

Complementary and alternative medicine (CAM) use is common among adults, and recent reports suggest that 25%–50% of prostate cancer (PCa) patients use at least one CAM modality. The most common CAM modalities used by PCa patients are vitamin and herbal preparations (e.g., common antioxidants, pomegranate extract, green tea, turmeric, resveratrol, silibinin, and herbal combination preparations) with purported antitumor effects despite only modest underlying preclinical or clinical evidence of efficacy. From the available data we conclude that there is insufficient data to support the use of CAMs for the treatment of prostate cancer patients outside of a clinical trial.
Keywords: Prostate cancer, Pomegranate, Curcumin, Green tea, Complementary and alternative medicine, Resveratrol, Silibinin
Abstract
Complementary and alternative medicine (CAM) use is common among adults, and recent reports suggest that 25%–50% of prostate cancer (PCa) patients use at least one CAM modality. The most common CAM modalities used by PCa patients are vitamin and herbal preparations with purported antitumor effects despite only modest underlying preclinical or clinical evidence of efficacy. In this review we provide a brief overview of the basic scientific and clinical studies underlying the most common herbal and vitamin preparations including common antioxidants, pomegranate extract, green tea, turmeric, resveratrol, silibinin, and herbal combination preparations. When available, prostate cancer clinical trial data are reviewed. Importantly, we have compared the concentration of these agents used in in vitro experiments to that likely to be achievable in humans. From the available data we conclude that there is insufficient evidence to support the use of CAMs for the treatment of prostate cancer patients outside of a clinical trial. The purpose of this review is to more rigorously evaluate CAM therapy in prostate cancer and educate oncologists and patients. This review focuses on examples from the general classes of agents in common use.
Introduction
Complementary and alternative medicine (CAM) use is common among adults with some reports demonstrating 25%–50% of prostate cancer (PCa) patients use at least one CAM modality [1–4]. Adults in the U.S. spent $33.9 billion on CAM products and visits to CAM practitioners in 2007; and prostate cancer patients are among the largest users of these preparations [5]. Despite their common use, there have been few controlled clinical trials involving CAM therapies, and treatment is often undertaken without proven clinical efficacy. Moreover, although in many cases CAMs may be derived from natural substances, there is still the possibility of untoward effects. For example, the recently published update of the SELECT trial showed an increased risk of PCa in men randomized to vitamin E, even after subjects stopped this vitamin [6]. This finding alone should call into question the role of CAMs that are untested with respect to either treatment or chemoprevention for prostate cancer and other malignancies.
What is problematic to the CAM field is that although there are often multiple cell-based and even animal models demonstrating efficacy, there is a paucity of clinical trial data. Making trials more difficult to accomplish are a reliable source of the CAM, lot-to-lot variability, a lack of reliable biomarkers, and a paucity of pharmacokinetic data to base decisions regarding schedule or dose. Lastly, there is little incentive for manufacturers to participate in or sponsor these trials, given that the market, especially the Internet market, is already so large for their products, and FDA approval is not necessary. Despite the lack of rigorous clinical studies, oncologists are frequently approached with questions and for advice regarding CAM therapies from prostate cancer patients.
In this review we describe potential scientific and molecular mechanisms for some of the more commonly used herbal and supplemental substances used in prostate cancer. We focus on agents that have been tested, even nominally, in the clinical setting. We have also tried to compare the drug concentrations needed to produce an effect in vitro with published concentrations achieved in humans. In some cases, the scientific basis for the CAM's effectiveness and the potential to achieve active in vivo levels of the CAM might make further evaluation reasonable. In other cases there may be less data to support further clinical experimentation. In either case an understanding of the possible mechanisms of action and achievable drug concentrations in patients may make this difficult field more tractable for patients and clinicians alike.
Materials and Methods
Data reviewed are from experiments involving human prostate cancer cell lines, prostate cancer animal models, and human clinical trials. We have focused primarily on preclinical and clinical data that relates to treatment, rather than the prevention, of prostate cancer. The findings in all referenced articles have been confirmed in at least one other publication available through PubMed. When possible, drug doses have been converted to micromolar (μM) units or microgram per milliliter (μg/mL) concentrations to better compare CAM drug levels between cell lines, xenograft, and human data. There are no unpublished data or personal communications included in the preparation of this review.
Selenium, Vitamin E, and Other Antioxidants
The concept that antioxidants can prevent malignant transformation or treat established malignancy has been reported for many years and previously reviewed [7–9]. Briefly, this concept is based on the idea that free radicals (molecules with incomplete electron shells) are able to induce DNA damage that may ultimately lead to mutations that predispose cells to malignant transformation [7, 8]. In humans, the most common free radical is the hydroxyl radical (·OH) and the term reactive oxygen species is commonly used to encompass all oxygen-containing free radicals.
There are multiple preclinical models in prostate and other malignancies to suggest that antioxidants (also known as free-radical scavengers) can prevent malignant transformation and/or delay progression [8–12]. Antioxidants such as selenium and vitamin E have long been touted, and until quite recently, commonly used by PCa patients with both preventive and therapeutic intent.
In 2009 the SELECT prevention trial reported the results of a randomized trial of vitamin E, selenium, or both for the prevention of prostate cancer in men over 50 [13]. Although the SELECT trial was a prevention trial, the nature of prostate cancer is such that it is likely that many of the participants had prostate cancer at the time of randomization, especially since prostate biopsies were not required for entry. Therefore, an effect on existing cancers during the course of the 7-year study might have been reflected by a reduced frequency of cancer in patients receiving active treatment (selenium, vitamin E, or both) compared with placebo [13]. However, with longer follow-up, a significant and lasting increase in prostate cancer emerged in patients treated with vitamin E (hazard ratio 1.17) and no effect on prostate cancer incidence in patients treated with selenium [6, 13]. The finding that even two fairly robust antioxidant agents in combination were not effective, and potentially harmful, further highlights the potential harm of treatment with untested CAM supplements and the need for their rigorous study before their widespread use [6, 13].
Although the SELECT trial and Physicians' Health Study (PHS II) have shed doubt on the role of selenium and vitamin E in the prevention of prostate cancer, there continue to be ongoing National Cancer Institute-sponsored (NCI-sponsored) treatment trials (see NCT00736645 and NCT01155791 for references) based on preclinical evidence. The preclinical data for selenium and vitamin E has been previously well reviewed and will not be further discussed in this review [14–17]. Further antioxidant studies are complicated by clinically applicable methods and ranges of antioxidant action on which to base assessment of efficacy and harm. Although the degree of antioxidant capacity can be readily quantified under controlled experimental conditions, there are no reliable clinical correlates. On the basis of phase III data from the SELECT trial, it is likely that continued research into antioxidants, including lycopene, as PCa treatment will wane.
Pomegranate Extract
Pomegranate (Punica granatum) has been used in medicine for over 3,000 years, with applications ranging from hypertension to malignancy [18]. Prior studies have shown the ability of pomegranate to inhibit several human cancer cell lines [19]. The most active and abundant anti-neoplastic ingredients are the polyphenol punicalagins and specifically the ellagitannin polyphenol punicalagin, commonly measured as ellagic acid (EA) in bioavailability studies [20]. Punicalagins are isomers of 2,3-(S)-hexahydroxydiphenoyl-4,6-(S,S)-gallagyl-d-glucose and exist as both alpha and beta forms in pomegranate. Both forms are abundant and typically constitute 2 g/L of juice [21].
Pomegranate polyphenols have robust antioxidant properties with a reactive oxygen species capacity higher than either red wine or green tea [22]. However, as discussed, it is not at all certain that any degree of antioxidant capacity accounts for an anticancer effect in prostate cancer. Beyond their pure antioxidant capabilities, the punicalagins have been shown to have multiple effects on signal transduction. The PCa cell line PC3 cells, exposed to pomegranate extract (PE) at high concentrations ranging from 10 to 100 μg/mL, showed dose-dependent apoptosis through induction of p21/WAF1 and p27/KIP1, a decrease in B-cell lymphoma 2 (Bcl-2), and an increase in Bcl-2–associated X protein (Bax) (favoring apoptosis) [23]. These results were confirmed in vivo with CWR22Rv1 prostate cancer xenografts in nude mice fed pomegranate extract. However, the concentrations needed to achieve these effects in vitro and in nude mice are likely to be far greater than that which can be achieved in patients.
