

Abstract
Haematopoietic stem cells (HSCs) are a self-renewing population that continuously replenish all blood and immune cells during the lifetime of an individual1, 2. HSCs are used clinically to treat a wide array of diseases, including acute leukaemias and congenital blood disorders, but obtaining suitable numbers of cells and finding immune compatible donors remain serious problems. These concerns have led to an interest in the conversion of embryonic stem cells or induced pluripotent stem cells into HSCs, which is not possible using current methodologies. To accomplish this goal, it is critical to understand the native mechanisms involved in specification of HSCs during embryonic development. Here we demonstrate that Wnt16 controls a novel genetic regulatory network required for HSC specification. Non-canonical signaling by Wnt16 is required for somitic expression of the Notch ligands deltaC (dlc) and deltaD (dld), and these ligands are in turn required for establishment of definitive haematopoiesis. Notch signalling downstream of Dlc/Dld is earlier than, and distinct from known cell-autonomous requirements for Notch, strongly suggesting that novel Notch-dependent relay signal(s) induce the first HSCs in parallel to other established pathways. Our results demonstrate that somite-specific gene expression is required for the production of haemogenic endothelium.
We wished to better define the role, if any, of Wnt signalling in HSC specification during embryonic development. Although Wnt signalling can exert strong effects on adult HSCs, an in vivo regulatory function is controversial, and the role of Wnt signalling in establishing haematopoiesis during development is unclear2. Wnt signalling pathways have been loosely grouped into two families, “canonical” and “noncanonical”2, 3. Canonical signalling is defined by stabilization of β-catenin, which in cooperation with lymphoid-enhancer-binding factor/T cell factor (Lef/Tcf) DNA-binding proteins, activates transcription of Wnt target genes2, 3. Non-canonical signalling is β-catenin/Tcf-independent, with less well characterized intracellular pathways3. Disruption of canonical signalling by targeted deletion of Wnt3a results in HSC deficits2, but these animals have massive morphological defects, including near absence of the caudal tissues4 where HSCs arise during embryogenesis. Canonical Wnt signalling by unknown ligands also appears to play a role in maintaining and/or expanding very early HSCs in cooperation with prostaglandins5. At present, no studies have demonstrated an absolute requirement for Wnt signalling in the earliest specification of recognizable HSCs, and no requirement for β-catenin/Tcf-independent, non-canonical signalling has been reported.
We searched for candidate Wnts expressed near prehaematopoietic mesoderm and identified a previously uncharacterized zebrafish wnt16 orthologue (Fig. S1) expressed in the dorsoanterior portion of more rostral somites from 10 hours post-fertilization (hpf; tailbud-stage) to 24 hpf (Fig. S2), the time frame when prehaematopoietic mesoderm transitions to recognizable HSC precursors in the dorsal aorta. Wnt16 is conserved across vertebrate phyla (Fig. S1), and the human form was originally identified as a gene inappropriately expressed in pre-B-acute lymphoblastic leukaemia (ALL) cells containing the E2A-PBX1 t(1;19) translocation product6. In zebrafish, two wnt16 splice variants are produced, only one of which is active (Fig. S1, a, b; Fig. S3; Table S1).
Knock down of Wnt16 by injection of either of two “splice-blocking” morpholinos caused a striking haematopoietic phenotype. Morpholinos alone or in combination reduced functional wnt16 message (Figs. S1, a; S4), yielding highly similar phenotypes in comparison to a 5-basepair mismatch control morpholino, which had no effect. (Figs. S4, S5). HSC precursors, a population of runx1+ cells in the ventral floor of the dorsal aorta1, 7, 8, as in mouse9, are absent in embryos injected with W16MO at 24 hpf by whole mount in situ hybridization (WISH; red arrowhead; Fig. 1 a, b; Table S2). By 33 hpf, HSCs can be observed as a population of cmyb+ cells between the dorsal aorta and posterior cardinal vein1, and this population is also absent (red arrowhead, Fig. 1 c, d; Table S2). HSCs observable in living transgenic animals carrying GFP under the control of the cd41 (itga2b) promoter10-12 are absent or reduced in wnt16 morphants (red arrowheads, Fig. 1 e, f; Movies S1 and S2; Table S2), as are unique double-positive cells in kdrl:RFP and cmyb:GFP double transgenics7 (Fig. 1 g, h, yellow cells; Table S2), whereas unrelated GFP-labelled multi-ciliate cells in the pronephros are unaffected (yellow arrowheads, Fig. 1 e, f; green cells, Fig. 1 g, h). Generation of T-lymphocytes requires an HSC precursor10, 11, 13, providing a useful readout for whether HSCs have been specified or not. In cd41:GFP transgenic animals, GFP+ HSCs retain residual fluorescence as they differentiate into the first thymic lymphocytes around 3 days post fertilization (dpf)10, and these thymic immigrants are greatly reduced in wnt16 morphants (Movies S3 and S4). Accordingly, wnt16 morphants lack rag1+ T lymphocytes at 4.5 dpf (blue arrows, Fig. 1, i, j; Table S2), despite intact thymic epithelium (Fig. S6). Together, our results indicate that Wnt16 is required for specification of HSCs during embryonic development.
