

Abstract
Background
Patients with hematologic malignancies can be successfully treated with donor lymphocyte infusion after HLA-matched allogeneic hematopoietic stem cell transplantation. The effect of donor lymphocyte infusion is mediated by donor T cells recognizing minor histocompatibility antigens. T cells recognizing hematopoietic restricted minor histocompatibility antigens may induce selective graft-versus-leukemia reactivity, whereas broadly-expressed antigens may be targeted in graft-versus-host disease.
Design and Methods
We analyzed in detail CD8+ T-cell immunity in a patient with relapsed chronic myelogenous leukemia who responded to donor lymphocyte infusion with minimal graft-versus-host disease of the skin. CD8+ T-cell clones specific for 4 HLA-B*40:01 restricted minor histocompatibility antigens were isolated which were identified by screening a plasmid cDNA library and whole genome association scanning. Detailed T-cell reactivity and monitoring experiments were performed to estimate the clinical and therapeutic relevance of the novel antigens.
Results
Three antigens were demonstrated to be expressed on primary leukemic cells of various origins as well as subtypes of non-malignant hematopoietic cells, whereas one antigen was selectively recognized on malignant hematopoietic cells with antigen presenting cell phenotype. Skin derived fibroblasts were only recognized after pre-treatment with IFN-γ by two T-cell clones.
Conclusions
Our data show evidence for different roles of the HLA-B*40:01 restricted minor histocompatibility antigens in the onset and execution of the anti-tumor response. All antigens may have contributed to a graft-versus-leukemia effect, and one minor histocompatibility antigen (LB-SWAP70-1Q) has specific therapeutic value based on its in vivo immunodominance and strong presentation on leukemic cells of various origins, but absence of expression on cytokine-treated fibroblasts.
Keywords: HLA-matched transplantation, novel minor histocompatibility antigens, CD8 positive, graft-versus-leukemia effect, T-cell reactivity
Introduction
Patients with hematologic malignancies can be successfully treated with HLA-matched allogeneic hematopoietic stem cell transplantation (alloSCT).1 To reduce the development of graft-versus-host disease (GvHD), donor T cells can be depleted from the stem cell graft, and re-administered after alloSCT pre-emptively in case of persistent mixed chimerism or upon detection of relapsed or residual malignant cells.2 The risk and severity of GvHD after this 2-step procedure of T-cell depleted alloSCT and donor lymphocyte infusion (DLI) is reduced compared to non-T-cell depleted alloSCT.
The anti-tumor effect of DLI after HLA-matched alloSCT is mediated by donor T cells recognizing minor histocompatibility antigens (MiHA) in the context of HLA molecules.3–7 MiHA are peptides which are differentially expressed at the cell surface of patient and donor cells due to amino acid differences encoded by single nucleotide polymorphisms (SNP). Donor T cells recognizing MiHA with restricted expression to hematopoietic cells may selectively induce graft-versus-leukemia (GvL) reactivity, whereas donor T cells specific for MiHA with broad tissue distribution patterns may also mediate GvHD.
Although various HLA class I associated MiHA have been identified in the last decades,3–7 the number of MiHA with therapeutic relevance is still limited. We and others recently demonstrated that Whole Genome Association scanning (WGAs) is an efficient method for high throughput identification of MiHA,8–12 illustrated by the discovery of 10 novel MiHA as targets for CD8+ T cells in 2 different patients with leukemia who responded to DLI after HLA-matched alloSCT.12 Since these MiHA were all presented in HLA-A*02:01 and B*07:02, we selected 80 third-party EBV-B cell lines for co-expression of these HLA molecules, and genotyped all cell lines for more than one million SNPs. MiHA were identified by analysis of association between T-cell recognition of these EBV-B cells and individual SNP genotypes.
In this report, we identified 4 novel HLA-B*40:01 restricted MiHA as targets for CD8+ T cells in a patient who developed strong GvL reactivity with minimal GvHD of the skin after treatment with DLI for relapsed chronic myelogenous leukemia (CML) more than one year after HLA-matched alloSCT. One MiHA was identified by screening a plasmid cDNA expression library, whereas the other 3 MiHA were identified by WGAs. The data illustrate that our method of WGAs using a panel of SNP-genotyped EBV-B cells after retroviral transduction of the HLA restriction allele allows identification of MiHA. Detailed T-cell reactivity and monitoring experiments were performed to estimate the clinical and therapeutic relevance of the HLA-B*40:01 restricted MiHA. The data showed that all MiHA may have contributed to a GvL effect, and that one MiHA (LB-SWAP70-1Q) has specific therapeutic value based on its in vivo immunodominance and strong presentation on leukemic cells of various origins, but absence of expression on cytokine-treated fibroblasts.
Design and Methods
Study subject
A 47-year old woman with chronic phase CML was transplanted with hematopoietic stem cells from her HLA-identical sister (HLA-A*02:01, A*24:02, B*08:01, B*40:01, C*03:04, C*07:01).13 Nine months after alloSCT, BCR/ABL transcripts were detected, indicating a cytogenetic relapse. Retrospective molecular analysis of stored samples showed the presence of BCR/ABL transcripts at three and six months after alloSCT. In preparation for DLI, she was treated with 3×106 units of α-IFN daily s.c. Four weeks later the CML had progressed to a hematologic relapse, and 107 donor mononuclear cells per kg body weight were administered. After DLI she converted to 100% donor chimerism. The DLI was complicated by grade I GvHD of the skin and mouth for which no systemic immunosuppressive treatment was necessary. Currently, more than 12 years after DLI, she is still in good clinical condition without GvHD.