It has previously been shown that inflammation and prostate cancer may be linked, perhaps through constitutive activation of the nuclear factor-κB (NF-κB) pathway [24, 25]. Increased levels of NF-κB in primary prostate cancer specimens have been shown to be an independent risk factor for recurrence after definitive therapy [26, 27]. PE inhibited NF-κB expression in the androgen-independent DU145 prostate cancer cell line, in a dose-dependent fashion, but at doses estimated to be more than 10 times greater than what could be achieved in patients [28]. PE also delayed the appearance of androgen-independent tumors in LAPC4 xenografts, an effect at least partially dependent on the NF-κB inhibitory effects of PE [28].
Although it was not certain that adequate levels of PE could be achieved in vivo, the preclinical data showing growth inhibition and apoptosis induction led to initiation of human trials with pomegranate juice. Pantuck et al. demonstrate the remarkable initial clinical observation that 8 ounces of pomegranate juice per day prolonged the prostate-specific antigen (PSA) doubling time from a baseline of 15 months to 54 months post-PE treatment in hormone-naive non-metastatic patients with rising PSA after surgery or radiotherapy [29]. Importantly, this is one of the few studies to show a clinical benefit from a CAM in prostate cancer. This study potentially addresses a large unmet need for those prostate cancer patients who have biochemical (PSA only) recurrence after primary therapy. Patients and providers are motivated to delay or prevent the side effects of androgen-deprivation therapy for PSA-only recurrence. There are currently 11 prospective cancer trials registered with www.clinicaltrials.gov evaluating the impact of pomegranate supplementation (see NCT01100866, NCT00719030, NCT00336934, NCT00413530, and NCT01220817 for reference).
In trying to understand the mechanism of PE, it is helpful to first assess what levels can be reliably achieved in patients. Investigators found that the 8 ounces per day of pomegranate juice (POM Wonderful, Los Angeles) used in the Pantuck study standardized to 570 mg of total polyphenol gallic acid equivalents per day achieved serum ellagic acid (EA) concentrations of 0.14 ± 0.05 μM/L [21, 29] (Table 1). However, in a separate study EA concentrations achieved were only 0.06 μM/L 1 hour after consuming 180 mL (approximately 6 ounces) of pomegranate juice) [22]. The effect of possible polymorphisms in metabolic enzymes such as catechol-o-methytransferase and uridine 5′-diphospho-glucuronosyltransferases, as well as whether or not EA is the optimal metabolite to measure, remains to be determined [21]. Post-treatment tissue samples from prostatectomy specimens with measurement of EA concentrations at the tissue level from planned neoadjuvant trials, as is planned by Carducci and colleagues (NCT00719030), will also be informative.
Table 1.
Comparison of CAM concentrations used in cell line/xenograft and human studies
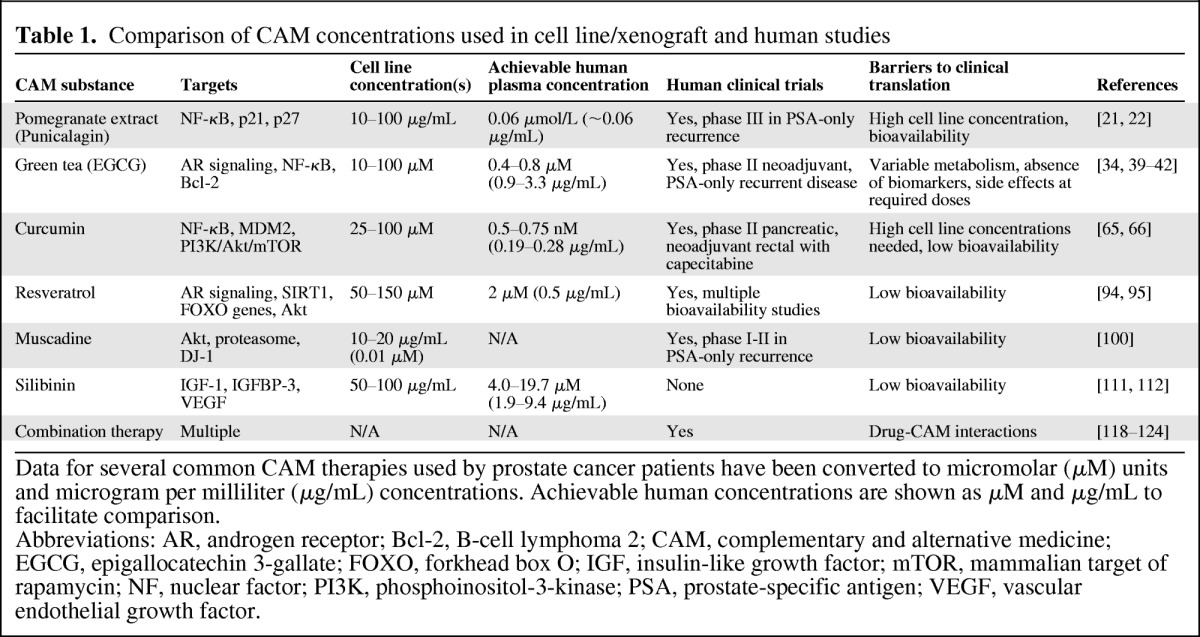
Data for several common CAM therapies used by prostate cancer patients have been converted to micromolar (μM) units and microgram per milliliter (μg/mL) concentrations. Achievable human concentrations are shown as μM and μg/mL to facilitate comparison.
Abbreviations: AR, androgen receptor; Bcl-2, B-cell lymphoma 2; CAM, complementary and alternative medicine; EGCG, epigallocatechin 3-gallate; FOXO, forkhead box O; IGF, insulin-like growth factor; mTOR, mammalian target of rapamycin; NF, nuclear factor; PI3K, phosphoinositol-3-kinase; PSA, prostate-specific antigen; VEGF, vascular endothelial growth factor.
Green Tea
Tea (Camellia sinensis) is one of the most widely consumed beverages in the world and epidemiologic studies have suggested varying effects on cancer incidence [30–34]. By far most of the attention for anticancer treatment and prevention has focused on green tea, and other tea preparations are not reviewed here.
Polyphenols account for up to 30% of green tea dry weight [35, 36]. The monomer flavan-3-ols are called catechins and account for the bitter taste of green tea as well as most of the biologic properties [37]. In green tea the most abundant catechin is epigallocatechin 3-gallate (EGCG), which may account for up to 50%–80% of the total catechin content [35, 36, 38]. One cup (7 ounces) of green tea contains roughly 142 mg of EGCG, and the equivalent of 8–16 cups per day is needed to achieve plasma free ECGC concentrations of 0.4–0.8 μM [34, 36, 39–42] (Table 1). However, variations in steeping will result in highly variable levels of serum and tissue polyphenols. Therefore, there are now standardized preparations such as polyphenon-E (poly-E) capsules (standardized for 400, 800, or 1200 mg of EGCG), which have been shown to achieve plasma free EGCG concentration of 1.5 μg/mL after single dosing (800 mg of EGCG) in human patients on an empty stomach [43]. These preparations may result in peak concentrations of up to 5 μM at best, although these levels, often reached with the 1200 mg poly-E capsules are commonly associated with gastrointestinal upset [43]. Plasma free EGCG concentrations have been highly variable in phase I trials possibly related to the green tea extract formulation, the presence or absence of recent food intake, metabolic polymorphisms, and/or the methodology used to measure plasma free EGCG concentrations [44].
In prostate cancer there has been intense investigation of the effects of EGCG on androgen receptor (AR) signaling in both cell line and xenograft experiments [45–50]. Siddiqui et al. demonstrated that EGCG is a direct antagonist of androgen action and physically interacts with the ligand binding domain of the androgen receptor [50]. EGCG at 40 and 60 μM doses both inhibited growth of hormone refractory C4-2 prostate cancer cells and decreased AR protein expression in EGCG-treated mice bearing a CWR22Rv1 xenograft [50]. In this context it is well to remember that serum levels achieved are likely to be an order of magnitude less than 40 μM.
