

Abstract
Toxoplasma gondii infections occur worldwide in humans and animals. In immunocompromised or prenatally infected humans, T. gondii can cause severe clinical symptoms. The identification of specific epitopes on T. gondii antigens is essential for the improvement and standardization of the serological diagnosis of toxoplasmosis. We selected 20 peptides mimicking linear epitopes on GRA1, GRA2, GRA4, and MIC3 antigenic T. gondii proteins in silico using the software ABCpred. A further 18 peptides representing previously published epitopes derived from GRA1, SAG1, NTPase1, and NTPase2 antigens were added to the panel. A peptide microarray assay was established to prove the diagnostic performance of the selected peptides with human serum samples. Seropositive human serum samples (n = 184) were collected from patients presenting with acute toxoplasmosis (n = 21), latent T. gondii infection (n = 53), and inactive ocular toxoplasmosis (n = 10) and from seropositive forest workers (n = 100). To adjust the cutoff values for each peptide, sera from seronegative forest workers (n = 75) and patients (n = 65) were used. Univariate logistic regression suggested the significant diagnostic potential of eight novel and two previously published peptides. A test based on these peptides had an overall diagnostic sensitivity of 69% (100% in ocular toxoplasmosis patients, 86% in acutely infected patients, 81% in latently infected patients, and 57% in seropositive forest workers). The analysis of seronegative sera performed with these peptides revealed a diagnostic specificity of 84%. The results of our study suggest that the use of a bioinformatic approach for epitope prediction in combination with peptide microarray testing is a powerful method for the selection of T. gondii epitopes as candidate antigens for serological diagnosis.
INTRODUCTION
Toxoplasma gondii is a widespread protozoan parasite. The seroprevalence of T. gondii infection in humans varies depending on the region or country and is reported to usually range from 30% to 60% in women of child-bearing age (17, 56). The infection is associated with a large spectrum of clinical diseases in both humans and animals. The progression and severity of toxoplasmosis are variable, presumably due to a combination of host and parasite factors (37, 55).
Infection of immunocompromised individuals or prenatally infected persons with the parasite can have severe consequences (43). Congenital transmission from a nonimmune mother to the fetus may cause abortion or may result in the birth of a prenatally infected child. Clinical signs of toxoplasmosis may be evident at birth (e.g., neurological disorders) but can also develop later (e.g., retinochorioiditis). In immunocompetent individuals, T. gondii infection often occurs unnoticed, eventually being associated with lymphadenitis or other flu-like symptoms, but may also cause infectious retinochorioiditis, accounting for 30% to 50% of all cases of posterior uveitis (15, 30).
Serological methods play a major role in the diagnosis of toxoplasmosis. Assays for the detection of T. gondii-specific antibodies are often based on crude antigen extracts (e.g., tachyzoite lysates) or purified native antigenic proteins such as the major surface antigen SAG1 (12, 18, 20, 38). Production of these native antigens is limited, and amounts are difficult to standardize. To avoid these limitations, recombinant antigens were produced and successfully applied in diagnostic assays (32). Potential contaminations with proteins of the organisms used for the production of recombinant antigens (e.g., Escherichia coli) may be a constraint for the use of such preparations (34). Moreover, some recombinant antigens show lower reactivity with specific antibodies than the corresponding native antigens, mainly because of the differences in protein folding that can result in altered epitope presentation (9, 19). Variations in sensitivity and specificity were also observed with respect to the level of recognition of recombinant antigens by human serum samples. This effect might be the result of differences in the methods of production of recombinant proteins in various studies (32).
Several studies have shown that synthetic peptides can be used for the serological diagnosis of viral and bacterial as well as parasitic diseases (1, 31, 36, 44, 46, 54). Large amounts of synthetic peptides can be produced with a high level of purity within a relative short period of time. However, to achieve a satisfactory diagnostic sensitivity and a high specificity, it is necessary to use optimized peptide combinations, mimicking reactive epitopes on natural antigens (36, 53). Peptides can be arranged in microarrays, thus allowing simultaneous analysis of up to several thousands of peptides with a single serum sample. With this technique, global patterns of antibody responses to various infectious agents were studied (3, 35, 36, 42, 44).
Several experimental approaches have been used to identify protein regions and epitopes suitable for the serological diagnosis of T. gondii, including phage display of cDNA libraries, epitope mapping, and reactivity with monoclonal antibodies (5, 11, 26, 29, 41, 49). For this study, a bioinformatic approach was used for in silico prediction of 20 B-cell linear epitopes on the T. gondii antigenic proteins GRA1, GRA2, GRA4, and MIC3. These 20 novel peptides and 18 peptides reported in studies previously published by others (6, 11, 25, 26) were printed into a peptide microarray and validated by analyzing a large number of well-characterized human serum samples.
MATERIALS AND METHODS
Patient sera.
A total of 84 T. gondii-positive human patient serum samples were provided by the Institute of Medical Microbiology and Hospital Hygiene, Heinrich-Heine-University, Düsseldorf, Germany, and the Department of Medical Microbiology and the National Reference Center for Systemic Mycoses, University Medical Center, Göttingen, Germany. Of these, 21 originated from patients with acute toxoplasmosis, 53 from patients with latent T. gondii infection, and 10 from patients with latent ocular toxoplasmosis showing typical lesions in the retina. These institutions also provided 65 samples from serologically T. gondii-negative patients for the study. Detailed information about the serological status of patients with acute or latent T. gondii infection and T. gondii-seronegative patients was published previously by Maksimov et al. (39). Sera collected from serologically T. gondii-positive (n = 100) or -negative (n = 75) forest workers were also used and were described in detail previously (39, 40). The presence or absence of T. gondii-specific antibodies in the sera was confirmed by a latex agglutination test (LAT; see Table S1 in the supplemental material) and T. gondii surface antigen 1 (SAG1) immunoblot analysis (39).
Ethical considerations.
The study reported here was a collaborative work of the Toxonet01 and Toxonet02 projects of the National Research Platform for Zoonoses and was approved by the respective ethical committees of the medical faculties of the University of Düsseldorf (3174; 20 January 2009) and the University of Göttingen (8 June 2009) and by the State Medical Association of Brandenburg (19 April 2010). Serum samples were collected using approved protocols. For the anonymized patient sera provided by the Institute of Medical Microbiology and Hospital Hygiene, Heinrich-Heine-University Düsseldorf, Düsseldorf, Germany, and the Department of Medical Microbiology and the National Reference Center for Systemic Mycoses, University Medical Center Göttingen, Göttingen, Germany, informed consent was obtained verbally that was in agreement with the ethical committee's approved guidelines. All volunteers (forest workers) were included in the study on the basis of written informed consent as described in detail in reference 40.
In silico prediction of T. gondii linear B-cell epitopes by the use of ABCpred.
For the prediction of linear B-cell epitopes on T. gondii protein sequences, the bioinformatic tool ABCpred (http://www.imtech.res.in/raghava/abcpred) was used. This software has been developed with a combination of recurrent neural networks (RNN) and standard feed-forward networks (52). Four T. gondii antigens, i.e., GRA1, GRA2, GRA4, and MIC3, were analyzed in ABCpred for the presence of linear epitopes. Protein sequences were obtained either from the immune epitope database (http://www.immuneepitope.org) or from GenBank.
