

Abstract
Defining the regulatory molecular networks involved in patterning the developing anterior endoderm is essential to understanding how the pancreas, liver, stomach and duodenum are discretely specified from each other. In this study, we analyzed the expression and function of the double-stranded RNA-binding protein Staufen2 in Xenopus laevis endoderm. We found that staufen2 was broadly expressed within the developing endoderm beginning at gastrulation becoming localized to the anterior endoderm at later stages. Through morpholino-mediated knockdown, we demonstrate that Staufen2 function is required for proper formation of the stomach, liver and pancreas. We define that its function is required during gastrulation for proper patterning of the dorsal-ventral axis and that it acts to regulate expression of BMP signaling components.
Keywords: Pancreas, RNA binding protein, Organizer, BMP, morpholino
Introduction
The vertebrate endoderm contributes to the epithelia lining the respiratory and digestive tract and the organs associated with these systems. This germ layer is derived from cells located in the vegetal pole of the early Xenopus embryo (Dale and Slack, 1987). Initiation of its development requires the maternal T domain transcription factor, VegT, which activates expression of many signaling molecules and transcription factors necessary for subsequent endoderm formation (Lustig et al., 1996; Stennard et al., 1996; Zhang and King, 1996; Horb and Thomsen, 1997; Xanthos et al., 2001). To date, many studies have focused on how regional specification of the endoderm is regulated and recent advances have provided a deeper understanding to how signaling molecules and gene regulatory networks control endoderm development (Sinner et al., 2006; McLin et al., 2007; Pan et al., 2007; Pearl et al., 2009; Zorn and Wells, 2009). However, increasing evidence suggests a vital role for post-translational control during differentiation and specification of the endoderm (Mayer and Fishman, 2003; Spagnoli and Brivanlou, 2006; Horb and Horb, 2010).
RNA binding proteins play a vital role during embryogenesis in the regulation of gene expression through translational control, subcellular localization or mRNA stability (Kuersten and Goodwin, 2003). Double-stranded RNA binding proteins (dsRBP) are present in all eukaryotes, suggesting they play a critical function in controlling gene expression within the cell (Saunders and Barber, 2003; Tian et al., 2004). Staufen-related proteins are one family of dsRBPs that have been shown to play crucial roles during development. Originally identified in Drosophila, Staufen was shown to be essential for anterior-posterior axis formation, by acting to localize oskar and bicoid mRNAs to their respective poles (St Johnston et al., 1989; St Johnston et al., 1991; Ferrandon et al., 1994; St Johnston, 1995). It was also shown to participate in embryonic neuroblast differentiation by mediating asymmetric segregation of prospero mRNA (Broadus et al., 1998; Schuldt et al., 1998; Shen et al., 1998).
In vertebrates, there are two Staufen homologs, Staufen1 and Staufen2 that are expressed in a variety of tissues, including primordial germ cells and neural tissue. Mouse staufen1 and staufen2 are expressed in germ cells during oogenesis and embryogenesis (Saunders et al., 2000). Similarly, zebrafish staufen1 and staufen2 are expressed in primordial germ cells and necessary for their proper migration and survival (Ramasamy et al., 2006). Staufen proteins have been identified in RNA-containing granules that co-localize to microtubules and function in mRNA transport within the dendrites (Buchner et al., 1999; Roegiers and Jan, 2000; Macchi et al., 2003; Kanai et al., 2004; Kim et al., 2005a). Staufen1 has also been implicated in a new mRNA decay mechanism (Kim et al., 2005b). In Xenopus, Staufen1 and Staufen2 proteins are vegetally localized in a complex containing Vg1 mRNA, and may regulate proper localization of Vg1 mRNA within the oocyte (Allison et al., 2004; Yoon and Mowry, 2004). In the adult, Staufen1 and 2 expression is largely confined to the testis and ovary, though Staufen2 protein was also detected less abundantly in adult spleen, pancreas and intestine (Allison et al., 2004).
Here we report that Xenopus staufen2 is expressed early in the developing endoderm and that its function was required for proper formation of anterior organ derivatives (pancreas, stomach and liver). We demonstrate that knockdown of Staufen2 inhibited the expression of the BMP antagonist Chordin resulting in early disruption of dorsal-ventral axis specification. Our results suggest that Staufen2 has a unique role in patterning and development of the anterior endoderm during embryogenesis.
