ, Boucher CA. Host-parasite dynamics and outgrowth of virus containing a single K70R amino acid change in reverse transcriptase are responsible for the loss of human immunodeficiency virus type 1 RNA load suppression by zidovudine. Proc Natl Acad Sci U S A. 1996;93:5501–6. doi: 10.1073/pnas.93.11.5501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.De Luca A, Antinori A, Cingolani A, Rizzo MG, Murri R, Ammassari A, Baldini F, Di Giambenedetto S, Marconi P, Ciancio B, Cauda R. A prospective randomized study on the usefulness of genotypic resistance testing and the assessment of patient-reported adherence in unselected patients failing potent HIV therapy (ARGENTA): Final 6-month results [abstract 433]. 8th Conference on Retroviruses and Opportunistic Infections.2001. [Google Scholar]
- 51.De Smet K, Celis L, Schmit J, Colebunders R, Miller V, Soriano V, Quinn TAM, Stuyver L. Identification of key mutations in the HIV-1 protease using the INNO-LiPA HIV protease assay [abstract 50] Antivir Ther. 2000;5:172–173. [Google Scholar]
- 52.Deeks SG. Nonnucleoside reverse transcriptase inhibitor resistance. J Acquir Immune Defic Syndr. 2001;26(Suppl 1):S25–33. doi: 10.1097/00042560-200103011-00004. [DOI] [PubMed] [Google Scholar]
- 53.DeGruttola V, Dix L, D’Aquila R, Holder D, Phillips A, Ait-Khaled M, Baxter J, Clevenbergh P, Hammer S, Harrigan R, Katzenstein D, Lanier R, Miller M, Para M, Yerly S, Zolopa A, Murray J, Patick A, Miller V, Castillo S, Pedneault L, Mellors J. The relation between baseline HIV drug resistance and response to antiretroviral therapy: re-analysis of retrospective and prospective studies using a standardized data analysis plan. Antivir Ther. 2000;5:41–48. doi: 10.1177/135965350000500112. [DOI] [PubMed] [Google Scholar]
- 54.Delassus S, Cheynier R, Wain-Hobson S. Evolution of human immunodeficiency virus type 1 nef and long terminal repeat sequences over 4 years in vivo and in vitro. J Virol. 1991;65:225–31. doi: 10.1128/jvi.65.1.225-231.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Delaugerre C, Mouroux M, Yvon-Groussin A, Simon A, Angleraud F, Huraux JM, Agut H, Katlama C, Calvez V. Prevalence and conditions of selection of E44D/A and V118I human immunodeficiency virus type 1 reverse transcriptase mutations in clinical practice. Antimicrob Agents Chem other. 2001;45:946–948. doi: 10.1128/AAC.45.3.946-948.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Demeter L, Haubrich R. Phenotypic and genotypic resistance assays: methodology, reliability, and interpretations. J Acquir Immune Defic Syndr. 2001;26(Suppl 1):S3–9. doi: 10.1097/00042560-200103011-00002. [DOI] [PubMed] [Google Scholar]
- 57.Demeter LM, D’Aquila R, Weislow O, Lorenzo E, Erice A, Fitzgibbon J, Shafer R, Richman D, Howard TM, Zhao Y, Fisher E, Huang D, Mayers D, Sylvester S, Arens M, Sannerud K, Rasheed S, Johnson V, Kuritzkes D, Reichelderfer P, Japour A. Interlaboratory concordance of DNA sequence analysis to detect reverse transcriptase mutations in HIV-1 proviral DNA. ACTG Sequencing Working Group AIDS Clinical Trials Group. J Virol Methods. 1998;75:93–104. doi: 10.1016/s0166-0934(98)00100-1. [DOI] [PubMed] [Google Scholar]
- 58.Demeter LM, Shafer RW, Meehan PM, Holden-Wiltse J, Fischl MA, Freimuth WW, Para MF, Reichman RC. Delavirdine susceptibilities and associated reverse transcriptase mutations in human immunodeficiency virus type 1 isolates from patients in a phase I/II trial of delavirdine monotherapy (ACTG 260) Antimicrob Agents Chem other. 2000;44:794–797. doi: 10.1128/aac.44.3.794-797.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Descamps DAC, Collins G, Damond F, Simon F, Brun-Vizinet F. In vitro drug susceptibility and sequence analysis of HIV-1 group M isolates belonging to different subtypes. 2nd International Workshop on HIV Drug Resistance, Treatment, and Eradication.1998a. [Google Scholar]
- 60.Descamps D, Apetrei C, Collin G, Damond F, Simon F, Brun-Vezinet F. Naturally occurring decreased susceptibility of HIV-1 subtype G to protease inhibitors [letter] AIDS. 1998b;12:1109–11. [PubMed] [Google Scholar]
- 61.Descamps D, Collin G, Letourneur F, Apetrei C, Damond F, Loussert-Ajaka I, Simon F, Saragosti S, Brun-Vezinet F. Susceptibility of human immunodeficiency virus type 1 group O isolates to antiretroviral agents: in vitro phenotypic and genotypic analyses. J Virol. 1997;71:8893–8898. doi: 10.1128/jvi.71.11.8893-8898.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Di Stefano M, Fiore JR, Monno L, Lepera A, Pastore G, Angarano G. Detection of multiple drug-resistance-associated pol mutations in cervicovaginal secretions from women largely treated with antiretroviral agents [letter] AIDS. 1999;13:992–4. doi: 10.1097/00002030-199905280-00018. [DOI] [PubMed] [Google Scholar]
- 63.Di Stefano M, Sabri F, Leitner T, Svennerholm B, Hagberg L, Norkrans G, Chiodi F. Reverse transcriptase sequence of paired isolates of cerebrospinal fluid and blood from patients infected with human immunodeficiency virus type 1 during zidovudine treatment. J Clin Microbiol. 1995;33:352–5. doi: 10.1128/jcm.33.2.352-355.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Diaz RS, Sabino EC, Mayer A, Mosley JW, Busch MP. Dual human immunodeficiency virus type 1 infection and recombination in a dually exposed transfusion recipient. The Transfusion Safety Study Group. J Virol. 1995;69:3273–81. doi: 10.1128/jvi.69.6.3273-3281.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Diaz RS, Zhang L, Busch MP, Mosley JW, Mayer A. Divergence of HIV-1 quasispecies in an epidemiologic cluster. AIDS. 1997;11:415–22. doi: 10.1097/00002030-199704000-00003. [DOI] [PubMed] [Google Scholar]
- 66.Dileanis J, Kunstman K, Wolinsky S, Palumbo P, Huang D, Brown RC, Buimer M, Schuurman R, Van Laethem K, Vandamme A-M. Multi-center testing of the Applied Biosystems ViroSeq HIV-1 genotyping kit version 2. XIII International AIDS Conference.2000a. [Google Scholar]
- 67.Dileanis J, Marlowe N, Hoo B, Brown RC, Buimer M, Huang D, Palumbo P, Schuurman R, Van Laethem K, Vandamme A-M, Elbeik T. Performance of the new ViroSeq genotyping system (version 2) with group M subtype panel and with subtype B clinical research samples at test sites. 5th International Congress on Drug Therapy in HIV Infection.2000b. [Google Scholar]
- 68.Ding J, Das K, Hsiou Y, Sarafianos SG, Clark AD, Jr, Jacobo-Molina A, Tantillo C, Hughes SH, Arnold E. Structure and functional implications of the polymerase active site region in a complex of HIV-1 RT with a double-stranded DNA template-primer and an antibody Fab fragment at 2.8 A resolution. J Mol Biol. 1998;284:1095–111. doi: 10.1006/jmbi.1998.2208. [DOI] [PubMed] [Google Scholar]
- 69.Doyon L, Croteau G, Thibeault D, Poulin F, Pilote L, Lamarre D. Second locus involved in human immunodeficiency virus type 1 resistance to protease inhibitors. J Virol. 1996;70:3763–9. doi: 10.1128/jvi.70.6.3763-3769.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Dueweke TJ, Pushkarskaya T, Poppe SM, Swaney SM, Zhao JQ, Chen IS, Stevenson M, Tarpley WG. A mutation in reverse transcriptase of bis(heteroaryl)piperazine- resistant human immunodeficiency virus type 1 that confers increased sensitivity to other nonnucleoside inhibitors. Proc Natl Acad Sci USA. 1993;90:4713–4717. doi: 10.1073/pnas.90.10.4713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Durant J, Clevenbergh P, Halfon P, Delgiudice P, Porsin S, Simonet P, Montagne N, Boucher CA, Schapiro JM, Dellamonica P. Drug-resistance genotyping in HIV-1 therapy: the VIRADAPT randomised controlled trial. Lancet. 1999;353:2195–2199. doi: 10.1016/s0140-6736(98)12291-2. [DOI] [PubMed] [Google Scholar]
- 72.Duwe S, Brunn M, Altmann D, Hamouda O, Schmidt B, Walter H, Pauli G, Kucherer C. Frequency of genotypic and phenotypic drug-resistant HIV-1 among therapy-naive patients of the german seroconverter study. J Acquir Immune Defic Syndr. 2001;26:266–73. doi: 10.1097/00042560-200103010-00010. [DOI] [PubMed] [Google Scholar]
- 73.Dybul M, Chun TW, Ward DJ, Hertogs K, Larder B, Fox CH, Orenstein JM, Baird BF, Li Y, Green LG, Engel D, Liu S, Mican JM, Fauci AS. Evaluation of lymph node virus burden in human immunodeficiency virus- infected patients receiving efavirenz-based protease inhibitor--sparing highly active antiretroviral therapy. J Infect Dis. 2000;181:1273–9. doi: 10.1086/315407. [DOI] [PubMed] [Google Scholar]
- 74.Eastman PS, Mittler J, Kelso R, Gee C, Boyer E, Kolberg J, Urdea M, Leonard JM, Norbeck DW, Mo H, Markowitz M. Genotypic changes in human immunodeficiency virus type 1 associated with loss of suppression of plasma viral RNA levels in subjects treated with ritonavir (Norvir) monotherapy. J Virol. 1998;72:5154–5164. doi: 10.1128/jvi.72.6.5154-5164.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Embretson J, Zupancic M, Ribas JL, Burke A, Racz P, Tenner-Racz K, Haase AT. Massive covert infection of helper T lymphocytes and macrophages by HIV during the incubation period of AIDS. Nature. 1993;362:359–62. doi: 10.1038/362359a0. [DOI] [PubMed] [Google Scholar]
- 76.Erali M, Hillyard DR. Evaluation of the ultrasensitive Roche Amplicor HIV-1 monitor assay for quantitation of human immunodeficiency virus type 1 RNA. J Clin Microbiol. 1999;37:792–5. doi: 10.1128/jcm.37.3.792-795.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Erice A, Li W, Balfour HH, Jr, Boies LR, Jr, Melroe H, Henry K. Analysis of HIV-1 reverse transcriptase and protease sequences in paired plasma and lymphoid tissue specimens from HIV-1 infected individuals. Aids. 2001;15:831–836. doi: 10.1097/00002030-200105040-00003. [DOI] [PubMed] [Google Scholar]
- 78.Erickson JW, Gulnik SV, Markowitz M. Protease inhibitors: resistance, cross-resistance, fitness and the choice of initial and salvage therapies. AIDS. 1999;13 (Suppl A):S189–204. [PubMed] [Google Scholar]
- 79.Eron JJ, Vernazza PL, Johnston DM, Seillier-Moiseiwitsch F, Alcorn TM, Fiscus SA, Cohen MS. Resistance of HIV-1 to antiretroviral agents in blood and seminal plasma: implications for transmission. AIDS. 1998;12:F181–F189. doi: 10.1097/00002030-199815000-00003. [DOI] [PubMed] [Google Scholar]
- 80.Eshleman SH, Mracna M, Guay L, Deseyve M, Cunningham S, Musoke P, Mmiro F, Jackson JB. Selection of nevirapine resistance (NVPR) mutations in Ugandan women and infants receiving NVP prophylaxis to prevent HIV-1 vertical transmission (HIVNET-012) [abstract 516]. 8th Conf. on Retroviruses and Opportunistic Infections; Chicago, IL. 2001. [Google Scholar]