Nonhormonal effects have also been shown for EGCG [51, 52]. EGCG demonstrated a dose-responsive effect for increasing p53 expression and stabilization via serine phosphorylation in LNCaP cells [51]. EGCG has been shown to downregulate transcriptional activity of NF-κB, which creates a proapoptotic environment, and to increase the sensitivity of LNCaP cells to tumor necrosis factor [48, 51]. Overall, the data supports EGCG-mediated anticancer effects via multiple mechanisms with multiple targets, but the concentrations used in cell line and xenograft experiments range from 10–200 μM [51–56]. These levels are significantly higher than the 5 μM concentration that can be achieved in patients. There is also preclinical data for EGCG (at somewhat lower concentrations) used to sensitize cells for treatment in combination with taxanes in prostate and breast cancer cell lines, as well as combinations with doxorubicin (ovarian sarcoma) [57, 58].
In a small study of patients with the potential prostate cancer precursor lesion, high-grade prostatic intraepithelial neoplasia (HG-PIN) patients were randomized into a control and an EGCG-treated group. Patients had routine prostate biopsies every 3–6 months for 1 year. Patients treated with EGCG had a dramatic reduction in the detection of cancer on subsequent prostate biopsies (30% in the control arm to 3% in the EGCG-treated group) [59]. Potential anticancer effects of EGCG were also detected in a small phase II trial of 42 patients with castrate refractory PCa that demonstrated a low, but discernable, PSA response [60].
As with most anticancer herbal/vitamin preparations, there is difficulty translating cell line and xenograft data into disease-specific activity in patient studies. One method to overcome this problem is to ascertain levels and target effects of EGCG in prostate tissue. The current NCI-sponsored phase II trial of neoadjuvant green tea treatment in men treated prior to planned prostatectomy (NCT00685516) may be very informative if tissue levels can be adequately measured and equated with molecular or antitumor effects. There is also an ongoing study of EGCG for men following biochemical recurrence (NCT00669656).
Curcumin
Curcumin (isolated from Curcuma longa) is a major component of the popular spice turmeric, and curcuminoids are responsible for the yellow color of curry. The major component of curcumin is the lipophylic polyphenol curcumin I (diferuloylmethane) [61]. Curcumin has been used for treatment of many inflammatory conditions, and more recently for malignancies including prostate cancer [62]. The interest in the antineoplastic properties of curcumin grew out of epidemiologic studies correlating gastrointestinal and prostate cancer incidence and dietary turmeric intake [63, 64].
The safety of high doses of curcumin up to 8 grams per day is well described in humans [65]. Newer nanoparticle formulations of curcumin are able to achieve plasma curcumin concentrations in the range of 0.5–0.75 nM (189 and 275 ng/mL) for oral preparations of 150 and 210 mg, respectively [66] (Table 1). However, as has been the case with other CAMs, in vitro experiments with curcumin have typically been conducted at micromolar levels. For example, the levels used in DU145 prostate cell line to suppress NF-κB and AP-1 expression were 75 and 50 μM, respectively [67]. Curcumin has been shown to downregulate cyclin D1 in a dose-dependent manner in LNCaP cells as a result of activation of proteases involved in the ubiquitin-dependent proteasomal pathway [68]. Others have shown that curcumin at concentrations of 15 μmol/L inhibits MDM2 expression independent of p53 status, possibly related to modulation of the erythroblastosis virus transcription factor 2 (ETS2), a transcription factor that can be activated as a result of the TMPRSS2-erg translocation in some prostate cancers [69]. Curcumin-mediated inhibition of the PI3K/Akt/mTOR pathway was demonstrated in PC-3 cells as a result of dephosphorylation of Akt and mTOR, but again at a high concentration (40 μM) [70, 71]. In C4–2 and LNCaP lines curcumin treatment has also been shown to downregulate the AR regulated TMPRSS2-ERG fusion transcript associated with prostate cancer progression at curcumin concentrations starting at 5 μM [72, 73].
Curcumin has been tested in combination with cytotoxic chemotherapies in prostate cancer cell lines. Synergism between 1–10 μM curcumin and 5-fluorouracil (5-FU) and paclitaxel in PC-3 cells has been observed [74]. The ability of curcumin to potentiate the cytotoxicity of chemotherapy agents has also been documented in other cell lines [75–79]. Curcumin-mediated MDM2 downregulation sensitized the PC3 prostate cancer cell line to both gemcitabine and radiation in cell line and mouse xenograft models [80].
The observed synergism of curcumin with cytotoxic therapies has prompted several combination trials currently ongoing. Curcumin is being investigated in combination with neoadjuvant capecitabine and radiation in rectal cancer (NCT00745134) and combined with FOLFOX in inoperable colon cancer (NCT01490996). There is also an ongoing study of curcumin monotherapy in advanced pancreatic cancer (NCT00094445).
Although it is apparent that curcumin can exert anticancer properties via multiple targets, the concentration needed to achieve these results may not be achievable in vivo. Therefore, there are several ongoing efforts evaluating newer nanoparticle curcumin formulations as well as combinations with piperine to improve bioavailability.
Resveratrol and Other Grape Skin Extracts
Resveratrol (RSV) is a phytoalexin found in high concentrations in grapes and has been shown to inhibit chemical carcinogenesis in several models [81]. Interest in RSV as a chemotherapeutic agent increased after the publication of a seminal 1997 paper by Jang et al. [81]. A recent review described the preclinical, animal model, and human studies across several malignancies [82].
Specific interest for the treatment of prostate cancer was enhanced by a study in which resveratrol was shown to inhibit AR target genes due, at least in part, to androgen receptor transcriptional repression [83]. Other groups have shown resveratrol treatment leads to accelerated AR degradation, as well as modulating AR co-activator molecules such as SIRT1 at RSV concentrations of 50 μM [84–86]. In addition to resveratrol's influence on AR function, there is evidence for its nongenomic effects through the PI3K pathway in steroid hormone responsive tumor cell lines (estrogen receptor + breast and AR+ prostate) [87–89]. High concentrations of 50–150 μM have been shown to result in significant inhibition of PKB/Akt phosphorylation in a concentration-dependent manner in both PC-3 and LNCaP cell lines [90]. The cellular effects of resveratrol-mediated PI3K inhibition may be partly mediated by inhibition of FOXO family tumor suppressor phosphorylation, thereby allowing upregulation of proapoptotic and antiproliferative FOXO targets such as Bim, TRAIL, DR5, DR4, and p27 [91]. Very recently, Park et al. elegantly demonstrated that resveratrol, at concentrations of 50 μM, directly inhibits cyclic-AMP dependent phosphodiesterases and ultimately leads to activation of SIRT1 and AMP kinase through the cyclic-AMP effector protein Epac1 [92].
Pharmacokinetic studies of resveratrol at doses up to 5 grams per day have demonstrated poor bioavailability due to extensive glucuronidation and sulfation as well as metabolism by gut bacterial enzymes [82, 93]. Estimates using liquid chromatography, mass spectrometry, and radiolabeled doses estimate oral bioavailability of 1% and peak plasma concentrations of 500 ng/mL (approximately 2 μM), which is well below the doses used in most cell line experiments [94, 95] (Table 1). However, low micromolar levels of 2.5–10 μM have been shown to sensitize prostate cancer cells to ionizing radiation without affecting normal prostate epithelial cells [96]. There is also data for the effect of RSV on inhibition of angiogenesis in lung, breast, and glioma xenograft models [97–99]. Interest in resveratrol as a therapeutic adjuvant to current therapy persists due to accumulating cell line data and ongoing efforts to improve oral bioavailability.
Other grape skin extracts such as muscadine grape (Vitis rotundifolia) skin extract, which does not contain resveratrol, have also been investigated in prostate cancer. Hudson and colleagues demonstrated 10–20 μg/mL muscadine grape skin extract (approximately 10–100 nM) induced apoptosis in multiple prostate cell lines via decreased Akt signaling [100]. The Department of Defense Prostate Cancer Consortium has recently begun a phase I/II study of muscadine for men with biochemical recurrence of prostate cancer (NCT01317199).
Silibinin
Silibinin is thought to be the major active ingredient in Milk Thistle extract (Silybum marianum), a supplement often used by prostate cancer patients and previously shown to be nontoxic in humans [101, 102]. Structurally, silibinin is a polyphenolic flavanoid and has historically been used in various liver tonic preparations on account of its antihepatotoxic effects [103, 104]. Silibinin may have a mechanism of action different from the previously reviewed CAMs in that it can modulate the insulin-like growth factor 1 (IGF-1) signaling pathway, known to be upregulated in prostate cancer [105, 106]. IGF ligand levels have been shown to be associated with prostate cancer risk, and IGF signaling can have downstream effects on the prosurvival and antiapoptotic Ras/Raf/MAPK and PI3K/Akt pathways [107, 108]. Mouse models have shown that oral silibinin increases insulin-like growth factor binding protein 3 (IGFBP-3) plasma concentrations, thereby possibly inhibiting IGF binding to IGF receptors [101, 105, 106, 108]. Silibin has demonstrated synergy with doxorubicin and mitoxantrone in prostate cell lines at concentrations readily achievable in humans [109, 110].