For the GRA1 protein, a sequence at the amino-terminal region between amino acid positions 160 and 189 was used (linear sequence source: GenBank accession no. ADK88943.1). This region was described as immunogenic for human T. gondii-specific B cells (6).
The GRA2 protein sequence (57) was analyzed for linear epitopes within the signal peptide (amino acid positions 1 to 23) and the amino-terminal region (amino acid positions 24 to 94). This protein sequence was described previously as a region containing major epitopes (29) (linear sequence source: http://www.immuneepitope.org/assayId/8305).
The GRA4 protein sequence, including amino acids 297 to 345 (linear sequence source: http://www.immuneepitope.org/assayId/1244669) at the carboxy-terminal region, was used for selecting specific epitopes. This region has been reported to be well recognized by IgG antibodies from T. gondii-infected humans, mice, and sheep (41).
The entire MIC3 protein sequences (linear sequence source: GenBank accession no. CAB56644 and EEB02693) were also analyzed for potential linear epitopes.
For the prediction of potential epitopes, a threshold value of 0.6 and a window length of 16 amino acids with overlapping filters were used (48).
Published peptide sequences.
A total of 18 previously described peptides representing four different antigens, i.e., SAG1, GRA1, NTP1, and NTP2, were also included in the study.
Two peptides derived from NTPase1 (length = 20 amino acids) and from NTPase2 (length = 18 amino acids) (25) were selected (Table 1).
Table 1.
Peptides used in this study and their diagnostic propertiesa
| Peptide designation | Peptide sequence | Protein | Sensitivity (%) | Specificity (%) | Cutoff value | Reference or source |
|---|---|---|---|---|---|---|
| Peptides from literature | ||||||
| SAG1-48 | SDPPLVANQVVTCPDKKS | SAG1 | 4.35 | 3.57 | 1.87 | 26 |
| dSAG1-62 | DKKSTAAVILTPTENHFT | SAG1 | 3.26 | 3.57 | 1.62 | 26 |
| SAG1-76 | NHFTLKCPKTALTEPPTLA | SAG1 | 2.72 | 3.57 | 2.81 | 26 |
| SAG1-90 | PPTLAYSPNRQICPAGTTS | SAG1 | 3.26 | 3.57 | 2.03 | 26 |
| SAG1-104 | AGTTSSCTSKAVTLSSLIP | SAG1 | 1.63 | 3.57 | 1.29 | 26 |
| SAG1-146 | PIEKFPVTTQTFFWGCIKG | SAG1 | 3.26 | 3.57 | 1.30 | 26 |
| SAG1-273 | AFPAESKSVIIGCTGGSPE | SAG1 | 2.17 | 3.57 | 1.84 | 26 |
| SAG1-287 | GGSPEKHHCTVKLEFAGAA | SAG1 | 5.43 | 3.57 | 0.54 | 26 |
| dSAG1-301 | FAGAAGSAKSAAGTASHVS | SAG1 | 1.09 | 3.57 | 2.28 | 26 |
| SAG1-315 | ASHVSIFAMVIGLIGSIAACVA | SAG1 | 16.85 | 3.57 | 1.13 | 26 |
| SAG1-61 | DKKSTAAVILTPTENHFTL | SAG1 | 9.78 | 3.57 | 1.53 | 11 |
| SAG1-181 | SVVNNVARCSYGADSTLGP | SAG1 | 1.63 | 3.57 | 2.06 | 11 |
| SAG1-241 | SFKDILPKLTENPWQGNAS | SAG1 | 2.72 | 3.57 | 2.60 | 11 |
| SAG1-261 | DKGATLTIKKEAFPAESKS | SAG1 | 2.17 | 3.57 | 1.48 | 11 |
| SAG1-301 | AGAAGSAKSAAGTASHVSI | SAG1 | 0.54 | 3.57 | 1.71 | 11 |
| GRA1-100 | EEVIDTMKSMQRDED | GRA1 | 2.72 | 3.57 | 2.81 | 6 |
| NTP1 | CKAPMIVTGGGMLAA INTLK | NTP1 | 4.35 | 3.57 | 1.82 | 25 |
| NTP2 | APMFITGREMLASIDTLK | NTP2 | 3.26 | 3.57 | 1.91 | 25 |
| Novel peptides | ||||||
| GRA1-162 | MKVIDDVQQLEKDKQQ | GRA1 | 34.24 | 3.57 | 2.83 | This study |
| GRA2-61 | DERQQEPEEPVSQRAS | GRA2 | 30.98 | 3.57 | 1.28 | This study |
| GRA2-28 | GVVNQGPVDVPFSGKP | GRA2 | 50.00 | 3.57 | 0.98 | This study |
| GRA2-67 | PEEPVSQRASRVAEQL | GRA2 | 4.89 | 3.57 | 3.39 | This study |
| GRA2-38 | PFSGKPLDERAVGGKG | GRA2 | 9.24 | 3.57 | 1.71 | This study |
| GRA4-300 | GGTRTSTAPAEAGKTE | GRA4 | 9.24 | 3.57 | 1.06 | This study |
| GRA4-316 | LDDGYRPPPFNPRPSP | GRA4 | 4.35 | 3.57 | 0.95 | This study |
| GRA4-309 | PAEAGKTELDDGYRPP | GRA4 | 33.15 | 3.57 | 1.13 | This study |
| GRA4-321 | PPPFNPRPSPYAELLK | GRA4 | 13.04 | 3.57 | 2.33 | This study |
| MIC3-219 | GETLVNLPEGGQGCKR | MIC3 | 2.72 | 3.57 | 3.19 | This study |
| MIC3-15 | SGAVWMCTPAEALPIQ | MIC3 | 0.00 | 3.57 | 3.08 | This study |
| MIC3-88 | RQLHTDNGYFIGASCP | MIC3 | 1.09 | 3.57 | 2.82 | This study |
| MIC3-324 | GSEGSLSEKMNIVFKC | MIC3 | 10.33 | 3.57 | 1.37 | This study |
| MIC3-238 | HAFRENCSPGRCIDDA | MIC3 | 2.17 | 3.57 | 2.64 | This study |
| MIC3-282 | GVEVTLAEKCEKEFGI | MIC3 | 19.57 | 3.57 | 2.82 | This study |
| MIC3-191 | SKRGNAKCGPNGTCIV | MIC3 | 31.52 | 3.57 | 1.22 | This study |
| MIC3-26 | ALPIQKSVQLGSFDKV | MIC3 | 2.17 | 3.57 | 1.66 | This study |
| MIC3-291 | CEKEFGISASSCKCDN | MIC3 | 7.61 | 3.57 | 1.73 | This study |
| MIC3-8 | LLHALTFSGAVWMCTP | MIC3 | 1.63 | 3.57 | 2.25 | This study |
| MIC3-39 | DKVVPSREVVSESLAP | MIC3 | 2.17 | 3.57 | 1.26 | This study |
Peptide sequences were either previously published (n = 18) or selected after in silico epitope prediction (n = 20). The number at the end of the peptide names indicates the position of the first amino acid of the peptide in the protein sequence. The individual cutoff value for each peptide was selected using the mean index values obtained for a panel of 140 T. gondii-seronegative human sera and accepting a maximum of 4% false-positive reactions.