Patient and donor samples
Peripheral blood (PB) and bone marrow (BM) samples and skin biopsies from patients with CML, acute myeloid leukemia (AML), acute lymphoblastic leukemia (ALL), chronic lymphocytic leukemia (CLL) and multiple myeloma (MM), and PB and BM samples from healthy individuals were obtained after receiving approval from the LUMC Institutional Review Board and informed consent according to the Declaration of Helsinki. Mononuclear cells were isolated by Ficoll-Isopaque gradient centrifugation and cryopreserved.
Isolation and culture of T-cell clones
Peripheral blood mononuclear cells (PBMC) obtained six weeks after DLI were stimulated overnight with irradiated BM cells obtained from the patient prior to alloSCT, and single IFN-γ producing CD8+ T cells were isolated by flowcytometry after staining with PE-conjugated antibody against IFN-γ (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany). In addition, PBMC were stained with FITC-labeled anti-HLA-DR and APC-conjugated anti-CD8 antibodies (BD Biosciences, Breda, The Netherlands), and in vivo activated HLA-DR+ CD8+ T cells were single cell sorted by flowcytometry. T-cell clones were cultured as previously described.12
Cell culture
EBV-transformed B cells (EBV-B) and COS-7 cells were cultured in IMDM with 10% FCS. PHA-T blasts were generated by stimulating PBMC with IL-2 and PHA similar as described for T-cell clones. Primary fibroblasts (FB) and keratinocytes (KC) were cultured from skin biopsies in DMEM with low glucose (Cambrex) and 10% FCS supplemented with and without IFN-γ (100 IU/mL; Immukine; Boehringer Ingelheim, Alkmaar, The Netherlands) for four days. Proximal tubular epithelial cells (PTEC) cultured with and without IFN-γ (100 IU/mL) were kindly provided by Dr C van Kooten (Dept. Nephrology, LUMC, Leiden, The Netherlands).
Results
Isolation of CD8+ T-cell clones for minor histocompatibility antigens
A detailed analysis of CD8+ T-cell immunity was made in a patient who developed a strong GvL response with only limited GvHD after treatment with DLI for relapsed CML more than one year after alloSCT. In previous experiments, the GvL response in this patient was shown to coincide with a peak response in numbers of CD8+ T cells specific for hematopoietic restricted MiHA HA-1 and HA-2 between 4–8 weeks after DLI.13 To investigate whether, in addition to HA-1 and HA-2, other MiHA were targeted in this GvL response, single CD8+ T cells were isolated by flowcytometry from patient PBMC obtained six weeks after DLI. T cells were isolated based on specific production of IFN-γ after overnight stimulation with irradiated BM cells obtained from the patient prior to alloSCT,14 and based on expression of activation marker HLA-DR as previously described.12,15 CD8+ T-cell clones showing specific lysis and recognition of patient, but not donor, EBV-B cells in 4 h 51Cr-release assays and IFN-γ ELISA (Figure 1A) were selected and tested against a panel of EBV-B cells sharing one or more HLA class I alleles with the patient. The data demonstrated that the T-cell clones (ZRZ16, ZRZ25, 12A2 and 3H1) were specific for 4 unknown MiHA in HLA-B*40:01, as confirmed by retroviral transfer of the HLA restriction allele (Figure 1B).
Figure 1.

Isolation of CD8+ T-cell clones specific for HLA-B*40:01 restricted MiHA. (A) A number of selected CD8+ T-cell clones showed reactivity against patient, but not donor, EBV-B cells. Mean percentage of specific lysis in triplicate wells is shown at E:T ratios of 10:1 in 4 h 51Cr-release assays (upper) and of IFN-γ (ng/mL) in 50 μL culture supernatants of duplicate wells in ELISA (lower). Reactivity against patient and donor EBV-B cells by an allo-HLA-A*02:01 reactive T-cell clone is shown as control. (B) Selected CD8+ T-cell clones were specific for MiHA in HLA-B*40:01, as demonstrated by specific recognition of MiHA+ EBV-B cells after retroviral transfer of MP71-HLA-B*40:01-IRES-NGFR, but not mock MP71 vector, in IFN-γ ELISA. Specific production of IFN-γ (ng/mL) in 50 μL culture supernatants of duplicate wells in ELISA is shown.
Identification of minor histocompatibility antigens by screening a plasmid cDNA expression library
To identify the unknown MiHA, a plasmid cDNA expression library was constructed from patient derived EBV-B cells as previously described.16 COS-7 cells stably expressing HLA-B*40:01 were transfected with pools of 50 different cDNAs, and screened for T-cell recognition in IFN-γ ELISA using different mixtures of the MiHA specific CD8+ T-cell clones. By screening COS-7 cells, one positive cDNA pool was recognized by clone ZRZ16. In a second round of screening of single plasmids, a cDNA was isolated encompassing the 1102 – 2029 bp region of the gene encoding thyroid hormone receptor interactor protein 10 (TRIP10). By cloning different cDNA regions of the TRIP10 gene, the MiHA was shown to be encoded within the 1574–1810 bp region which contains the stop codon of the TRIP10 coding sequence at 1704 bp (Online Supplementary Figure S1A). This region was sequenced in patient and donor cDNA, and 3 SNP differences were demonstrated in the 3′ untranslated region (UTR) of the gene at 1761 bp (rs1049229), 1763 bp (rs1049230) and 1775 bp (rs1049232). Long 19–22 AA peptides comprising protein sequences in 3 different reading frames were synthesized, pulsed on donor EBV-B cells, and tested for T-cell recognition in IFN-γ ELISA (Online Supplementary Figure S1B). One of the long peptides (ARF1) comprising AA sequences in a reading frame different from the upstream encoded TRIP10 protein was recognized. The minimal epitope LB-TRIP10-1EPC was identified as a 9-mer peptide comprising 3 AA encoded by SNP differences (GEPQDLCTL), of which the E and C most significantly contributed to T-cell recognition, as determined by analysis of patient type peptides containing single AA of donor origin.
