

Abstract
Myotonic Dystrophy Type-2 (DM2) is an autosomal dominant disease caused by the expansion of a CCTG tetraplet repeat. It is a multisystemic disorder, affecting skeletal muscles, the heart, the eye, the central nervous system and the endocrine system. Since microRNA (miRNA) expression is disrupted in Myotonic Dystrophy Type-1 and many other myopathies, miRNAs deregulation was studied in skeletal muscle biopsies of 13 DM2 patients and 13 controls. Eleven miRNAs were deregulated: 9 displayed higher levels compared to controls (miR-34a-5p, miR-34b-3p, miR-34c-5p, miR-146b-5p, miR-208a, miR-221-3p and miR-381), while 4 were decreased (miR-125b-5p, miR-193a-3p, miR-193b-3p and miR-378a-3p). To explore the relevance of DM2 miRNA deregulation, the predicted interactions between miRNA and mRNA were investigated. Global gene expression was analyzed in DM2 and controls and bioinformatic analysis identified more than 1,000 miRNA/mRNA interactions. Pathway and function analysis highlighted the involvement of the miRNA-deregulated mRNAs in multiple aspects of DM2 pathophysiology. In conclusion, the observed miRNA dysregulations may contribute to DM2 pathogenetic mechanisms.
Introduction
Myotonic Dystrophy (DM) is an autosomal dominant disease and the most common muscular dystrophy of the adult, with a prevalence of 1/8,000 worldwide [1]. DM is a progressive multisystemic disorder characterized by a variety of manifestations that include muscle hyperexcitability (myotonia), muscular dystrophy, cardiac conduction defects, dilated cardiomyopathy, insulin-resistance, cataracts, neurological involvement, and a multiplicity of serological abnormalities. The severity and the number of symptoms are highly variable and can range from minimal signs, to facial and muscle wasting and weakness, to severe congenital disorder [1]. Respiratory failure and cardiac arrhythmias are among the main causes of premature death in DM patients [1]. At the histological level, pathological features of the DM skeletal muscle include the concomitant presence of atrophic and hypertrophic myofibers, increased number of central nuclei, and presence of fibers with nuclear clumps [2].
Two types of DM have been described. The most common form of DM, named type 1 or Steinert’s disease (DM1, OMIM 160900), is caused by an expanded (CTG)n in the 3′ untranslated region of the Dystrophia Myotonica Protein Kinase gene [3], [4], [5]. The second form of DM is named type 2 or PROximal Myotonic Myopathy-PROMM (DM2, OMIM 602688): it displays a prevalently proximal impairment and milder clinical symptoms than DM1, since it does not show a congenital form or anticipation i.e. a phenomenon of progressively earlier and more severe manifestation of the disease. DM2 is caused by the expansion of a tetranucleotidic repetition (CCTG)n in the first intron of the CCHC-type zinc finger, nucleic acid binding protein (CNBP) [6].
Several lines of evidence indicate that an RNA-mediated toxic gain-of-function is the common pathogenetic mechanism of the two DM types. In both DM1 and DM2, the mutation causes the accumulation of the expanded CUG/CCUG transcripts into discrete nuclear RNA foci. The expanded RNA that is trapped in the nucleus, deregulates the interaction of RNA binding proteins, which are essential for mRNA splicing and other cell processes, leading to aberrant RNA maturation and cell dysfunction [7].
MicroRNAs (miRNAs) are short non-coding RNAs that regulate the stability and/or the translational efficiency of target mRNAs. miRNAs have a very pervasive role: bioinformatic studies estimate that >60% of all transcripts are regulated by one or more miRNA [8]. Indeed, miRNAs have a demonstrated impact on virtually all cell functions, including differentiation, proliferation and apoptosis [9], [10]. Specifically, miRNA sequences can regulate skeletal and cardiac muscle function in both development and disease, such as ischemia and muscular dystrophy [11], [12].
We and others demonstrated miRNA deregulation in Duchenne Muscular Dystrophy patients and in mdx mice, both lacking a functional dystrophin gene [13], [14], [15], [16]. Moreover, Eisenberg and collaborators described miRNA expression profiles in skeletal muscle from various human primary muscle disorders and identified a series of miRNAs that are deregulated in almost all myopathies analyzed [14]. Finally, we and others analyzed the expression of miRNAs in DM1 and identified specific miRNAs displaying altered expression and localization [17], [18], [19].
In this study, miRNA expression profiles were measured in the skeletal muscle of DM2 patients. We identified a subset of miRNAs that are specifically deregulated potentially contributing to DM2 pathogenetic mechanisms.
Materials and Methods
Patient Selection and Skeletal Muscle Biopsies
This study was authorized by the Institutional Ethics Committee (ASL MI2) and was conducted according to the principles expressed in the Declaration of Helsinki, the institutional regulation and Italian laws and guidelines. All biopsy specimens were taken after specific written informed consent was obtained. Human muscle biopsies from biceps brachii were harvested under sterile conditions and snap-frozen in liquid nitrogen. Clinical diagnosis of DM2 patients was based upon the criteria set by the International Consortium for Myotonic Dystrophies [20]. Fluorescence in situ hybridization was performed on frozen muscle sections to confirm DM2 diagnosis according to Cardani et al. [21]. Control (CTR) biopsies were from subjects admitted with suspected neuromuscular disorder of undetermined nature. CTR biopsies did not show overt signs of muscle pathology on histological and immunohistochemical examination. All muscle biopsies were processed by the same pathology team. Immunohistochemical analysis of atrophy and hypertrophy in fast and slow fibers was carried out as previously described [22].