Results
Staufen2 is required for anterior endoderm organogenesis
In Xenopus, staufen2 was shown to be expressed almost exclusively in reproductive tissues, but low levels were also detected in pancreas and spleen (Allison et al., 2004). However, whether it was expressed within the endoderm during development was not examined. We initially identified staufen2 as one gene that was down regulated in Ptf1a knockdown embryos, suggesting that it was expressed in the developing endoderm, and we therefore examined its endodermal expression in detail. Initial expression of staufen2 was detected during gastrulation throughout the developing endoderm. Expression was punctate throughout the endoderm, including within the dorsal marginal zone (Fig. 1A). During neurulation, expression remained punctate throughout the endoderm, and was also detected in the archenteron roof as well as the mesoderm (Fig. 1B arrowheads). At later tadpole stages, expression was localized to the anterior endoderm, with expression also detected in the spinal cord (Fig. 1C, D). At NF40, when the anterior endoderm has been patterned into discrete organs, Staufen2 continued to be expressed in the stomach, liver and pancreas (Fig. 1E), and expression remained restricted to these regions at later stages as well (Fig. 1F). No expression was detected in posterior endoderm tissues at any stage. The restricted expression pattern of Staufen2 within the anterior endoderm during development suggested a possible role in differentiation of endodermal organs.
Fig. 1. Staufen2 expression in the endoderm and isolated gut tissue.

A: Transverse section through embryo at NF10 showing punctate expression of Staufen 2 throughout the mesendodermal precursor cells (arrowheads) as well as in the dorsal marginal zone (DMZ). B: Transverse section through embryo at NF15 showing Staufen2 expression within the archenteron roof (Ac) endoderm (arrowheads). Expression is also detected in the mesoderm (Mes). C: NF35 embryos showing Staufen2 expression throughout the head, in the spinal cord and in the dorsal (dp) and ventral (vp) pancreatic buds. D: Sagittal section through Staufen2 expressing region of NF35 embryo showing broad expression in the endoderm (en) and in the spinal cord (sc). E,F: Whole guts from NF40 and NF44 tadpole, expression was found throughout the pancreas, liver and stomach, becoming slightly stronger at later stages. St, stomach; L, liver; dp, dorsal pancreas; vp, ventral pancreas.
To investigate whether Staufen2 was necessary for endoderm development we created a knockdown phenotype using antisense morpholinos. The antisense morpholino was designed to the 5′UTR and injected (40ng) into the dorsal-vegetal blastomeres at the eight-cell stage targeting the anterior endoderm. Embryos injected with antisense Staufen2 morpholino developed normally during the first 3 days, without any overt defects. Beginning at NF40 we noticed a lack of discernable outgrowth of the stomach, liver and pancreas (Fig. 2). This phenotype was observed in 100% of embryos where the morpholino was targeted to the anterior endoderm (n=162). The fact that the embryos only displayed a phenotype at the stage when discrete organs are first observed in tadpoles suggested that functional Staufen2 was required for specification or patterning of the anterior endoderm.
Fig. 2. Staufen2 is required for anterior endodermal organ formation.

A,B: Insulin expression at NF44 is almost completely abolished (75%, n=16). C,D: Glucagon expression at NF44 is almost entirely lost in the stomach and pancreas (67%, n=6). E,F: Expression of PDIp is almost completely lost in Staufen2 knockdown animals at NF44 (75%, n=12). G,H: tm4sf3 gene expression is lost at NF44 (78%, n=9). I,J: Expression of hex is lost at NF44 (100%, n=11). K,L: Real-time PCR analysis of endocrine and exocrine markers in Staufen2 knockdown embryos for amylase in control and Staufen2 knockdown whole guts at NF44 and insulin expression in whole embryos at NF35. Each bar is an average of 8 individual whole tadpoles. Amylase expression is almost lost, and insulin expression is decreased by more than 2-fold. M–P: Flag-Staufen2 mRNA rescues the morpholino knockdown phenotype. M: Control NF45 whole gut stained for ptf1a mRNA showing expression in the pancreas. N: Whole gut from embryo injected with 40 ng of Staufen2 morpholino. Ptf1a expression is almost completely lost in injected embryos (n=20). O: Isolated whole gut stained for ptf1a expression from embryo injected with 40 ng of Staufen2 morpholino and 2000 pg of flag-Staufen2 mRNA. Ptf1a expression was restored in 31/45 embryos. P: In vitro transcription translation of staufen2 mRNA with either the morpholino (MO) or the mismatch morpholino (MM).
To define which organs were affected by the knockdown of Staufen2 protein, we examined the expression of pancreas, stomach and liver markers. Within the pancreas, both endocrine and exocrine markers were considerably reduced. At NF44 expression of insulin, which is normally punctate throughout the pancreas, was significantly diminished (Fig. 2A, B). We have previously observed a similar phenotype in Ptf1a knockdown embryos, where there was no apparent pancreatic tissue, yet insulin-expressing cells are detected. Since morpholino knockdown is not 100% effective, it is possible that an endocrine cell fate can still be initiated from the residual, lower level of Staufen2 (Jarikji et al., 2007). Expression of glucagon, which is expressed in both pancreatic and gastrointestinal endocrine cells was also substantially reduced (Fig. 2C, D). Similarly, expression of the exocrine marker XPDIp was almost entirely lost (Fig. 2E, F). We confirmed these results using quantitative RT-PCR; amylase expression was almost completely abolished whereas insulin expression was reduced more than 2-fold (Fig. 2K, L). In addition to the pancreas, we found expression of stomach marker, tm4sf3, to be significantly reduced in the anterior endoderm, while expression of the liver maker hex was absent in Staufen2 morphants (Fig. 2G–J). Taken together, these results demonstrate that Staufen2 function was required for proper development of pancreas, stomach and liver prior to NF40.