Oral silibinin administration achieves measurable concentrations of between 0.3 and 4.0 μmol/L depending on silibinin dose (range 360-1440 mg daily), which is roughly comparable to concentrations tested in mouse xenograft doses [111] (Table 1). In a small high-dose oral (13 grams per day) neoadjuvant trial for men with localized prostate cancer planned for radical prostatectomy, mean plasma silibinin levels of 19.7 μM were achieved, but silibinin was not detected in significant accumulation in prostate tissue [112]. Furthermore, there was no significant change in serum IGF or IGFBP-3 levels, although the treatment was of short duration (mean of 20 days) [112].
Silibinin has demonstrated interesting preventative and anticancer properties in prostate cancer animal models. It appears to be more bioavailable than some of the other CAMs, but there may be major differences in serum compared with tissue levels, making its future in clinical treatment uncertain.
Combination CAM Therapy
Although there is a strong preclinical rationale for the use of several CAMs, the concentrations needed to achieve an in vivo effect are not certain. One method to overcome this problem is to combine several CAMs with overlapping targets within the same preparation.
PC-SPES, a previously patented blend of 8 herbs including baikal skullcap (Scutellaria baicalensis Georgi), chrysanthemum (Dendranthema morifolium), ganoderma (Ganoderma lucidum), isatis (Isatis indigotica fortune), licorice (Glycyrrhiza glabra licorice), Panax ginseng (Panax pseudoginseng var.), Hara (Rabdosia rubescens), and saw palmetto (Serenoa repens), was one of the early combination therapies in prostate cancer [113]. In preclinical models PC-SPES demonstrated activity in androgen-dependent and androgen-independent in vitro and in vivo models [114–117]. Our group with others embarked on a randomized phase II trial of PC-SPES compared with diethylstilbestrol (DES) in patients with castrate refractory PCa. Over 40% of patients randomized to PC-SPES demonstrated a 50% decline in PSA and a median time to progression of 5.5 months compared with 2.9 months for DES [118]. However, concentrations up to 3.1% of DES were detected in PC-SPES formulations, which potentially confounded the trial and led to its early termination [118]. Subsequent analyses also detected low concentrations of warfarin and indomethacin contaminants in PC-SPES, and it was taken off even the Internet market in 2002.
A more recent herbal combination therapy common to many prostate cancer patients is Zyflamend (New Chapter, Brattleboro, Vermont), a proprietary blend of 10 standardized herbal extracts (turmeric, holy basil, green tea, hu zhang, ginger, Chinese goldthread, oregano, Scutellaria baicalensis, and barberry) [119]. In LNCAP cells Zyflamend has been shown to inhibit COX-1 and COX-2, induce cell cycle inhibitory proteins including p21, and suppress AR expression [120, 121]. Consistent with its anti-inflammatory actions, Zyflamend downregulates NF-κB gene products and NF-κB activation in lung cancer and leukemia cell lines at concentrations of 0.8 mg/mL [122]. Recently, Zyflamend demonstrated synergism with Bicalutamide in LNCaP cells and with gemcitabine in a mouse pancreatic cancer model [123, 124]. Combination preparations are currently in clinical trials in prostate cancer (NCT00669656). As with other CAMs discussed, the dosing and measurement of plasma concentrations in preformulated herbal combinations is complicated, and the potential interaction between constituents is often not well established.
Summary
CAM use is common among prostate cancer patients and is expected to continue to increase over time. The preclinical anticancer mechanistic data for many naturally occurring supplements can be impressive, but there is insufficient attention paid to meticulous establishment of meaningful biologic concentrations and whether these can be achieved in patients. Prior to mounting phase II-III clinical trials with these agents, rigorous phase I and pharmacokinetic data from standardized preparations will be needed to determine an appropriate dose and schedule for testing. Beyond this concern there are the issues of a more clear understanding of target-interactions, response biomarkers, and compound purity. Finally, there are likely to be issues about investigational new drug concerns for CAMs that are not controlled by established pharmaceutical companies. Most studies will of necessity be investigator-initiated, but without a funding source or even a reliable source of drug, these trials will continue to be problematic.
Here we have provided an overview of the preclinical data and proposed mechanisms for selected compounds in prostate cancer. It behooves the translational and practicing community oncologist to have a familiarity with CAMs because of their widespread use. On the basis of preclinical studies, there is a rationale for CAM use in prostate cancer, but it is difficult if not impossible for clinicians to sanction their use because of all the concerns above. For example, some preparations of Prostasol, a combination CAM widely used, may be associated with an increase in venous thrombosis, possibly because of estrogenic properties [125]. Nonetheless, for a well-studied, well-tolerated, and bioavailable CAM, studies in combination with established therapies such as docetaxel or as single agents aimed at slowing PSA progression in patients with biochemical recurrence are appropriate and much needed.
Author Contributions
Conception/Design: Samuel J. Klempner, Glenn Bubley
Collection and/or assembly of data: Samuel J. Klempner, Glenn Bubley
Data analysis and interpretation: Samuel J. Klempner, Glenn Bubley
Manuscript writing: Samuel J. Klempner, Glenn Bubley
Final approval of manuscript: Samuel J. Klempner, Glenn Bubley
References
- 1.Barnes PM, Bloom B, Nahin RL. Complementary and alternative medicine use among adults and children: United States, 2007. Natl Health Stat Report. 2008;12:1–23. [PubMed] [Google Scholar]
- 2.Nam RK, Fleshner N, Rakovitch E, et al. Prevalence and patterns of the use of complementary therapies among prostate cancer patients: an epidemiologic analysis. J Urol. 1999;161:1521–1524. [PubMed] [Google Scholar]
- 3.McDermott CL, Blough DK, Fedorenko CR, et al. Complementary and alternative medicine use among newly diagnosed prostate cancer patients. Support Care Cancer. 2012;20:65–73. doi: 10.1007/s00520-010-1055-y. [DOI] [PubMed] [Google Scholar]
- 4.Jacobson JS, Chetty AP. Complementary and alternative medicine in prostate cancer. Curr Oncol Rep. 2001;3:448–452. doi: 10.1007/s11912-001-0032-5. [DOI] [PubMed] [Google Scholar]
- 5.Nahin RL, Barnes PM, Stussman BJ, et al. Costs of complementary and alternative medicine (CAM) and frequency of visits to CAM practitioners: United States, 2007. Natl Health Stat Report. 2009;18:1–14. [PubMed] [Google Scholar]
- 6.Klein EA, Thompson IM, Jr., Tangen CM, et al. Vitamin E and the risk of prostate cancer: the Selenium and Vitamin E Cancer Prevention Trial (SELECT) JAMA. 2011;306:1549–1556. doi: 10.1001/jama.2011.1437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Evans MD, Dizdaroglu M, Cooke MS. Oxidative DNA damage and disease: induction, repair and significance. Mutat Res. 2004;567:1–61. doi: 10.1016/j.mrrev.2003.11.001. [DOI] [PubMed] [Google Scholar]