RNA Isolation and miRNA Profiling
Total RNA was extracted by Trizol (Invitrogen) and the TissueLyser system (Qiagen). TaqMan Low Density miRNA arrays v.2 (Applied Biosystems) were used to measure 365 human mature miRNAs, as previously described [23]. After median Ct value normalization, relative miRNA expression was calculated with the 2−ΔΔCt method [24]. We further analyzed those miRNAs with a Ct≤33, displaying a significant fold change ≥2. Data distribution was evaluated by the Kolmogorov-Smirnov’s normality test. Permutational multivariate analysis of variance was performed by the multivariate non-parametric PERMANOVA test, using 9,999 permutations, Bonferroni post-hoc test and Gower test for distance measurement. Adjusted p<0.05 was considered as statistically significant.
mRNA Profiling
Gene expression profiles were measured using the Gene Chip Human Exon 1.0 ST Array (Affymetrix). One hundred ng of total RNA were analyzed according to the manufacturer instructions. Data analysis was computed using Partek Genomics Suite (Partek Inc.) according to MIAME guidelines. The output signals were normalized using the Partek Robust Multi-Chip Average analysis adjusted for the GC content of probe sequence (GCRMA). The genes that were differentially expressed among the two classes were identified using the Analysis of Variance (ANOVA) as implemented in the Partek Gene Expression tool. An estimate of the associated False Discovery Rate (FDR) was computed per gene using the method of Benjamini and Hochberg [25]. mRNA transcripts were considered to be significantly differentially expressed if they obtained ANOVA p-value<0.005 and FDR<0.15.
qPCR Assay
Individual mature miRNAs were measured using TaqMan MicroRNA single assays (Applied Biosystems) and samples were normalized to miR-16 expression [26]. MiRNA profiling indicated that miR-16 was not-modulated in DM2 patients. mRNA levels were measured using the SYBR-GREEN qPCR method (Applied Biosystems) and samples were normalized to GAPDH. For both miRNAs and mRNAs, relative expression was calculated using the comparative Ct method 2–ΔΔCt [24]. Each sample was measured in triplicate and values were averaged.
DM2 miRNA Score
To calculate the DM2 miRNA scores, the median fold change of all the deregulated miRNAs was calculated for each subject. For negatively regulated miRNAs, the minus sign was inverted before calculating the median.
Bioinformatic Analysis
Bioinformatic prediction of miRNA target genes was performed using miRonTop [27]. Predicted miRNA target genes were compared to differently expressed genes (p≤0.01) displaying an opposite modulation with their corresponding miRNA. The correlation analysis between mRNA and miRNA expression was evaluated by the quantitative trait tool in BRB ArrayTools (Richard Simon and BRB-ArrayTools Development Team) (p≤0.01).
Pathway analysis was performed using Ingenuity Pathways Knowledge Base (version 8.8, Ingenuity Systems) as reference set and assuming direct and indirect relationships. A Fisher’s exact test p-value <0.05 was deemed as statistically significant.
Statistics
Continuous variables are expressed as mean±standard error (SE), unless indicated differently. For group-wise comparisons, Mann–Whitney test or t-test were used as appropriate. For the DM2 miRNAs-score, Mann-Whitney’s variance analysis was used. The ability to discriminate between the DM2 and control groups was characterized by the receiver operating characteristic (ROC) curve, and the area under the ROC curve (AUC) was calculated. Spearman rank correlation was used to compare the DM2 miRNA score with atrophy/hypertrophy fiber index. All tests were performed 2-sided and a p≤0.05 was considered as statistically significant. For statistical analysis and heat map data visualization, GraphPad Prism v.4.03 (GraphPad Software Inc.) and Genesis 1.7.2 (Graz University of Technology, Institute for Genomics and Bioinformatics) softwares were used, respectively.
Results
Identification of Differentially Expressed miRNAs in DM2 Patients
Patients were selected so that DM2 and controls (CTR), displayed similar age and sex distribution (Table 1). Most DM2 patients had myotonia and cataract, two disease hallmarks, while differences in other clinical parameters were not significant. The experimental plan is schematized in Figure 1.
Table 1. Characteristics of Profiling Cohorts (Mean±Se).
| PARAMETER | DM2 (N = 13) | CONTROLS (N = 13) | p |
| AGE (years) | 56.9±1.9 | 49.3±2.5 | ns |
| SEX (f/m) | 5/8 | 5/8 | ns |
| STRENGHT (biopsied muscle) (MRC scale) | 4.5±0.1 | 4.9±0.04 | ns |
| STRENGHT (Megascore on 15 muscles, MRC scale) | 137.4±9.1 | 145.6±6.8 | ns |
| MYOTONIA (%) | 85 | 0 | <0.0001 |
| GLUCOSE (mg/dl) (n.v.70–110) | 101.4±16.0 | 86.0±8.5 | ns |
| DIABETES MELLITUS (%) | 18 | 0 | ns |
| CPK (IU/l) (n.v. m<190, f<167) | 292.7±3.5 | 236.4±67.1 | ns |
| FT3 (pg/ml) (n.v. 1.5–4.1) | 3.0±3.1 | 1.7±0.5 | ns |
| FT4 (ng/dl) (n.v. 0.8–1.9) | 1.3±0.3 | 1.2±0.3 | ns |
| TSH (mIU/ml) (n.v. 0.4–4.0) | 1.0±0.2 | 0.7±0.2 | ns |
| ECG-QRS duration (msec) (n.v. 60–100) | 94.3±16.4 | 85.9±12.1 | ns |
| EF (%) (n.v.>50) | 54.5±9.7 | 64.0±19.3 | ns |
| ICM (%) | 9 | 0 | ns |
| SERIC β-GLOBULIN (%) | 12.3±1.7 | 14.1±2.0 | ns |
| CATARACT (%) | 70 | 0 | <0.0005 |
Abbreviations: MRC scale = Medical Research Council scale (0–5 grade); CPK = Creatine PhosphoKinase; FT3 = free-triiodothyronine; FT4 = free-thyroxine; TSH = thyroid stimulating hormone; ECG = electrocardiogram; QRS duration = QRS complex of ECG corresponds to the depolarization of the right and left ventricles of the human heart; EF = ejection fraction; ICM = ischemic cardiomyopathy.
Figure 1. Experimental Plan.