To confirm that the observed phenotype was directly related to the inhibition of endogenous Staufen2 we attempted to rescue the morpholino-induced phenotype through co-injection of a flag-Staufen2 mRNA lacking the 5′UTR sequence to which the morpholino was designed. Co-injection of 2000 pg of flag-Staufen2 was sufficient to rescue the differentiation of the endoderm as judged by expression of ptf1a (Fig. 2M–O). In control embryos, the stomach, liver and pancreas were all of normal size and abundant ptf1a expression was detected in the pancreas (Fig. 2M). In the morpholino-injected embryos, the expression of ptf1a was reduced in 100% of the injected samples (Fig. 2N, n=20). In embryos injected with the morpholino and flag-Staufen2 mRNA the morphology of the stomach, liver and pancreas were restored along with the expression of ptf1a in 69% of the injected embryos (Fig. 2O, n=45). In vitro transcription and translation confirmed that the morpholino blocked translation of Staufen2 mRNA, while the mismatch did not (Fig. 2P). These results demonstrate that the morpholino-induced phenotype was directly due to the loss of Staufen2.
Staufen2 is required for the patterning of the developing endoderm
To determine at which stage Staufen2 function was required for endoderm development, we studied the effect of Staufen2 knockdown at earlier stages, before overt morphogenesis of the organs has occurred. At NF35, we examined whether the anterior endoderm was properly patterned and specified by determining whether expression of pdx1 and ptf1a was affected. In Staufen2 knockdown embryos we found a complete absence of both pdx1 and ptf1a expression, indicating that the pancreas and duodenum was not properly induced (Fig. 3A–D). Similarly, we found expression of the endocrine progenitor marker, insm1, and the beta cell marker insulin to be diminished, reflecting the reduction of endocrine cells observed in older tadpoles (Fig. 4A, B, data not shown). Expression of the early acinar cell marker, XPDIp was also completely lost in the developing endoderm of the knockdown embryo (Fig. 4C, D). As we also observed a lost of stomach and liver organs, we studied the expression of frp5 and hnf6, early markers for the stomach and liver, respectively, to determine whether Staufen2 function was also required for their specification. We found expression of frp5 and hnf6 to be almost completely absent within the developing anterior endoderm at NF35 (Fig. 4E–H). The loss of early expression of genes required for the development of stomach, liver and pancreas indicated that patterning of the anterior endoderm into discrete organ domains did not properly occur, and suggested that Staufen2 function was required at a much earlier stage.
Fig. 3. Staufen2 is required for the patterning of the developing anterior endoderm.
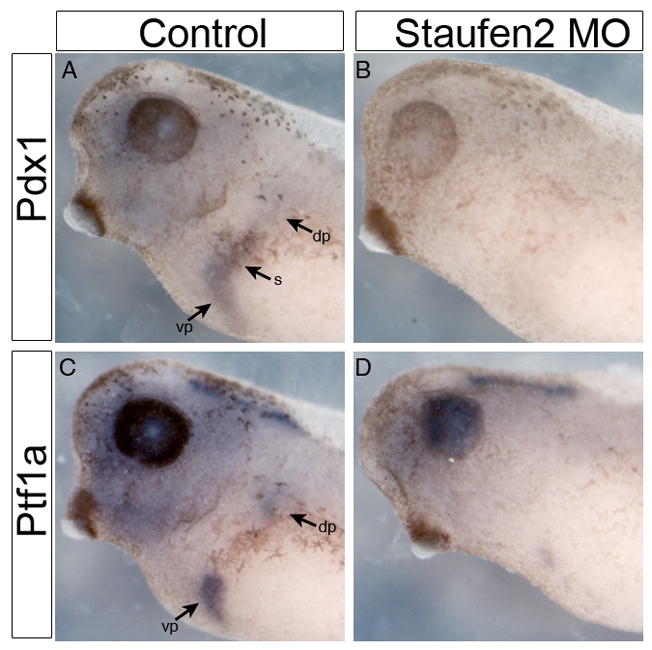
A,B: Pdx1 expression by whole mount in situ hybridization in control and Staufen2-MO-injected embryos at NF35, expression of pdx1 is lost in the developing stomach and pancreatic anlagen (83%, n=12).. C,D: NF35 embryo, expression of ptf1a is almost completely lost in the developing dorsal and ventral pancreatic bud (80%, n=10).