Figure 1. Wnt16 is required for specification of HSCs.
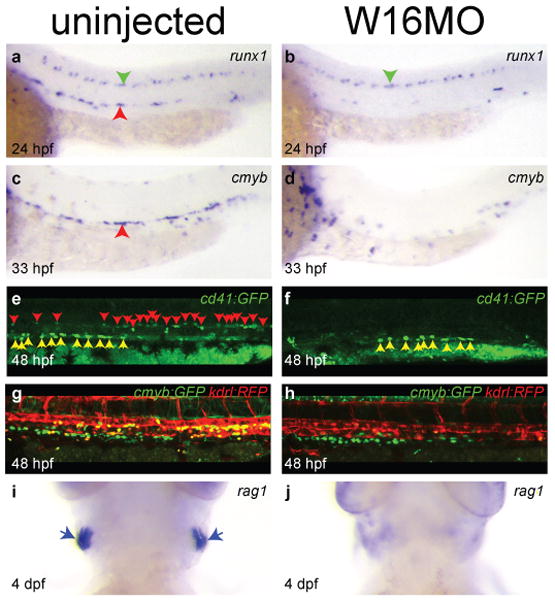
Expression of the HSC markers runx1 (a-b) and cmyb (c-d). Fluorescently labelled HSCs in cd41:gfp (e-f) and cmyb:GFP;kdrl:RFP transgenics (g-h). Expression of the lymphocyte marker rag1 (i-j). Embryos are either uninjected (left column) or injected with 5 ng W16MO (right column). Red arrowheads identify the aorta region (a, c) or individual HSCs (e). Green arrowheads identify unaffected runx1+ neurons (a-b). Yellow arrowheads identify unaffected GFP+ multiciliate cells of the pronephros (e-f). Yellow cells are HSCs (g). Blue arrows identify thymic T cells (i). a-h dorsal up, anterior left. i-j ventral views, anterior up.
To determine the specificity of W16MO-induced defects, we examined the integrity of non-hematopoietic tissues by morphology and expression of marker genes (Fig. 2; Table S3). Wnt16 morphants are grossly normal (Fig. 2 a, b), with intact and functional vasculature, as evidenced by beating hearts and circulating primitive, non-HSC-derived erythroid cells, although there appear to be subtle patterning defects in the number and position of intersegmental vessels. By WISH, W16MO-injected embryos have primitive blood (gata1; Fig. 2 c, d; Table S3), somites (myod; Fig. 2 e, f; Table S3), vasculature (tll1, cdh5, and flk1; Fig. 2 g-l; Table S3), dorsal aorta (efnb2a; Fig. 2 m, n; notch1b, notch3, dll4 Fig. S7, a-d, o, p; Tables S3, S5), hypochord (col2a1a; Fig. 2 o, p; Table S3), notochord and floorplate (shha; Fig. 2 q, r; Table S3), and pronephros (cdh17; Fig. 2 s, t; Table S3). Thus, defects in HSC specification in wnt16 morphant animals are highly specific and not due to wholesale failure in specification of nearby tissues.
Figure 2. Wnt16 loss of function phenotype is specific.
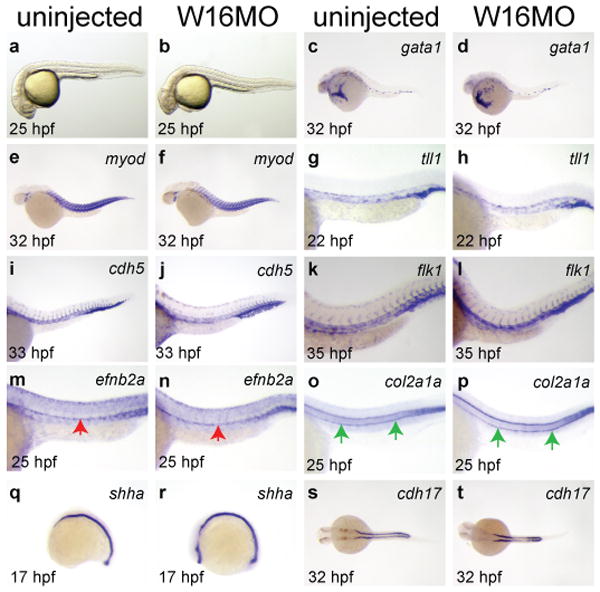
Uninjected or W16MO-injected embryos in brightfield (a-b) or processed by WISH for tissue specific genes: primitive blood (gata1, c-d), somites (myod, e-f), vasculature (tll1, g-h; cdh5, i-j; flk1, k-l), dorsal aorta (efnb2a, m-n), hypochord (col2a1a, o-p), floorplate and notochord (shha, q-r), or pronephros (cdh17, s-t) at the developmental times indicated. Red arrows, dorsal aorta (m-n); green arrows, hypochord (o-p). Anterior left. a-r, dorsal up. s-t, dorsal views.