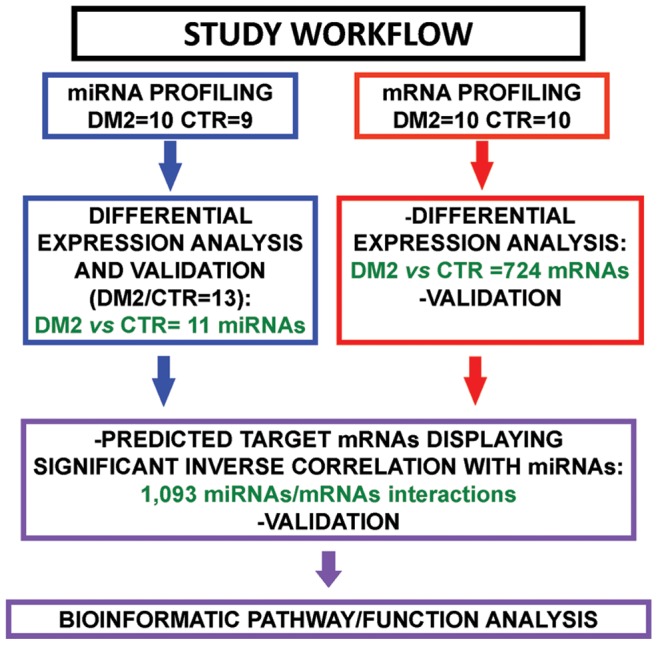
miRNA (blue) and mRNA (red) expression patterns were determined in DM2 and CTR, and significantly modulated miRNAs and mRNAs were identified. Validation was performed by qPCR for both miRNAs and mRNAs in 13 DM2 patients and 13 CTR. miRNA targets among the mRNAs identified by Class Comparison analysis were predicted by dedicated softwares. Only miRNA/mRNA couples displaying inverse significant correlation were selected and then analyzed by Ingenuity Pathway Analysis software that allowed to identify enriched molecular pathways and functions.
Skeletal muscle biopsies were harvested from 10 DM2 patients and 9 CTR, and total RNA was extracted. MiRNA profiling showed that, out of 365 miRNA measured, 203 and 199 were readily detectable (Ct<33) in DM2 and CTR groups, respectively (see Gene Expression Omnibus database, GEO, GSE37794 for a complete gene list). Statistical analysis showed that 20 miRNAs were significantly modulated ≥2-fold in DM2 compared to CTR (Fig. 2) and these miRNAs were validated by more sensitive and specific qPCR assays. This validation step included the same patients and CTR used for profiling, and was extended to three more DM2 and four more CTR (n = 13 for each group). Moreover, we added six more miRNAs to the 20 modulated miRNAs: the muscle-specific miRNAs, miR-1 and miR-206 [28], [29], miR-221-3p, which is co-transcribed with miR-222-3p, miR-193a-3p and miR-146a since they belong to miR-193 and miR-146 families, respectively, and miR-499-5p, which is downstream of miR-208a network [30]. We found that 11 miRNAs were validated as significantly deregulated in DM2 patients (Fig. 3).
Figure 2. Profiling of miRNAs in DM2 patients and CTR.
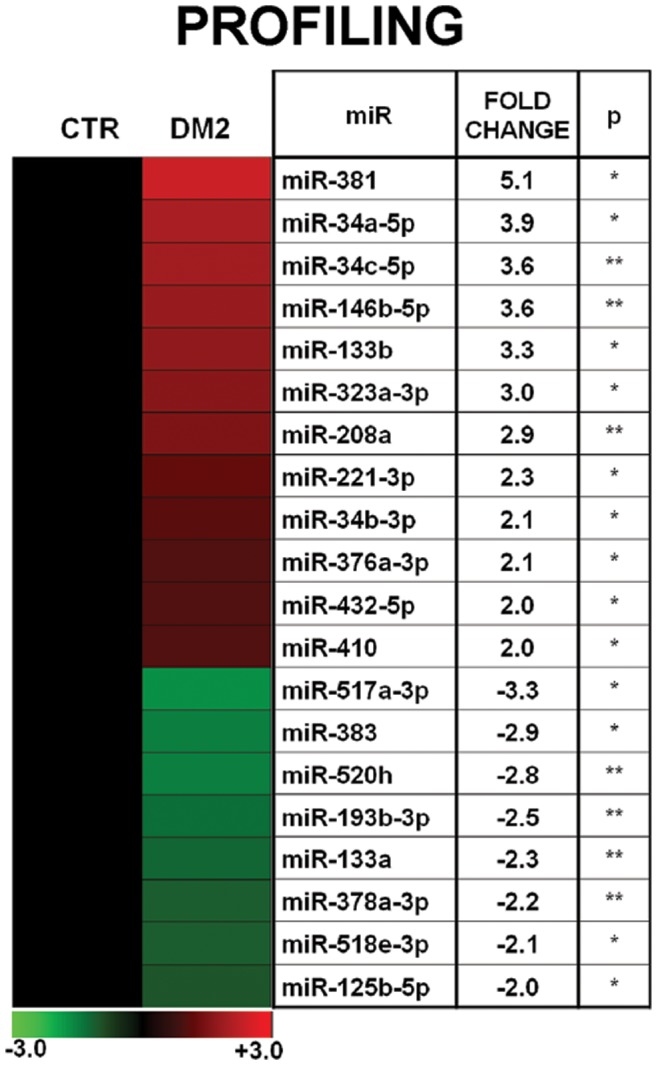
In the heat map on the left, mean miRNAs expression values are shown in a log2 scale (-ΔΔCt), where red and green indicate positive and negative modulation respectively. The table on the right shows the same values in a linear scale (DM2 = 13, CTR = 13; *p≤0.05 **p≤0.01).
Figure 3. Validation of miRNA modulations in DM1 and DM2 patients.

In the heat map on the left, the mean values of miRNA expression in both DM1 and DM2 compared to controls are shown in a log2 scale (-ΔΔCt), where red and green indicate positive and negative modulation respectively. On the right, the table shows the same values in a linear scale; statistically significant differences are highlighted in bold (DM2 n = 13, DM1 = 16, CTR n = 13; *p≤0.05 **p≤0.01 ***p≤0.001).
These DM2 miRNAs were also tested in an age and sex matched cohort of DM1 patients. We found that miR-193b-3p, miR-208a and miR-381 showed a similar significant modulation also in these patients (Fig. 3).