A peptide mimicking a GRA1-specific epitope (named “epi24”) derived from protein residues 172 to 186 of GRA1 and published by Beghetto et al. was also included in the peptide panel (6) (Table 1).
Finally, a total of 15 SAG1-specific epitopes, which have previously been described by other workers (11, 26), were also selected for the peptide panel (Table 1).
Preparation of peptide microarray slides.
Peptides were synthesized and printed on peptide microarray slides by JPT Peptide Technologies GmbH, Berlin, Germany, essentially as described by Maksimov et al. (39).
Examination of sera by peptide microarray.
Each array block on the slides was first framed with a “Pap Pen” (Kisker Biotech GmbH & Co. KG, Steinfurt, Germany). Arrays were then incubated (60 μl/well) with blocking solution (phosphate-buffered saline [PBS], 0.05% Tween 20, 0.2% I-Block [Applied Biosystems, Bedford, MA]) for 30 min.
Human serum samples (60 μl/well), diluted 1:200 in blocking solution, were incubated at 37°C for 1 h and washed seven times for 3 min each time with PBS-T (PBS [pH 7.2], 0.5% Tween 20) at room temperature. Conjugate (Cy5-AffiniPure donkey anti-human IgG; Fcγ fragment specific; and minimal cross reactions to bovine, horse, and mouse serum proteins) (Jackson ImmunoResearch Laboratories, West Grove, PA) diluted 1:1,000 (1 μg/ml) was added to the wells (60 μl/well), incubated at 37°C for 30 min, and washed as indicated above, followed by three additional washing steps (1 min each) with sterile filtered Milli-Q water. Afterwards, the slides were spun dry for 10 s using a slide spinner (DW-41MA-230; Qualitron Inc./Eppendorf, Berzdorf, Germany).
Scanning and measurement of spot signal intensities and data extraction.
Scanning and evaluation of microarray slides as well as data extraction were performed essentially as described previously (39, 44, 45).
Peptide microarray data analysis.
To analyze the raw data (median of signal intensity) in GPR files, index values (IVs) were recovered for each peptide spot as the log2 of the quotient of the medians of foreground and background values (39, 44, 45). Each serum sample was analyzed on a single block, with the peptides printed in triplicate on each block. To obtain the serum-specific spectrum for each peptide, the means of the IVs for each peptide spot per block (mean sample index value [MSIV]) were calculated using the “corrected mean” formula (Microsoft Office Excel 2003) to exclude artifacts, i.e., false-positive and -negative signals within the replicas in each block (39). The peptide microarrays used in this study failed to meet the criteria required for submission to MIAME-based public databases, as the data types of biomolecular interactions and parameters studied and the protocols, as well as the types of information extracted from the microarray experiment, are different from those of standard DNA microarray experiments (47, 58). Therefore, the MSIVs for all sera and peptides are presented as supplemental material (see Table S1 in the supplemental material).
To ensure the specificity of all of the peptides used, we established an individual cutoff value for each peptide to classify the result of the reaction performed with each particular peptide as positive or negative. The cutoff value was selected for each peptide separately by using the MSIVs obtained for a panel of 140 T. gondii-seronegative human serum samples and accepting a maximum of 4% false-positive reactions.
Statistical analysis.
Fisher's exact test and logistic regression (LR) were computed with R, version 2.8.1 (R Foundation for Statistical Computing, Vienna, Austria; ISBN 3-900051-07-0; http://www.R-project.org). Linear regression analysis and the Wilcoxon signed-rank test were performed with STATISTICA 8 (StatSoft, Tulsa, OK). This program was also used to produce figures. P values < 0.05 were regarded as statistically significant. Univariate LR analysis (R, package “epicalc”) was performed to assess the diagnostic potential of tested peptides. A P value (Wald's test) < 0.05 was regarded as statistically significant (13). The R-package “stats” was used for analysis of variance (ANOVA) in calculations of peptide and serum reactivity.
To perform multiple comparisons of means determined from all index values, a post hoc test (LSD [least significant difference]-Bonferroni test) of ANOVA results was applied using the R package “agricolae.” The differences between means of positive peptide reaction values determined in the analysis of peptide and serum reactivities within tested groups were accepted as significant when the differences were equal to or higher than the LSD values as corrected by the Bonferroni method (10).
RESULTS
Diagnostic specificity and peptide-specific reactivity of sera.
Twenty candidate B-cell linear epitopes (Table 1) derived from the antigenic proteins GRA1, GRA2, GRA4, and MIC3 were selected in silico using the computer program ABCpred. The respective peptides containing these epitopes (referred to here as novel peptides) and 18 additional peptides with published sequences were synthesized.
All sera of seronegative patients (n = 65) and 75 randomly selected sera of seronegative forest workers were used to establish cutoff values for each peptide. Individual application of these peptide-specific cutoff values resulted in a diagnostic specificity for T. gondii-seronegative sera of 96.6% for each individual peptide.
Sera of patients with acute signs of T. gondii infection recognized statistically significantly more peptides than the sera of seropositive forest workers (Wilcoxon signed-rank test, P = 0.0003; Fig. 1). Similarly, sera of patients with latent T. gondii infection or ocular signs of toxoplasmosis recognized a significantly higher number of peptides than sera of serologically positive forest workers (Wilcoxon signed-rank test; P < 0.000 or P = 0.001, respectively) (Fig. 1). The differences between the results determined for the groups of patients with ocular, acute, or latent toxoplasmosis were not statistically significant (Wilcoxon signed-rank test, P > 0.6) (Fig. 1).
Fig 1.

Number of peptides recognized by individual T. gondii-positive sera. Sera from forest workers recognized a significantly lower number of peptides per serum sample compared to sera from acutely or latently infected patients or patients with ocular toxoplasmosis (OT patients) (Wilcoxon signed-rank test; P ≤ 0.001). No significant differences were observed within the three patient groups (acute, latent, and ocular toxoplasmosis) concerning the number of recognized peptides per serum sample.
The sera of 72 of 84 (86%) T. gondii-seropositive patients with acute, latent, or ocular toxoplasmosis and 66 of 100 (66%) T. gondii-seropositive forest workers recognized at least 1 of the 38 peptides. The remaining 46 of 184 (25%) sera failed to react with any of the 38 peptides. Linear regression analysis revealed a significant positive correlation between reciprocal LAT titers and the number of novel peptides recognized by the individual sera (r2 = 0.24, P < 0.0001; Fig. 2A). No significant correlation was found between the reciprocal LAT titers and the number of peptides taken from the literature that were recognized by the individual sera (r2 = 0.008, P = 0.1; Fig. 2B).
Fig 2.