Canonical, β-catenin/Tcf-dependent Wnt signalling has been reported to be involved in HSC specification in mouse2 and very early maintenance in zebrafish5. We therefore wanted to determine if Wnt16 is canonical. Overexpression of Wnt16 caused phenotypic defects demonstrating active protein (Fig. S3), but in comparison to Wnt314, did not cause ectopic expression of canonical targets (Fig. S8). Conversely, Wnt16 knock down caused no decrease in canonical reporter activity (Fig. S9). Thus, Wnt16 does not act via β-catenin/Tcf and must act via a non-canonical pathway as has been suggested for human WNT16b15, 16. Loss of other non-canonical Wnt ligands causes distinct phenotypes. Loss of Wnt5b is much more severe causing nearly complete absence of primary trunk vasculature17, whereas loss of Wnt11 had no effects on HSC specification (Fig. S10, Table S4). We therefore conclude that the HSC defects caused by loss of wnt16 are not a general consequence of loss of non-canonical Wnt signalling.
Notch signalling is required across phyla for developmental specification of HSCs1. Global inhibition of Notch signalling by mutation or targeted deletion of Mindbomb (Mib), which is required for Notch ligand activity, or the essential Notch DNA-binding co-factor Rbpjκ results in total loss of HSCs, as well as severe vascular defects1. If Wnt16 were to regulate Notch pathway genes, this regulation might explain the loss of HSCs in wnt16 morphant animals. The specific ligands and receptors required in zebrafish for specification of HSCs by the Notch pathway have not been determined. We therefore examined comparative expression of Notch ligands and receptors that might participate in HSC specification in uninjected or W16MO-injected animals. Most Notch receptors and ligands were either unaffected or very weakly affected (Fig. S7; Fig. 4 c,d; Table S5). However, expression of two Notch ligands, dlc and dld, was markedly decreased in somites at 17 hpf (16-ss; Fig. 3 a-d, Table S5). In accord with decreased somitic ligand expression, Notch reporter activity was decreased in somites at 17.5 hpf (Fig. S11). Interestingly, diminution of dlc and dld was tissue-specific, because expression was maintained in presomitic mesoderm (Fig. 4 a-d), and dlc expression in the dorsal aorta appeared relatively normal at 22 hpf (Fig. S12, Table S5). Although Wnt16 function is required for somitic expression of dlc and dld, we see no evidence for reciprocal regulation of wnt16 by Dlc and Dld (Fig. S13), or indeed by Notch at all, as expression is unaffected in mib mutants (not shown).
Figure 4. Non-cell-autonomous requirement for Notch in HSC specification.