A DM2 miRNA Score Discriminates between Patients and Controls and Correlates with Myofiber Atrophy and Hypertrophy
DM2 deregulated miRNAs were also considered collectively: median fold change of the differentially expressed miRNAs was used to calculate a “DM2 miRNAs score” in each patient or CTR subject. The DM2 miRNA score allowed a significant separation between the DM2 and the CTR groups, with a median score of 2.3 in the DM2 group and 0.6 in the CTR group (Figure 4A). The ability of the DM2 miRNA score to discriminate DM2 patients from CTR was also demonstrated by the Receiver-Operator Characteristic (ROC) curve (Figure 4B). A ROC curve allows to visualize the reciprocal relationship between sensitivity and specificity when discriminating between control and DM2 values. The calculation of sensitivity and specificity depends on the threshold value used to separate the two classes; if the threshold value used is high, the specificity of the test increases, but the sensitivity decreases, conversely if the threshold is low, the test’s sensitivity is increased [31], [32]. By using a threshold score of 1.15 we achieved a sensitivity of 100% and a specificity of 92.3% for the identification of DM2 patients. Moreover, the Area Under Curve (AUC), which quantifies the overall ability of the test to discriminate between normal and diseased individuals, was 0.96, where perfect test (with zero false positives and zero false negatives) has an area of 1.00 [33].
Figure 4. Discrimination between DM2 and control groups using the DM2 miRNA score.

(A) Dot plot of the DM2 miRNA score in DM2 patients and CTR. The segments among the dots indicate median values for each group and the difference is statistically significant (DM2 = 13, CTR n = 13; p<0.0001). (B) ROC curve displaying DM2 miRNA score discrimination between the DM2 and CTR groups (AUC = 0.96).
At the histological level, a DM2 hallmark is represented by the concomitant presence of atrophic and hypertrophic myofibers, preferentially in type 2 (fast) fibers [2], [34], [35] (Figure 5A). We found that DM2 miRNA score correlated with both the atrophy (AR) and the hypertrophy (HR) index, but only in type 2 myofibers (Figure 5B).
Figure 5. Correlation of atrophy and hypertrophy with DM2 miRNAs score.

A) Representative stainings of transverse sections of frozen DM2 muscle biopsies. Top panel shows a hematoxylin and eosin staining; bottom panel shows an immunostaining for fast myosin heavy chain. Atrophic (blue arrows) and hypertrophic (asterisks) myofibers, as well as nuclear clumps (red arrows) and internal nuclei (black arrows) are highlighted. B) Spearman’s correlation between DM2 miRNAs score and Atrophy (AR) or Hypertrophy (HR) indexes in fast and slow fibers (DM2 n = 13; CTR n = 9).
Transcriptome Analysis and Predicted miRNA/mRNA Interactions
To investigate the implications of miRNA dysregulations identified in DM2 patients, we tested their impact on the transcriptome using high-density oligonucleotide microarrays. The same RNAs used for miRNA profiling were analyzed, with the addition of one extra control (total, 10 DM2 and 10 CTR) (see Gene Expression Omnibus database, GEO, GSE37084 for a complete gene list). As expected, unsupervised hierarchical clustering analysis segregated the profiles of DM2 and CTR (Fig. S1). Class Comparison analysis versus the CTR group was performed identifying 724 non redundant transcripts differentially expressed in DM2 patients. qPCR validation allowed to confirm 10 out of 12 tested genes (not shown).
Next, we attempted to identify the contribution of miRNA deregulation to differential gene expression. Given the inhibitory nature of miRNAs [8], [36], mRNA displaying reciprocal modulation to DM2 miRNAs were selected. These transcripts were further filtered using target identification algorithms, to predict direct binding events. The application of these three sequential filers allowed the identification of 1,093 miRNA/mRNA interactions (Table S1).
It is worth noting that several mRNAs were predicted targets of more than one miRNA deregulated in DM2, with a potential signal reinforcement. A randomly selected subset of the identified miRNA-target modulations was further tested by qPCR and 15 out of 22 were validated (Fig. 6). The remaining 7 interactions originated from 3 transcripts (CARM1, NXN and YIPF7) that were not detectable in most controls and DM2 samples.
Figure 6. Validation of miRNA/mRNA interactions in DM2 patients.

Validation by qPCR of the expression of randomly selected mRNAs either up-(A) and down-(B) regulated in DM2 patients vs CTR. Expression level compared to CTR of the targeting miRNA is indicated for each mRNA (DM2 = 12, CTR = 10;*p≤0.05; **p≤0.01; ***p≤0.001; ****p≤0.0001).
Pathways Analysis of Gene Expression Changes
To facilitate the interpretation of the complex gene expression changes observed, we used Ingenuity Pathway Analysis software (IPA) exploring enriched biological functions and pathways. Both global gene expression changes and DM2 miRNA/mRNA interactions were analyzed.
To assess the potential functional relevance of the overall gene expression changes observed in DM2 patients, differentially expressed genes, analyzed by Class Comparison, were uploaded to the IPA database. Several interesting pathways and functions were enriched significantly in the DM2 group compared to CTR, such as “Mitochondrial Dysfunctions”, “TGF-β Signaling”, “Insulin Receptor Signaling”, “Calcium Signaling” for the pathways and “Skeletal and Muscular System Development”, “Cell Cycle”, “Cell Death”, “Skeletal and Muscular Disorders” for the enriched functions (Table S2 and S3). Moreover, “Cardiac Dilation”, “Cardiac Hypoplasia” and “Cardiac Artheriopathy” among the toxic functions were also found enriched (Table S4). Complete lists of enriched categories can be found in Table S8.
IPA analysis of miRNAs predicted targets differentially modulated in DM2 patients identified several important pathways enriched, such as: “Insulin Receptor Signaling”, “Integrin Signaling”, “PI3K/AKT Signaling” and “TGF-β Signaling” (Table 2 and Table S5). “Skeletal and Muscular Disorders”, “Ophthalmic Diseases” and “Endocrine System Disorders” and “Cardiac Damage”, “Cardiac Dilation” and “Heart Failure” were relevant enriched functions (Table 2 and Table S6) and toxic functions (Table 2 and Table S7), respectively, and all the enriched categories are showed in Table S9, where the selected categories of Table 2 are highlighted in bold.