Fig. 4. Patterning of anterior endoderm is perturbed in Staufen 2 morpholino injected embryos.

Sagittal sections through the anterior endoderm of NF35 control and Staufen 2 morpholino injected embryos. A,B: Insulin expression is almost completely abolished (93%, n=14). Arrowhead in B points to the small remaining insulin expression in the dorsal pancreas. C,D: Expression of PDIp is lost in the future dorsal and ventral anlagen (100%, n=6). E,F: Expression of frp5 is almost completely lost (100%, n = 7). G,H: Expression of hnf6 is lost (63%, n=8). dp, dorsal pancreas; vp, ventral pancreas; st/du, stomach/duodenum; li, liver
Staufen2 acts upstream of the endoderm transcription factor network
We next examined whether Staufen2 function was required at earlier stages for the commitment of cells to an endodermal fate or for early anterior-posterior patterning of the endoderm. To determine if commitment to an endoderm fate was affected, we examined whether expression of the general endoderm marker sox17beta expression was disrupted. In control embryos sox17beta was expressed in a punctate pattern throughout the endoderm, and no change was seen in Staufen2 knockdown embryos (Fig. 5A, B). This confirmed that commitment to an endodermal cell fate was not dependent on Staufen2 function and that perhaps it was early anterior-posterior (AP) patterning that was affected. To ascertain whether AP patterning was altered, we examined whether expression of key anterior endoderm markers gata4, sizzled, foxA2, hnf6, and hex were reduced in Staufen morphants. In Staufen2 knockdown embryos, expression of gata4, sizzled, foxA2, hnf6, and hex were all lost within the anterior endoderm (Fig. 5C–H, data not shown). These results demonstrated that Staufen2 was required at gastrula and neurula stages when the endoderm is specified into anterior and posterior domains.
Fig. 5. Staufen2 is required for the earliest stages of endoderm regionalization.

A–L: Transverse section through the embryo at NF18. A,B: Sox17β expression by whole mount in situ hybridization in control and Staufen2-MO-injected embryos, expression of sox17β remains unchanged throughout the endoderm (80%, n=20). C,D: Expression of gata4 is lost in the anterior endoderm (82%, n=17). E,F: Expression of hnf6 is lost in the anterior endoderm (67%, n=9). G,H: Expression of hex is almost completely lost (72%, n=18). Brackets identify anterior endoderm
Staufen2 regulates early endoderm patterning through the BMP pathway
Expression of endogenous staufen2 within the dorsal marginal zone and perturbed endoderm axis development suggests it may be involved in the formation or function of the organizer. To define the mechanism by which Staufen2 inhibits anterior endoderm development, we analyzed gastrula stage embryos for altered expression of opposing signaling pathway components involved in the organizer activity. In embryos injected with Staufen2 morpholino, there was a loss of dorsal expression of chordin at NF10 (Fig. 6A, B). Chordin is secreted from the Spemann organizer and acts as a BMP antagonist, to keep BMP signaling levels lowest near the organizer. Conversely, expression of bmp4 and vent2, a downstream target of BMP4, were upregulated in the anterior endoderm of Staufen2 knockdown embryos (Fig. 6C, D, data not shown). To determine whether loss of Staufen2 is acting specifically on chordin expression or whether it has a general effect on the organizer we studied the expression of cerberus, a multivalent inhibitor secreted by the organizer and known to antagonize Nodal, BMP and Wnt. Cerberus expression however was unaffected in embryos lacking Staufen2 (Fig. 6E, F). Taken together, these results indicate that Staufen2 is necessary during gastrulation to ensure the expression of chordin within the developing embryo, leading to the appropriate establishment of the BMP signaling gradient.
Fig. 6. Staufen2 regulates early endoderm patterning through the BMP pathway.

A,B: Chd expression by whole mount in situ hybridization in control and Staufen2-MO-injected embryos at NF10 transverse section, expression of chd is lost in the dorsal half of the embryo (80%, n=10). Arrows indicate dorsal lip. C,D: Vent2 expression is significantly upregulated in the anterior half of the embryo at NF18, transverse section (75%, n=12). Anterior is to the left and dorsal is up. E,F: Expression of cerberus is unaffected in NF10 embryos (100%, n=10). Arrows indicate dorsal lip.
Discussion
In this study, we identified a new function for Staufen2 in endoderm specification. We showed that Staufen2 expression within the endoderm and mesoderm occurred early in development beginning at gastrulation, and is expressed in the dorsal marginal zone. By performing loss-of-function experiments, we demonstrated that Staufen2 is required for expression of chordin. Knockdown of Staufen2 resulted in increased expression of BMP4 signaling components and altered regionalization of the developing anterior endoderm. This led to an almost complete loss of pancreas, stomach and liver differentiation, which ultimately led to hypoplasia or aplasia of these organs. Based on these results, we conclude that Staufen2 is an indispensable mediator of anterior endoderm patterning in X. laevis.