- 81.Esnouf R, Ren J, Ross C, Jones Y, Stammers D, Stuart D. Mechanism of inhibition of HIV-1 reverse transcriptase by non- nucleoside inhibitors. Nat Struct Biol. 1995;2:303–8. doi: 10.1038/nsb0495-303. [DOI] [PubMed] [Google Scholar]
- 82.EuroGuidelines Group for HIV Resistance. Clinical and laboratory guidelines for the use of HIV-1 drug resistance testing as part of treatment management: recommendations for the European setting. AIDS. 2001;15:309–320. doi: 10.1097/00002030-200102160-00003. [DOI] [PubMed] [Google Scholar]
- 83.Falloon J, Piscitelli S, Vogel S, Sadler B, Mitsuya H, Kavlick MF, Yoshimura K, Rogers M, LaFon S, Manion DJ, Lane HC, Masur H. Combination therapy with amprenavir, abacavir, and efavirenz in human immunodeficiency virus (HIV)-infected patients failing a protease- inhibitor regimen: pharmacokinetic drug interactions and antiviral activity. Clin Infect Dis. 2000;30:313–8. doi: 10.1086/313667. [DOI] [PubMed] [Google Scholar]
- 84.Felsenstein J. PHYLIP (phylogeny inference package) version 3.5. University of Washington; Seattle: 1993. [Google Scholar]
- 85.Finzi D, Blankson J, Siliciano JD, Margolick JB, Chadwick K, Pierson T, Smith K, Lisziewicz J, Lori F, Flexner C, Quinn TC, Chaisson RE, Rosenberg E, Walker B, Gange S, Gallant J, Siliciano RF. Latent infection of CD4+ T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy. Nat Med. 1999;5:512–7. doi: 10.1038/8394. [DOI] [PubMed] [Google Scholar]
- 86.Finzi D, Hermankova M, Pierson T, Carruth LM, Buck C, Chaisson RE, Quinn TC, Chadwick K, Margolick J, Brookmeyer R, Gallant J, Markowitz M, Ho DD, Richman DD, Siliciano RF. Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science. 1997;278:1295–1300. doi: 10.1126/science.278.5341.1295. [DOI] [PubMed] [Google Scholar]
- 87.Fischer M, Huber W, Kallivroussis A, Ott P, Opravil M, Luthy R, Weber R, Cone RW. Highly sensitive methods for quantitation of human immunodeficiency virus type 1 RNA from plasma, cells, and tissues. J Clin Microbiol. 1999;37:1260–4. doi: 10.1128/jcm.37.5.1260-1264.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Fitzgibbon JE, Howell RM, Haberzettl CA, Sperber SJ, Gocke DJ, Dubin DT. Human immunodeficiency virus type 1 pol gene mutations which cause decreased susceptibility to 2¢,3¢-dideoxycytidine. Antimicrob Agents Chem other. 1992;36:153–157. doi: 10.1128/aac.36.1.153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Fontane E, Riva C, Peeters M, Schmit J-C, Delaporte E, VanLaethem K, VanVaerenbergh K, VanWijngaerden E, VanRanst M, Vandamme A-M. Evaluation of two commercial kits for the detection of genotypic drug-resistance on a panel of HIV-1 subtypes A-J. 5th International Conf. on Drug Therapy in HIV Infection.2000. [Google Scholar]
- 90.Fransen K, Mortier D, Heyndrickx L, Verhofstede C, Janssens W, van der Groen G. Isolation of HIV-1 RNA from plasma: evaluation of seven different methods for extraction (part two) J Virol Methods. 1998;76:153–7. doi: 10.1016/s0166-0934(98)00115-3. [DOI] [PubMed] [Google Scholar]
- 91.Frenkel LM, Mullins JI, Learn GH, Manns-Arcuino L, Herring BL, Kalish ML, Steketee RW, Thea DM, Nichols JE, Liu SL, Harmache A, He X, Muthui D, Madan A, Hood L, Haase AT, Zupancic M, Staskus K, Wolinsky S, Krogstad P, Zhao J, Chen I, Koup R, Ho D, Roberts NJ, Jr, et al. Genetic evaluation of suspected cases of transient HIV-1 infection of infants. Science. 1998;280:1073–7. doi: 10.1126/science.280.5366.1073. [DOI] [PubMed] [Google Scholar]
- 92.Frost SD, Nijhuis M, Schuurman R, Boucher CA, Brown AJ. Evolution of lamivudine resistance in human immunodeficiency virus type 1-infected individuals: the relative roles of drift and selection. J Virol. 2000;74:6262–8. doi: 10.1128/jvi.74.14.6262-6268.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Fujiwara T, Sato A, el-Farrash M, Miki S, Abe K, Isaka Y, Kodama M, Wu Y, Chen LB, Harada H, Sugimoto H, Hatanaka M, Hinuma Y. S-1153 inhibits replication of known drug-resistant strains of human immunodeficiency virus type 1. Antimicrob Agents Chem other. 1998;42:1340–1345. doi: 10.1128/aac.42.6.1340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Furtado MR, Callaway DS, Phair JP, Kunstman KJ, Stanton JL, Macken CA, Perelson AS, Wolinsky SM. Persistence of HIV-1 transcription in peripheral-blood mononuclear cells in patients receiving potent antiretroviral therapy [see comments] N Engl J Med. 1999;340:1614–22. doi: 10.1056/NEJM199905273402102. [DOI] [PubMed] [Google Scholar]
- 95.Gao WY, Shirasaka T, Johns DG, Broder S, Mitsuya H. Differential phosphorylation of azidothymidine, dideoxycytidine, and dideoxyinosine in resting and activated peripheral blood mononuclear cells. J Clin Invest. 1993;91:2326–2333. doi: 10.1172/JCI116463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Gonzales MJ, Machekano R, Shafer RW. HIV-1 reverse transcriptase and protease subtypes: Classification, amino acid patterns, and prevalence in a Northern California clinic-based population. J Infect Dis. 2001 doi: 10.1086/323601. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Goudsmit J, de RA, de RE, de BR. Broad spectrum of in vivo fitness of human immunodeficiency virus type 1 subpopulations differing at reverse transcriptase codons 41 and 215. J Virol. 1997;71:4479–4484. doi: 10.1128/jvi.71.6.4479-4484.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Goudsmit J, de RA, Ho DD, Perelson AS. Human immunodeficiency virus fitness in vivo: calculations based on a single zidovudine resistance mutation at codon 215 of reverse transcriptase. J Virol. 1996;70:5662–5664. doi: 10.1128/jvi.70.8.5662-5664.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Graham N, Peeters M, Verbeist W, Harrigan R, Larder B. The virtual phenotype is an independent predictor of clinical response [abstract 524]. 8th Conference on Retroviruses and Opportunistic Infections.2001. [Google Scholar]
- 100.Grant RM, Hecht F, Petropoulos C, Hellmann N, Warmerdam M, Bandrapalli NI, Gittens T, Chesney M, Kahn J. Trends in prevalence of primary HIV-1 drug resistance among recenty infected persons in San Francisco [abstract 142] Antivir Ther. 1999;4 (Supplement 1):98–99. [Google Scholar]
- 101.Gu Z, Fletcher RS, Arts EJ, Wainberg MA, Parniak MA. The K65R mutant reverse transcriptase of HIV-1 cross-resistant to 2¢, 3¢-dideoxycytidine, 2¢,3¢-dideoxy- 3¢-thiacytidine, and 2¢,3¢-dideoxyinosine shows reduced sensitivity to specific dideoxynucleoside triphosphate inhibitors in vitro. J Biol Chem. 1994a;269:28118–28122. [PubMed] [Google Scholar]
- 102.Gu Z, Gao Q, Fang H, Salomon H, Parniak MA, Goldberg E, Cameron J, Wainberg MA. Identification of a mutation at codon 65 in the IKKK motif of reverse transcriptase that encodes human immunodeficiency virus resistance to 2¢,3¢-dideoxycytidine and 2¢,3¢-dideoxy-3¢-thiacytidine. Antimicrob Agents Chem other. 1994b;38:275–281. doi: 10.1128/aac.38.2.275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Gu Z, Gao Q, Li X, Parniak MA, Wainberg MA. Novel mutation in the human immunodeficiency virus type 1 reverse transcriptase gene that encodes cross-resistance to 2¢,3¢-dideoxyinosine and 2¢,3¢-dideoxycytidine. J Virol. 1992;66:7128–7135. doi: 10.1128/jvi.66.12.7128-7135.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Gulnik SV, Suvorov LI, Liu B, Yu B, Anderson B, Mitsuya H, Erickson JW. Kinetic characterization and cross-resistance patterns of HIV-1 protease mutants selected under drug pressure. Biochemistry. 1995;34:9282–9287. doi: 10.1021/bi00029a002. [DOI] [PubMed] [Google Scholar]
- 105.Gunthard HF, Havlir DV, Fiscus S, Zhang ZQ, Eron J, Mellors J, Gulick R, Frost SD, Brown AJ, Schleif W, Valentine F, Jonas L, Meibohm A, Ignacio CC, Isaacs R, Gamagami R, Emini E, Haase A, Richman DD, Wong JK. Residual human immunodeficiency virus (hiv) type 1 rna and dna in lymph nodes and hiv rna in genital secretions and in cerebrospinal fluid after suppression of viremia for 2 years. J Infect Dis. 2001;183:1318–27. doi: 10.1086/319864. [DOI] [PubMed] [Google Scholar]
- 106.Gunthard HF, Leigh-Brown AJ, D’Aquila RT, Johnson VA, Kuritzkes DR, Richman DD, Wong JK. Higher selection pressure from antiretroviral drugs in vivo results in increased evolutionary distance in HIV-1 pol [published erratum appears in Virology 1999 Sep 1;261(2):367] Virology. 1999;259:154–165. doi: 10.1006/viro.1999.9774. [DOI] [PubMed] [Google Scholar]
- 107.Gunthard HF, Wong JK, Ignacio CC, Guatelli JC, Riggs NL, Havlir DV, Richman DD. Human immunodeficiency virus replication and genotypic resistance in blood and lymph nodes after a year of potent antiretroviral therapy. J Virol. 1998a;72:2422–2428. doi: 10.1128/jvi.72.3.2422-2428.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Gunthard HF, Wong JK, Ignacio CC, Havlir DV, Richman DD. Comparative performance of high-density oligonucleotide sequencing and dideoxynucleotide sequencing of HIV type 1 pol from clinical samples. AIDS Res Hum Retroviruses. 1998b;14:869–876. doi: 10.1089/aid.1998.14.869. [DOI] [PubMed] [Google Scholar]
- 109.Haddad DN, Birch C, Middleton T, Dwyer DE, Cunningham AL, Saksena NK. Evidence for late stage compartmentalization of HIV-1 resistance mutations between lymph node and peripheral blood mononuclear cells. Aids. 2000;14:2273–81. doi: 10.1097/00002030-200010200-00008. [DOI] [PubMed] [Google Scholar]
- 110.Hammond J, Calef C, Larder B, Schinazi RF, Mellors J. Mutations in retroviral genes associated with drug resistance. In: Kuiken CL, Foley B, Hahn BH, Marx P, McCutchan FE, Mellors J, Mullins JI, Wolinsky S, Korber B, editors. Human retroviruses and AIDS: a compilation and analysis of nucleic and amino acid sequences. Los Alamos National Laboratory; Los Alamos, NM: 1999. [Google Scholar]
- 111.Hanna GJ, D’Aquila RT. Clinical use of genotypic and phenotypic drug resistance testing to monitor antiretroviral chemotherapy. Clin Infect Dis. 2001;32:774–82. doi: 10.1086/319231. [DOI] [PubMed] [Google Scholar]
- 112.Hanna GJ, Johnson VA, Kuritzkes DR, Richman DD, Brown AJ, Savara AV, Hazelwood JD, D’Aquila RT. Patterns of resistance mutations selected by treatment of human immunodeficiency virus type 1 infection with zidovudine, didanosine, and nevirapine. J Infect Dis. 2000a;181:904–911. doi: 10.1086/315329. [DOI] [PubMed] [Google Scholar]