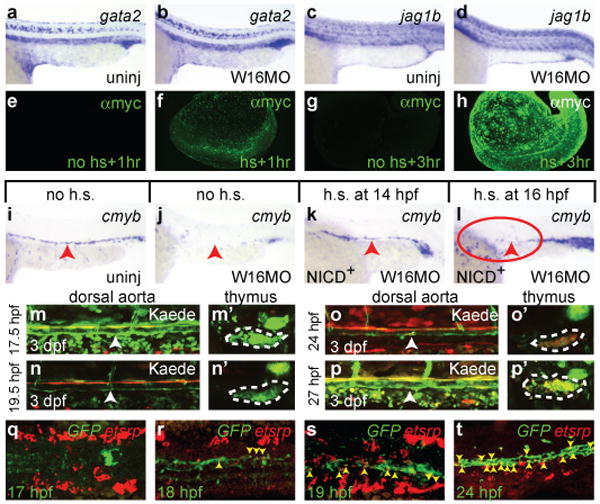
Expression of the Notch target gata2 (a-b) and the Notch ligand jag1b (c-d) in uninjected (a, c) or W16MO-injected (b, d) embryos at 22 hpf. Whole mount immunofluorescence visualization of the myc-tagged NICD at 1 (e-f) and 3 (g-h) hours following either no induction (e, g) or heat-shock induction (f, h). Cmyb expression at 36 hpf in transgenic animals carrying a heat-shock inducible dominant activator of Notch signalling (NICD) in uninjected (i), W16MO-injected (j), W16MO-injected and heat-shock induced at 14 hpf (10-ss; k) or 16 hpf (14-ss; l). Red arrowheads indicate the dorsal aorta region. Red circle where HSCs should normally be expressed (h). Green-to-red photoconvertible Kaede Notch reporter animals were entirely photoconverted at the times indicated at left of each panel pair and imaged at 3 dpf (m-p′). Confocal images of the dorsal aorta (white arrowheads; m-p) and thymus (dashed white outline; m′-p′) reveal photoconverted cells only in the thymi of fish converted at 24 and 27 hpf (o′, p′). Max-projection confocal images of the trunk region of embryos processed by double fluorescence in situ for a Notch reporter GFP transgene (green) and the haematopoietic mesoderm marker etsrp (red) at the times indicated (q-t). Yellow arrowheads indicate double-positive cells (r-t). a-d, i-l close up lateral views of the trunk region, anterior left, dorsal up. e-h, whole embryo views. m-p close up lateral views of the dorsal aorta. m′-p′ single thymic lobes. q-s, dorsal views, anterior left. t, close up lateral aorta view, anterior left, dorsal up.
Figure 3. Wnt16 acts upstream of Notch ligands Dlc and Dld.
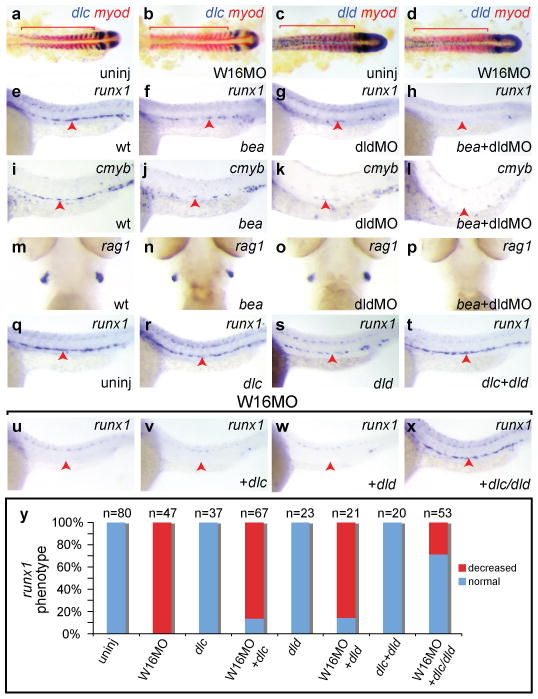
Expression of somitic dlc (a-b) and dld (c-d) but not myod (red) is decreased at 17 hpf in W16MO-injected embryos. Red bars indicate somites. Expression of the HSC markers runx1 at 24 hpf (e-h) and cmyb at 36 hpf (i-l) is reduced in dlc mutant (bea) embryos (f, j) or dldMO-injected embryos (g, k), and eliminated in the combined animals (h, l). The lymphocyte marker rag1 at 4.5 dpf is present in wild-type (m), bea (n), and dldMO-injected animals (o), but eliminated in bea embryos injected with dldMO (p). Combined injection of dlc/dld rescues HSCs in wnt16 morphants (q-x). Runx1 at 24 hpf. One group of embryos was uninjected, or injected with the indicated Notch ligand mRNAs alone (q-t). A second group of W16MO-injected embryos was co-injected with Notch ligand mRNAs (u-x). Percentages of embryos displaying the depicted phenotypes (y). a-d flat mounts, anterior left. e-l, q-x close up lateral views, anterior left, dorsal up. m-p ventral head views, anterior up. Red arrowheads indicate the aorta region.
To determine whether alterations in the expression of dlc and/or dld might explain the decrease in HSCs we observe in wnt16 morphant animals, we tested whether loss of function in these genes alone or in combination could alter HSC specification. Beamter (bea) mutants carry a predicted null allele of dlc18. We compared HSC and T lymphocyte marker expression in wild-type embryos, homozygous bea mutants, embryos injected with dld morpholino (dldMO)19, and homozygous bea mutants injected with dldMO. We found that runx1 transcripts at 24 hpf, and cmyb at 36 hpf, were greatly reduced in bea homozygous mutants and embryos injected with dldMO (Fig. 3, e-g, i-k: Table S6), but an apparent recovery of HSCs had occurred by 4.5 dpf, when rag1 transcripts, revealing the thymic progeny of HSCs, were readily visible in both groups (Fig. 3 m-o; Table S6). Combinatorial reduction of Dlc and Dld activity, however, eliminated HSCs, as revealed by elimination of runx1 at 24 hpf (Fig. 3, h; Table S6), cmyb at 36 hpf (Fig. 3, l; Table S6), and rag1 transcripts at 4.5 dpf (Fig. 3, p, Table S6). Thus, dlc and dld are combinatorially required for specification of HSCs in zebrafish, and their diminished expression in the somites of wnt16 morphant animals can, in principle, explain the observed loss of HSCs.