The role of RNA binding proteins in endoderm development has not been profoundly investigated, though increased focus towards understanding the molecular networks involved in the development of the endoderm has led to a recent studies detailing RNA binding protein function. Our lab recently identified another RNA binding protein involved in endoderm development, BrunoL1, which was shown to regulate proliferation of endoderm cells through translational control of cyclin A2 mRNA (Horb and Horb, 2010). Another study in Xenopus showed that Vg1RBP was essential for pancreas development (Spagnoli and Brivanlou, 2006), while in zebrafish, nil per os (npo) was shown to be required for gut morphogenesis and exocrine pancreas development (Mayer and Fishman, 2003). While these results suggest that modulating the activity of target RNAs may control organogenesis in part, what these targets are and how they are involved in endoderm development remains to be elucidated.
It is unclear how Staufen2 mediates its effect on chordin expression, but data from other organisms showed that Staufen2 acts to regulate mRNA localization and translation. In zebrafish, Staufen proteins seem to be required for migration and survival of primordial germ cells through mRNA localization (Ramasamy et al., 2006). It is possible that in Xenopus endoderm development Staufen2 may be involved in the asymmetric localization of specific mRNAs, which in turn regulate activation of Chordin, though we do not necessarily agree with this hypothesis. In mammals, Staufen1 protein has been shown to be apically distributed within differentiated Caco-2 intestinal cells (Gautrey et al., 2005). Finally, in Drosophila, interaction between Staufen and the 3′UTR of propero is required for basal localization during embryonic neuroblast differentiation (Schuldt et al., 1998; Shen et al., 1998).
Alternatively, Staufen1 has been shown to mediate mRNA decay for conditional down regulation of genes encoding mRNAs that contain a STAU1 binding site in their 3′UTR (Kim et al., 2005b; Gong et al., 2009). In C2C12 myoblast cells, Stau1-mediated mRNA decay (SMD) increases in efficiency as these cells differentiate into myotubes, and Pax3 mRNA, which encodes a transcription factor that inhibits myogenesis, has been identified as a target of SMD, therefore its down-regulation allows for myogenesis to occur (Epstein et al., 1995; Gong et al., 2009). While Staufen2 has yet to be studied as a mediator of mRNA decay, it may be possible that instead of playing a role in the localization and translation of mRNA required for endoderm development, Staufen2 may be required to down regulate genes that are involved in the specification and regionalization of the endoderm.
We have shown here that Staufen2 is required for the proper expression of chordin during gastrulation. Although we have not conclusively addressed whether loss of Chordin is directly responsible for the observed foregut defects, there is increasing evidence suggesting regionalization of the endoderm occurs as the germ layers are being formed at this early stage in development, and it was previously shown that knockdown of Chordin resulted in reduced expression of pdx1 (Zorn et al., 1999; Wells and Melton, 2000; Spagnoli and Brivanlou, 2008). Recently, the inhibition of BMP signaling within the dorsal endoderm was shown to be required for specification of the of the pancreatic field. Knockdown of Chordin, leads to dorsal overexpression of BMP, and ultimately in a loss of Pdx1 expression (Spagnoli and Brivanlou, 2008). While, it has been shown that this inhibition is endogenously mediated throughout endoderm development by Chordin and TGIF2, we have identified a novel role of Staufen2 that appears to be upstream of these factors. We believe that Staufen2 acts to mediate translation of specific targets at this early stage rather than asymmetric localization or mRNA decay. We are currently attempting to identify Staufen2 mRNA targets and ascertain whether this is indeed the case.
In conclusion, we have shown that Staufen 2 function is essential for proper development of anterior endodermal organs. In particular, we showed that knockdown of Staufen 2 resulted in very early patterning defects, resulting in decreased expression of chordin and a concomitant increase in Vent2 expression. In contrast, there was no change in expression of Cerberus. Although, we identified very early molecular changes, we were surprised by the restricted phenotypes observed at later stages in Staufen 2 morpholino injected tadpoles. After identifying that Staufen 2 was required for proper development of anterior endodermal organs, but not for head or other tissues, we expected to find molecular changes only after gastrulation, during neurula stages. At the moment we cannot explain these discrepancies, but it remains possible that chordin is expressed for a short period and that this is sufficient for maintaining its function during gastrulation. Identification of Staufen 2 mRNA targets should help resolve this issue.