- 113.Hanna GJ, Johnson VA, Kuritzkes DR, Richman DD, Martinez-Picado J, Sutton L, Hazelwood JD, D’Aquila RT. Comparison of sequencing by hybridization and cycle sequencing for genotyping of human immunodeficiency virus type 1 reverse transcriptase. J Clin Microbiol. 2000b;38:2715–2721. doi: 10.1128/jcm.38.7.2715-2721.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Harrigan PR, Kinghorn I, Bloor S, Kemp SD, Najera I, Kohli A, Larder BA. Significance of amino acid variation at human immunodeficiency virus type 1 reverse transcriptase residue 210 for zidovudine susceptibility. J Virol. 1996;70:5930–5934. doi: 10.1128/jvi.70.9.5930-5934.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Harrigan PR, Stone C, Griffin P, Najera I, Bloor S, Kemp S, Tisdale M, Larder B. Resistance profile of the human immunodeficiency virus type 1 reverse transcriptase inhibitor abacavir (1592U89) after monotherapy and combination therapy. CNA2001 Investigative Group. J Infect Dis. 2000;181:912–920. doi: 10.1086/315317. [DOI] [PubMed] [Google Scholar]
- 116.Harzic M, Deveau C, Pellegrin I, Dubeaux b, Sageat P, Ngo N, Fleury H, Hoen B, Sereni D, Delfraissy JF. Prevalence of drug resistance mutations in patients with primary HIV-1 infection [abstract 91] Antivir Ther. 1999;4 (Supplement 1):61–62. [Google Scholar]
- 117.Haubrich R, Demeter L. Clinical utility of resistance testing: retrospective and prospective data supporting use and current recommendations. J Acquir Immune Defic Syndr. 2001;26 (Suppl 1):S51–9. doi: 10.1097/00042560-200103011-00006. [DOI] [PubMed] [Google Scholar]
- 118.Haubrich R, Keiser P, Kemper C, Witt M, Leedom J, Forthal D, Leibowitz M, Hwang J, Seefried E, McCutchan JA, Hellemann N, Richman D. CCTG 575: a randomized, prospective study of phenotype testing versus standard of care for patients failing antiretroviral therapy. Antivir Ther. 2001;6:63. [Google Scholar]
- 119.Havlir DV, Eastman S, Gamst A, Richman DD. Nevirapine-resistant human immunodeficiency virus: kinetics of replication and estimated prevalence in untreated patients. Antimicrob Agents Chemother. 1996;70:7894–7899. doi: 10.1128/jvi.70.11.7894-7899.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Hertogs K, Bloor S, De Vroey V, van Den Eynde C, Dehertogh P, van Cauwenberge A, Sturmer M, Alcorn T, Wegner S, van Houtte M, Miller V, Larder BA. A novel human immunodeficiency virus type 1 reverse transcriptase mutational pattern confers phenotypic lamivudine resistance in the absence of mutation 184V. Antimicrob Agents Chemother. 2000a;44:568–73. doi: 10.1128/aac.44.3.568-573.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Hertogs K, Bloor S, Kemp SD, Van den Eynde C, Alcorn TM, Pauwels R, Van Houtte M, Staszewski S, Miller V, Larder BA. Phenotypic and genotypic analysis of clinical HIV-1 isolates reveals extensive protease inhibitor cross-resistance: a survey of over 6000 samples. AIDS. 2000b;14:1203–10. doi: 10.1097/00002030-200006160-00018. [DOI] [PubMed] [Google Scholar]
- 122.Hertogs K, de Bethune MP, Miller V, Ivens T, Schel P, Van Cauwenberge A, Van Den Eynde C, Van Gerwen V, Azijn H, Van Houtte M, Peeters F, Staszewski S, Conant M, Bloor S, Kemp S, Larder B, Pauwels R. A rapid method for simultaneous detection of phenotypic resistance to inhibitors of protease and reverse transcriptase in recombinant human immunodeficiency virus type 1 isolates from patients treated with antiretroviral drugs. Antimicrob Agents Chem other. 1998;42:269–276. doi: 10.1128/aac.42.2.269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Hirsch MS, Brun-Vezinet F, D’Aquila RT, Hammer SM, Johnson VA, Kuritzkes DR, Loveday C, Mellors JW, Clotet B, Conway B, Demeter LM, Vella S, Jacobsen DM, Richman DD. Antiretroviral drug resistance testing in adult HIV-1 infection: recommendations of an International AIDS Society-USA Panel. JAMA. 2000;283:2417–2426. doi: 10.1001/jama.283.18.2417. [DOI] [PubMed] [Google Scholar]
- 124.Ho DD, Toyoshima T, Mo H, Kempf DJ, Norbeck D, Chen CM, Wideburg NE, Burt SK, Erickson JW, Singh MK. Characterization of human immunodeficiency virus type 1 variants with increased resistance to a C2-symmetric protease inhibitor. J Virol. 1994;68:2016–20. doi: 10.1128/jvi.68.3.2016-2020.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Holodniy M, Katzenstein D, Mole L, Winters M, Merigan T. Human immunodeficiency virus reverse transcriptase codon 215 mutations diminish virologic response to didanosine-zidovudine therapy in subjects with non-syncytium-inducing phenotype. J Infect Dis. 1996;174:854–857. doi: 10.1093/infdis/174.4.854. [DOI] [PubMed] [Google Scholar]
- 126.Hooker DJ, Tachedjian G, Solomon AE, Gurusinghe AD, Land S, Birch C, Anderson JL, Roy BM, Arnold E, Deacon NJ. An in vivo mutation from leucine to tryptophan at position 210 in human immunodeficiency virus type 1 reverse transcriptase contributes to high- level resistance to 3¢-azido-3¢-deoxythymidine. J Virol. 1996;70:8010–8018. doi: 10.1128/jvi.70.11.8010-8018.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Huang D, Bremer J, Brambilla D, Eshleman S, Nutter R, Hart S, Wantman M, Palumbo P. Proficiency testing of sequence-based genotypic assays [abstract 792]. 7th Conf. on Retroviruses and Opportunistic Infections; San Francisco, CA. 2000a. [Google Scholar]
- 128.Huang H, Chopra R, Verdine GL, Harrison SC. Structure of a covalently trapped catalytic complex of HIV-1 reverse transcriptase: implications for drug resistance. Science. 1998;282:1669–1675. doi: 10.1126/science.282.5394.1669. [DOI] [PubMed] [Google Scholar]
- 129.Huang W, Gamarnik A, Wrin T, Limoli K, Petropoulos C, Whitcomb J. NNRTI-resistance profiles, replicative capacity and protease processing defects in HIV-1 that contain RT mutations at amino acid 190 [abstract 29] Antivir Ther. 2000b;5(Supplement 3):23. [Google Scholar]
- 130.Huang W, Parkin NT, Lie YS, Wrin T, Haubrich R, Deeks S, Hellmann N, Petropoulos CJ, Whitcomb JM. A novel HIV-1 RT mutation (M230L) confers NNRTI resistance and dose-dependent stimulation of replication [abstract 30] Antivir Ther. 2000c;5 (Supplement 3):24–25. [Google Scholar]
- 131.Imamichi H, Crandall KA, Natarajan V, Jiang MK, Dewar RL, Berg S, Gaddam A, Bosche M, Metcalf JA, Davey RT, Jr, Lane HC. Human Immunodeficiency Virus Type 1 Quasi Species That Rebound after Discontinuation of Highly Active Antiretroviral Therapy Are Similar to the Viral Quasi Species Present before Initiation of Therapy. J Infect Dis. 2001;183:36–50. doi: 10.1086/317641. [DOI] [PubMed] [Google Scholar]
- 132.Iversen AK, Shafer RW, Wehrly K, Winters MA, Mullins JI, Chesebro B, Merigan TC. Multidrug-resistant human immunodeficiency virus type 1 strains resulting from combination antiretroviral therapy. J Virol. 1996;70:1086–1090. doi: 10.1128/jvi.70.2.1086-1090.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Izopet J, Bicart-See A, Pasquier C, Sandres K, Bonnet E, Marchou B, Puel J, Massip P. Mutations conferring resistance to zidovudine diminish the antiviral effect of stavudine plus didanosine. J Med Virol. 1999;59:507–11. [PubMed] [Google Scholar]
- 134.Jackson JB, Becker-Pergola G, Guay LA, Musoke P, Mracna M, Fowler MG, Mofenson LM, Mirochnick M, Mmiro F, Eshleman SH. Identification of the K103N resistance mutation in Ugandan women receiving nevirapine to prevent HIV-1 vertical transmission. AIDS. 2000;14:F111–5. doi: 10.1097/00002030-200007280-00001. [DOI] [PubMed] [Google Scholar]
- 135.Jacobo-Molina A, Ding J, Nanni RG, Clark AD, Jr, Lu X, Tantillo C, Williams RL, Kamer G, Ferris AL, Clark P, Hizi A, Hughes SH, Arnold E. Crystal structure of human immunodeficiency virus type 1 reverse transcriptase complexed with double-stranded DNA at 3.0 A resolution shows bent DNA. Proc Natl Acad Sci U S A. 1993;90:6320–4. doi: 10.1073/pnas.90.13.6320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Jacobsen H, Yasargil K, Winslow DL, Craig JC, Krohn A, Duncan IB, Mous J. Characterization of human immunodeficiency virus type 1 mutants with decreased sensitivity to proteinase inhibitor Ro 31–8959. Virology. 1995;206:527–534. doi: 10.1016/s0042-6822(95)80069-7. [DOI] [PubMed] [Google Scholar]
- 137.Japour AJ, Welles S, D’Aquila RT, Johnson VA, Richman DD, Coombs RW, Reichelderfer PS, Kahn JO, Crumpacker CS, Kuritzkes DR. Prevalence and clinical significance of zidovudine resistance mutations in human immunodeficiency virus isolated from patients after long-term zidovudine treatment. AIDS Clinical Trials Group 116B/117 Study Team and the Virology Committee Resistance Working Group. J Infect Dis. 1995;171:1172–1179. doi: 10.1093/infdis/171.5.1172. [DOI] [PubMed] [Google Scholar]
- 138.Kantor R, Machekano R, Gonzales MJ, Dupnik BS, Schapiro JM, Shafer RW. Human immunodeficiency virus reverse transcriptase and protease sequence database: An expanded model integrating natural language text and sequence analysis. Nucleic Acids Res. 2001;29:296–299. doi: 10.1093/nar/29.1.296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Kellam P, Boucher CA, Larder BA. Fifth mutation in human immunodeficiency virus type 1 reverse transcriptase contributes to the development of high-level resistance to zidovudine. Proc Natl Acad Sci USA. 1992;89:1934–1938. doi: 10.1073/pnas.89.5.1934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Kemp SD, Shi C, Bloor S, Harrigan PR, Mellors JW, Larder BA. A novel polymorphism at codon 333 of human immunodeficiency virus type 1 reverse transcriptase can facilitate dual resistance to zidovudine and L-2¢,3¢-dideoxy-3¢-thiacytidine. J Virol. 1998;72:5093–5098. doi: 10.1128/jvi.72.6.5093-5098.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Kempf D, Isaacson J, King M, Brun S, Xu Y, Real K, Lie Y, Hellmann N, Hertogs K, Larder B, Bernstein B, Japour A, Sun E, Rode R. Genotypic correlates of reduced in vitro susceptibility to ABT-378 in HIV isolates from patients failing protease inhibitor therapy [abstract 38] Antivir Ther. 2000;5(Supplement 3):29–30. [Google Scholar]
- 142.King RW, Winslow DL, Garber S, Scarnati HT, Bachelor L, Stack S, Otto MJ. Identification of a clinical isolate of HIV-1 with an isoleucine at position 82 of the protease which retains susceptibility to protease inhibitors. Antiviral Res. 1995;28:13–24. doi: 10.1016/0166-3542(95)00033-i. [DOI] [PubMed] [Google Scholar]
- 143.Kleim JP, Bender R, Kirsch R, Meichsner C, Paessens A, Riess G. Mutational analysis of residue 190 of human immunodeficiency virus type 1 reverse transcriptase. Virology. 1994;200:696–701. doi: 10.1006/viro.1994.1233. [DOI] [PubMed] [Google Scholar]