Identification of minor histocompatibility antigens by whole genome association scanning
In addition to screening a plasmid cDNA expression library, WGAs was performed to identify the unknown MiHA in HLA-B*40:01. We retrovirally transduced third party EBV-B cells, which we had previously genotyped for more than one million SNPs,12 with MP71-HLA-B*40:01-IRES-NGFR. A total number of 60 SNP-genotyped EBV-B cell lines expressing HLA-B*40:01 endogenously or after retroviral transduction were analyzed for recognition by T-cell clones ZRZ25, 12A2 and 3H1 in IFN-γ ELISA (Figure 2A). For each T-cell clone, SNP-genotyped EBV-B cells were separated into MiHA positive and negative groups using a threshold of recognition of 5-fold the background production of IFN-γ. The T-cell recognition patterns of EBV-B cells were subsequently analyzed for association with all individual SNP genotypes as previously described,12 and the level of matching between both patterns was calculated according to Fisher’s exact test using Plink statistical analysis package (http://pngu.mgh.harvard.edu/purcell/plink/version1.03). Table 1 shows the strongest associating SNPs (P<10−9) for all T-cell clones. Associating missense SNPs in the genes encoding SON DNA-binding protein (SON), SWAP switching B-cell complex 70 kDa subunit (SWAP70) and nucleoporin 133 kDa (NUP133) were measured for T-cell clones ZRZ25, 12A2, and 3H1, respectively.
Figure 2.

Identification of MiHA by whole genome association scanning. (A) A panel of approx. 60 SNP-genotyped EBV-B cells expressing HLA-B*40:01 endogenously or after retroviral transfer of MP71-HLA-B*40:01-IRES-NGFR with more than 20% of marker gene positive cells were tested for recognition by T-cell clones ZRZ25, 3H1 and 12A2 in IFN-γ ELISA. EBV-B cells were divided into MiHA positive and negative groups based on a threshold of 5-fold the background production of IFN-γ (horizontal lines). Mean release of IFN-γ (ng/mL) in 50 μL culture supernatants of duplicate wells is shown. (B) Peptides comprising patient (filled symbols) type amino acids with predicted binding to HLA-B*40:01 as well as donor type peptide variants (gray symbols) were pulsed on donor EBV-B cells and tested for T-cell recognition in IFN-γ ELISA. LB-TRIP10-1EPC (GEPQDLCTL) contains 3 patient type amino acids and has been identified by cDNA library screening. LB-SON-1R (SETKQRTVL), LB-SWAP70-1Q (MEQLEQLEL) and LB-NUP133-1R (SEDLILCRL) contain single patient type amino acids encoded by exon SNPs identified by WGAs based on significant association with T-cell recognition. Donor type peptides SETKQCTVL and MEQLEELEL encoded by the genes for SON and SWAP70, respectively, were not recognized by the T-cell clones, whereas donor type peptide SEDLILCQL encoded by the NUP133 gene was similarly recognized as LB-NUP133-1R. Mean production of IFN-γ (ng/mL) in 50 μL culture supernatants of duplicate wells at various peptide concentrations (μg/mL) is shown. (C) NUP133 genes were isolated from patient and donor derived cDNA and cloned into expression vector pcDNA-3. Hela cells stably expressing HLA-B*40:01 were transiently transfected with patient and donor derived NUP133 genes encoding the R and Q at position 406, respectively, and incubated with T-cell clone 3H1. Mean production of IFN-γ (ng/mL) in 50 μL culture supernatants of duplicate wells is shown.
Table 1.
Associating SNPs encoding candidate MiHA.
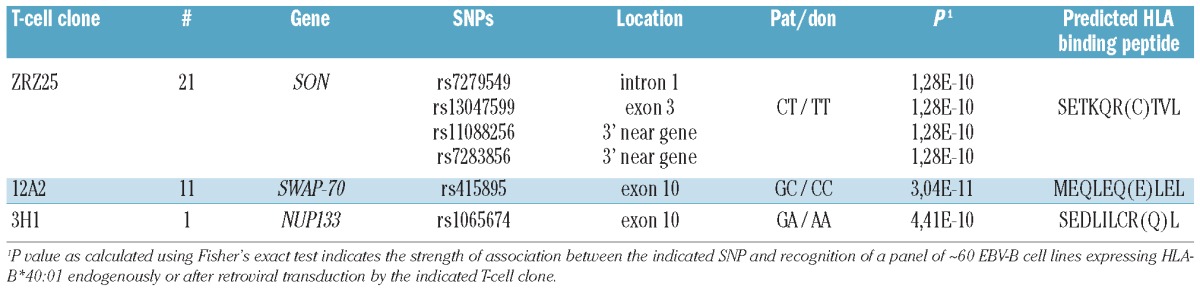
To identify the minimal MiHA epitopes, amino acid sequences comprising the associating missense SNPs were searched for peptides with predicted binding to HLA-B*40:01 using the algorithm of NetMHC (http://www.cbs.dtu.dk/services/NetMHC). For all associating missense SNPs, 9-mer peptides with predicted binding to HLA-B*40:01 were synthesized, pulsed on donor EBV-B cells, and shown to be recognized by the T-cell clones. This validated the identification of 3 HLA-B*40:01 restricted MiHA encoded by the genes for SON (LB-SON-1R; SETKQRTVL), SWAP70 (LB-SWAP70-1Q; MEQLEQLEL) and NUP133 (LB-NUP133-1R; SEDLILCRL) recognized by T-cell clones ZRZ25, 12A2 and 3H1, respectively (Figure 2B). Donor type peptide variants were not recognized, except for the donor peptide encoded by the NUP133 gene. We amplified the NUP133 genes from patient and donor derived cDNA by PCR, and cloned the genes into expression vector pcDNA-3. Cells transfected with the patient derived NUP133 gene encoding the R at position 406 were strongly recognized by clone 3H1, whereas cells transfected with the donor derived NUP133 gene encoding the Q at this position were not recognized (Figure 2C). These data demonstrated that differential recognition of patient and donor derived EBV-B cells by clone 3H1 is mediated by differences between LB-NUP133-1R and its donor variant in intracellular processing and presentation, and not by differential recognition by the T-cell receptor (TCR). In conclusion, by screening a plasmid cDNA expression library and WGAs, we identified 4 novel MiHA in HLA-B*40:01. The population frequencies of the HLA-B*40:01 associated MiHA are 96% (LB-TRIP10-1EPC; rs1049229, rs1049230, rs1049232), 48% (LB-SON-1R; rs13047599), 69% (LB-SWAP70-1Q; rs415895) and 48% (LB-NUP133-1R; rs1065674) calculated based on allele frequencies of 79%, 28%, 45% and 28% as reported in the SNP database (www.ncbi.nlm.nih.gov/snp), respectively.