- 8.Cooke MS, Evans MD, Dizdaroglu M, et al. Oxidative DNA damage: mechanisms, mutation, and disease. FASEB J. 2003;17:1195–1214. doi: 10.1096/fj.02-0752rev. [DOI] [PubMed] [Google Scholar]
- 9.Tan AC, Konczak I, Sze DM, et al. Molecular pathways for cancer chemoprevention by dietary phytochemicals. Nutr Cancer. 2011;63:495–505. doi: 10.1080/01635581.2011.538953. [DOI] [PubMed] [Google Scholar]
- 10.Corcoran NM, Najdovska M, Costello AJ. Inorganic selenium retards progression of experimental hormone refractory prostate cancer. J Urol. 2004;171:907–910. doi: 10.1097/01.ju.0000092859.16817.8e. [DOI] [PubMed] [Google Scholar]
- 11.Zu K, Ip C. Synergy between selenium and vitamin E in apoptosis induction is associated with activation of distinctive initiator caspases in human prostate cancer cells. Cancer Res. 2003;63:6988–6995. [PubMed] [Google Scholar]
- 12.Neuzil J, Weber T, Gellert N, et al. Selective cancer cell killing by a-tocopheryl succinate. Br J Cancer. 2001;84:87–89. doi: 10.1054/bjoc.2000.1559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lippman SM, Klein EA, Goodman PJ, et al. Effect of selenium and vitamin E on risk of prostate cancer and other cancers. The Selenium and Vitamin E Cancer Prevention Trial (SELECT) JAMA. 2009;301:39–51. doi: 10.1001/jama.2008.864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ganther HE. Selenium metabolism, selenoprotein and mechanisms of cancer prevention: complexities and thioredoxin reductase. Carcinogenesis. 1999;20:1657–1666. doi: 10.1093/carcin/20.9.1657. [DOI] [PubMed] [Google Scholar]
- 15.Combs GF., Jr Status of selenium in prostate cancer prevention. Br J Cancer. 2004;91:195–199. doi: 10.1038/sj.bjc.6601974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Venkateswaran V, Fleshner NE, Klotz LH. Modulation of cell proliferation and cell cycle regulators by vitamin E in human prostate carcinoma cell lines. J Urol. 2002;168:1578–1582. doi: 10.1016/S0022-5347(05)64524-7. [DOI] [PubMed] [Google Scholar]
- 17.Qi Y, Schoene NW, Lartey FM, et al. Selenium compunds activate ATM-dependent DNA damage response via the mismatch repair protein hMLH1 in colorectal cancer cells. J Biol Chem. 2010;285:33010–33017. doi: 10.1074/jbc.M110.137406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Langley P. Why a pomegranate? BMJ. 2000;321:1153–1154. doi: 10.1136/bmj.321.7269.1153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Longtin R. The pomegranate: nature's power fruit? J Natl Cancer Inst. 2003;95:346–348. doi: 10.1093/jnci/95.5.346. [DOI] [PubMed] [Google Scholar]
- 20.Gil MI, Tomas-Barberan FA, Hess-Pierce B, et al. Antioxidant activity of pomegranate juice and its relationship with phenolic composition and processing. J Agric Food Chem. 2000;48:4581–4589. doi: 10.1021/jf000404a. [DOI] [PubMed] [Google Scholar]
- 21.Seeram NP, Zhang Y, McKeever R, et al. Pomegranate juice and extracts provide similar levels of plasma and urinary ellagitannin metabolites in human subjects. J Med Food. 2008;11:390–394. doi: 10.1089/jmf.2007.650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Seeram NP, Henning SM, Zhang Y, et al. Pomegranate juice ellagitannin metabolites are present in human plasma and some persist in urine for up to 48 hours. J Nutr. 2006;136:2481–2485. doi: 10.1093/jn/136.10.2481. [DOI] [PubMed] [Google Scholar]
- 23.Malik A, Afaq F, Sarfaraz S, et al. Pomegranate fruit juice for chemoprevention and chemotherapy of prostate cancer. Proc Natl Acad Sci U S A. 2005;102:14813–14818. doi: 10.1073/pnas.0505870102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Irani J, Goujon JM, Ragni E, et al. High-grade inflammation in prostate cancer as a prognostic factor for biochemical recurrence after radical prostatectomy. Pathologist Multi Center Study Group. Urology. 1999;54:467–472. doi: 10.1016/s0090-4295(99)00152-1. [DOI] [PubMed] [Google Scholar]
- 25.De Marzo AM, Marchi VL, Epstein JI, et al. Proliferative inflammatory atrophy of the prostate: implications for prostatic carcinogenesis. Am J Pathol. 1999;155:1985–1992. doi: 10.1016/S0002-9440(10)65517-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Domingo-Domenech J, Mellado B, Ferrer B, et al. Activation of nuclear factor-kB in human prostate carcinogenesis and association to biochemical relapse. Br J Cancer. 2005;93:1285–1294. doi: 10.1038/sj.bjc.6602851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Fradet V, Lessard L, Begin LR, et al. Nuclear factor-kB nuclear localization is predictive of biochemical recurrence in patients with positive margin prostate cancer. Clin Cancer Res. 2004;10:8460–8464. doi: 10.1158/1078-0432.CCR-04-0764. [DOI] [PubMed] [Google Scholar]
- 28.Rettig MB, Heber D, An J, et al. Pomegranate extract inhibits androgen-independent prostate cancer growth through a nuclear factor-KB-dependent mechanism. Mol Cancer Ther. 2008;7:2662–2671. doi: 10.1158/1535-7163.MCT-08-0136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pantuck AJ, Leppert JT, Zomorodian N, et al. Phase II study of pomegranate juice for men with rising prostate-specific antigen following surgery or radiation for prostate cancer. Clin Cancer Res. 2006;12:4018–4026. doi: 10.1158/1078-0432.CCR-05-2290. [DOI] [PubMed] [Google Scholar]
- 30.Kuriyama S, Shimazu T, Ohmori K, et al. Green tea consumption and mortality due to cardiovascular disease, cancer, and all causes in Japan: the Ohsaki study. JAMA. 2006;296:1255–1265. doi: 10.1001/jama.296.10.1255. [DOI] [PubMed] [Google Scholar]
- 31.Katiyar SK, Mukhtar H. Tea in chemoprevention of cancer: epidemiologic and experimental studies. Int J Oncol. 1996;8:221–238. doi: 10.3892/ijo.8.2.221. [DOI] [PubMed] [Google Scholar]
- 32.Rosenberg L. Coffee and tea consumption in relation to the risk of large bowel cancer: a review of epidemiologic studies. Cancer Lett. 1990;52:163–171. doi: 10.1016/0304-3835(90)90183-x. [DOI] [PubMed] [Google Scholar]
- 33.Kinlen LJ, McPherson K. Pancreas cancer and coffee and tea consumption: a case-control study. Br J Cancer. 1984;49:93–96. doi: 10.1038/bjc.1984.14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Yang G, Zheng W, Xiang YB, et al. Green tea consumption and colorectal cancer risk: a report from the Shanghai Men's Health Study. Carcinogenesis. 2011;32:1684–1688. doi: 10.1093/carcin/bgr186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Yang CS, Wang ZY. Tea and cancer. J Natl Cancer Inst. 1993;85:1038–1049. doi: 10.1093/jnci/85.13.1038. [DOI] [PubMed] [Google Scholar]
- 36.Graham HN. Green tea composition, consumption, and polyphenol chemistry. Prev Med. 1992;21:334–350. doi: 10.1016/0091-7435(92)90041-f. [DOI] [PubMed] [Google Scholar]
- 37.Adhami VM, Malik A, Zaman N, et al. Combined inhibitory effects of green tea polyphenols and selective cyclooxygenase-2 inhibitors on the growth of human prostate cancer cells both in vitro and in vivo. Clin Cancer Res. 2007;13:1611–1619. doi: 10.1158/1078-0432.CCR-06-2269. [DOI] [PubMed] [Google Scholar]
- 38.Park AM, Dong Z. Signal transduction pathways: targets for green and black tea polyphenols. J Biochem Mol Biol. 2003;36:66–77. [PubMed] [Google Scholar]