- 144.Kleim JP, Rosner M, Winkler I, Paessens A, Kirsch R, Hsiou Y, Arnold E, Riess G. Selective pressure of a quinoxaline nonnucleoside inhibitor of human immunodeficiency virus type 1 (HIV-1) reverse transcriptase (RT) on HIV- 1 replication results in the emergence of nucleoside RT-inhibitor- specific (RT Leu-74-->Val or Ile and Val-75-->Leu or Ile) HIV-1 mutants. Proc Natl Acad Sci USA. 1996;93:34–38. doi: 10.1073/pnas.93.1.34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Koch N, Yahi N, Ariasi F, Fantini J, Tamalet C. Comparison of human immunodeficiency virus type 1 (HIV-1) protease mutations in HIV-1 genomes detected in plasma and in peripheral blood mononuclear cells from patients receiving combination drug therapy. J Clin Microbiol. 1999;37:1595–7. doi: 10.1128/jcm.37.5.1595-1597.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Kohlstaedt LA, Steitz TA. Reverse transcriptase of human immunodeficiency virus can use either human tRNA(3Lys) or Escherichia coli tRNA(2Gln) as a primer in an in vitro primer-utilization assay. Proc Natl Acad Sci U S A. 1992;89:9652–6. doi: 10.1073/pnas.89.20.9652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Kohlstaedt LA, Wang J, Friedman JM, Rice PA, Steitz TA. Crystal structure at 3.5 A resolution of HIV-1 reverse transcriptase complexed with an inhibitor. Science. 1992;256:1783–90. doi: 10.1126/science.1377403. [DOI] [PubMed] [Google Scholar]
- 148.Korber B, Myers G. Signature pattern analysis: a method for assessing viral sequence relatedness. AIDS Res Hum Retroviruses. 1992;8:1549–1560. doi: 10.1089/aid.1992.8.1549. [DOI] [PubMed] [Google Scholar]
- 149.Kozal MJ, Shafer RW, Winters MA, Katzenstein DA, Merigan TC. A mutation in human immunodeficiency virus reverse transcriptase and decline in CD4 lymphocyte numbers in long-term zidovudine recipients. J Infect Dis. 1993;167:526–532. doi: 10.1093/infdis/167.3.526. [DOI] [PubMed] [Google Scholar]
- 150.Kozal MJ, Kroodsma K, Winters MA, Shafer RW, Efron B, Katzenstein DA, Merigan TC. Didanosine resistance in HIV-infected patients switched from zidovudine to didanosine monotherapy. Ann Intern Med. 1994;121:263–268. doi: 10.7326/0003-4819-121-4-199408150-00005. [DOI] [PubMed] [Google Scholar]
- 151.Kuiken CL, Foley B, Hahn B, Korber B, McCutchan F, Marx PA, Mellors JW, Mullins JI, Sodroski J, Wolinksy S. Human retroviruses and AIDS 1999: a compilation and analysis of nucleic and amino acid sequences. Theoretical Biology and Biophysics Group, Los Alamos National Laboratory; Los Alamos, NM: 1999. [Google Scholar]
- 152.Kuritzkes DR, Shugarts D, Bakhtiari M, Poticha D, Johnson J, Rubin M, Gingeras TR, Kennedy M, Eron JJ. Emergence of dual resistance to zidovudine and lamivudine in HIV-1- infected patients treated with zidovudine plus lamivudine as initial therapy. J Acquir Immune Defic Syndr. 2000;23:26–34. doi: 10.1097/00126334-200001010-00004. [DOI] [PubMed] [Google Scholar]
- 153.Kusumi K, Conway B, Cunningham S, Berson A, Evans C, Iversen AK, Colvin D, Gallo MV, Coutre S, Shpaer EG, et al. Human immunodeficiency virus type 1 envelope gene structure and diversity in vivo and after cocultivation in vitro. J Virol. 1992;66:875–85. doi: 10.1128/jvi.66.2.875-885.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Kwok S, Higuchi R. Avoiding false positives with PCR. Nature. 1989;339:237–8. doi: 10.1038/339237a0. [DOI] [PubMed] [Google Scholar]
- 155.Lacey SF, Larder BA. Novel mutation (V75T) in human immunodeficiency virus type 1 reverse transcriptase confers resistance to 2¢,3¢-didehydro-2¢,3¢-dideoxythymidine in cell culture. Antimicrob Agents Chem other. 1994;38:1428–1432. doi: 10.1128/aac.38.6.1428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Lambert C, Fontaine E, Servais J, Robert I, Arendt V, Staub T, Hemmer R, Schmit JC, Schneider F. Drug resistance-related mutations in the pol gene of non-subtype B HIV-1 patient strains. Antivir Ther. 2000;5(Supplement 3) [Google Scholar]
- 157.Lanier R, Scott J, Steel H, Hetherington B, Ait-Khaled M, Pearce G, Spreen W, Lafon S. Multivariate analysis of predictors of response to abacavir: comparison of prior antiretroviral therapy, baseline HIV RNA, CD4 count and viral resistance [abstract 82] Antivir Ther. 1999;4(Suppl 1):56. [Google Scholar]
- 158.Larder BA, Kemp SD. Multiple mutations in HIV-1 reverse transcriptase confer high-level resistance to zidovudine (AZT) Science. 1989;246:1155–1158. doi: 10.1126/science.2479983. [DOI] [PubMed] [Google Scholar]
- 159.Larder BA. 3¢-Azido-3¢-deoxythymidine resistance suppressed by a mutation conferring human immunodeficiency virus type 1 resistance to nonnucleoside reverse transcriptase inhibitors. Antimicrob Agents Chem other. 1992;36:2664–2669. doi: 10.1128/aac.36.12.2664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Larder BA, Kellam P, Kemp SD. Zidovudine resistance predicted by direct detection of mutations in DNA from HIV-infected lymphocytes. AIDS. 1991;5:137–44. doi: 10.1097/00002030-199102000-00002. [DOI] [PubMed] [Google Scholar]
- 161.Larder BA, Kellam P, Kemp SD. Convergent combination therapy can select viable multidrug-resistant HIV-1 in vitro. Nature. 1993a;365:451–453. doi: 10.1038/365451a0. [DOI] [PubMed] [Google Scholar]
- 162.Larder BA, Kohli A, Kellam P, Kemp SD, Kronick M, Henfrey RD. Quantitative detection of HIV-1 drug resistance mutations by automated DNA sequencing. Nature. 1993b;365:671–673. doi: 10.1038/365671a0. [DOI] [PubMed] [Google Scholar]
- 163.Larder BA. Interactions between drug resistance mutations in human immunodeficiency virus type 1 reverse transcriptase. J Gen Virol. 1994;75:951–957. doi: 10.1099/0022-1317-75-5-951. [DOI] [PubMed] [Google Scholar]
- 164.Larder BA, Kemp SD, Harrigan PR. Potential mechanism for sustained antiretroviral efficacy of AZT-3TC combination therapy. Science. 1995;269:696–699. doi: 10.1126/science.7542804. [DOI] [PubMed] [Google Scholar]
- 165.Larder BA, Stammers DK. Closing in on HIV drug resistance. Nat Struct Biol. 1999;6:103–6. doi: 10.1038/5787. [DOI] [PubMed] [Google Scholar]
- 166.Learn GH, Jr, Korber BT, Foley B, Hahn BH, Wolinsky SM, Mullins JI. Maintaining the integrity of human immunodeficiency virus sequence databases. J Virol. 1996;70:5720–5730. doi: 10.1128/jvi.70.8.5720-5730.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Lee SY, Sandhu M, Griswold MP, Van Gorder M. Validation of the ultrasensitive HIV-1 genotyping assay [abstract 249]. 8th Conference on Retroviruses and Opportunistic Infections.2001. [Google Scholar]
- 168.Lennerstrand J, Stammers DK, Larder BA. Biochemcial mechanism of human immunodeficiency virus type 1 reverse transcriptase resistance to stavudine. Antimicrob Agents Chemother. 2001;45:2144–2146. doi: 10.1128/AAC.45.7.2144-2146.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Little SJ. Transmission and prevalence of HIV resistance among treatment-naive subjects. Antivir Ther. 2000;5:33–40. doi: 10.1177/135965350000500111. [DOI] [PubMed] [Google Scholar]
- 170.Little SJ, Daar ES, D’Aquila RT, Keiser PH, Connick E, Whitcomb JM, Hellmann NS, Petropoulos CJ, Sutton L, Pitt JA, Rosenberg ES, Koup RA, Walker BD, Richman DD. Reduced antiretroviral drug susceptibility among patients with primary HIV infection. JAMA. 1999;282:1142–1149. doi: 10.1001/jama.282.12.1142. [DOI] [PubMed] [Google Scholar]
- 171.Liu SL, Schacker T, Musey L, Shriner D, McElrath MJ, Corey L, Mullins JI. Divergent patterns of progression to AIDS after infection from the same source: human immunodeficiency virus type 1 evolution and antiviral responses. J Virol. 1997;71:4284–95. doi: 10.1128/jvi.71.6.4284-4295.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Long EM, Martin HL, Jr, Kreiss JK, Rainwater SM, Lavreys L, Jackson DJ, Rakwar J, Mandaliya K, Overbaugh J. Gender differences in HIV-1 diversity at time of infection. Nat Med. 2000;6:71–5. doi: 10.1038/71563. [DOI] [PubMed] [Google Scholar]
- 173.Loveday C. International perspectives on antiretroviral resistance. Nucleoside reverse transcriptase inhibitor resistance. J Acquir Immune Defic Syndr. 2001;26(Suppl 1):S10–24. doi: 10.1097/00042560-200103011-00003. [DOI] [PubMed] [Google Scholar]
- 174.Lukashov VV, Kuiken CL, Goudsmit J. Intrahost human immunodeficiency virus type 1 evolution is related to length of the immunocompetent period. J Virol. 1995;69:6911–6916. doi: 10.1128/jvi.69.11.6911-6916.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Mahalingam B, Louis JM, Reed CC, Adomat JM, Krouse J, Wang YF, Harrison RW, Weber IT. Structural and kinetic analysis of drug resistant mutants of HIV-1 protease. Eur J Biochem. 1999;263:238–45. doi: 10.1046/j.1432-1327.1999.00514.x. [DOI] [PubMed] [Google Scholar]
- 176.Mammano F, Petit C, Clavel F. Resistance-associated loss of viral fitness in human immunodeficiency virus type 1: phenotypic analysis of protease and gag coevolution in protease inhibitor-treated patients. J Virol. 1998;72:7632–7. doi: 10.1128/jvi.72.9.7632-7637.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Mansky LM. Retrovirus mutation rates and their role in genetic variation. J Gen Virol. 1998;79:1337–1345. doi: 10.1099/0022-1317-79-6-1337. [DOI] [PubMed] [Google Scholar]
- 178.Marlowe N, Dileanis J, Elbeik T. Evaluation of Applied Biosystems ViroSeq HIV-1 Genotyping System Version 2 for the detection of drug resistance mutations in research clinical plasma samples (TuPeA3006). XIII International AIDS Conference; Durban, South Africa. 2000. [Google Scholar]
- 179.Masquelier B, Descamps D, Carriere I, Ferchal F, Collin G, Denayrolles M, Ruffault A, Chanzy B, Izopet J, Buffet-Janvresse C, Schmitt MP, Race E, Fleury HJ, Aboulker JP, Yeni P, Brun-Vezinet F. Zidovudine resensitization and dual HIV-1 resistance to zidovudine and lamivudine in the delta lamivudine roll-over study. Antivir Ther. 1999;4:69–77. [PubMed] [Google Scholar]
- 180.Mayers D, Merigan T, Gilbert P. T215Y/F mutation associated with zidovudine (ZDV) resistance leads to poor response to ZDV+ddI or ZDV+ddI+NVP: ACTG244/RV79 [abstract 129]. 6th Conference on Retroviruses and Opportunistic Infections; Chicago, U.S. 1999. [Google Scholar]
- 181.Melnick D, Rosenthal J, Cameron M, Snyder M, Griffith-Howard S, Hertogs K, Verbiest W, Graham N, Pham S. Impact of phenotypic antiretroviral drug resistance testing on the response to salvage antiretroviral therapy (ART) in heavily experienced patients [abstract 786]. 7th Conference on Retroviruses and Opportunistic Infections; San Francisco, U.S. 2000. Abstract 786. [Google Scholar]