T-cell recognition of non-malignant hematopoietic and non-hematopoietic cells
To investigate the cell type specificity in more detail, all MiHA specific CD8+ T-cell clones were tested for reactivity against patient derived EBV-B and PHA-T cells as well as primary B cells, T cells and monocytes isolated from patient PBMC pior to alloSCT by flowcytometry based on expression of CD19, CD3 and CD14, respectively (Figure 3A). The data showed that all MiHA specific CD8+ T-cell clones recognized specific types of primary hematopoietic cells, except for the T-cell clone for LB-TRIP10-1EPC, which showed reactivity against EBV-B cells, but failed to recognize all other non-malignant hematopoietic cells tested. The T-cell clone for LB-SWAP70-1Q showed clear recognition of hematopoietic cells of B-cell origin, whereas no or only weak reactivity was observed against T cells and monocytes.
Figure 3.

T-cell recognition of non-malignant hematopoietic and non-hematopoietic cells. CD8+ T-cell clones specific for LB-TRIP10-1EPC, LB-SON-1R, LB-SWAP70-1Q and LB-NUP133-1R were tested for recognition of non-malignant hematopoietic and non-hematopoietic cells in IFN-γ ELISA. (A) T-cell clones were tested against patient derived EBV-B cells and PHA-T blasts, as well as primary B cells, T cells and monocytes isolated from patient PBMC prior to alloSCT. (B) T-cell clones were tested against FB cultured from a skin biopsy obtained from the patient after alloSCT. FB were cultured with and without IFN-γ (100 IU/mL) for four days. Reactivity of T-cell clone 4D8 recognizing an unknown MiHA in HLA-B*08:01 is shown as control. Mean production of IFN-γ (ng/mL) in 50 μL culture supernatants of duplicate wells is shown.
In addition to hematopoietic cells, we tested T-cell recognition of non-hematopoietic FB cultured from a skin biopsy from the patient. FB were treated with IFN-γ to mimick the pro-inflammatory cytokine milieu of the early post transplantation period. In the absence of IFN-γ, FB were only recognized by a control T-cell clone (4D8) recognizing an unknown MiHA in HLA-B*08:01 (Figure 3B). After pre-treatment with IFN-γ, reactivity against FB was measured for the T-cell clones for LB-SON-1R and LB-NUP133-1R, whereas no or only weak recognition of cytokine-treated FB was observed for the T-cell clones for LB-TRIP10-1EPC and LB-SWAP70-1Q. We, therefore, concluded that LB-TRIP10-1EPC and LB-SWAP70-1Q are MiHA with potential therapeutic value based on lack of T-cell recognition of cytokine-treated FB.
T-cell recognition of leukemic cells of different origins
Since the HLA-B*40:01 restricted MiHA were identified as targets for CD8+ T cells in a patient with CML, we tested the capacity of the T-cell clones to recognize CD34+ CML cells in IFN-γ ELISA. CD34+ CML cells were isolated from BM samples from 3 HLA-B*40:01-positive patients, and tested for T-cell recognition directly after isolation as well as after in vitro culture with growth factors to generate CML cells with professional antigen presenting cell phenotype (CML-APC). All T-cell clones recognized freshly isolated CML cells as well as in vitro cultured CML-APC, except for the T-cell clone for LB-TRIP10-1EPC. This T-cell clone showed strong reactivity against CML-APC, whereas freshly isolated CML cells were weakly recognized or not recognized at all (Figure 4A). A similar difference in recognition was observed between monocytes and monocyte-derived dendritic cells (DC) (data not shown). The data demonstrated that LB-SON-1R, LB-SWAP70-1Q and LB-NUP133-1R are expressed at the surface of CML cells suggesting that these MiHA served as direct targets for CD8+ T cells in the GvL response after DLI. The data also showed that LB-TRIP10-1EPC is selectively expressed on professional APC, suggesting that T cells specific for this MiHA were induced in the onset of the immune response but did not contribute to direct lysis of leukemic cells in the execution phase of the response. In addition to CML cells, we analyzed T-cell recognition of ALL and AML cells of different subtypes (M0–M6). As shown in Figure 4B and C, the T-cell clone for LB-TRIP10-1EPC selectively recognized AML cells of monocytic (M4/M5) origin, whereas it failed to recognize ALL and AML cells of other subtypes. The T-cell clones for LB-SON-1R, LB-SWAP70-1Q and LB-NUP133-1R recognized HLA-B*40:01 and MiHA positive ALL and AML cells irrespective of their subtype origin. All T-cell clones were capable of mediating cytolysis of primary leukemic blasts in 51Cr-release assays (Online Supplementary Figure S2).