- 39.Chow HH, Cai Y, Alberts DS, et al. Phase I pharmacokinetic study of tea polyphenols following single-dose administration of epigallocatechin gallate and polyphenon E. Cancer Epidemiol Biomark Prev. 2001;10:53–58. [PubMed] [Google Scholar]
- 40.Chow HH, Cai Y, Hakim IA, et al. Pharmacokinetics and safety of green tea polyphenols after multipledose administration of epigallocatechin gallate and polyphenon E in healthy individuals. Clin Cancer Res. 2003;9:3312–3319. [PubMed] [Google Scholar]
- 41.Yang CS, Maliakal P, Meng X. Inhibition of carcinogenesis by tea. Annu Rev Pharmacol Toxicol. 2002;42:25–54. doi: 10.1146/annurev.pharmtox.42.082101.154309. [DOI] [PubMed] [Google Scholar]
- 42.Jankun J, Selman SH, Swiercz R, et al. Why drinking green tea could prevent cancer. Nature. 1997;387:561. doi: 10.1038/42381. [DOI] [PubMed] [Google Scholar]
- 43.Chow HH, Hakim IA, Vining DR, et al. Effects of dosing condition on the oral bioavailability of green tea catechins after single-dose administration of polyphenon E in healthy individuals. Clin Cancer Res. 2005;11:4627–4633. doi: 10.1158/1078-0432.CCR-04-2549. [DOI] [PubMed] [Google Scholar]
- 44.Pisters KM, Newman RA, Coldman B, et al. Phase I trial of oral green tea extract in adult patients with solid tumors. J Clin Oncol. 2001;19:1830–1838. doi: 10.1200/JCO.2001.19.6.1830. [DOI] [PubMed] [Google Scholar]
- 45.Cao Y, Cao R. Angiogenesis inhibited by drinking tea. Nature. 1999;398:381. doi: 10.1038/18793. [DOI] [PubMed] [Google Scholar]
- 46.Lin JK, Liang YC, Lin-Shiau SY. Cancer chemoprevention by tea polyphenols through mitotic signal transduction blockade. Biochem Pharmacol. 1999;58:911–915. doi: 10.1016/s0006-2952(99)00112-4. [DOI] [PubMed] [Google Scholar]
- 47.Yang GY, Liao J, Kim K, et al. Inhibition of growth and induction of apoptosis in human cancer cell lines by tea polyphenols. Carcinogenesis. 1998;19:611–616. doi: 10.1093/carcin/19.4.611. [DOI] [PubMed] [Google Scholar]
- 48.Siddiqui IA, Malik A, Adhami VM, et al. Green tea polyphenol EGCG sensitizes human prostate carcinoma LNCaP cells to TRAIL-mediated apoptosis and synergistically inhibits biomarkers associated with angiogenesis and metastasis. Oncogene. 2008;27:2055–2063. doi: 10.1038/sj.onc.1210840. [DOI] [PubMed] [Google Scholar]
- 49.Liao X, Tang S, Thrasher JB, et al. Small-interfering RNA-induced androgen receptor silencing leads to apoptotic cell death in prostate cancer. Mol Cancer Ther. 2005;4:505–515. doi: 10.1158/1535-7163.MCT-04-0313. [DOI] [PubMed] [Google Scholar]
- 50.Siddiqui IA, Asim M, Hafeez BB, et al. Green tea polyphenol EGCG blunts androngen receptor function in prostate cancer. FASEB J. 2011;25:1198–1207. doi: 10.1096/fj.10-167924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Hastak K, Gupta S, Ahmad N, et al. Role of p53 and NF-kB in epigallocatechin-3-gallate-induced apoptosis of LNCaP cells. Oncogene. 2003;22:4851–4859. doi: 10.1038/sj.onc.1206708. [DOI] [PubMed] [Google Scholar]
- 52.Gupta S, Ahmad N, Nieminen, et al. Growth inhibition, cell-cycle dysregulation, and induction of apoptosis by green tea constituent (−)-epigallocatechin-3-gallate in androgen-sensitive and androgen-insensitive human prostate carcinoma cells. Toxicol Appl Pharmacol. 2000;164:82–90. doi: 10.1006/taap.1999.8885. [DOI] [PubMed] [Google Scholar]
- 53.Shammas M, Neri P, Koley H, et al. Specific killing of multiple myeloma cells by (−)-epigallocatechin-3-gallate extracted from green tea: biologic activity and therapeutic implications. Blood. 2006;108:2804–2810. doi: 10.1182/blood-2006-05-022814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Shimizu M, Deguchi A, Lim JT, et al. (−)-Epigallocatechin gallate and polyphenon E inhibit growth and activation of the epidermal growth factor receptor and human epidermal growth factor receptor-2 signaling pathways in human colon cancer cells. Clin Cancer Res. 2005;11:2735–2746. doi: 10.1158/1078-0432.CCR-04-2014. [DOI] [PubMed] [Google Scholar]
- 55.Ahn WS, Huh SW, Bae SM, et al. A major constituent of green tea, EGCG, inhibits the growth of a human cervical cancer cell line, CaSki cells, through apoptosis, G(1) arrest, and regulation of gene expression. DNA Cell Biol. 2003;22:217–224. doi: 10.1089/104454903321655846. [DOI] [PubMed] [Google Scholar]
- 56.Stearns ME, Wang M. Synergistic effects of the green tea extract epigallocatechin-3-gallate in eradication of malignant human prostate tumors. Transl Oncol. 2011;4:147–156. doi: 10.1593/tlo.10286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Luo T, Wang J, Yin Y, et al. (−)-Epigallocatechin gallate sensitizes bre ast cancer cells to paclitaxel in a murine model of breast carcinoma. Breast Cancer Res. 2010;12:R8. doi: 10.1186/bcr2473. doi: 10.1186/bcr2473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Sadzuka Y, Sugiyama T, Hirota S. Modulation of cancer chemotherapy by green tea. Clin Cancer Res. 1998;4:153–156. [PubMed] [Google Scholar]
- 59.Bettuzzi S, Brausi M, Rizzi F, et al. Chemoprevention of human prostate cancer by oral administration of green tea catechins in volunteers with high-grade prostate intraepithelial neoplasia: a preliminary report from a one-year proof-of-principle study. Cancer Res. 2006;66:1234–1240. doi: 10.1158/0008-5472.CAN-05-1145. [DOI] [PubMed] [Google Scholar]
- 60.Jatoi A, Ellison N, Burch PA, et al. A phase II trial of green tea in the treatment of patients with androgen independent metastatic prostate carcinoma. Cancer. 2003;97:1442–1446. doi: 10.1002/cncr.11200. [DOI] [PubMed] [Google Scholar]
- 61.Ruby AJ, Kuttan G, Babu KD, et al. Anti-tumour and antioxidant activity of natural curcuminoids. Cancer Lett. 1995;94:79–83. doi: 10.1016/0304-3835(95)03827-j. [DOI] [PubMed] [Google Scholar]
- 62.Ruby AJ, Kuttan G, Babu KD, et al. Antitumour and antioxidant activity of natural curcuminoids. Cancer Lett. 1995;94:79–83. doi: 10.1016/0304-3835(95)03827-j. [DOI] [PubMed] [Google Scholar]
- 63.Aggarwal BB, Sundaram C, Malani N, et al. Curcumin: the Indian solid gold. Adv Exp Med Biol. 2007;595:1–75. doi: 10.1007/978-0-387-46401-5_1. [DOI] [PubMed] [Google Scholar]
- 64.Hebert JR, Hurley TG, Olendzki BC, et al. Nutritional and socioeconomic factors in relation to prostate cancer mortality: a crossnational study. J Natl Cancer Inst. 1998;90:1637–1647. doi: 10.1093/jnci/90.21.1637. [DOI] [PubMed] [Google Scholar]
- 65.Cheng AL, Hsu CH, Lin JK, et al. Phase I clinical trial of curcumin, a chemopreventive agent, in patients with high-risk or pre-malignant lesions. Anticancer Res. 2001;21:2895–2900. [PubMed] [Google Scholar]
- 66.Kanai M, Imaizumi A, Otsuka Y, et al. Dose-escalation and pharmacokinetic study of nanoparticle curcumin, a potential anticancer agent with improved bioavailability, in healthy human volunteers. Cancer Chemother Pharmacol. 2012;69:65–70. doi: 10.1007/s00280-011-1673-1. [DOI] [PubMed] [Google Scholar]
- 67.Mukhopadhyay A, Bueso-Ramos C, Chatterjee D, et al. Curcumin downregulates cell survival mechanisms in human prostate cancer cell lines. Oncogene. 2001;20:7597–7609. doi: 10.1038/sj.onc.1204997. [DOI] [PubMed] [Google Scholar]
- 68.Mukhopadhyay A, Banerjee S, Stafford LJ, et al. Curcumin-induced suppression of cell proliferation correlates with down-regulation of cyclin D1 expression and CDK4-mediated retinoblastoma protein phosphorylation. Oncogene. 2002;21:8852–8861. doi: 10.1038/sj.onc.1206048. [DOI] [PubMed] [Google Scholar]