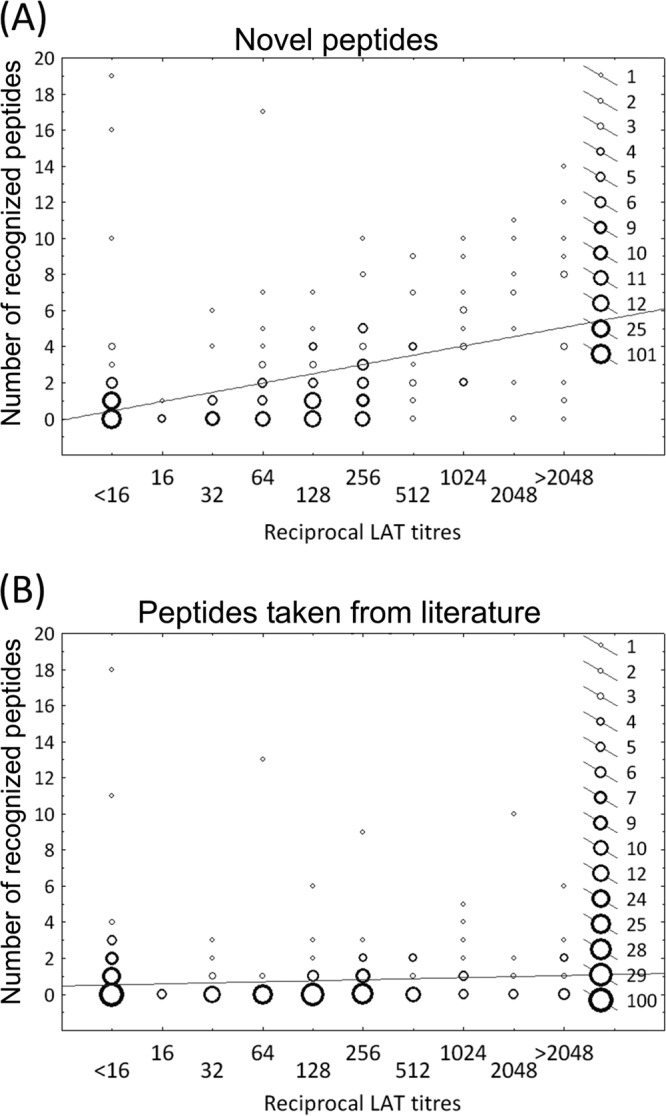
Latex agglutination test (LAT) titers and the numbers of recognized peptides in all seronegative and seropositive human serum samples (n = 324) were analyzed for potential correlations by linear regression analysis. (A) Novel peptides: statistically significant correlation (r2 = 0.24; P < 0.0001). (B) Peptides taken from the literature: no significant correlation (r2 = 0.008; P = 0.1).
Individual diagnostic sensitivities of novel peptides predicted in silico.
Fourteen of the 20 novel peptides were recognized by at least one serum sample from the patients with acute and ocular toxoplasmosis. Individual peptide sensitivities in these two serum panels (acute and ocular) ranged from 0% to 86% and from 0% to 100%, respectively (Fig. 3A and B).
Fig 3.

Diagnostic sensitivity of 20 novel peptides predicted in silico in this study in groups of human serum samples (A) from patients with acute toxoplasmosis (n = 21), (B) from patients with nonactive ocular toxoplasmosis (OT patients) (n = 10), (C) from patients with latent toxoplasmosis (n = 53), and (D) from serologically positive forest workers (n = 100).
Sera from patients with latent T. gondii infection recognized 19 of the 20 novel peptides. The individual diagnostic sensitivities of the peptides ranged from 0% to 60% (Fig. 3C). The lowest number of peptides (13 of 20) was recognized by sera from forest workers, with a range of individual peptide sensitivities of 1% to 30% (Fig. 3D). Taken together for all sera (n = 184), the individual diagnostic sensitivities determined for each novel peptide ranged from 0% to 49% (Table 1).
The GRA2–28 peptide was the most reactive of the novel peptides and was recognized by 100% of the serum samples from the ocular toxoplasmosis patients, 85.7% of the serum samples from patients with acute infections, 60.3% of the serum samples from latently infected patients, and 30% of the serum samples from T. gondii-seropositive forest workers (Fig. 3A, B, C, and D, respectively). GRA-derived peptides were statistically significantly more often recognized than MIC3-derived peptides (Wilcoxon signed-rank test, P < 0.0001). One peptide (MIC3–15) failed to show a positive reaction with any of the analyzed sera (Table 1).
Individual diagnostic sensitivities of peptides taken from the literature.
All 18 peptides taken from the literature were recognized by sera from latently infected patients, with peptide-specific sensitivities of 2% to 30% (Fig. 4C). Sera from ocular toxoplasmosis patients and forest workers recognized 11 of these 18 peptides, with individual peptide sensitivities of 0% to 20% (Fig. 4B and D). The lowest number of peptides (7 of 18) was recognized by sera from patients with acute signs of toxoplasmosis, with individual peptide sensitivities ranging from 0% to 38% (Fig. 4A).
Fig 4.

Diagnostic sensitivity of 18 peptides taken from the literature in groups of human serum samples (A) from patients with acute toxoplasmosis (n = 21), (B) from patients with nonactive ocular toxoplasmosis (OT patients) (n = 10), (C) from patients with latent toxoplasmosis (n = 53), and (D) from serologically positive forest workers (n = 100).
Among the 18 previously published peptides, SAG1–315 was the peptide most often recognized in all tested serum groups. The diagnostic sensitivities seen with this peptide ranged from 5% (forest workers) to 38% (acutely infected patients). The individual diagnostic sensitivities of each peptide with respect to all sera (184 sera per peptide) ranged between 1% and 17%.
SAG1-derived peptides showed reactivity that was statistically significantly higher than the reactivity shown by the NTPase-derived peptides (Wilcoxon signed-rank test, P < 0.0001; Table 1).
Diagnostic sensitivity and specificity of an optimized peptide array.
The diagnostic potential of all 38 tested peptides was analyzed in relation to LAT seropositivity by univariate logistic regression (LR). LR revealed a statistically significant association of the results obtained for eight novel and two published peptides in relation to the serological status of sera in LAT (Table 2). For all serologically positive individuals, the peptide-specific diagnostic sensitivity within this panel of 10 peptides ranged from 9.78% (SAG1–60) to 50% (GRA2–28) (Table 1).
Table 2.