Figure 4.

T-cell recognition of leukemic cells of different origins. CD8+ T-cell clones specific for LB-TRIP10-1EPC, LB-SON-1R, LB-SWAP70-1Q and LB-NUP133-1R were tested for recognition of malignant hematopoietic cells in IFN-γ ELISA. (A) T-cell clones were tested against CD34+ CML cells directly after flowcytometric isolation from BM cells from 3 HLA-B*40:01 positive patients (gray bars) as well as after in vitro culture with growth factors to generate CML-APC (filled bars). (B) T-cell clones were tested against ALL cells from PB and BM samples from 5 HLA-B*40:01 positive patients directly after isolation by flowcytometry based on expression of CD19. (C) T-cell clones were tested against PB and BM samples from 10 HLA-B*40:01 positive patients with more than 40% CD33+ AML cells of different subtypes (M0–M6). Two HLA-B*40:01 negative AML-M4 and M5 samples were included as negative controls. The MiHA status of the samples is shown as +/+, +/− and −/−. Mean production of IFN-γ (ng/mL) in 50 μL culture supernatants of duplicate wells. Unpaired Student’s t-test showed a significant difference in T-cell recognition between MiHA positive (+/− or +/+) and MiHA negative (−/−) CML, ALL and AML samples for LB-SON-1R (P=5.6×10−5), LB-SWAP70-1Q (P=1.2×10−4), and LB-NUP133-1R (P=4.0×10−3). A significant difference for LB-TRIP10-1EPC (P=3.7×10−3) was demonstrated between MiHA positive (+/− and +/+) AML-M4/M5 and MiHA positive (+/+ and +/−) CML, ALL and AML samples of other subtypes.
We also investigated whether the MiHA specific T-cell clones were capable of lysing hematopoietic progenitor cells in colony forming cell (CFC) assays. For this purpose, a BM sample obtained from the patient from whom the T-cell clones were isolated was selected at the time of relapse of the CML after alloSCT prior to DLI. The T-cell clones for the HLA-B*40:01 restricted MiHA failed to lyse the CML progenitor cells, whereas substantial inhibition in numbers of CFU-GM and erythroid colonies was observed for the T-cell clone for HA-1 (Online Supplementary Figure S3A). The cytolytic capacity of the T-cell clone for LB-SWAP70-1Q was also investigated in a BM sample from a patient with transformed juvenile CML and BM samples from 2 healthy individuals. The T-cell clone failed to lyse hematopoietic progenitor cells from healthy individuals, whereas substantial inhibition in numbers of CFU-GM was observed when the T-cell clone for LB-SWAP70-1Q was pre-incubated with the juvenile CML progenitor cells (Online Supplementary Figure S3B). Outgrowth of erythroid colonies was not observed, compatible with the malignant transformed nature of the juvenile CML progenitor cells.
In conclusion, the data showed that LB-SON-1R, LB-SWAP70-1Q and LB-NUP133-1R are relevant targets in GvL reactivity against CML, AML and ALL, whereas LB-TRIP10-1EPC may only be relevant as a direct target for AML-M4/M5. Furthermore, the data suggest that the HLA-B*40:01 restricted MiHA may not have played a dominant role in sustaining long-term clinical remission of the patient with CML from whom the respective T-cell clones were isolated based on lack of cytolysis of the malignant hematopoietic progenitor cells. Finally, we conclude that LB-SWAP70-1Q is a MiHA with therapeutic potential based on strong presentation on leukemic cells of different origins and lack of T-cell recognition of cytokine treated FB.
Expression patterns of the genes encoding the HLA-B*40:01 restricted MiHA
The expression patterns of the TRIP10, SON, NUP133 and SWAP70 genes was investigated by microarray gene expression analysis using a panel of selected (malignant) hematopoietic and (cytokine treated) non-hematopoietic cell samples. For each malignancy (CML, AML, ALL, CLL and MM), malignant cells were isolated by flowcytometry from 5 different patients. Non-malignant hematopoietic cells (PBMC, B cells, T cells, monocytes, immature and mature DC, HSC) were isolated by flowcytometry from 3 different healthy individuals. Non-hematopoietic FB, KC and PTEC were derived from 3 healthy individuals. These were cultured with and without IFN-γ to mimick the pro-inflammatory cytokine milieu of the early post transplantation period. The data showed broad expression patterns and significant expression of all genes in cytokine treated non-hematopoietic cell types (Online Supplementary Figure S4). The data, therefore, demonstrate that the threshold for T-cell recognition of the HLA-B*40:01 restricted MiHA, and in particular of LB-SWAP70-1Q, the MiHA with selective GvL effect, is not only determined by gene expression, but rather by intracellular processing and presentation mechanisms and/or surface expression of adhesion and co-stimulatory molecules.