- 69.Li M, Zhang Z, Hill DL, et al. Curcumin, a dietary component, has anticancer, chemosensitization, and radiosensitization effects by down-regulating the MDM2 oncogene through the PI3K/mTOR/ETS2 pathway. Cancer Res. 2007;67:1988–1996. doi: 10.1158/0008-5472.CAN-06-3066. [DOI] [PubMed] [Google Scholar]
- 70.Yu S, Shen G, Khor TO, et al. Curcumin inhibits Akt/mammalian target of rapamycin signaling through protein phosphatase-dependent mechanism. Mol Cancer Ther. 2008;7:2609–2620. doi: 10.1158/1535-7163.MCT-07-2400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Beevers CS, Li F, Liu L, et al. Curcumin inhibits the mammalian target of rapamycin-mediated signaling pathways in cancer cells. Int J Cancer. 2006;119:757–764. doi: 10.1002/ijc.21932. [DOI] [PubMed] [Google Scholar]
- 72.Thangapazham RL, Shaheduzzaman S, Kim KH, et al. Androgen responsive and refractory prostate cancer cells exhibit distinct curcumin regulated transcriptome. Cancer Biol Ther. 2008;7:1427–1435. doi: 10.4161/cbt.7.9.6469. [DOI] [PubMed] [Google Scholar]
- 73.Tomlins SA, Rhodes DR, Perner S, et al. Recurrent fusion of TMPRSS2 and ETS transcription factor genes in prostate cancer. Science. 2005;310:644–648. doi: 10.1126/science.1117679. [DOI] [PubMed] [Google Scholar]
- 74.Hour TC, Chen J, Huang CY, et al. Curcumin enhances cytotoxicity of chemotherapeutic agents in prostate cancer cells by inducing p21WAF1/CIPI and C/EBPβ expressions and suppressing NF-κB activation. Prostate. 2002;51:211–218. doi: 10.1002/pros.10089. [DOI] [PubMed] [Google Scholar]
- 75.Bava SV, Puliappadamba VT, Deepti, et al. Sensitization of taxol-induced apoptosis by curcumin involves down-regulation of nuclear factor-κB and the serine/threonine kinase Akt and is independent of tubulin polymerization. J Biol Chem. 2005;280:6301–6308. doi: 10.1074/jbc.M410647200. [DOI] [PubMed] [Google Scholar]
- 76.Kamat AM, Sethi G, Aggarwal BB, et al. Curcumin potentiates the apoptotic effects of chemotherapeutic agents and cytokines through down-regulation of nuclear factor-KB and nuclear factor-KB–regulated gene products in IFN-A–sensitive and IFN-A–resistant human bladder cancer cells. Mol Cancer Ther. 2007;6:1022–1030. doi: 10.1158/1535-7163.MCT-06-0545. [DOI] [PubMed] [Google Scholar]
- 77.Lee JY, Lee YM, Chang GC, et al. Curcumin induces EGFR degradation in lung adenocarcinoma and modulates p38 activation in intestine: the versatile adjuvant for gefitinib therapy. PLoS One. 2011;6:e23756. doi: 10.1371/journal.pone.0023756. doi: 10.1371/journal.pone. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kunnumakkara AB, Guha S, Krishnan S, et al. Curcumin potentiates antitumor activity of gemcitabine in an orthotopic model of pancreatic cancer through suppression of proliferation, angiogenesis, and inhibition of nuclear factor-kappaB-regulated gene products. Cancer Res. 2007;67:3853–3861. doi: 10.1158/0008-5472.CAN-06-4257. [DOI] [PubMed] [Google Scholar]
- 79.Ali S, Ahmad A, Banerjee S, et al. Gemcitabine sensitivity can be induced in pancreatic cancer cells through modulation of miR-200 and miR-21 expression by curcumin or its analogue CDF. Cancer Res. 2010;70:3606–3617. doi: 10.1158/0008-5472.CAN-09-4598. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 80.Anand P, Sundaram C, Jhurani S, et al. Curcumin and cancer: an “old-age” disease with an “age-old”solution. Cancer Lett. 2008;267:133–164. doi: 10.1016/j.canlet.2008.03.025. [DOI] [PubMed] [Google Scholar]
- 81.Jang M, Cai L, Udeani GO, et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science. 1997;275:218–220. doi: 10.1126/science.275.5297.218. [DOI] [PubMed] [Google Scholar]
- 82.Subramanian L, Youssef S, Bhattacharya S, et al. Resveratrol: challenges in translation to the clinic–a critical discussion. Clin Cancer Res. 2010;16:5942–5948. doi: 10.1158/1078-0432.CCR-10-1486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Shi WF, Leong M, Cho E, et al. Repressive effects of resveratrol on androgen receptor transcriptional activity. PLoS One. 2009;4:e7398. doi: 10.1371/journal.pone.0007398. doi: 10.1371/journal.pone. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Harada N, Murata Y, Yamaji R, et al. Resveratrol down-regulates the androgen receptor at the post-translational level in prostate cancer cells. J Nutr Sci Vitaminol (Tokyo) 2007;53:556–560. doi: 10.3177/jnsv.53.556. [DOI] [PubMed] [Google Scholar]
- 85.Fu M, Liu M, Sauve AA, et al. Hormonal control of androgen receptor function through SIRT1. Mol Cell Biol. 2006;26:8122–8135. doi: 10.1128/MCB.00289-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Faus H, Haendler B. Androgen receptor acetylation sites differentially regulate gene control. J Cell Biochem. 2008;104:511–524. doi: 10.1002/jcb.21640. [DOI] [PubMed] [Google Scholar]
- 87.Kuwajerwala N, Cifuentes E, Gautam S, et al. Resveratrol induces prostate cancer cell entry into s phase and inhibits DNA synthesis. Cancer Res. 2002;62:2488–2492. [PubMed] [Google Scholar]
- 88.Gao S, Liu GZ, Wang Z. Modulation of androgen receptor-dependent transcription by resveratrol and genistein in prostate cancer cells. Prostate. 2004;59:214–225. doi: 10.1002/pros.10375. [DOI] [PubMed] [Google Scholar]
- 89.Benitez DA, Pozo-Guisado E, Alvarez-Barrientos A, et al. Mechanisms involved in resveratrol-induced apoptosis and cell cycle arrest in prostate cancer-derived cell lines. J Androl. 2007;28:282–293. doi: 10.2164/jandrol.106.000968. [DOI] [PubMed] [Google Scholar]
- 90.Benitez DA, Pozo-Guisado E, Clementi M, et al. Non-genomic action of resveratrol on androgen and oestrogen receptors in prostate cancer: modulation of the phosphoinositide 3-kinase pathway. Br J Cancer. 2007;96:1595–1604. doi: 10.1038/sj.bjc.6603755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Chen Q, Ganapathy S, Singh KP, et al. Resveratrol induces growth arrest and apoptosis through activation of FOXO transcription factors in prostate cancer cells. PLoS One. 2010;5:e15288. doi: 10.1371/journal.pone.0015288. doi: 10.1371/journal.pone. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Park SJ, Ahmad F, Philip A, et al. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phospodiesterases. Cell. 2012;148:421–433. doi: 10.1016/j.cell.2012.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Walle T. Bioavailability of resveratrol. Ann N Y Acad Sci. 2011;1215:9–15. doi: 10.1111/j.1749-6632.2010.05842.x. [DOI] [PubMed] [Google Scholar]
- 94.Walle T, Hsieh F, DeLegge MH, et al. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab Dispos. 2004;32:1377–1382. doi: 10.1124/dmd.104.000885. [DOI] [PubMed] [Google Scholar]
- 95.Goldberg DM, Yan J, Soleas GJ. Absorption of three wine-related polyphenols in three different matrices by healthy subjects. Clin Biochem. 2003;36:79–87. doi: 10.1016/s0009-9120(02)00397-1. [DOI] [PubMed] [Google Scholar]
- 96.Rashid A, Liu C, Sanli T, et al. Resveratrol enhances prostate cancer cell response to ionizing radiation. Modulation of the AMPK, Akt and mTOR pathways. Radiat Oncol. 2011;6:144–156. doi: 10.1186/1748-717X-6-144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Tseng SH, Lin SM, Chen JC, et al. Resveratrol suppresses the angiogenesis and tumor growth of gliomas in rats. Clin Cancer Res. 2004;10:2190–2202. doi: 10.1158/1078-0432.ccr-03-0105. [DOI] [PubMed] [Google Scholar]