Table 2. Pathways and Functions Analysis by IPA.
| PATHWAYS | p-value | ratio | enrichment index |
| Insulin Receptor Signaling | 0.002 | 11/139 | 263 |
| Integrin Signaling | 0.003 | 14/209 | 223 |
| FAK Signaling | 0.02 | 7/102 | 345 |
| PI3K/AKT Signaling | 0.03 | 8/140 | 285 |
| RAR Activation | 0.03 | 10/183 | 275 |
| TGF-β Signaling | 0.04 | 6/89 | 478 |
| FUNCTIONS | p-value | ratio | enrichment index |
| Skeletal and Muscular System Development and Function | 7.8E−06 | 50/2849 | 15 |
| Skeletal and Muscular Disorders | 2.7E−04 | 142/3851 | 11 |
| Neurological Disease | 2.4E−04 | 179/4743 | 9 |
| Cardiovascular Disease | 9.4E−04 | 110/2663 | 159 |
| Metabolic Disease | 9.4E−04 | 124/2496 | 17 |
| Endocrine System Disorders | 1.4E−03 | 10/2271 | 19 |
| Cell Growth and Proliferation | 2.1E−03 | 36/2884 | 15 |
| Cardiovascular System Development and Function | 2.7E−03 | 11/2738 | 15 |
| Ophthalmic Disease | 2.7E−03 | 3/987 | 43 |
| TOXIC FUNCTIONS | p-value | ratio | enrichment index |
| Cardiac Damage | 3.0E−02 | 1/88 | 481 |
| Cardiac Dilation | 3.0E−02 | 4/83 | 510 |
| Heart Failure | 4.8E−02 | 11/330 | 128 |
Table legend:
ratio = number of modulated genes/total number of genes present in the relevant IPA category.
enrichment index = ratio between observed genes and number of genes expected by chance in each category.
Discussion
Understanding the molecular mechanisms triggered by the tetranucleotide expansion in the Zinc Finger Protein-9 gene is of great importance to gain insight into the molecular mechanisms underpinning DM2 disease. For the first time, we measured the miRNA profile of skeletal muscle biopsies of DM2 patients, identifying 11 deregulated miRNAs. We and others found that miR-221-3p, miR-381, miR-34a-5p, miR-34c-5p, miR-146b-5p, and miR-193b-3p were similarly modulated in muscular diseases other than DM2 [13], [14] (Table S10), suggesting potential similarities in certain aspects of the pathogenetic mechanisms. The DM2 miRNAs were also measured in DM1 patients and miR-193b-3p, miR-208a and miR-381 were similarly modulated. The differences displayed by the remaining miRNAs did not reach statistical significance, possibly due to insufficient patient numerosity.
We found that a DM2 miRNA score allowed to discriminate DM2 patients from CTR with a good sensitivity and specificity. Furthermore, DM2 miRNA score correlated to atrophy and hypertrophy of type 2, fibers that are particularly affected in DM2, indicating that the deregulated miRNAs were related to the disease pathogenetic mechanism. Further studies are granted to explore DM2 miRNA score diagnostic potential.
The up-regulation of all members of miR-34 family appears particularly significant, given miR-34 role in cell differentiation, apoptosis and senescence [37]. It is also worth noting the up-regulation of miR-221-3p, playing an important role in myotube maturation and in the maintenance of the myofibrillar organization [38]. Finally, we demonstrated the down-regulation of miR-125b-5p, which negatively regulates myoblast differentiation and muscle regeneration [39].
In an attempt to gain further insight into the underlying molecular mechanisms, we investigated the transcripts predicted to be regulated by the DM2 miRNAs. In most circumstances, miRNA binding induces the degradation of the target mRNA [8], [40]. Thus, we measured the transcriptome of DM2 patients and used target prediction softwares [27] to identify the genes potentially regulated by the DM2 miRNAs, directly. To minimize the number of false positive miRNA/mRNA interactions, we used three selection criteria: 1) only significantly modulated miRNAs and mRNAs were considered; 2) given the inhibitory nature of miRNA function [8], [36], only miRNA/mRNAs couples displaying a statistically significant reciprocal correlation of their expression levels were included; 3) among the genes passing filters 1 and 2, predicted targets were identified by MiRonTop [27]. After this highly selective selection, 1,093 miRNA/mRNA interactions were identified. Several mRNAs were targeted by more than one miRNA, leading to a potential reinforcement of the final deregulation. For instance, EGF was a predicted target not only of the three members of the miR-34 family, but also of miR-208a and miR-381.
The increase of MBNL2, a miR-378a-3p predicted target, was another interesting finding. Indeed, a hallmark of DM2 is the nuclear accumulation of (CCUG)n repeats that bind and sequester muscleblind-like proteins MBNL1, MBNL2 and MBNL3 [41], [42], [43]. The observed induction of MBNL2 mRNA expression in DM2 may be an adaptive response counteracting the decreased bioavailability of MBNL.
To the best of our knowledge, this represents the first study investigating miRNA/transcriptome interactions in DM2 patients, paving the way to further investigations that are necessary to formally demonstrate cause/effect relationships between the DM2 miRNAs and their targets. Moreover, a potential caveat of our analysis is represented by the heterogeneity of the cell populations composing skeletal muscle tissue. Albeit myofibers constitute the majority of the tissue mass, it is possible that certain miRNAs and target mRNAs are deregulated in different cell types.
A bioinformatic analysis of the pathways and functions revealed that DM2 miRNA targets were potentially involved in most DM2 pathogenetic mechanisms. The functions “Skeletal and Muscular System Development and Function”, “Neurological Disease” and “Skeletal and Muscular Disorders” are of obvious relevance. The “PI3K/AKT signaling “and the “TGF-β signaling” pathways seem particularly important. Indeed, dysregulation of TGF-β signaling has been implicated in various pathological conditions affecting skeletal muscle, both inherited and acquired [44], and Akt2 expression is greatly increased during skeletal muscle cell differentiation and myocyte growth, suggesting a critical role of Akt2 in myogenesis [45], [46], [47]. Thus, both PI3K/AKT and TGF-β signaling may relate to the atrophic/hypertrophic myofiber alterations characterizing DM2.
DM2 is a multisystemic disease. Indeed, other pathways and functions are linked to aspects of DM2 that do not affect the skeletal muscle directly. The “Insulin Receptor Signaling” pathway as well as the “Metabolic Disease” and “Endocrine System Disorders” functions suggest the involvement of the relevant miRNA/mRNA couples in insulin resistance, a classic DM clinical feature, predisposing to Type 2 diabetes [1], [34], [48].
Albeit the assayed tissue was skeletal muscle, many functions and toxic functions related to heart and cardiovascular disease, in keeping with the cardiac conduction defects and the dilated cardiomyopathy commonly observed in DM. Another DM hallmark is cataract; thus, it was interesting that the “Ophthalmic Disease” function was also represented.