In vivo monitoring for MiHA specific T cells
We investigated the in vivo immunodominance of the HLA-B*40:01 restricted MiHA in the patient with CML from whom the T-cell clones were isolated. We previously measured a peak response in HA-1 and HA-2 tetramer positive T cells in this patient, and 2 PB samples obtained during this peak response at six and seven weeks after treatment with DLI were selected for tetramer analysis. The data showed a dominance for LB-SWAP70-1Q and LB-NUP133-1R in the immune response with 0.94% and 0.42% of tetramer positive cells within the PBMC population gated for negative expression of CD4, CD14, CD19 and CD56 at week 6 after DLI, respectively (Online Supplementary Figure S5). These frequencies exceeded the T-cell frequencies for HA-2 (0.33%) and HA-1 (0.13%) as measured at weeks 6 and 7, respectively. In addition to LB-SWAP70-1Q and LB-NUP133-1R, a low frequency of T cells specific for LB-SON-1R (0.03%) was detected at week 7 after DLI. T cells specific for LB-TRIP10-1EPC could not be detected at significant frequencies.
The in vivo immunogenicity of LB-SWAP70-1Q, the MiHA with selective GvL effect, was also evaluated in another HLA-B*40:01 and MiHA positive patient transplanted with an HLA-matched MiHA negative donor. The frequency of HLA-B*40:01 in our transplanted patient and donor population is 12.5%, which is higher than the 2–9% reported for various European countries (www.allelefrequencies.net). We screened 1,069 genomic DNA samples for the SNP encoding LB-SWAP70-1Q. A total number of 612 individuals were positive for LB-SWAP70-1Q, indicating a frequency of 57% in our patient and donor population. DNA samples from 398 patient-donor pairs included 297 (75%) transplantations with sibling donors and 101 (25%) transplantations with matched unrelated donors (MUD). Of the 398 patient-donor pairs, 56 LB-SWAP70-1Q positive patients were transplanted with MiHA negative donors, including 37 (12%) sibling and 19 (19%) MUD donors. Since HLA-B*40:01 is expressed in 12.5% of the human population, our data indicate a chance for mismatching for LB-SWAP70-1Q of 1.5% and 2.3% after sibling and MUD transplantation, respectively. Of the 56 LB-SWAP70-1Q positive patients transplanted with MiHA negative donors, 7 patient-donor pairs were positive for HLA-B*40:01. From 4 patients, PB and BM samples were available for T-cell monitoring. Three of these patients, however, were not treated with DLI and no obvious immune responses were ongoing at the available time points after alloSCT. One patient with CLL was successfully treated with DLI, and PB samples pre-DLI and six and eight weeks post-DLI were available for T-cell monitoring. These samples were screened for the presence of T cells for LB-SWAP70-1Q directly ex vivo as well as after one week of in vitro stimulation with peptide, but no tetramerpositive T cells could be detected (data not shown).
In conclusion, the data show that LB-SWAP70-1Q, the MiHA with selective GvL effect, was dominant in the immune response in the patient with CML from whom the T-cell clones were isolated. Since only one other patient could be monitored for T cells specific for LB-SWAP70-1Q, conclusions as to whether LB-SWAP70-1Q is immunogenic in a broader patient population cannot be drawn.
Discussion
Although an increasing number of MiHA in different HLA molecules have been identified in the last decades, the number of MiHA with therapeutic relevance is still limited.3–7 We previously demonstrated induction of CD8+ T cells specific for hematopoietic restricted MiHA HA-1 and HA-2 in a patient who developed a strong GvL response with limited GvHD of the skin after treatment with DLI for relapsed CML more than one year after HLA-matched alloSCT.13 We also isolated CD4+ T cells from this patient, and showed recognition of 6 HLA class II associated MiHA, of which 5 MiHA have been identified by screening a recombinant bacteria cDNA library.15,17 In this report, we demonstrated that in addition to HA-1 and HA-2, which are both presented by HLA-A*02:01, CD8+ T cells were induced against 4 MiHA in HLA-B*40:01 and one MiHA in HLA-B*08:01, illustrating the diversity of MiHA targeted in this GvL response. The HLA-B*40:01 restricted MiHA were identified by screening a plasmid cDNA expression library and WGAs, and detailed T-cell reactivity and monitoring experiments were performed to estimate the clinical and therapeutic relevance of the MiHA.
T cells specific for LB-SON-1R, LB-SWAP70-1Q and LB-NUP133-1R were shown to recognize CML, ALL and AML cells, illustrating the relevance of these MiHA as direct targets in GvL reactivity. LB-TRIP10-1EPC, however, was expressed on AML-M4/M5 of monocytic origin, but not on ALL and AML cells of other subtypes. Surface expression of LB-TRIP10-1EPC was also demonstrated for professional APC, including in vitro cultured CML-APC and monocyte derived DC, but expression could not be measured on freshly isolated CML progenitor cells and monocytes, nor on (specific types of) peripheral blood cells (B cells, T cells or monocytes). Based on these T-cell recognition data, we speculate that T cells specific for LB-TRIP10-1EPC were induced by professional leukemic APC at the onset of the immune response, but that these T cells did not mediate direct cytolysis of CML cells in the execution phase of the anti-tumor response. Although all T-cell clones for the HLA-B*40:01 restricted MiHA required similar amounts of peptide to be activated, surface expression of endogenous LB-TRIP10-1EPC in HLA-B*40:01 may be low, and the affinity of the T-cell receptor as expressed on the T-cell clone may not be sufficiently high to recognize endogenous LB-TRIP10-1EPC in the absence of high expression of costimulatory and/or adhesion molecules. Significant expression of the co-stimulatory molecule B7-2 (CD86) has been reported for AML-M4/M5,18 and may contribute to recognition of these leukemic subtypes by T cells specific for LB-TRIP10-1EPC.
Despite clear recognition and lysis of primary leukemic cells in IFN-γ ELISA and 51Cr-release assays, all T-cell clones for the HLA-B*40:01 restricted MiHA failed to lyse the malignant progenitor cells of the patient with CML from whom the T-cell clones were isolated in CFC assays. Our data, therefore, suggest that the HLA-B*40:01 restricted MiHA are relevant targets in GvL reactivity, but that they may not have played a dominant role in sustaining long-term clinical remission of the patient with CML.