- 182.Meyer PR, Matsuura SE, Mian AM, So AG, Scott WA. A mechanism of AZT resistance: an increase in nucleotide-dependent primer unblocking by mutant HIV-1 reverse transcriptase. Mol Cell. 1999;4:35–43. doi: 10.1016/s1097-2765(00)80185-9. [DOI] [PubMed] [Google Scholar]
- 183.Meyer PR, Matsuura SE, Schinazi RF, So AG, Scott WA. Differential removal of thymidine nucleotide analogues from blocked DNA chains by human immunodeficiency virus reverse transcriptase in the presence of physiological concentrations of 2¢-deoxynucleoside triphosphates. Antimicrob Agents Chem other. 2000a;44:3465–72. doi: 10.1128/aac.44.12.3465-3472.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Meyer PR, Matsuura SE, So AG, Scott WA. Unblocking of chain-terminated primer by HIV-1 reverse transcriptase through a nucleotide-dependent mechanism. Proc Natl Acad Sci USA. 1998;95:13471–13476. doi: 10.1073/pnas.95.23.13471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Meyer PR, Pfeifer I, Matsuura SE, Bazmi H, So AG, Mellors JW, Scott WA. Effects of M41L and T215Y mutations in HIV-1 reverse transcriptase on removal of chain-terminators from blocked primer/templates. Antiviral Therapy. 2000b;5(Supplement 3):14. Abstract 15. [Google Scholar]
- 186.Meyerhans A, Cheynier R, Albert J, Seth M, Kwok S, Sninsky J, Morfeldt-Manson L, Asjo B, Wain-Hobson S. Temporal fluctuations in HIV quasispecies in vivo are not reflected by sequential HIV isolations. Cell. 1989;58:901–10. doi: 10.1016/0092-8674(89)90942-2. [DOI] [PubMed] [Google Scholar]
- 187.Meynard JL, Vray M, Monard-Joubert L, Matheron S, Peytavin G, Clavel F, Brun-Vezinet F, Girard PM. Impact of treatment guided by phenotypic or genotypic resistance tests on the response to antiretroviral therapy (ART): a randomized trial (NARVAL, ANRS 088) [abstract 698]. 40th Interscience Conference on Antimicrobial Agents and Chemotherapy; Toronto, Canada. 2000. p. 294. [Google Scholar]
- 188.Miller V. Resistance to protease inhibitors. J Acquir Immune Defic Syndr. 2001;26(Suppl 1):S34–50. doi: 10.1097/00042560-200103011-00005. [DOI] [PubMed] [Google Scholar]
- 189.Molla A, Korneyeva M, Gao Q, Vasavanonda S, Schipper PJ, Mo HM, Markowitz M, Chernyavskiy T, Niu P, Lyons N, Hsu A, Granneman GR, Ho DD, Boucher CA, Leonard JM, Norbeck DW, Kempf DJ. Ordered accumulation of mutations in HIV protease confers resistance to ritonavir. Nat Med. 1996;2:760–766. doi: 10.1038/nm0796-760. [DOI] [PubMed] [Google Scholar]
- 190.Montaner JS, Mo T, Raboud JM, Rae S, Alexander CS, Zala C, Rouleau D, Harrigan PR. Human immunodeficiency virus-infected persons with mutations conferring resistance to zidovudine show reduced virologic responses to hydroxyurea and stavudine-lamivudine. J Infect Dis. 2000;181:729–32. doi: 10.1086/315243. [DOI] [PubMed] [Google Scholar]
- 191.Mulder J, Resnick R, Saget B, Scheibel S, Herman S, Payne H, Harrigan R, Kwok S. A rapid and simple method for extracting human immunodeficiency virus type 1 RNA from plasma: enhanced sensitivity. J Clin Microbiol. 1997;35:1278–80. doi: 10.1128/jcm.35.5.1278-1280.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Myers T, Birch D, Bodepudi V, Fong D, Gelfand D, LAK, Nersesian R, Shahinian R, Sigua C, Schonbrunner N, Resnick R, Wu K, Ryder T. Microarray-based HIV-1 drug resistance/genotyping assay [abstract 49] Antivir Ther. 2000;5 (Supplement 3):172. [Google Scholar]
- 193.Najera I, Holguin A, Quinones-Mateu ME, Munoz-Fernandez MA, Najera R, Lopez-Galindez C, Domingo E. Pol gene quasispecies of human immunodeficiency virus: mutations associated with drug resistance in virus from patients undergoing no drug therapy. J Virol. 1995;69:23–31. doi: 10.1128/jvi.69.1.23-31.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Nei M, Gojobori T. Simple methods for estimating the numbers of synonymous and nonsynonymous nucleotide substitutions. Mol Biol Evol. 1986;3:418–26. doi: 10.1093/oxfordjournals.molbev.a040410. [DOI] [PubMed] [Google Scholar]
- 195.Niubo J, Li W, Henry K, Erice A. Recovery and analysis of human immunodeficiency virus type 1 (HIV) RNA sequences from plasma samples with low HIV RNA levels. J Clin Microbiol. 2000;38:309–12. doi: 10.1128/jcm.38.1.309-312.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Notermans DW, Jurriaans S, de WF, Foudraine NA, de JJ, Cavert W, Schuwirth CM, Kauffmann RH, Meenhorst PL, McDade H, Goodwin C, Leonard JM, Goudsmit J, Danner SA. Decrease of HIV-1 RNA levels in lymphoid tissue and peripheral blood during treatment with ritonavir, lamivudine and zidovudine. Ritonavir/3TC/ZDV Study Group. AIDS. 1998;12:167–173. doi: 10.1097/00002030-199802000-00006. [DOI] [PubMed] [Google Scholar]
- 197.Ouellette BF. The GenBank sequence database. In: Baxevanis AD, Fouellette B, editors. BioInformatics: A Practical Guide to the Analysis of Genes and Proteins. Wiley-Interscience; New York: 1998. pp. 16–34. [Google Scholar]
- 198.Page RD. TreeView: an application to display phylogenetic trees on personal computers. Comput Appl Biosci. 1996;12:357–8. doi: 10.1093/bioinformatics/12.4.357. [DOI] [PubMed] [Google Scholar]
- 199.Palmer S, Alaeus A, Albert J, Cox S. Drug susceptibility of subtypes A, B, C, D, and E human immunodeficiency virus type 1 primary isolates. AIDS Res Hum Retroviruses. 1998;14:157–162. doi: 10.1089/aid.1998.14.157. [DOI] [PubMed] [Google Scholar]
- 200.Pang S, Shlesinger Y, Daar ES, Moudgil T, Ho DD, Chen IS. Rapid generation of sequence variation during primary HIV-1 infection. Aids. 1992;6:453–60. doi: 10.1097/00002030-199205000-00003. [DOI] [PubMed] [Google Scholar]
- 201.Pantaleo G, Cohen OJ, Schacker T, Vaccarezza M, Graziosi C, Rizzardi GP, Kahn J, Fox CH, Schnittman SM, Schwartz DH, Corey L, Fauci AS. Evolutionary pattern of human immunodeficiency virus (HIV) replication and distribution in lymph nodes following primary infection: implications for antiviral therapy. Nat Med. 1998;4:341–345. doi: 10.1038/nm0398-341. [DOI] [PubMed] [Google Scholar]
- 202.Panther LA, Tucker L, Xu C, Tuomala RE, Mullins JI, Anderson DJ. Genital tract human immunodeficiency virus type 1 (HIV-1) shedding and inflammation and HIV-1 env diversity in perinatal HIV-1 transmission. J Infect Dis. 2000;181:555–63. doi: 10.1086/315230. [DOI] [PubMed] [Google Scholar]
- 203.Para MF, Glidden DV, Coombs R, Collier A, Condra J, Craig C, Bassett R, Leavitt R, Snyder S, McAuliffe VJ, Boucher C. Baseline human immunodeficiency virus type I phenotype, genotype, and RNA response after switching from long-term hard-capsule saquinavir to indinavir or soft-gel-capsule saquinavir in AIDS clinical trials group protocol 333. J Infect Dis. 2000;182:733–43. doi: 10.1086/315769. [DOI] [PubMed] [Google Scholar]
- 204.Parkin NT, Chappey C, Maranta M, Whitehurst N, Petropoulos CJ. Genotypic and phenotypic analysis of a large database of patient samples reveals distinct patterns of protease inhibitor cross-resistance [abstract 65] Antivir Ther. 2001;6(Suppl 1):49–50. [Google Scholar]
- 205.Partaledis JA, Yamaguchi K, Tisdale M, Blair EE, Falcione C, Maschera B, Myers RE, Pazhanisamy S, Futer O, Cullinan AB, et al. In vitro selection and characterization of human immunodeficiency virus type 1 (HIV-1) isolates with reduced sensitivity to hydroxyethylamino sulfonamide inhibitors of HIV-1 aspartyl protease. J Virol. 1995;69:5228–35. doi: 10.1128/jvi.69.9.5228-5235.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Patick AK, Duran M, Cao Y, Shugarts D, Keller MR, Mazabel E, Knowles M, Chapman S, Kuritzkes DR, Markowitz M. Genotypic and phenotypic characterization of human immunodeficiency virus type 1 variants isolated from patients treated with the protease inhibitor nelfinavir. Antimicrob Agents Chem other. 1998;42:2637–2644. doi: 10.1128/aac.42.10.2637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Patick AK, Mo H, Markowitz M, Appelt K, Wu B, Musick L, Kalish V, Kaldor S, Reich S, Ho D, Webber S. Antiviral and resistance studies of AG1343, an orally bioavailable inhibitor of human immunodeficiency virus protease. Antimicrob Agents Chem other. 1996;40:292–297. doi: 10.1128/aac.40.2.292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Pelemans H, Esnouf RM, Parniak MA, Vandamme AM, De Clercq E, Balzarini J. A proline-to-histidine substitution at position 225 of human immunodeficiency virus type 1 (HIV-1) reverse transcriptase (RT) sensitizes HIV-1 RT to BHAP U-90152. J Gen Virol. 1998;79:1347–52. doi: 10.1099/0022-1317-79-6-1347. [DOI] [PubMed] [Google Scholar]
- 209.Pellegrin I, Izopet J, Reynes J, Denayrolles M, Montes B, Pellegrin JL, Massip P, Puel J, Fleury H, Segondy M. Emergence of zidovudine and multidrug-resistance mutations in the HIV-1 reverse transcriptase gene in therapy-naive patients receiving stavudine plus didanosine combination therapy. STADI Group. AIDS. 1999;13:1705–9. doi: 10.1097/00002030-199909100-00014. [DOI] [PubMed] [Google Scholar]
- 210.Perelson AS, Neumann AU, Markowitz M, Leonard JM, Ho DD. HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time. Science. 1996;271:1582–1586. doi: 10.1126/science.271.5255.1582. [DOI] [PubMed] [Google Scholar]
- 211.Petropoulos CJ, Parkin NT, Limoli KL, Lie YS, Wrin T, Huang W, Tian H, Smith D, Winslow GA, Capon DJ, Whitcomb JM. A novel phenotypic drug susceptibility assay for human immunodeficiency virus type 1. Antimicrob Agents Chem other. 2000;44:920–8. doi: 10.1128/aac.44.4.920-928.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Pieniazek D, Janini LM, Ramos A, Tanuri A, Schechter M, Peralta JM, Vicente AC, Pieniazek NK, Schochetman G, Rayfield MA. HIV-1 patients may harbor viruses of different phylogenetic subtypes: implications for the evolution of the HIV/AIDS pandemic. Emerg Infect Dis. 1995;1:86–88. doi: 10.3201/eid0103.950303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Pieniazek D, Rayfield M, Hu DJ, Nkengasong J, Wiktor SZ, Downing R, Biryahwaho B, Mastro T, Tanuri A, Soriano V, Lal R, Dondero T. Protease sequences from HIV-1 group M subtypes A-H reveal distinct amino acid mutation patterns associated with protease resistance in protease inhibitor-naive individuals worldwide. HIV Variant Working Group. AIDS. 2000;14:1489–95. doi: 10.1097/00002030-200007280-00004. [DOI] [PubMed] [Google Scholar]
- 214.Poss M, Martin HL, Kreiss JK, Granville L, Chohan B, Nyange P, Mandaliya K, Overbaugh J. Diversity in virus populations from genital secretions and peripheral blood from women recently infected with human immunodeficiency virus type 1. J Virol. 1995;69:8118–22. doi: 10.1128/jvi.69.12.8118-8122.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Prado JG, Wrin T, Beauchaine J, Ruiz L, Petropoulos CJ, Clotet B, D’Aquila R, Martinez-Picado J. Lopinavir resistance of amprenavir-selected, replication-impaired mutants of HIV-1 [abstract 67] Antivir Ther. 2001;6(Suppl 1):51. [Google Scholar]