To confirm that loss of HSCs in wnt16 morphants is due to loss of dlc and dld, we performed a rescue experiment. We injected embryos with W16MO, and in some cases co-injected mRNA encoding full-length Dlc and Dld ligands singly or in combination. Injection of dlc and dld mRNA did not alone, or in combination have a strong effect on HSC numbers, as measured by runx1+ cells in the dorsal aorta (Fig. 3, q-t, y). Co-injection of individual mRNAs with W16MO was unable to restore runx1 in (Fig. 3, u-w, y). However, when injected together, dlc and dld restored runx1+ HSCs in a high percentage of W16MO-injected animals (Fig. 3, x, y). These results confirm that decreased dlc and dld expression in wnt16 morphants is responsible for loss of HSCs.
Studies in zebrafish and mouse have shown that Notch specification of HSCs is regulated by Shh and VegfA1. Shh regulates expression of vegfa, and VegfA signalling is in turn required for vascular expression of Notch receptor genes1. Notch signalling is required for both arteriovenous and HSC specification1, 20-24. In accord with these observations, loss of Shh signalling causes loss of both artery and HSCs1. Taken together, one level of control over HSC specification appears to be through a Shh/VegfA/Notch signalling pathway1. Shh/VegfA/Notch specification of HSCs appears to be distinct from Wnt16/Dlc/Dld effects because vascular and arterial specification is unaffected in W16MO animals (Fig. 2, i-n; Fig. S7, a-d, o, p). Moreover shha and Notch receptor expression are unaffected in W16MO animals (Fig. 2, q, r, Fig. S7, a-l). To confirm these pathways are discrete, we examined expression of six known Shh target genes, with particular interest in vegfaa. None of the Shh targets examined showed significantly altered expression (Fig. S14; Table S7). Our results suggest that the Wnt16 and Shh pathways act in parallel upstream of HSC specification.
The predominant model for how Notch signalling regulates HSC specification is that endothelial cells of the dorsal aorta, receive a requisite Notch1-mediated signal1, 21-24. Chimeric mice generated using Notch1-deleted cells show no contribution of knockout cells to the adult haematopoietic system, demonstrating that Notch1 signalling is required cell-autonomously for specification of HSCs1, 21. The relevant Notch ligand(s) are thought to be expressed in the formed dorsal aorta and/or immediately surrounding mesenchyme1, 22-24. A contributing ligand appears to be Jag1, because knockout mice have severely impaired, but not totally abrogated development of HSCs1, 24. Notch1 activation of Runx1 is thought to occur indirectly through Gata2, since the Runx1 promoter does not contain identifiable Rbpjκ binding sites1, 21, 23, 24. Notch signalling defects in wnt16 morphants suggests that the requirement for Dlc/Dld is not the same as this cell-autonomous requirement, because decreased Notch ligand expression is specific to somites (Fig. 3, a-d), whereas expression of dlc is normal in the dorsal aorta of wnt16 morphants (Fig. S12, Table S5) and dld is not natively expressed near the formed dorsal aorta. Moreover, expression of jag1b and its putative target gata2 are unaffected in Wnt16 knock down animals (Fig. 4, a-d).
To better understand whether Dlc/Dld specification of HSCs work cell-autonomously or not, we sought to define the timing of the Notch signalling event lacking in W16MO-injected animals using animals carrying transgenes that allow heat-shock induction of the Notch intracellular domain (NICD), which is a dominant activator of the Notch pathway20, 25. We induced NICD expression at different time points to identify the temporal window when enforced Notch activity could rescue HSC expression in wnt16 morphants. Nuclear, myc-tagged NICD protein is present in transgenic animals by 1 hour post heat shock (hphs), and is strong from 3 hphs (Fig. 4, e-h) through at least 24 hphs (not shown), consistent with prior reports26. Heat-shock induction of NICD at 14 hpf (10-ss) rescued cmyb expression at 36 hpf (Fig. 4, i-k; Table S8), as has been seen previously for rescue of mib20. Surprisingly, heat shock just two hours later (14-ss) did not rescue cmyb expression along the dosal aorta (red circle, Fig. 4, l, Table S8). These results indicate that the critical phase of Notch signalling required for HSC specification downstream of Wnt16, occurs between 15-17 hpf (12-16-ss) and abruptly terminates by about 18-19 hpf (18-20-ss).