Looking at the global alterations of the transcriptome, to the best of our knowledge, only one previous study exists [22]. However, a direct comparison with our investigation is limited by the fact that Vihola et al. narrowed their scope to 327 probesets functionally associated with muscle a priori. Nevertheless, it is interesting to note that both studies identified several altered genes involved in Calcium handling and signaling, that may relate to the abnormal calcium handling and myotonia observed in DM [49], [50], [51]. Moreover, in keeping with increased oxidative stress levels measured in DM1 patients [52], we found that “NRF2-mediated Ox-Stress Response” and “mitochondrial dysfunction” pathways were also enriched.
Conclusions
In conclusion, we identified a small subset of miRNA whose expression was deregulated in DM2. These findings may improve our understanding of the molecular mechanisms linking (CCTG)n expansion to disease.
Supporting Information
Unsupervised hierarchical clustering of mRNA expression differentiating DM2 (n = 10) from controls (n = 10). Each row represents an mRNA, and each column represents an individual. A color code represents the relative intensity of the expression signal, with red indicating high expression and blue indicating low expression.
(PPT)
Transcriptome analysis identified 1,093 miRNA/mRNA interactions. Table represents the identified miRNAs/mRNA interactions, where green and red colours indicate down- or up-regulation of miRNA or mRNA compared to controls, respectively.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in transcriptome. Some important enriched pathways, functions and toxic functions and relative miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S8.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in transcriptome. Some important enriched pathways, functions and toxic functions and relative miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S8.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in transcriptome. Some important enriched pathways, functions and toxic functions and relative miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S8.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in miRNAs targets analysis. Some important enriched pathways, functions and toxic functions in miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S9.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in miRNAs targets analysis. Some important enriched pathways, functions and toxic functions in miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S9.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in miRNAs targets analysis. Some important enriched pathways, functions and toxic functions in miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S9.
(XLS)
Ingenuity Pathway Analysis (IPA) of enriched categories in transcriptome. Statistical significance and complete list of involved molecules in each enriched categories are shown.
(XLS)
Ingenuity Pathway Analysis (IPA) of enriched categories in miRNAs targets analysis. Statistical significance and complete list of involved molecules in each enriched categories are shown.
(XLS)
Comparison of miRNAs dysregulations with previous studies. The modulation of dysregulated miRNAs in DM2 patients were compared to previously published data on miRNAs dysregulation in muscular diseases. Green and red colours indicate down- or up-regulation of miRNAs, respectively.
(XLS)
Acknowledgments
The authors thank Dr. Enrico Bugiardini, IRCCS Policlinico San Donato for his help in patient’s characterization.
Footnotes
Competing Interests: F. Martelli and M.C. Capogrossi are members of the PLoS One Editorial Board. This does not alter the authors’ adherence to all the PLoS ONE policies on sharing data and materials. All the other authors have declared that no competing interests exist.
Funding: This research was funded by Ministero della Salute, FMM-Fondazione Malattie Miotoniche, and Association Francaise contre les Myopathies. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Turner C, Hilton-Jones D. The myotonic dystrophies: diagnosis and management. J Neurol Neurosurg Psychiatry. 2010;81:358–367. doi: 10.1136/jnnp.2008.158261. [DOI] [PubMed] [Google Scholar]
- 2.Vihola A, Bassez G, Meola G, Zhang S, Haapasalo H, et al. Histopathological differences of myotonic dystrophy type 1 (DM1) and PROMM/DM2. Neurology. 2003;60:1854–1857. doi: 10.1212/01.wnl.0000065898.61358.09. [DOI] [PubMed] [Google Scholar]
- 3.Brook JD, McCurrach ME, Harley HG, Buckler AJ, Church D, et al. Molecular basis of myotonic dystrophy: expansion of a trinucleotide (CTG) repeat at the 3′ end of a transcript encoding a protein kinase family member. Cell. 1992;69:385. doi: 10.1016/0092-8674(92)90418-c. [DOI] [PubMed] [Google Scholar]
- 4.Fu YH, Pizzuti A, Fenwick RG, Jr, King J, Rajnarayan S, et al. An unstable triplet repeat in a gene related to myotonic muscular dystrophy. Science. 1992;255:1256–1258. doi: 10.1126/science.1546326. [DOI] [PubMed] [Google Scholar]
- 5.Mahadevan M, Tsilfidis C, Sabourin L, Shutler G, Amemiya C, et al. Myotonic dystrophy mutation: an unstable CTG repeat in the 3′ untranslated region of the gene. Science. 1992;255:1253–1255. doi: 10.1126/science.1546325. [DOI] [PubMed] [Google Scholar]
- 6.Liquori CL, Ricker K, Moseley ML, Jacobsen JF, Kress W, et al. Myotonic dystrophy type 2 caused by a CCTG expansion in intron 1 of ZNF9. Science. 2001;293:864–867. doi: 10.1126/science.1062125. [DOI] [PubMed] [Google Scholar]
- 7.Wojciechowska M, Krzyzosiak WJ. Cellular toxicity of expanded RNA repeats: focus on RNA foci. Hum Mol Genet. 2011;20:3811–3821. doi: 10.1093/hmg/ddr299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136:215–233. doi: 10.1016/j.cell.2009.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kloosterman WP, Plasterk RH. The diverse functions of microRNAs in animal development and disease. Dev Cell. 2006;11:441–450. doi: 10.1016/j.devcel.2006.09.009. [DOI] [PubMed] [Google Scholar]
- 10.Mendell JT. MicroRNAs: critical regulators of development, cellular physiology and malignancy. Cell Cycle. 2005;4:1179–1184. doi: 10.4161/cc.4.9.2032. [DOI] [PubMed] [Google Scholar]