- 98.Garvin S, Ollinger K, Dabrosin C. Resveratrol induces apoptosis and inhibits angiogenesis in human breast cancer xenografts in vivo. Cancer Lett. 2006;231:113–122. doi: 10.1016/j.canlet.2005.01.031. [DOI] [PubMed] [Google Scholar]
- 99.Kimura Y, Okuda H. Resveratrol isolated from polygonum cuspidatum root prevents tumor growth and metastasis to lung and tumorinduced neovascularization in lewis lung carcinoma-bearing mice. J Nutr. 2001;131:1844–1849. doi: 10.1093/jn/131.6.1844. [DOI] [PubMed] [Google Scholar]
- 100.Hudson TS, Hartle DK, Hursting SD, et al. Inhibition of prostate cancer growth by muscadine grape skin extract and resveratrol through distinct mechanisms. Cancer Res. 2007;67:8396–8405. doi: 10.1158/0008-5472.CAN-06-4069. [DOI] [PubMed] [Google Scholar]
- 101.Singh RP, Dhanalakshmi S, Tyagi AK, et al. Dietary feeding of silibinin inhibits advance human prostate carcinoma growth in athymic nude mice and increases plasma insulin-like growth factor binding protein-3 levels. Cancer Res. 2002;62:3063–3069. [PubMed] [Google Scholar]
- 102.Wellington K, Jarwis B. Silymarin: a review of its clinical properties in the management of hepatic disorders. BioDrugs. 2001;15:465–489. doi: 10.2165/00063030-200115070-00005. [DOI] [PubMed] [Google Scholar]
- 103.Ferenci P, Dragosics B, Dittrich H, et al. Randomized controlled trial of silymarin treatment in patients with cirrhosis of the liver. J Hepatol. 1989;9:105–113. doi: 10.1016/0168-8278(89)90083-4. [DOI] [PubMed] [Google Scholar]
- 104.Letteron P, Labbe G, Degott C, et al. Mechanism for the protective effects of silymarin against carbon tetrachloride-induced lipid peroxidation and hepatotoxicity in mice. Evidence that silymarin acts both as an inhibitor of metabolic activation and as a chain-breaking antioxidant. Biochem Pharmacol. 1990;39:2027–2034. doi: 10.1016/0006-2952(90)90625-u. [DOI] [PubMed] [Google Scholar]
- 105.Stattin P, Bylund A, Rinaldi S, et al. Plasma insulin-like growth factor-I insulin-like growth factor binding proteins, and prostate cancer risk: a prospective study. J Natl Cancer Inst. 2000;92:1910–1917. doi: 10.1093/jnci/92.23.1910. [DOI] [PubMed] [Google Scholar]
- 106.Zi X, Zhang J, Agarwal R, et al. Silibinin up-regulates insulin-like growth factor-binding protein 3 expression and inhibits proliferation of androgen-independent prostate cancer cells. Cancer Res. 2000;60:5617–5620. [PubMed] [Google Scholar]
- 107.Roddam AW, Allen NE, Appleby PE, et al. Insulin like growth factors, their binding proteins, and prostate cancer risk: analysis of individual patient data from 12 prospective studies. Ann Intern Med. 2008;149:461–471. doi: 10.7326/0003-4819-149-7-200810070-00006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Grimberg A, Cohen P. Role of insulin-like growth factors and their binding proteins in growth control and carcinogenesis. J Cell Physiol. 2000;183:1–9. doi: 10.1002/(SICI)1097-4652(200004)183:1<1::AID-JCP1>3.0.CO;2-J. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Tyagi AK, Singh RP, Agarwal C, et al. Silibinin strongly synergizes human prostate carcinoma DU145 cells to doxorubicin-induced growth inhibition, G2-M arrest, and apoptosis. Clin Cancer Res. 2002;8:3512–3519. [PubMed] [Google Scholar]
- 110.Flaig TW, Su LJ, Harrison G, et al. Silibinin synergizes with mitoxantrone to inhibit cell growth and induce apotosis in human prostate cancer cells. Int J Cancer. 2007;120:2028–2033. doi: 10.1002/ijc.22465. [DOI] [PubMed] [Google Scholar]
- 111.Hoh C, Boocock D, Marczylo T, et al. Pilot study of oral silibinin, a putative chemopreventive agent, in colorectal cancer patients: silibinin levels in plasma, colorectum, and liver and their pharmacodynamic consequences. Clin Cancer Res. 2006;12:2944–2950. doi: 10.1158/1078-0432.CCR-05-2724. [DOI] [PubMed] [Google Scholar]
- 112.Flaig TW, Glode M, Gustafson D, et al. A study of high-dose oral silybin-phytosome followed by prostatectomy in patients with localized prostate cancer. Prostate. 2010;70:848–855. doi: 10.1002/pros.21118. [DOI] [PubMed] [Google Scholar]
- 113.Chen S, Wang X. Herbal Composition for Treating Prostate Carcinoma. U.S. Patent 5665393. 1997 Sep 9;
- 114.Halicka H, Ardelt B, Juan G, et al. Apoptosis and cell cycle effects induced by extracts of the Chinese herbal preparation PC-SPES. Int J Oncol. 1997;11:437–448. doi: 10.3892/ijo.11.3.437. [DOI] [PubMed] [Google Scholar]
- 115.Tiwari RK, Geliebter J, Garikapaty VP, et al. Anti-tumor effects of PC-SPES, an herbal formulation in prostate cancer. Int J Oncol. 1999;14:713–719. doi: 10.3892/ijo.14.4.713. [DOI] [PubMed] [Google Scholar]
- 116.Oh WK, George DJ, Hackmann K, et al. Activity of the herbal combination, PC-SPES, in the treatment of patients with androgen independent prostate cancer. Urology. 2001;57:122–126. doi: 10.1016/s0090-4295(00)00986-9. [DOI] [PubMed] [Google Scholar]
- 117.de la Taille A, Buttyan R, Hayek O, et al. Herbal therapy PC-SPES: in vitro effects and evaluation of its efficacy in 69 patients with prostate cancer. J Urol. 2000;164:1229–1234. doi: 10.1097/00005392-200010000-00021. [DOI] [PubMed] [Google Scholar]
- 118.Oh WK, Kantoff PW, Weinberg V, et al. Prospective, multicenter, randomized phase II trial of the herbal supplement, PC-SPES, and diethylstilbestrol in patients with androgen-independent prostate cancer. J Clin Oncol. 2004;22:3705–3712. doi: 10.1200/JCO.2004.10.195. [DOI] [PubMed] [Google Scholar]
- 119.Zyflamend product info. [Accessed 12/7/2011]. Available at http://www.newchapter.com/zyflamend/zyflamend#supplement-facts.
- 120.Bemis DL, Capodice JL, Anastasiadis AG, et al. Zyflamend®, a unique herbal preparation with nonselective COX inhibitory activity, induces apoptosis of prostate cancer cells that lack COX-2 expression. Nutr Cancer. 2005;52:202–212. doi: 10.1207/s15327914nc5202_10. [DOI] [PubMed] [Google Scholar]
- 121.Huang EC, Chen G, Baek SJ, et al. Zyflamend reduces the expression of androgen receptor in a model of castrate resistant prostate cancer. Nutr Cancer. 2011;63:1287–1296. doi: 10.1080/01635581.2011.606956. [DOI] [PubMed] [Google Scholar]
- 122.Sandur SK, Ahn KS, Ichikawa H, et al. Zyflamend, a polyherbal preparation, inhibits invasion, suppresses osteoclastogenesis, and potentiates apoptosis through down-regulation of NF-κB activation and NF-κB–regulated gene products. Nutr Cancer. 2007;57:78–87. doi: 10.1080/01635580701268295. [DOI] [PubMed] [Google Scholar]
- 123.Yan J, Xie B, Capodice JL, et al. Zyflamend inhibits the expression and function of androgen receptor and acts synergistically with bicalutimide to inhibit prostate cancer cell growth. Prostate. 2012;72:244–252. doi: 10.1002/pros.21426. doi: 10.1002/pros.21426. [DOI] [PubMed] [Google Scholar]
- 124.Kunnumakkara AB, Sung B, Ravindran J, et al. Zyflamend suppresses growth and sensitizes human pancreatic tumors to gemcitabine in an orthotopic mouse model through modulation of multiple targets. Int J Cancer. 2011 Sep 20; doi: 10.1002/ijc.26442. [E-pub ahead of print]. doi: 10.1002/ijc.26442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Clement J, Bubley G. Prostasol and venous thromboembolism. Urology. 2008;72:664–666. doi: 10.1016/j.urology.2008.04.045. [DOI] [PubMed] [Google Scholar]