Methods
Plasmids, RT-PCR and Whole Mount In Situ Hybridization
For all constructs, primers were designed based on full-length sequence information obtained from GenBank, and the PCR products were cloned into pCRII (Invitrogen). The GenBank accession number for Staufen2 is BC046732.1. RT-PCR was performed on isolated whole guts for amylase and NF35 whole embryos for insulin, and normalized to EF1α as described (Jarikji et al., 2007). Whole-mount in situ hybridizations with single probes were performed as described using BM Purple (Horb et al., 2003). Antisense digoxigenin probe for Staufen2 was synthesized from Staufen2 in pCRII linearized with XbaI and transcribed with SP6 RNA polymerase. Probes for other pancreatic markers were prepared as previously described (Horb et al., 2003; Jarikji et al., 2007). Complete information for each clone is available upon request. For whole-mount in situ analysis in the endoderm at tail bud and early tadpole stages was prepared as described (Horb et al., 2009).
Antisense morpholino
Antisense morpholino oligonucleotides were designed by Gene Tools, LLC. The antisense morpholinos were designed in the 5′UTR. Morpholinos were injected into the dorsal vegetal blastomeres at the eight-cell stage. For functional analysis, we selected only those samples where the morpholino targeted the entire anterior part of the gut. Targeting to the stomach, liver and pancreas was confirmed by monitoring fluorescence from labeled oligonucleotides, only after isolation of whole guts from injected embryos. The sequences of the antisense morpholinos used was Staufen2 5′-CCGGGCTGCTCACACCTCCAACATT-3, and the sequence of the mismatch morpholino used was Staufen2-mismatch 5′-TGGCTCTGCGTTAATCTCTGCGATC-3′ The specificity of the morpholinos was tested by in vitro transcription/translation reactions using a FLAG antibody to detect the Staufen2 protein as described (Horb et al., 2009).
Acknowledgments
This work was supported by grants from the National Institutes of Health (DK077197) to M.E.H. and from the Canadian Institute of Health Research (Frederick Banting and Charles Best Canada Graduate Scholarships - Doctoral Awards) to C.K.B. Special thanks go to Lori Dawn Horb for her help with injections, Zeina Jarikji for cloning the full length Staufen2, and to Frédéric Bourque for his care of the frogs. We would also like to thank the MBL for their support and help in setting up the new lab.
References
- Allison R, Czaplinski K, Git A, Adegbenro E, Stennard F, Houliston E, Standart N. Two distinct Staufen isoforms in Xenopus are vegetally localized during oogenesis. Rna. 2004;10:1751–1763. doi: 10.1261/rna.7450204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broadus J, Fuerstenberg S, Doe CQ. Staufen-dependent localization of prospero mRNA contributes to neuroblast daughter-cell fate. Nature. 1998;391:792–795. doi: 10.1038/35861. [DOI] [PubMed] [Google Scholar]
- Buchner G, Bassi MT, Andolfi G, Ballabio A, Franco B. Identification of a novel homolog of the Drosophila staufen protein in the chromosome 8q13–q21.1 region. Genomics. 1999;62:113–118. doi: 10.1006/geno.1999.6015. [DOI] [PubMed] [Google Scholar]
- Dale L, Slack JM. Fate map for the 32-cell stage of Xenopus laevis. Development. 1987;99:527–551. doi: 10.1242/dev.99.4.527. [DOI] [PubMed] [Google Scholar]
- Epstein JA, Lam P, Jepeal L, Maas RL, Shapiro DN. Pax3 inhibits myogenic differentiation of cultured myoblast cells. J Biol Chem. 1995;270:11719–11722. doi: 10.1074/jbc.270.20.11719. [DOI] [PubMed] [Google Scholar]
- Ferrandon D, Elphick L, Nüsslein-Volhard C, St Johnston D. Staufen protein associates with the 3′UTR of bicoid mRNA to form particles that move in a microtubule-dependent manner. Cell. 1994;79:1221–1232. doi: 10.1016/0092-8674(94)90013-2. [DOI] [PubMed] [Google Scholar]
- Gautrey H, McConnell J, Hall J, Hesketh J. Polarised distribution of the RNA-binding protein Staufen in differentiated intestinal epithelial cells. FEBS Lett. 2005;579:2226–2230. doi: 10.1016/j.febslet.2005.02.074. [DOI] [PubMed] [Google Scholar]