- 11.Fasanaro P, Greco S, Ivan M, Capogrossi MC, Martelli F. microRNA: emerging therapeutic targets in acute ischemic diseases. Pharmacol Ther. 2010;125:92–104. doi: 10.1016/j.pharmthera.2009.10.003. [DOI] [PubMed] [Google Scholar]
- 12.Almeida MI, Reis RM, Calin GA. MicroRNA history: discovery, recent applications, and next frontiers. Mutat Res. 2011;717:1–8. doi: 10.1016/j.mrfmmm.2011.03.009. [DOI] [PubMed] [Google Scholar]
- 13.Greco S, De Simone M, Colussi C, Zaccagnini G, Fasanaro P, et al. Common micro-RNA signature in skeletal muscle damage and regeneration induced by Duchenne muscular dystrophy and acute ischemia. Faseb J. 2009;23:3335–3346. doi: 10.1096/fj.08-128579. [DOI] [PubMed] [Google Scholar]
- 14.Eisenberg I, Eran A, Nishino I, Moggio M, Lamperti C, et al. Distinctive patterns of microRNA expression in primary muscular disorders. Proc Natl Acad Sci U S A. 2007;104:17016–17021. doi: 10.1073/pnas.0708115104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cacchiarelli D, Incitti T, Martone J, Cesana M, Cazzella V, et al. miR-31 modulates dystrophin expression: new implications for Duchenne muscular dystrophy therapy. EMBO Rep. 2011;12:136–141. doi: 10.1038/embor.2010.208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cacchiarelli D, Martone J, Girardi E, Cesana M, Incitti T, et al. MicroRNAs involved in molecular circuitries relevant for the Duchenne muscular dystrophy pathogenesis are controlled by the dystrophin/nNOS pathway. Cell Metab. 2010;12:341–351. doi: 10.1016/j.cmet.2010.07.008. [DOI] [PubMed] [Google Scholar]
- 17.Gambardella S, Rinaldi F, Lepore SM, Viola A, Loro E, et al. Overexpression of microRNA-206 in the skeletal muscle from myotonic dystrophy type 1 patients. J Transl Med. 2010;8:48. doi: 10.1186/1479-5876-8-48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Perbellini R, Greco S, Sarra-Ferraris G, Cardani R, Capogrossi MC, et al. Dysregulation and cellular mislocalization of specific miRNAs in myotonic dystrophy type 1. Neuromuscul Disord. 2011;21:81–88. doi: 10.1016/j.nmd.2010.11.012. [DOI] [PubMed] [Google Scholar]
- 19.Rau F, Freyermuth F, Fugier C, Villemin JP, Fischer MC, et al. Misregulation of miR-1 processing is associated with heart defects in myotonic dystrophy. Nat Struct Mol Biol. 2011;18:840–845. doi: 10.1038/nsmb.2067. [DOI] [PubMed] [Google Scholar]
- 20.Udd B, Meola G, Krahe R, Wansink DG, Bassez G, et al. Myotonic dystrophy type 2 (DM2) and related disorders report of the 180th ENMC workshop including guidelines on diagnostics and management 3–5 December 2010, Naarden, The Netherlands. Neuromuscul Disord. 2011;21:443–450. doi: 10.1016/j.nmd.2011.03.013. [DOI] [PubMed] [Google Scholar]
- 21.Cardani R, Mancinelli E, Sansone V, Rotondo G, Meola G. Biomolecular identification of (CCTG)n mutation in myotonic dystrophy type 2 (DM2) by FISH on muscle biopsy. Eur J Histochem. 2004;48:437–442. doi: 10.4081/918. [DOI] [PubMed] [Google Scholar]
- 22.Vihola A, Bachinski LL, Sirito M, Olufemi SE, Hajibashi S, et al. Differences in aberrant expression and splicing of sarcomeric proteins in the myotonic dystrophies DM1 and DM2. Acta Neuropathol. 2010;119:465–479. doi: 10.1007/s00401-010-0637-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Greco S, Fasanaro P, Castelvecchio S, D’Alessandra Y, Arcelli D, et al. MicroRNA Dysregulation in Diabetic Ischemic Heart Failure Patients. Diabetes. 2012. [DOI] [PMC free article] [PubMed]
- 24.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 25.Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Roy Statist Soc Ser B. 1995;57:289–300. [Google Scholar]
- 26.Mestdagh P, Van Vlierberghe P, De Weer A, Muth D, Westermann F, et al. A novel and universal method for microRNA RT-qPCR data normalization. Genome Biol. 2009;10:R64. doi: 10.1186/gb-2009-10-6-r64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Le Brigand K, Robbe-Sermesant K, Mari B, Barbry P. MiRonTop: mining microRNAs targets across large scale gene expression studies. Bioinformatics. 2010;26:3131–3132. doi: 10.1093/bioinformatics/btq589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ge Y, Chen J. MicroRNAs in skeletal myogenesis. Cell Cycle. 2011;10:441–448. doi: 10.4161/cc.10.3.14710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.McCarthy JJ, Esser KA, Peterson CA, Dupont-Versteegden EE. Evidence of MyomiR network regulation of beta-myosin heavy chain gene expression during skeletal muscle atrophy. Physiol Genomics. 2009;39:219–226. doi: 10.1152/physiolgenomics.00042.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.van Rooij E, Quiat D, Johnson BA, Sutherland LB, Qi X, et al. A family of microRNAs encoded by myosin genes governs myosin expression and muscle performance. Dev Cell. 2009;17:662–673. doi: 10.1016/j.devcel.2009.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Erdreich LS, Lee ET. Use of relative operating characteristic analysis in epidemiology. A method for dealing with subjective judgement. Am J Epidemiol. 1981;114:649–662. doi: 10.1093/oxfordjournals.aje.a113236. [DOI] [PubMed] [Google Scholar]
- 32.Henderson AR. Assessing test accuracy and its clinical consequences: a primer for receiver operating characteristic curve analysis. Ann Clin Biochem 30 (Pt. 1993;6):521–539. doi: 10.1177/000456329303000601. [DOI] [PubMed] [Google Scholar]
- 33.Hanley JA, McNeil BJ. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology. 1982;143:29–36. doi: 10.1148/radiology.143.1.7063747. [DOI] [PubMed] [Google Scholar]
- 34.Day JW, Ricker K, Jacobsen JF, Rasmussen LJ, Dick KA, et al. Myotonic dystrophy type 2: molecular, diagnostic and clinical spectrum. Neurology. 2003;60:657–664. doi: 10.1212/01.wnl.0000054481.84978.f9. [DOI] [PubMed] [Google Scholar]
- 35.Schoser BG, Schneider-Gold C, Kress W, Goebel HH, Reilich P, et al. Muscle pathology in 57 patients with myotonic dystrophy type 2. Muscle Nerve. 2004;29:275–281. doi: 10.1002/mus.10545. [DOI] [PubMed] [Google Scholar]
- 36.Chekulaeva M, Filipowicz W. Mechanisms of miRNA-mediated post-transcriptional regulation in animal cells. Curr Opin Cell Biol. 2009;21:452–460. doi: 10.1016/j.ceb.2009.04.009. [DOI] [PubMed] [Google Scholar]