All T cells specific for the novel HLA-B*40:01 restricted MiHA were tested for recognition of skin derived FB. These cells were cultured with and without IFN-γ to mimick the pro-inflammatory cytokine milieu of the early post transplantation period.19 In the absence of IFN-γ, all T cells failed to recognize FB. After treatment with IFN-γ, the only T cells which failed to recognize FB were specific for LB-SWAP70-1Q and LB-TRIP10-1EPC. Lack of T-cell reactivity against FB and normal hematopoietic progenitor cells cannot merely be explained by gene expression and, therefore, the immunological threshold for T-cell recognition of the HLA-B*40:01 restricted MiHA rather seems to be determined by intracellular antigen processing and presentation mechanisms and/or surface expression of adhesion and costimulatory molecules.
In vivo monitoring of the patient with CML from whom the T-cell clones were isolated demonstrated that 2 HLA-B*40:01 restricted MiHA (LB-SWAP70-1Q and LB-NUP133-1R) were dominant in the immune response, and exceeded the T-cell frequencies for the well-known hematopoietic restricted MiHA HA-1 and HA-2. One of these MiHA (LB-SWAP70-1Q) has been shown to be recognized on leukemic cells of various origins, but not on cytokine treated non-hematopoietic FB, illustrating the therapeutic potential of this MiHA as target for GvL reactivity. However, although the T-cell clone for LB-SWAP70-1Q as described in this study failed to recognize cytokine treated FB, various non-hematopoietic cells were shown to express the SWAP70 gene at significant levels. It can not, therefore, be completely excluded that in vitro or in vivo strategies to induce LB-SWAP70-1Q specific T cells with high avidities may have the potential to mediate GvHD due to reactivity towards non-hematopoietic tissues. Expression analysis also demonstrated overexpression of the SWAP70 gene in B cells, and its protein product has been described to exert relevant functions in nuclear and signal transduction events in activated B cells.20–22 The SWAP70 protein has been found to be a component of a complex that promotes recombination between DNA regions during heavy chain class switching,20,21 and it transduces signals from tyrosine kinase receptors to Rac in a Ras-independent manner.22
Recently, the first clinical study on adoptive transfer of MiHA specific CD8+ T cells to patients with acute leukemia who relapsed after alloSCT has been reported by Warren et al.23 The clinical results are encouraging since several patients achieved complete remissions, although at the price of significant GvHD. The MiHA specific CD8+ T cells as administered in the clinical study were selected for cytolytic activity against patient, but not donor derived EBV-B cells, and lack of reactivity to patient derived dermal FB, but only in the absence of cytokines. Our data, however, demonstrated that 2 HLA-B*40:01 restricted MiHA were presented on patient derived FB after pre-treatment with IFN-γ. We, therefore, support a more stringent selection strategy of MiHA as targets for T-cell therapies for selective stimulation of GvL reactivity without GvHD.
Acknowledgments
The authors would like to thank Menno van der Hoorn and Guido de Roo for flowcytometric sorting.
Footnotes
Funding: this work was supported by a grant from the European Union 6th Framework Programme, Allostem project n. 503319.
The online version of this article has a Supplementary Appendix.
Authorship and Disclosures
The information provided by the authors about contributions from persons listed as authors and in acknowledgments is available with the full text of this paper at www.haematologica.org.
Financial and other disclosures provided by the authors using the ICMJE (www.icmje.org) Uniform Format for Disclosure of Competing Interests are also available at www.haematologica.org.
References
- 1.Appelbaum FR. The current status of hematopoietic cell transplantation. Annu Rev Med. 2003;54:491–512. doi: 10.1146/annurev.med.54.101601.152456. [DOI] [PubMed] [Google Scholar]
- 2.Kolb HJ. Graft-versus-leukemia effects of transplantation and donor lymphocytes. Blood. 2008;112(12):4371–83. doi: 10.1182/blood-2008-03-077974. [DOI] [PubMed] [Google Scholar]
- 3.Falkenburg JHF, van de Corput L, Marijt WAF, Willemze R. Minor histocompatibility antigens in human stem cell transplantation. Exp Hematol. 2003;31(9):743–51. doi: 10.1016/s0301-472x(03)00190-5. [DOI] [PubMed] [Google Scholar]
- 4.Mullally A, Ritz J. Beyond HLA: the significance of genomic variation for allogeneic hematopoietic stem cell transplantation. Blood. 2007;109(4):1355–62. doi: 10.1182/blood-2006-06-030858. [DOI] [PubMed] [Google Scholar]
- 5.Feng X, Hui KM, Younes HM, Brickner AG. Targeting minor histocompatibility antigens in graft versus tumor or graft versus leukemia responses. Trends Immunol. 2008;29(12):624–32. doi: 10.1016/j.it.2008.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dierselhuis M, Goulmy E. The relevance of minor histocompatibility antigens in solid organ transplantation. Curr Opin Organ Transplant. 2009;14(4):419–25. doi: 10.1097/MOT.0b013e32832d399c. [DOI] [PubMed] [Google Scholar]