Univariate logistic regression analysis to examine potential relationships between seropositivity in the latex agglutination test (LAT) and positive reactions with individual peptidesa
| Peptide name | Odds ratio (95% CI) | P (Wald's test) |
|---|---|---|
| Peptides from literature | ||
| SAG1-48 | 1.23 (0.39-3.84) | 0.725 |
| SAG1-62 | 0.91 (0.27-3.05) | 0.879 |
| SAG1-76 | 0.75 (0.21-2.66) | 0.661 |
| SAG1-90 | 0.91 (0.27-3.05) | 0.879 |
| SAG1-104 | 0.45 (0.11-1.91) | 0.337 |
| SAG1-146 | 0.45 (0.11-1.91) | 0.277 |
| SAG1-273 | 0.6 (0.16-2.28) | 0.453 |
| SAG1-287 | 1.55 (0.52-4.65) | 0.432 |
| dSAG1-301 | 0.3 (0.06-1.55) | 0.15 |
| SAG1-315 | 5.47 (2.07-14.47) | <0.001 |
| SAG1-61 | 2.93 (1.06-8.09) | 0.038 |
| SAG1-181 | 0.45 (0.11-1.91) | 0.277 |
| SAG1-241 | 0.75 (0.21-2.66) | 0.661 |
| SAG1-261 | 0.6 (0.16-2.28) | 0.453 |
| SAG1-301 | 0.15 (0.02-1.28) | 0.082 |
| GRA1-100 | 0.75 (0.21-2.66) | 0.661 |
| NTP1 | 1.23 (0.39-3.84) | 0.725 |
| NTP2 | 0.91 (0.27-3.05) | 0.879 |
| Novel peptides | ||
| GRA1-162 | 14.06 (5.47-36.1) | <0.001 |
| GRA2-61 | 12.12 (4.71-31.2) | 0.045 |
| GRA2-28 | 27 (10.57-68.99) | <0.001 |
| GRA2-67 | 1.39 (0.45-4.24) | 0.564 |
| GRA2-38 | 2.75 (0.99-7.64) | 0.053 |
| GRA4-300 | 2.75 (0.99-7.64) | 0.053 |
| GRA4-316 | 1.23 (0.39-3.84) | 0.725 |
| GRA4-309 | 13.39 (5.21-34.41) | <0.001 |
| GRA4-321 | 4.05 (1.5-10.9) | 0.006 |
| MIC3-219 | 0.75 (0.21-2.66) | 0.661 |
| MIC3-15 | 0 (0–infinity) | 0.981 |
| MIC3-88 | 0.3 (0.06-1.55) | 0.15 |
| MIC3-324 | 3.11 (1.13-8.55) | 0.028 |
| MIC3-238 | 0.6 (0.16-2.28) | 0.453 |
| MIC3-282 | 6.57 (2.5-17.22) | <0.001 |
| MIC3-191 | 12.43 (4.83-31.98) | <0.001 |
| MIC3-26 | 0.6 (0.16-2.28) | 0.453 |
| MIC3-291 | 2.22 (0.78-6.33) | 0.134 |
| MIC3-8 | 0.45 (0.11-1.91) | 0.277 |
| MIC3-39 | 0.6 (0.16-2.28) | 0.453 |
Statistically significant (P < 0.05) associations are marked (bold). CI, confidence interval.
An optimized array using a combination of these 10 peptides had a diagnostic sensitivity of 100% in ocular toxoplasmosis patients, 86% in acutely infected patients, 81% in latently infected patients, and 57% in seropositive forest workers when the array was regarded as positive when at least one of the peptides was recognized.
The overall sensitivity mounted to 69% when sera were not grouped.
Analysis of seronegative sera (n = 140) revealed a specificity of 84% for this optimized peptide array.
Reaction intensity of peptides with diagnostic potential.
Differences in the reaction intensities within the group of the 10 peptides with diagnostic potential were analyzed by ANOVA and an LSD post hoc test. Reactions performed with GRA1–162 showed the highest statistically significant intensities, i.e., the highest MSIVs, followed by MIC3–282 and GRA2–28 (LSD ≥ 0.458). Peptides SAG1–61, SAG1–315, and MIC3–324 displayed the lowest means (LSD ≥ 0.458) of the reaction intensities (Fig. 5A).
Fig 5.

Intensity index values for peptides with diagnostic potential among tested T. gondii-seropositive human serum samples. The differences between the means of the sample index values (MSIVs) for intensity for single peptides and those for human groups (acutely infected patients, latently infected patients, patients with ocular toxoplasmosis [OT], and seropositive forest workers) determined with 10 peptides with diagnostic potential were regarded as statistically significant when the differences were equal to or higher than the LSD values. Different letters above the whiskers indicate significant differences between the mean intensities in the post hoc LSD test ([A] LSD ≥ 0.458; [B] LSD ≥ 0.331). Whiskers in bar plots represent 95% confidence intervals of the MSIVs. (A) MSIVs of the reactions performed with 10 peptides with diagnostic potential observed for seropositive humans. (B) MSIVs for sera from different groups of humans determined with 10 peptides with diagnostic potential.
ANOVA and LSD test analyses further showed that sera from acutely infected patients recognized the 10 peptides with the highest statistically significant MSIVs (LSD ≥ 0.331) compared to the sera from the other groups (Fig. 5B). Patients presenting with latent and ocular T. gondii infection revealed MSIVs (LSD ≥ 0.331) that were significantly higher than those of serologically positive forest workers. No statistical differences were observed between patients presenting with ocular and latent toxoplasmosis (LSD ≥ 0.331) (Fig. 5B).
One novel peptide (GRA2–28) was recognized with higher statistically significant MSIVs (LSD ≥ 1.09) by sera of acutely infected patients compared to those determined for the members of the other groups (Fig. 6). Two further novel peptides (MIC3–282 and MIC3–191) were recognized with significantly higher MSIVs by sera of acutely infected patients compared to latently infected patients and seropositive forest workers (LSD ≥ 1.09 for MIC3–282; LSD > 0.95 for MIC3–191) (Fig. 6). Sera of patients with ocular toxoplasmosis showed no statistically significant differences in their reactivity with these two peptides compared to sera of acutely infected patients and of patients with latent toxoplasmosis (LSD ≥ 1.09 for MIC3–282; LSD > 0.95 for MIC3–191) (Fig. 6). For all 10 peptides with diagnostic potential, the sera from forest workers showed the lowest mean MSIV index values relative to those from the members of the other groups.
Fig 6.

Differences between various groups of humans regarding the reactivity of sera with peptides suitable for T. gondii diagnosis. The differences between the MSIVs among groups of humans (acutely infected patients, latently infected patients, patients with ocular toxoplasmosis [OT], and seropositive forest workers) for each peptide with diagnostic potential were regarded as statistically significant when the differences were equal to or higher than the LSD values. Different letters on the bars indicate statistically significant differences between the mean intensities in the post hoc LSD-Bonferroni test. Whiskers in bar plots represent 95% confidence intervals of the MSIVs. Peptide GRA2-28 was recognized by the sera from acutely infected patients with a statistically significant higher intensity than by sera from other groups. MIC3–282 and MIC3–191 peptides were recognized with statistically significant higher index values by sera from acutely infected patients than by sera from ocular toxoplasmosis patients and seropositive forest workers. For all 10 peptides with diagnostic potential, the sera from the forest workers displayed the lowest MSIVs relative to the sera from other groups.
DISCUSSION
A number of T. gondii proteins, including SAG1, NTPase1, NTPase2, GRA1, GRA2, and GRA4, have long been known to possess high antigenicity and are suitable antigens for the serological diagnosis of T. gondii infection (2, 7, 11, 16, 21, 24–27, 33, 41). We therefore selected these proteins to identify novel diagnostic peptides for the serological diagnosis of human toxoplasmosis.