Since this timing is well prior to the formation of the dorsal aorta and HSCs from its ventral endothelium, we sought to determine when cells fated to become HSCs first experience a cell-autonomous Notch signal. We first took advantage of a tp1:Kaede Notch reporter line expressing a green-to-red, photoconvertible Kaede protein under the control of a Notch-responsive promoter. Photoconversion of Notch-responsive, Kaede+ cells in the Dlc/Dld-critical window, before 19.5 hpf (21-ss) yielded converted, red fluorescent progeny that contributed to the region near the dorsal aorta at 3 dpf (Fig. 4, m, n), but these cells never became HSCs, as shown by their failure to produce labelled thymic progeny (Fig. 4, m′, n′). In contrast, photoconversion at 24 hpf and later produced both labelled dorsal aorta (Fig. 4, o, p) and thymic immigrants (Fig. 4, o′, p′). Because there is a lag between reception of a Notch signal and the production of mature Kaede protein in the reporter lines, we wanted to confirm when Notch activity appears in pre-haematopoietic cells by a more immediate read-out for Notch-responsiveness. We performed double in situs, in tp1:GFP Notch-reporter transgenic animals, for expression of GFP and etsrp, which labels pre-haematopoietic mesoderm and is required for HSC specification27. The very first double-labelled cells appeared at 18 hpf in the midline (Fig. 4, q, r), and increased in number through at least 24 hpf (Fig. 4, s, t). Taken together, our data suggest that Notch signalling in cells fated to become HSCs begins after 18 hpf and continues well into the second day of development. This timing fits well with the established model of cell-autonomous Notch specification of HSCs, where Notch signalling in nascent HSCs occurs after aortic specification via interactions between Jag1b and Notch11, 21-24. Overall, our results indicate that somitic signalling by Dlc/Dld downstream of Wnt16 is temporally and spatially distinct from the observable cell-autonomous requirement for Notch signalling. Wnt16 therefore controls a previously unappreciated environmental requirement for Notch signalling in the somites, most simply explained by regulation of an unidentified relay signal.
To better understand what cell population Wnt16-regulated somitic Dlc/Dld acts on, we examined fine somite patterning in Wnt16 and Dlc/Dld loss-of-function animals. Since somite segmentation (Fig. 2, a, b, e, f, Fig. 3, a-d) and myotomal specification (Fig. 2, e, f, Fig. 3, a-d) appear to be intact in wnt16 morphants, we examined specification of the sclerotomal somite compartment, which houses vertebral and vascular smooth muscle cell (VSMC) precursors. Sclerotomal markers displayed severely decreased expression, albeit to variable extents, in both W16MO and combined dlc/dld loss of function animals (Fig. S15; Table S9). Although hypochord (Fig. S15, u-y, green arrowheads), was abolished in the dlc/dld double loss of function animals (Fig. S15, x) as has been reported18, hypochord was specified normally in wnt16 morphants (Fig. 2, o, p, Fig. S15, y), emphasizing the fact that the Wnt16-dependent loss of somitic dlc/dld is distinct from global loss. Our results suggest that sclerotome specification or morphogenesis is required for HSC specification.
In sum, our results demonstrate that non-canonical signalling by Wnt16 is required genetically upstream of the combined actions of the Notch pathway ligands Dlc and Dld for HSC specification. Dlc/Dld-mediated Notch signalling is spatially and temporally distinct from previously described cell-autonomous requirements for Notch in HSC specification, pointing to the possibility of a novel relay signal. These data represent the first demonstration that non-canonical Wnt signalling activity is required for HSC specification in vertebrates. Given that Wnt16 in mouse is expressed at similar times of development28 and is expressed in embryoid bodies during commitment to blood and vasculature29, it is feasible that this function is conserved in mammals.
Methods summary
Zebrafish strains
The following strains were maintained in accord with IACUC approved procedures: AB*, Tg(TOP:GFP)w25, Tg(-6.0itga2b:eGFP)la2, Tg(hsp70l:Gal4)1.5kca4, Tg(UAS:myc-Notch1a-intra)kca3, Tg(tp1-MmHbb:EGFP)um14, Tg(tp1-MmHbb:Kaede)um15, Tg(cmyb:EGFP)zf169, Tg(kdrl:RFP)la4, and dlctit446/tit446.