- 216.Puchhammer-Stockl E, Schmied B, Mandl CW, Vetter N, Heinz FX. Comparison of line probe assay (LIPA) and sequence analysis for detection of HIV-1 drug resistance. J Med Virol. 1999;57:283–9. doi: 10.1002/(sici)1096-9071(199903)57:3<283::aid-jmv12>3.0.co;2-2. [DOI] [PubMed] [Google Scholar]
- 217.Quinones-Mateu ME, Albright JL, Mas A, Soriano V, Arts EJ. Analysis of pol gene heterogeneity, viral quasispecies, and drug resistance in individuals infected with group O strains of human immunodeficiency virus type 1. J Virol. 1998;72:9002–15. doi: 10.1128/jvi.72.11.9002-9015.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Quinones-Mateu ME, Holguin A, Dopazo J, Najera I, Domingo E. Point mutant frequencies in the pol gene of human immunodeficiency virus type 1 are two- to threefold lower than those of env. AIDS Res Hum Retroviruses. 1996;12:1117–1128. doi: 10.1089/aid.1996.12.1117. [DOI] [PubMed] [Google Scholar]
- 219.Quinones-Mateu ME, Soriano V, Domingo E, Menendez-Arias L. Characterization of the reverse transcriptase of a human immunodeficiency virus type 1 group O isolate. Virology. 1997;236:364–373. doi: 10.1006/viro.1997.8748. [DOI] [PubMed] [Google Scholar]
- 220.Ramratnam B, Mittler JE, Zhang L, Boden D, Hurley A, Fang F, Macken CA, Perelson AS, Markowitz M, Ho DD. The decay of the latent reservoir of replication-competent HIV-1 is inversely correlated with the extent of residual viral replication during prolonged anti-retroviral therapy. Nat Med. 2000;6:82–5. doi: 10.1038/71577. [DOI] [PubMed] [Google Scholar]
- 221.Ren J, Esnouf RM, Hopkins AL, Jones EY, Kirby I, Keeling J, Ross CK, Larder BA, Stuart DI, Stammers DK. 3¢-Azido-3¢-deoxythymidine drug resistance mutations in HIV-1 reverse transcriptase can induce long range conformational changes. Proc Natl Acad Sci U S A. 1998;95:9518–23. doi: 10.1073/pnas.95.16.9518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Rey D, Hughes M, Pi JT, Winters M, Merigan TC, Katzenstein DA. HIV-1 reverse transcriptase codon 215 mutation in plasma RNA: immunologic and virologic responses to zidovudine. The AIDS Clinical Trials Group Study 175 Virology Team. J Acquir Immune Defic Syndr Hum Retrovirol. 1998;17:203–208. doi: 10.1097/00042560-199803010-00003. [DOI] [PubMed] [Google Scholar]
- 223.Ribeiro RM, Bonhoeffer S. Production of resistant HIV mutants during antiretroviral therapy. Proc Natl Acad Sci U S A. 2000;97:7681–6. doi: 10.1073/pnas.97.14.7681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Richman DD. Principles of HIV resistance testing and overview of assay performance characteristics. Antivir Ther. 2000;5:27–31. [PubMed] [Google Scholar]
- 225.Rodrigo AG. HIV evolutionary genetics. Proc Natl Acad Sci U S A. 1999;96:10559–61. doi: 10.1073/pnas.96.19.10559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Rousseau MN, Vergne L, Montes B, Peeters M, Reynes J, Delaporte E, Segondy M. Patterns of resistance mutations to antiretroviral drugs in extensively treated HIV-1-infected patients with failure of highly active antiretroviral therapy. J Acquir Immune Defic Syndr. 2001;26:36–43. doi: 10.1097/00126334-200101010-00005. [DOI] [PubMed] [Google Scholar]
- 227.Rouzine IM, Coffin JM. Linkage disequilibrium test implies a large effective population number for HIV in vivo. Proc Natl Acad Sci U S A. 1999;96:10758–63. doi: 10.1073/pnas.96.19.10758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Ruiz L, Telles F, Smith M, Maillard A, Domingo C, Heiken H, Holtzer C. Inter-laboratory and inter-lot reproducibility and quality control of HIV-1 genotyping with a commercial kit [abstract 55] Antivir Ther. 2000;5:176–177. [Google Scholar]
- 229.Salomon H, Wainberg MA, Brenner B, Quan Y, Rouleau D, Cote P, LeBlanc R, Lefebvre E, Spira B, Tsoukas C, Sekaly RP, Conway B, Mayers D, Routy JP. Prevalence of HIV-1 resistant to antiretroviral drugs in 81 individuals newly infected by sexual contact or injecting drug use. Investigators of the Quebec Primary Infection Study. AIDS. 2000;14:F17–23. doi: 10.1097/00002030-200001280-00003. [DOI] [PubMed] [Google Scholar]
- 230.Sambrook S, Russell DW. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY: 2001. [Google Scholar]
- 231.Sarafianos SG, Das K, Clark AD, Jr, Ding J, Boyer PL, Hughes SH, Arnold E. Lamivudine (3TC) resistance in HIV-1 reverse transcriptase involves steric hindrance with beta-branched amino acids. Proc Natl Acad Sci U S A. 1999a;96:10027–32. doi: 10.1073/pnas.96.18.10027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Sarafianos SG, Das K, Ding J, Boyer PL, Hughes SH, Arnold E. Touching the heart of HIV-1 drug resistance: the fingers close down on the dNTP at the polymerase active site. Chem Biol. 1999b;6:R137–46. doi: 10.1016/s1074-5521(99)80071-4. [DOI] [PubMed] [Google Scholar]
- 233.Schapiro JM, Winters MA, Vierra M, Crawford S, Merigan TC. Lymph node human immunodeficiency virus RNA levels and resistance mutations in patients receiving high-dose saquinavir. J Infect Dis. 1998;177:477–80. doi: 10.1086/517380. [DOI] [PubMed] [Google Scholar]
- 234.Schmit JC, Van LK, Ruiz L, Hermans P, Sprecher S, Sonnerborg A, Leal M, Harrer T, Clotet B, Arendt V, Lissen E, Witvrouw M, Desmyter J, De CE, Vandamme AM. Multiple dideoxynucleoside analogue-resistant (MddNR) HIV-1 strains isolated from patients from different European countries. AIDS. 1998;12:2007–2015. doi: 10.1097/00002030-199815000-00012. [DOI] [PubMed] [Google Scholar]
- 235.Schuurman R, Brambilla DJ, de Groot T, Boucher C. Second worldwide evaluation of HIV-1 drug resistance genotyping quality, using the ENVA-2 panel [abstract 1168]. 39th Interscience Conference on Antimicrobial Agents and Chemotherapy; 1999a. p. 493. [Google Scholar]
- 236.Schuurman R, Demeter L, Reichelderfer P, Tijnagel J, de Groot T, Boucher C. Worldwide evaluation of DNA sequencing approaches for identification of drug resistance mutations in the human immunodeficiency virus type 1 reverse transcriptase. J Clin Microbiol. 1999b;37:2291–2296. doi: 10.1128/jcm.37.7.2291-2296.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Schuurman R, Nijhuis M, van Leeuwen R, Schipper P, de Jong D, Collis P, Danner SA, Mulder J, Loveday C, Christopherson C. Rapid changes in human immunodeficiency virus type 1 RNA load and appearance of drug-resistant virus populations in persons treated with lamivudine (3TC) J Infect Dis. 1995;171:1411–1419. doi: 10.1093/infdis/171.6.1411. [DOI] [PubMed] [Google Scholar]
- 238.Servais J, Lambert C, Fontaine E, Plesseria JM, Robert I, Arendt V, Staub T, Schneider F, Hemmer R, Burtonboy G, Schmit JC. Comparison of DNA Sequencing and a Line Probe Assay for Detection of Human Immunodeficiency Virus Type 1 Drug Resistance Mutations in Patients Failing Highly Active Antiretroviral Therapy. J Clin Microbiol. 2001a;39:454–459. doi: 10.1128/JCM.39.2.454-459.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Servais J, Lambert C, Fontaine E, Plesseria JM, Robert I, Arendt V, Staub T, Schneider F, Hemmer R, Burtonboy G, Schmit JC. Variant human immunodeficiency virus type 1 proteases and response to combination therapy including a protease inhibitor. Antimicrob Agents Chem other. 2001b;45:893–900. doi: 10.1128/AAC.45.3.893-900.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Shafer RW, Chuang TK, Hsu P, White CB, Katzenstein DA. Sequence and drug susceptibility of subtype C protease from human immunodeficiency virus type 1 seroconverters in Zimbabwe. AIDS Res Hum Retroviruses. 1999a;15:65–9. doi: 10.1089/088922299311727. [DOI] [PubMed] [Google Scholar]
- 241.Shafer RW, Eisen JA, Merigan TC, Katzenstein DA. Sequence and drug susceptibility of subtype C reverse transcriptase from human immunodeficiency virus type 1 seroconverters in Zimbabwe. J Virol. 1997;71:5441–8. doi: 10.1128/jvi.71.7.5441-5448.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Shafer RW, Hertogs K, Zolopa AR, Warford A, Bloor S, Betts BJ, Merigan TC, Harrigan R, Larder BA. High degree of interlaboratory reproducibility of human immunodeficiency virus type 1 protease and reverse transcriptase sequencing of plasma samples from heavily treated patients. J Clin Microbiol. 2001;39:1522–9. doi: 10.1128/JCM.39.4.1522-1529.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Shafer RW, Hsu P, Patick AK, Craig C, Brendel V. Identification of biased amino acid substitution patterns in human immunodeficiency virus type 1 isolates from patients treated with protease inhibitors. J Virol. 1999b;73:6197–202. doi: 10.1128/jvi.73.7.6197-6202.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Shafer RW, Jung DR, Betts BJ. Human immunodeficiency virus type 1 reverse transcriptase and protease mutation search engine for queries. Nat Med. 2000a;6:1290–1292. doi: 10.1038/81407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Shafer RW, Kantor R, Gonzales MJ. The genetic basis of HIV-1 resistance to reverse transcriptase and protease inhibitors. AIDS Rev. 2000b;2:211–218. [PMC free article] [PubMed] [Google Scholar]
- 246.Shafer RW, Kozal MJ, Winters MA, Iversen AK, Katzenstein DA, Ragni MV, Meyer WA, Gupta P, Rasheed S, Coombs R, Merigan TC. Combination therapy with zidovudine and didanosine selects for drug- resistant human immunodeficiency virus type 1 strains with unique patterns of pol gene mutations. J Infect Dis. 1994;169:722–729. doi: 10.1093/infdis/169.4.722. [DOI] [PubMed] [Google Scholar]
- 247.Shafer RW, Warford A, Winters MA, Gonzales MJ. Reproducibility of human immunodeficiency virus type 1 (HIV-1) protease and reverse transcriptase sequencing of plasma samples from heavily treated HIV-1-infected individuals. J Virol Methods. 2000c;86:143–153. doi: 10.1016/s0166-0934(00)00144-0. [DOI] [PubMed] [Google Scholar]
- 248.Shafer RW, Winters MA, Mayers DL, Japour AJ, Kuritzkes DR, Weislow OS, White F, Erice A, Sannerud KJ, Iversen A, Pena F, Dimitrov D, Frenkel LM, Reichelderfer PS. Interlaboratory comparison of sequence-specific PCR and ligase detection reaction to detect a human immunodeficiency virus type 1 drug resistance mutation. The AIDS Clinical Trials Group Virology Committee Drug Resistance Working Group. J Clin Microbiol. 1996;34:1849–1853. doi: 10.1128/jcm.34.7.1849-1853.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Shafer RW, Winters MA, Palmer S, Merigan TC. Multiple concurrent reverse transcriptase and protease mutations and multidrug resistance of HIV-1 isolates from heavily treated patients. Ann Intern Med. 1998;128:906–911. doi: 10.7326/0003-4819-128-11-199806010-00008. [DOI] [PubMed] [Google Scholar]