To identify T. gondii epitopes with diagnostic potential, we predicted potentially suitable epitopes by the use of the bioinformatic software ABCpred. A variety of computing methods for the prediction of epitopes (e.g., PREDITOPE, PEOPLE, BEPITOPE, BcePred, AAP, BepiPred, the Hopp and Woods method, the Jameson-Wolf antigenicity index, etc.) have been developed in the past (14, 50). Most of them utilize the physical and chemical properties of individual amino acids for the prediction of structural and functional properties of the peptide chain suggesting the localization of epitopes. However, the proportion of correct predictions made with these tools was only marginally better than that seen with random epitope selection (8).
Saha and Raghava (52) developed ABCpred, a software program designed to predict linear B-cell epitopes, using a combination of recurrent and artificial neural networks. This software was trained with a data set of 700 experimentally detected B-cell epitopes from the Bcipep database and 700 random peptides from the Swiss-Prot database. Thereafter, ABCpred showed a better prediction rate than other tools, and the results obtained were significantly better than those obtained by random selection (48, 52). Our study supports these findings, as almost all (19 of 20) ABCpred-selected T. gondii peptides were recognized by various groups of serologically T. gondii-positive human serum samples, and 40% of 20 ABCpred-predicted linear B-cell epitopes appeared to possess diagnostic potential. However, considerable variation in the respective diagnostic sensitivities was observed that depended on the individual peptide and the serum panel analyzed. The moderate success in predicting such epitopes was expected, since the ABCpred developers described a prediction accuracy of 65.93% by this method (52). Since we analyzed the protein sequences without prior knowledge of the linear B-cell epitopes and nonepitopes, we cannot directly compare our data on the proportion of correctly predicted epitopes with the prediction accuracy reported for ABCpred.
All previously described peptides, which included mainly SAG1-derived peptides, were recognized by the human serum samples with lower diagnostic sensitivity. This was unexpected, as a large proportion of anti-T. gondii antibodies in infected humans and animals are directed against SAG1. However, it is known that most B-cell epitopes on the SAG1 antigen are conformation dependent (9, 28, 33, 51), whereas we analyzed only linear epitopes in this study.
This could therefore be one explanation for the low reactivity of SAG1-derived peptides in our study. However, SAG1 peptide sequences had diagnostic sensitivity in our hands that was lower than that reported previously (11). Since Cardona et al. (11) conducted their study in South America, differences between South America and Europe in the prevailing clonal types of T. gondii appear to be the most likely reason for this discrepancy. Moreover, the results might also be biased by differences in the major histocompatibility complex (MHC) haplotype distributions in the human populations from South America and Europe (22, 23, 37).
We also observed lower reactivity of peptide GRA1–100 (also called “epi-24”), which was previously described as highly immunoreactive by Beghetto and colleagues (6). In our hands, GRA1–100 had a diagnostic sensitivity of only 2.7% (5/184), while 29 of 40 T. gondii-positive sera reacted with GRA1–100 in the study of Beghetto et al. (6). This discrepancy with respect to the diagnostic sensitivity of GRA1–100 might be explained by differences in the methods by which the antigens were produced. We applied synthetic peptides, whereas Beghetto and colleagues used a recombinant protein (6). The oligopeptide produced in an expression vector may be folded in a fashion different from that of synthetic peptides or might carry secondary modifications, which could also explain differences in antigenicity.
The diagnostic sensitivities of individual peptides analyzed in our study were limited, not exceeding 50% within the group of all serologically positive (i.e., all LAT-positive) human serum samples. Furthermore, we observed that each T. gondii-positive serum recognized an individual peptide pattern. Thus, our results suggest that it is necessary to use several peptides at the same time, i.e., a peptide array, for the serological diagnosis of T. gondii infections to achieve a level of sensitivity similar to the sensitivity of conventional serological diagnostic tests. Similar effects were also observed when synthetic peptides were used as diagnostic antigens for human echinococcosis and tuberculosis studies (36, 44, 53).
Univariate LR analysis identified 10 peptides for which the serological results showed a statistically significant concordance with those obtained with established serological tests. These peptides contain epitopes from the T. gondii antigenic proteins SAG1, GRA1, GRA2, and GRA4. The peptide-specific diagnostic sensitivities of the individual peptides ranged from 9.78% to 50%.The diagnostic sensitivity of an array based on the 10 peptides ranged between 79% and 100%, depending on the group of sera analyzed, while the diagnostic specificity was 84%. These values are acceptable, though far from optimal. In any case, the 10 peptides may in the future be included in a larger multiple antigenic peptide array for detecting T. gondii infection in patients. Further suitable peptides need to be identified to improve sensitivity. The specificity of a future peptide microarray could be increased by applying more stringent thresholds, i.e., by increasing the cutoffs for individual peptides or by increasing the number of peptides that must be recognized by an individual serum for it to be classified as testing positive. To increase the number of peptide candidates for an optimized peptide microarray, additional immunoreactive antigenic proteins, such as rhoptry (ROP1, ROP2), microneme (MIC2, MIC4, MIC5), dense granule (GRA1, GRA2, GRA4, GRA6, GRA7, GRA8), and surface antigens (SAG2A), should be taken into consideration for the identification of linear B-cell epitopes. If such an optimized array can be established, it may prove superior to conventional serological tests, because the epitopes recognized by sera from individual patients are unambiguously defined. The peptide recognition pattern may allow further conclusions regarding the status of the patient, e.g., differentiation of the acute stage versus chronic stage of infection. A T. gondii peptide array may also represent a powerful tool to compare serological reactions, e.g., reactions of a mother and her child, to find evidence for congenital transmission. Another field of application could be the differential examination of serum and intraocular fluids of patients with suspected ocular toxoplasmosis to confirm or rule out active infections of the eye.
Even within our small array, certain antipeptide reactions could be associated with the acute stage of infection. Sera from acutely infected patients generally recognized peptides with significantly higher index values than sera from other groups. Moreover, we found that particular peptides (e.g., GRA2–28, MIC3–282, MIC3–191) were recognized with a significantly higher intensity by sera from acutely infected patients than by sera from others, indicating that these peptides may be candidates for a predictive peptide panel for the diagnosis of acute toxoplasmosis in humans.
The present report demonstrates that ABCpred is an appropriate bioinformatical tool for use in the selection of potential epitope candidates. However, we also observed the prediction of epitopes that show low reactivity in the analysis with T. gondii-positive human serum samples. False-positive epitope prediction was also reported by the developers of the tool and can be explained, e.g., by the use of a fixed amino acid length in predicting epitopes (52). Moreover, most epitopes analyzed in this study were continuous, whereas the majority of naturally recognized epitopes are discontinuous (4). Because of this, it may also make sense to search for discontinuous epitopes, e.g., by using bioinformatical software for the analysis of the three-dimensional (3D) X-ray structure of an antigen of interest. Unfortunately, there are only limited 3D X-ray protein structure data for T. gondii antigenic proteins available. Nevertheless, our report shows that the use of a bioinformatic prediction method in combination with a peptide microarray assay is a powerful tool for selection and analysis of T. gondii epitopes as candidate antigens for serological diagnosis.
Supplementary Material
ACKNOWLEDGMENTS
This work was supported by the German Federal Ministry of Education and Research (Toxonet01 and Toxonet02) by funds to G.S. (01KI0765 and 01KI1002F), W.D. (01KI0764 and 01KI1002E), U.P. (01KI1002G), and U.G. (01KI0766 and 01KI1002B).