- Gong C, Kim YK, Woeller CF, Tang Y, Maquat LE. SMD and NMD are competitive pathways that contribute to myogenesis: effects on PAX3 and myogenin mRNAs. Genes & Development. 2009;23:54–66. doi: 10.1101/gad.1717309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horb LD, Horb ME. BrunoL1 regulates endoderm proliferation through translational enhancement of cyclin A2 mRNA. Dev Biol. 2010 doi: 10.1016/j.ydbio.2010.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horb LD, Jarkji ZH, Horb ME. Xenopus Insm1 is essential for gastrointestinal and pancreatic endocrine cell development. Dev Dyn. 2009;238:2505–2510. doi: 10.1002/dvdy.22071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horb ME, Shen CN, Tosh D, Slack JM. Experimental conversion of liver to pancreas. Curr Biol. 2003;13:105–115. doi: 10.1016/s0960-9822(02)01434-3. [DOI] [PubMed] [Google Scholar]
- Horb ME, Thomsen GH. A vegetally localized T-box transcription factor in Xenopus eggs specifies mesoderm and endoderm and is essential for embryonic mesoderm formation. Development. 1997;124:1689–1698. doi: 10.1242/dev.124.9.1689. [DOI] [PubMed] [Google Scholar]
- Jarikji ZH, Vanamala S, Beck CW, Wright CV, Leach SD, Horb ME. Differential ability of Ptf1a and Ptf1a-VP16 to convert stomach, duodenum and liver to pancreas. Dev Biol. 2007;304:786–799. doi: 10.1016/j.ydbio.2007.01.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanai Y, Dohmae N, Hirokawa N. Kinesin transports RNA: isolation and characterization of an RNA-transporting granule. Neuron. 2004;43:513–525. doi: 10.1016/j.neuron.2004.07.022. [DOI] [PubMed] [Google Scholar]
- Kim HK, Kim YB, Kim EG, Schuman E. Measurement of dendritic mRNA transport using ribosomal markers. Biochem Biophys Res Commun. 2005a;328:895–900. doi: 10.1016/j.bbrc.2005.01.041. [DOI] [PubMed] [Google Scholar]
- Kim YK, Furic L, Desgroseillers L, Maquat LE. Mammalian Staufen1 recruits Upf1 to specific mRNA 3′UTRs so as to elicit mRNA decay. Cell. 2005b;120:195–208. doi: 10.1016/j.cell.2004.11.050. [DOI] [PubMed] [Google Scholar]
- Kuersten S, Goodwin EB. The power of the 3′ UTR: translational control and development. Nat Rev Genet. 2003;4:626–637. doi: 10.1038/nrg1125. [DOI] [PubMed] [Google Scholar]
- Lustig KD, Kroll KL, Sun EE, Kirschner MW. Expression cloning of a Xenopus T-related gene (Xombi) involved in mesodermal patterning and blastopore lip formation. Development. 1996;122:4001–4012. doi: 10.1242/dev.122.12.4001. [DOI] [PubMed] [Google Scholar]
- Macchi P, Kroening S, Palacios IM, Baldassa S, Grunewald B, Ambrosino C, Goetze B, Lupas A, St Johnston D, Kiebler M. Barentsz, a new component of the Staufen-containing ribonucleoprotein particles in mammalian cells, interacts with Staufen in an RNA-dependent manner. J Neurosci. 2003;23:5778–5788. doi: 10.1523/JNEUROSCI.23-13-05778.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer AN, Fishman MC. Nil per os encodes a conserved RNA recognition motif protein required for morphogenesis and cytodifferentiation of digestive organs in zebrafish. Development. 2003;130:3917–3928. doi: 10.1242/dev.00600. [DOI] [PubMed] [Google Scholar]
- McLin VA, Rankin SA, Zorn AM. Repression of Wnt/beta-catenin signaling in the anterior endoderm is essential for liver and pancreas development. Development. 2007;134:2207–2217. doi: 10.1242/dev.001230. [DOI] [PubMed] [Google Scholar]
- Pan FC, Chen Y, Bayha E, Pieler T. Retinoic acid-mediated patterning of the pre-pancreatic endoderm in Xenopus operates via direct and indirect mechanisms. Mech Dev. 2007;124:518–531. doi: 10.1016/j.mod.2007.06.003. [DOI] [PubMed] [Google Scholar]
- Pearl EJ, Bilogan CK, Mukhi S, Brown DD, Horb ME. Xenopus pancreas development. Dev Dyn. 2009;238:1271–1286. doi: 10.1002/dvdy.21935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramasamy S, Wang H, Quach HN, Sampath K. Zebrafish Staufen1 and Staufen2 are required for the survival and migration of primordial germ cells. Dev Biol. 2006;292:393–406. doi: 10.1016/j.ydbio.2006.01.014. [DOI] [PubMed] [Google Scholar]
- Roegiers F, Jan YN. Staufen: a common component of mRNA transport in oocytes and neurons? Trends Cell Biol. 2000;10:220–224. doi: 10.1016/s0962-8924(00)01767-0. [DOI] [PubMed] [Google Scholar]