Constructs
Probes and mRNA were synthesized from published constructs or cloned de novo according to standard procedures.
Morpholinos
5 ng W16MO1 AGGTTAGTTCTGTCACCCACCTGTC, W16MO2 GCGTGGAATACTTACATCCAACTTC, W16CoMO AcGTTAGTTgTGTCAgCCAgCTcTC.
Genotyping and PCR
Performed by standard methods using primers and conditions described in the Complete Methods.
Whole mount in situ, antibody staining, microscopy
Performed according to standard methods and fully described in the Complete Methods.
Supplementary Material
Acknowledgments
The authors wish to thank L. Zon, K. Poss, D. Kimelman, M. Lardelli, B. Appel, C. Burns, J. Campos-Ortega, D. Ransom, N. Trede, J. Lewis, M. Pack, S. Holley, C. Moens, B. Paw, R. Karlström, and J. Waxman for probe constructs. L. Zon, R. Dorsky and S. Holley provided transgenic and mutant zebrafish lines. C. Weaver, K. Willert, K.J.P. Griffin, J. Bertrand, D. Stachura, and Y. Lee provided critical evaluation of the manuscript. This research was funded by an AHA Postdoctoral Fellowship 0725086Y to W.K.C., an AHA Predoctoral Founders Affiliate Fellowship 0815732D to J.C.M., NIH R01-HL093467 to N.L., and NIH R01-DK074482, CIRM New Investigator Award, and March of Dimes 6-FY09-508 to D.T.
Footnotes
Supplementary Information accompanies the paper on www.nature.com/nature.
Author Contributions W.K.C. and D.T. designed all experiments. Whole mount immunofluorescence, double fluorescence in situs, and Kaede-based fate mapping was performed by A.D.K. K.G.O. cloned and subcloned multiple constructs. J.C.M and N.L. generated Notch reporter lines. All other experiments were performed by W.K.C. The manuscript was written by W.K.C. and edited by N.L. and D.T., with critical input as described in the Acknowledgments.
Author Information Reprints and permissions information is available at www.nature.com/reprints.
The authors declare no competing interests.
References
- 1.Gering M, Patient R. Notch signalling and haematopoietic stem cell formation during embryogenesis. J Cell Physiol. 2010;222:11–6. doi: 10.1002/jcp.21905. [DOI] [PubMed] [Google Scholar]
- 2.Staal FJ, Luis TC. Wnt signaling in hematopoiesis: crucial factors for self-renewal, proliferation, and cell fate decisions. J Cell Biochem. 2010;109:844–9. doi: 10.1002/jcb.22467. [DOI] [PubMed] [Google Scholar]
- 3.Angers S, Moon RT. Proximal events in Wnt signal transduction. Nat Rev Mol Cell Biol. 2009;10:468–77. doi: 10.1038/nrm2717. [DOI] [PubMed] [Google Scholar]
- 4.Takada S, et al. Wnt-3a regulates somite and tailbud formation in the mouse embryo. Genes Dev. 1994;8:174–89. doi: 10.1101/gad.8.2.174. [DOI] [PubMed] [Google Scholar]
- 5.Goessling W, et al. Genetic interaction of PGE2 and Wnt signaling regulates developmental specification of stem cells and regeneration. Cell. 2009;136:1136–47. doi: 10.1016/j.cell.2009.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.McWhirter JR, et al. Oncogenic homeodomain transcription factor E2A-Pbx1 activates a novel WNT gene in pre-B acute lymphoblastoid leukemia. Proc Natl Acad Sci U S A. 1999;96:11464–9. doi: 10.1073/pnas.96.20.11464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bertrand JY, et al. Haematopoietic stem cells derive directly from aortic endothelium during development. Nature. 2010;464:108–11. doi: 10.1038/nature08738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kissa K, Herbomel P. Blood stem cells emerge from aortic endothelium by a novel type of cell transition. Nature. 2010;464:112–5. doi: 10.1038/nature08761. [DOI] [PubMed] [Google Scholar]
- 9.Boisset JC, et al. In vivo imaging of haematopoietic cells emerging from the mouse aortic endothelium. Nature. 2010;464:116–20. doi: 10.1038/nature08764. [DOI] [PubMed] [Google Scholar]
- 10.Bertrand JY, et al. Definitive hematopoiesis initiates through a committed erythromyeloid precursor in the zebrafish embryo. Development. 2007;134:4147–56. doi: 10.1242/dev.012385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kissa K, et al. Live imaging of emerging hematopoietic stem cells and early thymus colonization. Blood. 2008;111:1147–56. doi: 10.1182/blood-2007-07-099499. [DOI] [PubMed] [Google Scholar]