We acknowledge the excellent technical assistance of Andrea Bärwald, Lieselotte Minke, and Robert Carus. We thank Andreas Fröhlich for discussions about the statistical analysis of our study results.
Footnotes
Published ahead of print 11 April 2012
Supplemental material for this article may be found at http://cvi.asm.org/.
REFERENCES
- 1. Alcaro MC, Peroni E, Rovero P, Papini AM. 2003. Synthetic peptides in the diagnosis of HIV infection. Curr. Protein Pept. Sci. 4: 285–290 [DOI] [PubMed] [Google Scholar]
- 2. Altcheh J, et al. 2006. Kinetic analysis of the humoral immune response against 3 Toxoplasma gondii-recombinant proteins in infants with suspected congenital toxoplasmosis. Diagn. Microbiol. Infect. Dis. 56: 161–165 [DOI] [PubMed] [Google Scholar]
- 3. Bacarese-Hamilton T, Mezzasoma L, Ardizzoni A, Bistoni F, Crisanti A. 2004. Serodiagnosis of infectious diseases with antigen microarrays. J. Appl. Microbiol. 96: 10–17 [DOI] [PubMed] [Google Scholar]
- 4. Barlow DJ, Edwards MS, Thornton JM. 1986. Continuous and discontinuous protein antigenic determinants. Nature 322: 747–748 [DOI] [PubMed] [Google Scholar]
- 5. Beghetto E, et al. 2003. Use of an immunoglobulin G avidity assay based on recombinant antigens for diagnosis of primary Toxoplasma gondii infection during pregnancy. J. Clin. Microbiol. 41: 5414–5418 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Beghetto E, et al. 2001. Identification of a human immunodominant B-cell epitope within the GRA1 antigen of Toxoplasma gondii by phage display of cDNA libraries. Int. J. Parasitol. 31: 1659–1668 [DOI] [PubMed] [Google Scholar]
- 7. Beghetto E, et al. 2003. Molecular dissection of the human B-cell response against Toxoplasma gondii infection by lambda display of cDNA libraries. Int. J. Parasitol. 33: 163–173 [DOI] [PubMed] [Google Scholar]
- 8. Blythe MJ, Flower DR. 2005. Benchmarking B cell epitope prediction: underperformance of existing methods. Protein Sci. 14: 246–248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Burg JL, Perelman D, Kasper LH, Ware PL, Boothroyd JC. 1988. Molecular analysis of the gene encoding the major surface antigen of Toxoplasma gondii. J. Immunol. 141: 3584–3591 [PubMed] [Google Scholar]
- 10. Cabral HJ. 2008. Multiple comparisons procedures. Circulation 117: 698–701 [DOI] [PubMed] [Google Scholar]
- 11. Cardona N, de la Torre A, Siachoque H, Patarroyo MA, Gomez-Marin JE. 2009. Toxoplasma gondii: P30 peptides recognition pattern in human toxoplasmosis. Exp. Parasitol. 123: 199–202 [DOI] [PubMed] [Google Scholar]
- 12. Chaves-Borges FA, Souza MA, Silva DA, Kasper LH, Mineo JR. 1999. Detection of Toxoplasma gondii soluble antigen, SAG-1(p30), antibody and immune complex in the cerebrospinal fluid of HIV positive or negative individuals. Rev. Inst. Med. Trop. Sao Paulo 41: 329–338 [DOI] [PubMed] [Google Scholar]
- 13. Domínguez-Almendros S, Benítez-Parejo N, Gonzalez-Ramirez AR. 2011. Logistic regression models. Allergol. Immunopathol. (Madr.) 39: 295–305 [DOI] [PubMed] [Google Scholar]
- 14. El-Manzalawy Y, Honavar V. 2010. Recent advances in B-cell epitope prediction methods. Immunome Res. 6(Suppl. 2): S2 doi:10.1186/1745-7580-6-S2-S2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Garweg JG, et al. 2005. Congenital ocular toxoplasmosis—observations on the outcome after an early diagnosis. Klin. Monbl. Augenheilkd. 222: 721–727 [DOI] [PubMed] [Google Scholar]
- 16. Golkar M, et al. 2007. The dense granule protein GRA2, a new marker for the serodiagnosis of acute Toxoplasma infection: comparison of sera collected in both France and Iran from pregnant women. Diagn. Microbiol. Infect. Dis. 58: 419–426 [DOI] [PubMed] [Google Scholar]
- 17. Groß U. 2004. Prevalence and public-health-aspects of toxoplasmosis. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz 47: 692–697 (In German.) [DOI] [PubMed] [Google Scholar]
- 18. Gross U, Roos T, Appoldt D, Heesemann J. 1992. Improved serological diagnosis of Toxoplasma gondii infection by detection of immunoglobulin A (IgA) and IgM antibodies against P30 by using the immunoblot technique. J. Clin. Microbiol. 30: 1436–1441 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Harning D, Spenter J, Metsis A, Vuust J, Petersen E. 1996. Recombinant Toxoplasma gondii surface antigen 1 (P30) expressed in Escherichia coli is recognized by human Toxoplasma-specific immunoglobulin M (IgM) and IgG antibodies. Clin. Diagn. Lab. Immunol. 3: 355–357 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Hosseininejad M, Azizi HR, Hosseini F, Schares G. 2009. Development of an indirect ELISA test using a purified tachyzoite surface antigen SAG1 for sero-diagnosis of canine Toxoplasma gondii infection. Vet. Parasitol. 164: 315–319 [DOI] [PubMed] [Google Scholar]
- 21. Ismael AB, Sekkai D, Collin C, Bout D, Mévélec MN. 2003. The MIC3 gene of Toxoplasma gondii is a novel potent vaccine candidate against toxoplasmosis. Infect. Immun. 71: 6222–6228 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Jamieson SE, et al. 2009. Host genetic and epigenetic factors in toxoplasmosis. Mem. Inst. Oswaldo Cruz 104: 162–169 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Jamieson SE, et al. 2008. Genetic and epigenetic factors at COL2A1 and ABCA4 influence clinical outcome in congenital toxoplasmosis. PLoS One 3: e2285 doi:10.1371/journal.pone.0002285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Jiang T, et al. 2008. Evaluation of a recombinant MIC3 based latex agglutination test for the rapid serodiagnosis of Toxoplasma gondii infection in swines. Vet. Parasitol. 158: 51–56 [DOI] [PubMed] [Google Scholar]
- 25. Johnson MS, Broady KW, Johnson AM. 1999. Differential recognition of Toxoplasma gondii recombinant nucleoside triphosphate hydrolase isoforms by naturally infected human sera. Int. J. Parasitol. 29: 1893–1905 [DOI] [PubMed] [Google Scholar]
- 26. Kato M, et al. 2007. Reactivity of synthetic SAG1 (p30) peptide sequences with RH, S273 and Beverley strain-induced anti-Toxoplasma gondii antibodies. Pathobiology 74: 50–56 [DOI] [PubMed] [Google Scholar]