- Saunders LR, Barber GN. The dsRNA binding protein family: critical roles, diverse cellular functions. FASEB J. 2003;17:961–983. doi: 10.1096/fj.02-0958rev. [DOI] [PubMed] [Google Scholar]
- Saunders PT, Pathirana S, Maguire SM, Doyle M, Wood T, Bownes M. Mouse staufen genes are expressed in germ cells during oogenesis and spermatogenesis. Mol Hum Reorid. 2000;6:983–991. doi: 10.1093/molehr/6.11.983. [DOI] [PubMed] [Google Scholar]
- Schuldt AJ, Adams JH, Davidson CM, Micklem DR, Haseloff J, St Johnston D, Brand AH. Miranda mediates asymmetric protein and RNA localization in the developing nervous system. Genes & Development. 1998;12:1847–1857. doi: 10.1101/gad.12.12.1847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen CP, Knoblich JA, Chan YM, Jiang MM, Jan LY, Jan YN. Miranda as a multidomain adapter linking apically localized Inscuteable and basally localized Staufen and Prospero during asymmetric cell division in Drosophila. Genes & Development. 1998;12:1837–1846. doi: 10.1101/gad.12.12.1837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinner D, Kirilenko P, Rankin S, Wei E, Howard L, Kofron M, Heasman J, Woodland HR, Zorn AM. Global analysis of the transcriptional network controlling Xenopus endoderm formation. Development. 2006;133:1955–1966. doi: 10.1242/dev.02358. [DOI] [PubMed] [Google Scholar]
- Spagnoli FM, Brivanlou AH. The RNA-binding protein, Vg1RBP, is required for pancreatic fate specification. Dev Biol. 2006;292:442–456. doi: 10.1016/j.ydbio.2006.01.022. [DOI] [PubMed] [Google Scholar]
- Spagnoli FM, Brivanlou AH. The Gata5 target, TGIF2, defines the pancreatic region by modulating BMP signals within the endoderm. Development. 2008;135:451–461. doi: 10.1242/dev.008458. [DOI] [PubMed] [Google Scholar]
- St Johnston D. The intracellular localization of messenger RNAs. Cell. 1995;81:161–170. doi: 10.1016/0092-8674(95)90324-0. [DOI] [PubMed] [Google Scholar]
- St Johnston D, Beuchle D, Nüsslein-Volhard C. Staufen, a gene required to localize maternal RNAs in the Drosophila egg. Cell. 1991;66:51–63. doi: 10.1016/0092-8674(91)90138-o. [DOI] [PubMed] [Google Scholar]
- St Johnston D, Driever W, Berleth T, Richstein S, Nüsslein-Volhard C. Multiple steps in the localization of bicoid RNA to the anterior pole of the Drosophila oocyte. Development. 1989;107(Suppl):13–19. doi: 10.1242/dev.107.Supplement.13. [DOI] [PubMed] [Google Scholar]
- Stennard F, Carnac G, Gurdon JB. The Xenopus T-box gene, Antipodean, encodes a vegetally localised maternal mRNA and can trigger mesoderm formation. Development. 1996;122:4179–4188. doi: 10.1242/dev.122.12.4179. [DOI] [PubMed] [Google Scholar]
- Tian B, Bevilacqua PC, Diegelman-Parente A, Mathews MB. The double-stranded-RNA-binding motif: interference and much more. Nat Rev Mol Cell Biol. 2004;5:1013–1023. doi: 10.1038/nrm1528. [DOI] [PubMed] [Google Scholar]
- Wells JM, Melton DA. Early mouse endoderm is patterned by soluble factors from adjacent germ layers. Development. 2000;127:1563–1572. doi: 10.1242/dev.127.8.1563. [DOI] [PubMed] [Google Scholar]
- Xanthos JB, Kofron M, Wylie C, Heasman J. Maternal VegT is the initiator of a molecular network specifying endoderm in Xenopus laevis. Development. 2001;128:167–180. doi: 10.1242/dev.128.2.167. [DOI] [PubMed] [Google Scholar]
- Yoon YJ, Mowry KL. Xenopus Staufen is a component of a ribonucleoprotein complex containing Vg1 RNA and kinesin. Development. 2004;131:3035–3045. doi: 10.1242/dev.01170. [DOI] [PubMed] [Google Scholar]
- Zhang J, King ML. Xenopus VegT RNA is localized to the vegetal cortex during oogenesis and encodes a novel T-box transcription factor involved in mesodermal patterning. Development. 1996;122:4119–4129. doi: 10.1242/dev.122.12.4119. [DOI] [PubMed] [Google Scholar]
- Zorn AM, Butler K, Gurdon JB. Anterior endomesoderm specification in Xenopus by Wnt/beta-catenin and TGF-beta signalling pathways. Dev Biol. 1999;209:282–297. doi: 10.1006/dbio.1999.9257. [DOI] [PubMed] [Google Scholar]
- Zorn AM, Wells JM. Vertebrate endoderm development and organ formation. Annu Rev Cell Dev Biol. 2009;25:221–251. doi: 10.1146/annurev.cellbio.042308.113344. [DOI] [PMC free article] [PubMed] [Google Scholar]