- 37.Hermeking H. The miR-34 family in cancer and apoptosis. Cell Death Differ. 2010;17:193–199. doi: 10.1038/cdd.2009.56. [DOI] [PubMed] [Google Scholar]
- 38.Cardinali B, Castellani L, Fasanaro P, Basso A, Alema S, et al. Microrna-221 and microrna-222 modulate differentiation and maturation of skeletal muscle cells. PLoS One. 2009;4:e7607. doi: 10.1371/journal.pone.0007607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ge Y, Sun Y, Chen J. IGF-II is regulated by microRNA-125b in skeletal myogenesis. J Cell Biol. 2011;192:69–81. doi: 10.1083/jcb.201007165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat Rev Genet. 2010;11:597–610. doi: 10.1038/nrg2843. [DOI] [PubMed] [Google Scholar]
- 41.Fardaei M, Larkin K, Brook JD, Hamshere MG. In vivo co-localisation of MBNL protein with DMPK expanded-repeat transcripts. Nucleic Acids Res. 2001;29:2766–2771. doi: 10.1093/nar/29.13.2766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Fardaei M, Rogers MT, Thorpe HM, Larkin K, Hamshere MG, et al. Three proteins, MBNL, MBLL and MBXL, co-localize in vivo with nuclear foci of expanded-repeat transcripts in DM1 and DM2 cells. Hum Mol Genet. 2002;11:805–814. doi: 10.1093/hmg/11.7.805. [DOI] [PubMed] [Google Scholar]
- 43.Mankodi A, Urbinati CR, Yuan QP, Moxley RT, Sansone V, et al. Muscleblind localizes to nuclear foci of aberrant RNA in myotonic dystrophy types 1 and 2. Hum Mol Genet. 2001;10:2165–2170. doi: 10.1093/hmg/10.19.2165. [DOI] [PubMed] [Google Scholar]
- 44.Serrano AL, Munoz-Canoves P. Regulation and dysregulation of fibrosis in skeletal muscle. Exp Cell Res. 2010;316:3050–3058. doi: 10.1016/j.yexcr.2010.05.035. [DOI] [PubMed] [Google Scholar]
- 45.Altomare DA, Guo K, Cheng JQ, Sonoda G, Walsh K, et al. Cloning, chromosomal localization and expression analysis of the mouse Akt2 oncogene. Oncogene. 1995;11:1055–1060. [PubMed] [Google Scholar]
- 46.Altomare DA, Lyons GE, Mitsuuchi Y, Cheng JQ, Testa JR. Akt2 mRNA is highly expressed in embryonic brown fat and the AKT2 kinase is activated by insulin. Oncogene. 1998;16:2407–2411. doi: 10.1038/sj.onc.1201750. [DOI] [PubMed] [Google Scholar]
- 47.Rotwein P, Wilson EM. Distinct actions of Akt1 and Akt2 in skeletal muscle differentiation. J Cell Physiol. 2009;219:503–511. doi: 10.1002/jcp.21692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Harper PS. Myotonic dystrophy. Oxford; New York: Oxford University Press. xiii, 106 p. p. 2009.
- 49.Botta A, Vallo L, Rinaldi F, Bonifazi E, Amati F, et al. Gene expression analysis in myotonic dystrophy: indications for a common molecular pathogenic pathway in DM1 and DM2. Gene Expr. 2007;13:339–351. doi: 10.3727/000000006781510705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Osborne RJ, Lin X, Welle S, Sobczak K, O’Rourke JR, et al. Transcriptional and post-transcriptional impact of toxic RNA in myotonic dystrophy. Hum Mol Genet. 2009;18:1471–1481. doi: 10.1093/hmg/ddp058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Burge JA, Hanna MG. Novel insights into the pathomechanisms of skeletal muscle channelopathies. Curr Neurol Neurosci Rep. 2012;12:62–69. doi: 10.1007/s11910-011-0238-3. [DOI] [PubMed] [Google Scholar]
- 52.Toscano A, Messina S, Campo GM, Di Leo R, Musumeci O, et al. Oxidative stress in myotonic dystrophy type 1. Free Radic Res. 2005;39:771–776. doi: 10.1080/10715760500138932. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Unsupervised hierarchical clustering of mRNA expression differentiating DM2 (n = 10) from controls (n = 10). Each row represents an mRNA, and each column represents an individual. A color code represents the relative intensity of the expression signal, with red indicating high expression and blue indicating low expression.
(PPT)
Transcriptome analysis identified 1,093 miRNA/mRNA interactions. Table represents the identified miRNAs/mRNA interactions, where green and red colours indicate down- or up-regulation of miRNA or mRNA compared to controls, respectively.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in transcriptome. Some important enriched pathways, functions and toxic functions and relative miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S8.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in transcriptome. Some important enriched pathways, functions and toxic functions and relative miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S8.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in transcriptome. Some important enriched pathways, functions and toxic functions and relative miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S8.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in miRNAs targets analysis. Some important enriched pathways, functions and toxic functions in miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S9.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in miRNAs targets analysis. Some important enriched pathways, functions and toxic functions in miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S9.
(XLS)
Ingenuity Pathway Analysis (IPA) of important pathways, functions and toxic functions enriched in miRNAs targets analysis. Some important enriched pathways, functions and toxic functions in miRNA/mRNA interactions are shown; green and red colours indicate down- or up-regulation of miRNA or mRNA, respectively. Complete lists of enriched categories can be found in Table S9.
(XLS)
Ingenuity Pathway Analysis (IPA) of enriched categories in transcriptome. Statistical significance and complete list of involved molecules in each enriched categories are shown.
(XLS)
Ingenuity Pathway Analysis (IPA) of enriched categories in miRNAs targets analysis. Statistical significance and complete list of involved molecules in each enriched categories are shown.
(XLS)
Comparison of miRNAs dysregulations with previous studies. The modulation of dysregulated miRNAs in DM2 patients were compared to previously published data on miRNAs dysregulation in muscular diseases. Green and red colours indicate down- or up-regulation of miRNAs, respectively.
(XLS)