- 250.Sham HL, Kempf DJ, Molla A, Marsh KC, Kumar GN, Chen CM, Kati W, Stewart K, Lal R, Hsu A, Betebenner D, Korneyeva M, Vasavanonda S, McDonald E, Saldivar A, Wideburg N, Chen X, Niu P, Park C, Jayanti V, Grabowski B, Granneman GR, Sun E, Japour AJ, Leonard JM. ABT-378, a highly potent inhibitor of the human immunodeficiency virus protease. Antimicrob Agents Chem other. 1998;42:3218–3224. doi: 10.1128/aac.42.12.3218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Shankarappa R, Margolick JB, Gange SJ, Rodrigo AG, Upchurch D, Farzadegan H, Gupta P, Rinaldo CR, Learn GH, He X, Huang XL, Mullins JI. Consistent viral evolutionary changes associated with the progression of human immunodeficiency virus type 1 infection. J Virol. 1999;73:10489–502. doi: 10.1128/jvi.73.12.10489-10502.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Shirasaka T, Chokekijchai S, Yamada A, Gosselin G, Imbach JL, Mitsuya H. Comparative analysis of anti-human immunodeficiency virus type 1 activities of dideoxynucleoside analogs in resting and activated peripheral blood mononuclear cells. Antimicrob Agents Chem other. 1995a;39:2555–2559. doi: 10.1128/aac.39.11.2555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Shirasaka T, Kavlick MF, Ueno T, Gao WY, Kojima E, Alcaide ML, Chokekijchai S, Roy BM, Arnold E, Yarchoan R. Emergence of human immunodeficiency virus type 1 variants with resistance to multiple dideoxynucleosides in patients receiving therapy with dideoxynucleosides. Proc Natl Acad Sci USA. 1995b;92:2398–2402. doi: 10.1073/pnas.92.6.2398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Shulman N, Zolopa A, King H, Whitcomb J, Israelski D, Shafer R, Katzenstein D, Petropoulos C, Hellmann N. NNRTI hypersensitivity in a well-characterized cohort of treatment experienced patients receiving efavirenz and adefovir based salvage regimens. Antivir Ther. 2000;5(Supplement 3):78. Abstract 99. [Google Scholar]
- 255.Shulman NS, Shafer R, Winters M, Machekano R, Liou SH, Hughes M, Katzenstein D. Genotypic predictors of virologic response to stavudine after zidovudine monotherapy (ACTG 302) [abstract 437]. 8th Conference on Human Retroviruses and Opportunistic Infections; Chicago, U.S. 2001. [DOI] [PubMed] [Google Scholar]
- 256.Shulman NS, Zolopa AR, Passaro DJ, Murlidharan U, Israelski DM, Brosgart CL, Miller MD, Van Doren S, Shafer RW, Katzenstein DA. Efavirenz- and adefovir dipivoxil-based salvage therapy in highly treatment-experienced patients: clinical and genotypic predictors of virologic response. J Acquir Immune Defic Syndr. 2000;23:221–6. doi: 10.1097/00126334-200003010-00002. [DOI] [PubMed] [Google Scholar]
- 257.Simmonds P, Zhang LQ, McOmish F, Balfe P, Ludlam CA, Brown AJ. Discontinuous sequence change of human immunodeficiency virus (HIV) type 1 env sequences in plasma viral and lymphocyte-associated proviral populations in vivo: implications for models of HIV pathogenesis. J Virol. 1991;65:6266–76. doi: 10.1128/jvi.65.11.6266-6276.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Si-Mohamed A, Kazatchkine MD, Heard I, Goujon C, Prazuck T, Aymard G, Cessot G, Kuo YH, Bernard MC, Diquet B, Malkin JE, Gutmann L, Belec L. Selection of drug-resistant variants in the female genital tract of human immunodeficiency virus type 1-infected women receiving antiretroviral therapy. J Infect Dis. 2000;182:112–22. doi: 10.1086/315679. [DOI] [PubMed] [Google Scholar]
- 259.Simon V, Vanderhoeven J, Hurley A, Ramratnam B, Boden D, Louie M, Kost R, Dawson K, Parkin N, Routy JP, Sekaly RP, Markowitz M. Prevalence of drug-resistant HIV-1 variants in newly infected individuals during 1999–2000. 8th Conference on Retroviruses and Opportunistic Infections.2001. [Google Scholar]
- 260.Smith MS, Koerber KL, Pagano JS. Zidovudine-resistant human immunodeficiency virus type 1 genomes detected in plasma distinct from viral genomes in peripheral blood mononuclear cells. J Infect Dis. 1993;167:445–448. doi: 10.1093/infdis/167.2.445. [DOI] [PubMed] [Google Scholar]
- 261.Snowden W, Shortino D, Klein A, Harris W, Manohitharajah V, Elston R, Tisdale M, Maguire M. Development of amprenavir resistance in NRTI-experienced patients: alternative mechanisms and correlation with baseline resistance to concomitant NRTIs [abstract 108] Antivir Ther. 2000;5(Supplement 3):84. [Google Scholar]
- 262.Spence RA, Kati WM, Anderson KS, Johnson KA. Mechanism of inhibition of HIV-1 reverse transcriptase by nonnucleoside inhibitors. Science. 1995;267:988–93. doi: 10.1126/science.7532321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.St Clair M, Martin JL, Tudor-Williams G, Bach MC, Vavro CL, King DM, Kellam P, Kemp SD, Larder BA. Resistance to ddI and sensitivity to AZT induced by a mutation in HIV-1 reverse transcriptase. Science. 1991;253:1557–1559. doi: 10.1126/science.1716788. [DOI] [PubMed] [Google Scholar]
- 264.Stammers DK, Ren J, Nichols C, Chamberlain P, Douglas L, Lennerstrand J, Larder B, Stuart DI. Crystal structures of HIV-1 reverse transcriptase resistant to zidovudine or lamivudine containing mutations at 41, 184, and 215 [abstract 72] Antivr Ther. 2001;6(Suppl1):54–5. doi: 10.1128/JVI.76.19.10015-10019.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Stuyver L, Wyseur A, Rombout A, Louwagie J, Scarcez T, Verhofstede C, Rimland D, Schinazi RF, Rossau R. Line probe assay for rapid detection of drug-selected mutations in the human immunodeficiency virus type 1 reverse transcriptase gene. Antimicrob Agents Chem other. 1997;41:284–291. doi: 10.1128/aac.41.2.284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266.Swofford DL. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4. Sinauer Associates; Sunderland, Massachusetts: 1998. [Google Scholar]
- 267.Tamalet C, Pasquier C, Yahi N, Colson P, Poizot-Martin I, Lepeu G, Gallais H, Massip P, Puel J, Izopet J. Prevalence of drug resistant mutants and virological response to combination therapy in patients with primary HIV-1 infection. J Med Virol. 2000;61:181–6. doi: 10.1002/(sici)1096-9071(200006)61:2<181::aid-jmv2>3.0.co;2-t. [DOI] [PubMed] [Google Scholar]
- 268.Tamalet C, Izopet J, Koch N, Fantini J, Yahi N. Stable rearrangements of the beta3-beta4 hairpin loop of HIV-1 reverse transcriptase in plasma viruses from patients receiving combination therapy. AIDS. 1998;12:F161–6. doi: 10.1097/00002030-199814000-00003. [DOI] [PubMed] [Google Scholar]
- 269.Tanuri A, Vicente AC, Otsuki K, Ramos CA, Ferreira OC, Jr, Schechter M, Janini LM, Pieniazek D, Rayfield MA. Genetic variation and susceptibilities to protease inhibitors among subtype B and F isolates in Brazil. Antimicrob Agents Chem other. 1999;43:253–8. doi: 10.1128/aac.43.2.253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 270.Tisdale M, Alnadaf T, Cousens D. Combination of mutations in human immunodeficiency virus type 1 reverse transcriptase required for resistance to the carbocyclic nucleoside 1592U89. Antimicrob Agents Chem other. 1997;41:1094–1098. doi: 10.1128/aac.41.5.1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Tisdale M, Kemp SD, Parry NR, Larder BA. Rapid in vitro selection of human immunodeficiency virus type 1 resistant to 3¢-thiacytidine inhibitors due to a mutation in the YMDD region of reverse transcriptase. Proc Natl Acad Sci USA. 1993;90:5653–5656. doi: 10.1073/pnas.90.12.5653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272.Tural C, Ruiz L, Holtzer C, Viciana P, Gonzalez J, Ferrer E, Martinez-Picado J, Ruiz I, Dalmau D, Domingo P, Been A, Boucher C, Schapiro J, Romeu J, Sirera G, Clotet B. The potential role of resistance decision support software with or without expert advice in a trial of HIV genotyping versus standard of care - the Havanna trial [Abstract L-10]. 40th Interscience Conference on Antimicrobial Agents and Chemotherapy; Toronto, Canada. 2000. [Google Scholar]
- 273.UK Collaborative Group on Monitoring the Transmission of HIV Drug Resistance. Analysis of prevalence of HIV-1 drug resistance in primary infections in the United Kingdom. BMJ. 2001;322:1087–1088. doi: 10.1136/bmj.322.7294.1087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274.US Department of Health and Human Services Panel on Clinical Practices for Treatment of HIV Infection. Guidelines for the use of antiretroviral agents in HIV-1-infected adults and adolescents. The living document. 2000 January 28; doi: 10.1310/hct.2000.1.1.008. 2000. http://www.hivatis.org/trtgdlns.html. [DOI] [PubMed]
- 275.Ushiro-Lamb I, Norman J, Skinner C, Murphy M, Breuer J, Aitken C. Does HIV-1 subtype influence genotypic changes associated with ant-retroviral failure? AIDS. 2000;14 (suppl 4):S121. [Google Scholar]
- 276.Vahey M, Nau ME, Barrick S, Cooley JD, Sawyer R, Sleeker AA, Vickerman P, Bloor S, Larder B, Michael NL, Wegner SA. Performance of the Affymetrix GeneChip HIV PRT 440 platform for antiretroviral drug resistance genotyping of human immunodeficiency virus type 1 clades and viral isolates with length polymorphisms. J Clin Microbiol. 1999;37:2533–7. doi: 10.1128/jcm.37.8.2533-2537.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 277.Van de Peer Y, De Wachter R. Construction of evolutionary distance trees with TREECON for Windows: accounting for variation in nucleotide substitution rate among sites. Comput Appl Biosci. 1997;13:227–30. doi: 10.1093/bioinformatics/13.3.227. [DOI] [PubMed] [Google Scholar]
- 278.Van Laethem K, Van Vaerenbergh K, Schmit JC, Sprecher S, Hermans P, De Vroey V, Schuurman R, Harrer T, Witvrouw M, Van Wijngaerden E, Stuyver L, Van Ranst M, Desmyter J, De Clercq E, Vandamme AM. Phenotypic assays and sequencing are less sensitive than point mutation assays for detection of resistance in mixed HIV-1 genotypic populations. J Acquir Immune Defic Syndr. 1999;22:107–18. doi: 10.1097/00126334-199910010-00001. [DOI] [PubMed] [Google Scholar]
- 279.Van Laethem K, Witvrouw M, Balzarini J, Schmit JC, Sprecher S, Hermans P, Leal M, Harrer T, Ruiz L, Clotet B, Van Ranst M, Desmyter J, De Clercq E, Vandamme AM. Patient HIV-1 strains carrying the multiple nucleoside resistance mutations are cross-resistant to abacavir [letter] Aids. 2000;14:469–71. doi: 10.1097/00002030-200003100-00027. [DOI] [PubMed] [Google Scholar]