- 27. Kato M, et al. 2005. Toxoplasma gondii antigens GRA1 (p24) and SAG1 (p30): a comparison of their stimulatory influence on T-cell activation and cytokine expression in in vitro cultures. Pathobiology 72: 160–164 [DOI] [PubMed] [Google Scholar]
- 28. Kim K, Bülow R, Kampmeier J, Boothroyd JC. 1994. Conformationally appropriate expression of the Toxoplasma antigen SAG1 (p30) in CHO cells. Infect. Immun. 62: 203–209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Klaren VN, Peek R. 2001. Evidence for a compartmentalized B cell response as characterized by IgG epitope specificity in human ocular toxoplasmosis. J. Immunol. 167: 6263–6269 [DOI] [PubMed] [Google Scholar]
- 30. Kodjikian L, et al. 2006. Ocular manifestations in congenital toxoplasmosis. Graefes Arch. Clin. Exp. Ophthalmol. 244: 14–21 [DOI] [PubMed] [Google Scholar]
- 31. Kong JT, Grigg ME, Uyetake L, Parmley S, Boothroyd JC. 2003. Serotyping of Toxoplasma gondii infections in humans using synthetic peptides. J. Infect. Dis. 187: 1484–1495 [DOI] [PubMed] [Google Scholar]
- 32. Kotresha D, Noordin R. 2010. Recombinant proteins in the diagnosis of toxoplasmosis. APMIS 118: 529–542 [DOI] [PubMed] [Google Scholar]
- 33. Lekutis C, Ferguson DJ, Grigg ME, Camps M, Boothroyd JC. 2001. Surface antigens of Toxoplasma gondii: variations on a theme. Int. J. Parasitol. 31: 1285–1292 [DOI] [PubMed] [Google Scholar]
- 34. Li S, et al. 2000. Serodiagnosis of recently acquired Toxoplasma gondii infection with a recombinant antigen. J. Clin. Microbiol. 38: 179–184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Liang L, et al. 2011. Identification of potential serodiagnostic and subunit vaccine antigens by antibody profiling of toxoplasmosis cases in Turkey. Mol. Cell. Proteomics 10: M110.006916 doi:10.1074/mcp.M110.006916 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. List C, et al. 2010. Serodiagnosis of Echinococcus spp. infection: explorative selection of diagnostic antigens by peptide microarray. PLoS Negl. Trop. Dis. 4: e771 doi:10.1371/journal.pntd.0000771 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Lüder CG, Seeber F. 2001. Toxoplasma gondii and MHC-restricted antigen presentation: on degradation, transport and modulation. Int. J. Parasitol. 31: 1355–1369 [DOI] [PubMed] [Google Scholar]
- 38. Maksimov P, et al. 2011. Serological survey and risk factors for Toxoplasma gondii in domestic ducks and geese in Lower Saxony, Germany. Vet. Parasitol. 182: 140–149 [DOI] [PubMed] [Google Scholar]
- 39. Maksimov P, et al. 2012. Analysis of clonal type-specific antibody reactions in Toxoplasma gondii seropositive humans from Germany by peptide-microarray. PLoS One 7: e34212 doi:10.1371/journal.pone.0034212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Mertens M, et al. 2011. Seroprevalence study in forestry workers of a non-endemic region in eastern Germany reveals infections by Tula and Dobrava-Belgrade hantaviruses. Med. Microbiol. Immunol. 200: 263–268 [DOI] [PubMed] [Google Scholar]
- 41. Mévélec MN, et al. 1998. Mapping of B epitopes in GRA4, a dense granule antigen of Toxoplasma gondii and protection studies using recombinant proteins administered by the oral route. Parasite Immunol. 20: 183–195 [PubMed] [Google Scholar]
- 42. Mezzasoma L, et al. 2002. Antigen microarrays for serodiagnosis of infectious diseases. Clin. Chem. 48: 121–130 [PubMed] [Google Scholar]
- 43. Montoya JG, Liesenfeld O. 2004. Toxoplasmosis. Lancet 363: 1965–1976 [DOI] [PubMed] [Google Scholar]
- 44. Nahtman T, et al. 2007. Validation of peptide epitope microarray experiments and extraction of quality data. J. Immunol. Methods 328: 1–13 [DOI] [PubMed] [Google Scholar]
- 45. Ngo Y, et al. 2009. Identification and testing of control peptides for antigen microarrays. J. Immunol. Methods 343: 68–78 [DOI] [PubMed] [Google Scholar]
- 46. Noya O, Patarroyo ME, Guzmán F, Alarcón de Noya B. 2003. Immunodiagnosis of parasitic diseases with synthetic peptides. Curr. Protein Pept. Sci. 4: 299–308 [DOI] [PubMed] [Google Scholar]
- 47. Pamelard F, et al. 2009. PASE: a web-based platform for peptide/protein microarray experiments. Methods Mol. Biol. 570: 413–430 [DOI] [PubMed] [Google Scholar]
- 48. Reineke U, Schutkowski M. 2009. Epitope mapping protocols. Preface. Methods Mol. Biol. 524: v–vi [PubMed] [Google Scholar]
- 49. Reineke U. 2009. Antibody epitope mapping using de novo generated synthetic peptide libraries. Methods Mol. Biol. 524: 203–211 [DOI] [PubMed] [Google Scholar]
- 50. Roggen EL. 2006. Recent developments with B-cell epitope identification for predictive studies. J. Immunotoxicol. 3: 137–149 [DOI] [PubMed] [Google Scholar]
- 51. Saeij JP, Arrizabalaga G, Boothroyd JC. 2008. A cluster of four surface antigen genes specifically expressed in bradyzoites, SAG2CDXY, plays an important role in Toxoplasma gondii persistence. Infect. Immun. 76: 2402–2410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Saha S, Raghava GP. 2006. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins 65: 40–48 [DOI] [PubMed] [Google Scholar]
- 53. Shen G, Behera D, Bhalla M, Nadas A, Laal S. 2009. Peptide-based antibody detection for tuberculosis diagnosis. Clin. Vaccine Immunol. 16: 49–54 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Sousa S, et al. 2009. Selection of polymorphic peptides from GRA6 and′ GRA7 sequences of Toxoplasma gondii strains to be used in serotyping. Clin. Vaccine Immunol. 16: 1158–1169 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Suzuki Y. 2002. Host resistance in the brain against Toxoplasma gondii. J. Infect. Dis. 185(Suppl. 1): S58–S65 [DOI] [PubMed] [Google Scholar]
- 56. Tenter AM, Heckeroth AR, Weiss LM. 2000. Toxoplasma gondii: from animals to humans. Int. J. Parasitol. 30: 1217–1258 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Travier L, et al. 2008. Functional domains of the Toxoplasma GRA2 protein in the formation of the membranous nanotubular network of the parasitophorous vacuole. Int. J. Parasitol. 38: 757–773 [DOI] [PubMed] [Google Scholar]
- 58. Vigil A, et al. 2010. Identification of the feline humoral immune response to Bartonella henselae infection by protein microarray. PLoS One 5: e11447 doi:10.1371/journal.pone.0011447 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.