- 12.Lin HF, et al. Analysis of thrombocyte development in CD41-GFP transgenic zebrafish. Blood. 2005;106:3803–10. doi: 10.1182/blood-2005-01-0179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yokota T, et al. Tracing the first waves of lymphopoiesis in mice. Development. 2006;133:2041–51. doi: 10.1242/dev.02349. [DOI] [PubMed] [Google Scholar]
- 14.Clements WK, Ong KG, Traver D. Zebrafish wnt3 is expressed in developing neural tissue. Dev Dyn. 2009;238:1788–95. doi: 10.1002/dvdy.21977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nygren MK, et al. beta-catenin is involved in N-cadherin-dependent adhesion, but not in canonical Wnt signaling in E2A-PBX1-positive B acute lymphoblastic leukemia cells. Exp Hematol. 2009;37:225–33. doi: 10.1016/j.exphem.2008.10.007. [DOI] [PubMed] [Google Scholar]
- 16.Teh MT, et al. Role for WNT16B in human epidermal keratinocyte proliferation and differentiation. J Cell Sci. 2007;120:330–9. doi: 10.1242/jcs.03329. [DOI] [PubMed] [Google Scholar]
- 17.Cirone P, et al. A role for planar cell polarity signaling in angiogenesis. Angiogenesis. 2008;11:347–60. doi: 10.1007/s10456-008-9116-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Julich D, et al. beamter/deltaC and the role of Notch ligands in the zebrafish somite segmentation, hindbrain neurogenesis and hypochord differentiation. Dev Biol. 2005;286:391–404. doi: 10.1016/j.ydbio.2005.06.040. [DOI] [PubMed] [Google Scholar]
- 19.Holley SA, Julich D, Rauch GJ, Geisler R, Nusslein-Volhard C. her1 and the notch pathway function within the oscillator mechanism that regulates zebrafish somitogenesis. Development. 2002;129:1175–83. doi: 10.1242/dev.129.5.1175. [DOI] [PubMed] [Google Scholar]
- 20.Burns CE, Traver D, Mayhall E, Shepard JL, Zon LI. Hematopoietic stem cell fate is established by the Notch-Runx pathway. Genes Dev. 2005;19:2331–42. doi: 10.1101/gad.1337005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hadland BK, et al. A requirement for Notch1 distinguishes 2 phases of definitive hematopoiesis during development. Blood. 2004;104:3097–105. doi: 10.1182/blood-2004-03-1224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kumano K, et al. Notch1 but not Notch2 is essential for generating hematopoietic stem cells from endothelial cells. Immunity. 2003;18:699–711. doi: 10.1016/s1074-7613(03)00117-1. [DOI] [PubMed] [Google Scholar]
- 23.Robert-Moreno A, Espinosa L, de la Pompa JL, Bigas A. RBPjkappa-dependent Notch function regulates Gata2 and is essential for the formation of intra-embryonic hematopoietic cells. Development. 2005;132:1117–26. doi: 10.1242/dev.01660. [DOI] [PubMed] [Google Scholar]
- 24.Robert-Moreno A, et al. Impaired embryonic haematopoiesis yet normal arterial development in the absence of the Notch ligand Jagged1. Embo J. 2008;27:1886–95. doi: 10.1038/emboj.2008.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Scheer N, Campos-Ortega JA. Use of the Gal4-UAS technique for targeted gene expression in the zebrafish. Mech Dev. 1999;80:153–8. doi: 10.1016/s0925-4773(98)00209-3. [DOI] [PubMed] [Google Scholar]
- 26.Scheer N, Groth A, Hans S, Campos-Ortega JA. An instructive function for Notch in promoting gliogenesis in the zebrafish retina. Development. 2001;128:1099–107. doi: 10.1242/dev.128.7.1099. [DOI] [PubMed] [Google Scholar]
- 27.Ren X, Gomez GA, Zhang B, Lin S. Scl isoforms act downstream of etsrp to specify angioblasts and definitive hematopoietic stem cells. Blood. 2010;115:5338–46. doi: 10.1182/blood-2009-09-244640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kemp C, Willems E, Abdo S, Lambiv L, Leyns L. Expression of all Wnt genes and their secreted antagonists during mouse blastocyst and postimplantation development. Dev Dyn. 2005;233:1064–75. doi: 10.1002/dvdy.20408. [DOI] [PubMed] [Google Scholar]
- 29.Corrigan PM, Dobbin E, Freeburn RW, Wheadon H. Patterns of Wnt/Fzd/LRP gene expression during embryonic hematopoiesis. Stem Cells Dev. 2009;18:759–72. doi: 10.1089/scd.2008.0270. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.