- 280.Venturi G, Catucci M, Romano L, Corsi P, Leoncini F, Valensin PE, Zazzi M. Antiretroviral resistance mutations in human immunodeficiency virus type 1 reverse transcriptase and protease from paired cerebrospinal fluid and plasma samples. J Infect Dis. 2000;181:740–5. doi: 10.1086/315249. [DOI] [PubMed] [Google Scholar]
- 281.Verbiest W, Peeters M, Hertogs K, Schel P, Bloor S, Rinehart A, Graham N, Cohen C, Larder BA. VirtualPhenotype is concordant with actual phenotype: a retrospective analysis of data from VIRA3001 [abstract 81] Antivir Ther. 2000;5 (Suppl 3):62. [Google Scholar]
- 282.Vergne L, Peeters M, Mpoudi-Ngole E, Bourgeois A, Liegeois F, Toure-Kane C, Mboup S, Mulanga-Kabeya C, Saman E, Jourdan J, Reynes J, Delaporte E. Genetic diversity of protease and reverse transcriptase sequences in non-subtype-B human immunodeficiency virus type 1 strains: evidence of many minor drug resistance mutations in treatment-naive patients. J Clin Microbiol. 2000;38:3919–25. doi: 10.1128/jcm.38.11.3919-3925.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283.Verhofstede C, Fransen K, Marissens D, Verhelst R, van der Groen G, Lauwers S, Zissis G, Plum J. Isolation of HIV-1 RNA from plasma: evaluation of eight different extraction methods. J Virol Methods. 1996;60:155–9. doi: 10.1016/0166-0934(96)02062-9. [DOI] [PubMed] [Google Scholar]
- 284.Wang YM, Dyer WB, Workman C, Wang B, Sullivan JS, Saksena NK. Molecular evidence for drug-induced compartmentalization of HIV-1 quasispecies in a patient with periodic changes to HAART. AIDS. 2000;14:2265–72. doi: 10.1097/00002030-200010200-00007. [DOI] [PubMed] [Google Scholar]
- 285.Wei X, Ghosh SK, Taylor ME, Johnson VA, Emini EA, Deutsch P, Lifson JD, Bonhoeffer S, Nowak MA, Hahn BH, Shaw GM. Viral dynamics in human immunodeficiency virus type 1 infection. Nature. 1995;373:117–122. doi: 10.1038/373117a0. [DOI] [PubMed] [Google Scholar]
- 286.Weidle PJ, Kityo CM, Mugyenyi P, Downing R, Kebba A, Pieniazek D, Respess R, Hertogs K, De Vroey V, Dehertogh P, Bloor S, Larder B, Lackritz E. Resistance to antiretroviral therapy among patients in Uganda. J Acquir Immune Defic Syndr. 2001;26:495–500. doi: 10.1097/00126334-200104150-00017. [DOI] [PubMed] [Google Scholar]
- 287.Wilson JW, Bean P, Robins T, Graziano F, Persing DH. Comparative evaluation of three human immunodeficiency virus genotyping systems: the HIV-Genotype Method, the HIV PRT GeneChip Assay, and the HIV-1 RT Line Probe Assay. J Clin Microbiol. 2000;38:3022–3028. doi: 10.1128/jcm.38.8.3022-3028.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 288.Winslow DL, Garber S, Reid C, Scarnati H, Baker D, Rayner MM, Anton ED. Selection conditions affect the evolution of specific mutations in the reverse transcriptase gene associated with resistance to DMP 266. AIDS. 1996;10:1205–1209. doi: 10.1097/00002030-199609000-00005. [DOI] [PubMed] [Google Scholar]
- 289.Winters MA, Coolley KL, Girard YA, Levee DJ, Hamdan H, Shafer RW, Katzenstein DA, Merigan TC. A 6-basepair insert in the reverse transcriptase gene of human immunodeficiency virus type 1 confers resistance to multiple nucleoside inhibitors. J Clin Invest. 1998;102:1769–1775. doi: 10.1172/JCI4948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 290.Winters MA, Schapiro JM, Lawrence J, Merigan TC. Human immunodeficiency virus type 1 protease genotypes and in vitro protease inhibitor susceptibilities of isolates from individuals who were switched to other protease inhibitors after long-term saquinavir treatment. J Virol. 1998a;72:5303–5306. doi: 10.1128/jvi.72.6.5303-5306.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Winters MA, Merigan TC. Variants other than aspartic acid at codon 69 of the HIV-1 reverse transcriptase gene affect susceptibility to nucleoside analogues [abstract 75] Antivir Ther. 2001;6(Suppl 1):56–57. doi: 10.1128/AAC.45.8.2276-2279.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Wolfs TF, Zwart G, Bakker M, Goudsmit J. HIV-1 genomic RNA diversification following sexual and parenteral virus transmission. Virology. 1992;189:103–10. doi: 10.1016/0042-6822(92)90685-i. [DOI] [PubMed] [Google Scholar]
- 293.Wolinsky SM, Korber BT, Neumann AU, Daniels M, Kunstman KJ, Whetsell AJ, Furtado MR, Cao Y, Ho DD, Safrit JT. Adaptive evolution of human immunodeficiency virus-type 1 during the natural course of infection. Science. 1996;272:537–542. doi: 10.1126/science.272.5261.537. [DOI] [PubMed] [Google Scholar]
- 294.Wong JK, Gunthard HF, Havlir DV, Zhang ZQ, Haase AT, Ignacio CC, Kwok S, Emini E, Richman DD. Reduction of HIV-1 in blood and lymph nodes following potent antiretroviral therapy and the virologic correlates of treatment failure. Proc Natl Acad Sci USA. 1997a;94:12574–12579. doi: 10.1073/pnas.94.23.12574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Wong JK, Hezareh M, Gunthard HF, Havlir DV, Ignacio CC, Spina CA, Richman DD. Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science. 1997b;278:1291–1295. doi: 10.1126/science.278.5341.1291. [DOI] [PubMed] [Google Scholar]
- 296.Wong JK, Ignacio CC, Torriani F, Havlir D, Fitch NJ, Richman DD. In vivo compartmentalization of human immunodeficiency virus: evidence from the examination of pol sequences from autopsy tissues. J Virol. 1997c;71:2059–2071. doi: 10.1128/jvi.71.3.2059-2071.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297.Xu R, Andrews W, Spaltenstein A, Danger D, Dallas W, Carter L, Wright L, Furfine E. Molecular mechanism of I50V HIV-1 protease resistance and cross-resistance to protease inhibitors [abstract 466]. 8th Conference on Retroviruses and Opportunistic Infections; Chicago, IL. 2001. [Google Scholar]
- 298.Yahi N, Fantini J, Tourres C, Tivoli N, Koch N, Tamalet C. Use of drug resistance sequence data for the systematic detection of non-B human immunodeficiency virus type 1 (HIV-1) subtypes: How to create a sentinel site for monitoring the genetic diversity of HIV-1 at a country scale. J Infect Dis. 2001;183:1311–17. doi: 10.1086/319859. [DOI] [PubMed] [Google Scholar]
- 299.Yahi N, Tamalet C, Tourres C, Tivoli N, Ariasi F, Volot F, Gastaut JA, Gallais H, Moreau J, Fantini J. Mutation patterns of the reverse transcriptase and protease genes in human immunodeficiency virus type 1-infected patients undergoing combination therapy: survey of 787 sequences. J Clin Microbiol. 1999;37:4099–106. doi: 10.1128/jcm.37.12.4099-4106.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 300.Yerly S, Kaiser L, Race E, Bru JP, Clavel F, Perrin L. Transmission of antiretroviral-drug-resistant HIV-1 variants. Lancet. 1999;354:729–33. doi: 10.1016/S0140-6736(98)12262-6. [DOI] [PubMed] [Google Scholar]
- 301.Yerly S, Rakik A, De Loes SK, Hirschel B, Descamps D, Brun-Vezinet F, Perrin L. Switch to unusual amino acids at codon 215 of the human immunodeficiency virus type 1 reverse transcriptase gene in seroconvertors infected with zidovudine-resistant variants. J Virol. 1998;72:3520–3. doi: 10.1128/jvi.72.5.3520-3523.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 302.Young SD, Britcher SF, Tran LO, Payne LS, Lumma WC, Lyle TA, Huff JR, Anderson PS, Olsen DB, Carroll SS, et al. L-743, 726 (DMP-266): a novel, highly potent nonnucleoside inhibitor of the human immunodeficiency virus type 1 reverse transcriptase. Antimicrob Agents Chem other. 1995;39:2602–5. doi: 10.1128/aac.39.12.2602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 303.Zakeri H, Amparo G, Chen SM, Spurgeon S, Kwok PY. Peak height pattern in dichloro-rhodamine and energy transfer dye terminator sequencing. Biotechniques. 1998;25:406–10. 412–4. doi: 10.2144/98253st01. [DOI] [PubMed] [Google Scholar]
- 304.Zhang D, Caliendo AM, Eron JJ, DeVore KM, Kaplan JC, Hirsch MS, D’Aquila RT. Resistance to 2¢,3¢-dideoxycytidine conferred by a mutation in codon 65 of the human immunodeficiency virus type 1 reverse transcriptase. Antimicrob Agents Chem other. 1994;38:282–287. doi: 10.1128/aac.38.2.282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305.Zhang H, Dornadula G, Beumont M, Livornese LJ, Jr, Van UB, Henning K, Pomerantz RJ. Human immunodeficiency virus type 1 in the semen of men receiving highly active antiretroviral therapy. N Engl J Med. 1998;339:1803–1809. doi: 10.1056/NEJM199812173392502. [DOI] [PubMed] [Google Scholar]
- 306.Zhang L, Chung C, Hu BS, He T, Guo Y, Kim AJ, Skulsky E, Jin X, Hurley A, Ramratnam B, Markowitz M, Ho DD. Genetic characterization of rebounding HIV-1 after cessation of highly active antiretroviral therapy. J Clin Invest. 2000;106:839–45. doi: 10.1172/JCI10565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307.Zhang L, Ramratnam B, Tenner-Racz K, He Y, Vesanen M, Lewin S, Talal A, Racz P, Perelson AS, Korber BT, Markowitz M, Ho DD. Quantifying residual HIV-1 replication in patients receiving combination antiretroviral therapy. N Engl J Med. 1999;340:1605–13. doi: 10.1056/NEJM199905273402101. [DOI] [PubMed] [Google Scholar]
- 308.Zhang YM, Imamichi H, Imamichi T, Lane HC, Falloon J, Vasudevachari MB, Salzman NP. Drug resistance during indinavir therapy is caused by mutations in the protease gene and in its Gag substrate cleavage sites. J Virol. 1997;71:6662–6670. doi: 10.1128/jvi.71.9.6662-6670.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 309.Zhu T, Wang N, Carr A, Wolinsky S, Ho DD. Evidence for coinfection by multiple strains of human immunodeficiency virus type 1 subtype B in an acute seroconvertor. J Virol. 1995;69:1324–7. doi: 10.1128/jvi.69.2.1324-1327.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.Ziermann R, Limoli K, Das K, Arnold E, Petropoulos CJ, Parkin NT. A mutation in human immunodeficiency virus type 1 protease, N88S, that causes in vitro hypersensitivity to amprenavir. J Virol. 2000;74:4414–9. doi: 10.1128/jvi.74.9.4414-4419.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 311.Zolopa AR, Hertogs K, Shafer R, Dehertogh P, De Vroey V, Efron B, Bloor S, Larder B. A comparison of phenotypic, genotypic, and clinical/treatment history predictors of virologic response to saquinavir/ritonavir salvage therapy in a clinic-based cohort [abstract 68] Antivir Ther. 1999a;4(Supplement 1):47–48. [Google Scholar]
- 312.Zolopa AR, Shafer RW, Warford A, Montoya JG, Hsu P, Katzenstein D, Merigan TC, Efron B. HIV-1 genotypic resistance patterns predict response to saquinavir- ritonavir therapy in patients in whom previous protease inhibitor therapy had failed. Ann Intern Med. 1999b;131:813–821. doi: 10.7326/0003-4819-131-11-199912070-00003. [DOI] [PMC free article] [PubMed] [Google Scholar]