- 7.Bleakley M, Riddell SR. Exploiting T-cells specific for human minor histocompatibility antigens for therapy of leukemia. Immunol Cell Biol. 2011;89(3):396–407. doi: 10.1038/icb.2010.124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kawase T, Nannya Y, Torikai H, Yamamoto G, Onizuka M, Morishima S, et al. Identification of human minor histocompatibility antigens based on genetic association with highly parallel genotyping of pooled DNA. Blood. 2008;111(6):3286–94. doi: 10.1182/blood-2007-10-118950. [DOI] [PubMed] [Google Scholar]
- 9.Kamei M, Nannya Y, Torikai H, Kawase T, Taura K, Inamoto Y, et al. HapMap scanning of novel human minor histocompatibility antigens. Blood. 2009;113(21):5041–8. doi: 10.1182/blood-2008-07-171678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Spaapen RM, Lokhorst HM, van den Oudenalder K, Otterud BE, Dolstra H, Leppert MF, et al. Toward targeting B cell cancers with CD4+ CTLs: identification of a CD19-encoded minor histocompatibility antigen using a novel genome-wide analysis. J Exp Med. 2008;205(12):2863–72. doi: 10.1084/jem.20080713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Spaapen RM, de Kort RA, van den Ouden-alder K, van Elk M, Bloem AC, Lokhorst HM, et al. Rapid identification of clinical relevant minor histocompatibility antigens via genome-wide zygosity-genotype correlation analysis. Clin Cancer Res. 2009;15(23):7137–43. doi: 10.1158/1078-0432.CCR-09-1914. [DOI] [PubMed] [Google Scholar]
- 12.van Bergen CAM, Rutten CE, van der Meijden ED, van Luxemburg-Heijs SAP, Lurvink EGA, Houwing-Duistermaat JJ, et al. High-throughput characterization of 10 new minor histocompatibility antigens by whole genome association scanning. Cancer Res. 2010;70(22):9073–83. doi: 10.1158/0008-5472.CAN-10-1832. [DOI] [PubMed] [Google Scholar]
- 13.Marijt WAF, Heemskerk MHM, Kloosterboer FM, Goulmy E, Kester MGD, van der Hoorn MAWG, et al. Hematopoiesis-restricted minor histocompatibility antigens HA-1- or HA-2-specific T-cells can induce complete remissions of relapsed leukemia. Proc Natl Acad Sci USA. 2003;100(5):2742–7. doi: 10.1073/pnas.0530192100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kloosterboer FM, Luxemburg-Heijs SAP, van Soest RA, Barbui AM, van Egmond HM, Strijbosch MP, et al. Direct cloning of leukemia-reactive T-cells from patients treated with donor lymphocyte infusion shows a relative dominance of hematopoiesis-restricted minor histocompatibility antigen HA-1 and HA-2 specific T-cells. Leukemia. 2004;18(4):798–808. doi: 10.1038/sj.leu.2403297. [DOI] [PubMed] [Google Scholar]
- 15.Stumpf AN, van der Meijden ED, van Bergen CAM, Willemze R, Falkenburg JHF, Griffioen M. Identification of 4 new HLA-DR-restricted minor histocompatibility antigens as hematopoietic targets in antitumor immunity. Blood. 2009;114(17):3684–92. doi: 10.1182/blood-2009-03-208017. [DOI] [PubMed] [Google Scholar]
- 16.Slager EH, Honders MW, van der Meijden ED, Luxemburg-Heijs SAP, Kloosterboer FM, Kester MGD, et al. Identification of the angiogenic endothelial-cell growth factor-1/thymidine phosphorylase as a potential target for immunotherapy of cancer. Blood. 2006;107(12):4954–60. doi: 10.1182/blood-2005-09-3883. [DOI] [PubMed] [Google Scholar]
- 17.Griffioen M, van der Meijden ED, Slager EH, Honders MW, Rutten CE, van Luxemburg-Heijs SAP, et al. Identification of phosphatidylinositol 4-kinase type II beta as HLA class II-restricted target in graft versus leukemia reactivity. Proc Natl Acad Sci USA. 2008;105(10):3837–42. doi: 10.1073/pnas.0712250105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Maeda A, Yamamoto K, Yamashita K, Asagoe K, Nohgawa M, Kita K, et al. The expression of co-stimulatory molecules and their relationship to the prognosis of human acute myeloid leukaemia: poor prognosis of B7-2-positive leukaemia. Br J Haematol. 1998;102(5):1257–62. doi: 10.1046/j.1365-2141.1998.00901.x. [DOI] [PubMed] [Google Scholar]
- 19.Ferrara JL, Levine JE, Reddy P, Holler E. Graft-versus-host disease. Lancet. 2009;373(9674):1550–61. doi: 10.1016/S0140-6736(09)60237-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Borggrefe T, Wabl M, Akhmedov AT, Jessberger R. A B-cell-specific DNA recombination complex. J Biol Chem. 1998;273(27):17025–35. doi: 10.1074/jbc.273.27.17025. [DOI] [PubMed] [Google Scholar]
- 21.Borggrefe T, Masat L, Wabl M, Riwar B, Cattoretti G, Jessberger R. Cellular, intracellular, and developmental patterns of murine SWAP-70. Eur J Immunol. 1999;29(6):1812–22. doi: 10.1002/(SICI)1521-4141(199906)29:06<1812::AID-IMMU1812>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- 22.Shinohara M, Terada Y, Iwamatsu A, Shinohara A, Mochizuki N, Higuchi M, et al. SWAP-70 is a guanine-nucleotide-exchange factor that mediates signalling of membrane ruffling. Nature. 2002;416(6882):759–63. doi: 10.1038/416759a. [DOI] [PubMed] [Google Scholar]
- 23.Warren EH, Fujii N, Akatsuka Y, Chaney CN, Mito JK, Loeb KR, et al. Therapy of relapsed leukemia after allogeneic hematopoietic cell transplantation with T-cells specific for minor histocompatibility antigens. Blood. 2010;115(19):3869–78. doi: 10.1182/blood-2009-10-248997. [DOI] [PMC free article] [PubMed] [Google Scholar]