

Abstract
We addressed how restoration of forestry-drained peatlands affects CH4-cycling microbes. Despite similar community compositions, the abundance of methanogens and methanotrophs was lower in restored than in natural sites and correlated with CH4 emission. Poor establishment of methanogens may thus explain low CH4 emissions on restored peatlands even 10 to 12 years after restoration.
TEXT
Boreal peatlands have extensively been drained for forestry, agriculture, and peat mining. In Finland, for instance, more than half of the original peatland area has been lost (26). Restoration of drained peatlands involves raising the water table position and facilitating the return of peat-forming vegetation. With drainage, previously anoxic peat has been exposed to oxygen, which reduces CH4 production by methanogenic archaea and may limit CH4 consumption by aerobic methanotrophic alpha- and gammaproteobacteria through substrate depletion (9, 12, 16, 40). The objective of restoration is to revitalize the processes typical for pristine peatlands, including their function as carbon sinks and thus the microbial activity controlling carbon cycling. In restored mined peatlands, CH4 production and oxidation potentials have recovered and even exceeded those of natural sites in 4 to 30 years (1, 4, 10). On the other hand, the relatively few studies on CH4 emissions of restored drained peatlands indicate that although restoration increases the emissions (37), they may remain lower than on pristine mires during the first 2 to 3 years after restoration (18, 34). The low rates have been proposed to result from slow establishment of methanogens after prolonged aerobic conditions (34), but the effects of restoration on CH4-cycling microbial communities have not yet been demonstrated.
We addressed the response of methanogens, methanotrophs, and CH4 emission rates to restoration by comparing three natural peatlands and three restored forestry-drained peatlands in Finland (Table 1). The sites receive outflow water from the surrounding forested catchment (29, 36). Restoration of the drained peatlands was carried out 10 to 12 years before our study by filling in the drainage ditches with peat. The natural sites had vegetation typical of wet fen habitats, whereas species typical to drier and forestry-drained peatlands still occurred in the restored sites (see Fig. S1 in the supplemental material). In peat chemistry, site R2 differed from the other sites, because it had the lowest pH and the lowest Ca, K, and Mg levels (see Fig. S1).
Table 1.
Characteristics of peatland sites
| Site | Location | Yr constructed (age [yr] at sampling) | Water table (cm)a | Methane emission rate (mg m−2 day−1)a | pHb | Bulk density (g cm−3)b |
|---|---|---|---|---|---|---|
| Natural sites | ||||||
| N1, Asusuo | 60°26′N, 23°38′E | 1997 (10) | 2.3 ± 1.0 | 139 ± 67 | 4.6 ± 0.1 | 0.13 ± 0.02 |
| 4.7 ± 0.0 | 0.58 ± 0.13 | |||||
| N2, Kallioneva | 62°16′N, 23°48′E | 1998 (9) | −0.9 ± 0.6 | 707 ± 477 | 5.1 ± 0.0 | 0.08 ± 0.01 |
| 4.9 ± 0.1 | 0.07 ± 0.01 | |||||
| N3, Hirsikangas | 64°04′N, 26°40′E | 1999 (7) | 12.1 ± 0.7 | 227 ± 45 | 5.3 ± 0.1 | 0.12 ± 0.02 |
| 5.1 ± 0.1 | 0.15 ± 0.03 | |||||
| Restored sites | ||||||
| R1, Murtsuo | 61°01′N, 28°19′E | 1997 (11) | 3.6 ± 0.7 | 6.9 ± 3.7 | 5.1 ± 0.0 | 0.25 ± 0.02 |
| 5.2 ± 0.0 | 0.14 ± 0.00 | |||||
| R2, Konilamminsuo | 61°48′N, 24°17′E | 1995 (12) | −14.8 ± 2.1 | 3.0 ± 1.3 | 4.0 ± 0.1 | 0.08 ± 0.02 |
| 4.0 ± 0.1 | 0.08 ± 0.01 | |||||
| R3, Vanneskorpi | 61°51′N, 23°42′E | 1996–1997 (10–11) | −8.9 ± 1.1 | 10.8 ± 3.2 | 4.9 ± 0.1 | 0.35 ± 0.12 |
| 4.6 ± 0.1 | 0.15 ± 0.02 |
Growing season mean of eight sampling points ± SE. Water level values are negative below the peat surface.
For each site, the top row represents a depth of 0 to 7.5 cm and the bottom row represents a depth of 7.5 to 15 cm (mean ± SE).
Methane emission rates were measured at eight sampling points four or five times from May through September 2007 by the closed-chamber technique (11). The emission rates of the natural sites were at the higher end of the range reported for pristine boreal mires (Table 1) (25, 38). The restored sites had substantially lower CH4 emission rates (nested analysis of variance [ANOVA], P = 0.001). Even 10 to 12 years after restoration, their rates were more similar to those of drained, nutrient-poor, and forested peatlands (25, 28, 31). The emission rates in August 2007, when peat was sampled, were consistent with the rates for the whole growing season (data not shown).
Peat samples for microbial community analyses were taken from depths of 0 to 7.5 cm (aerobic surface peat for methanotrophs) and 7.5 to 15 cm (anaerobic layer for methanogens) next to the gas sampling positions (n = 8) at each site. The layer sampled for methanogens was below the water table level in all sites except R2 (Table 1). Samples were stored at −20°C. DNA was extracted with the Powersoil DNA isolation kit (Mo Bio Laboratories, Inc., Carlsbad, CA) with cell lysis in a FastPrep instrument (Thermo Savant, Holbrook, NY) with 3 rounds of 20 s at 4 m s−1. Quantitative PCR for methanogens with primers for methyl-coenzyme M reductase gene mcrA (30) (see the supplemental material) showed marginally larger mcrA copy numbers in natural than in restored sites (nested ANOVA, P = 0.065) (Fig. 1A). Methanogen abundance correlated with log-transformed CH4 emission rate (r = 0.884, P = 0.019) (Fig. 1A). Methanogen community composition was analyzed by mcrA terminal restriction fragment length polymorphism (T-RFLP) (40) (see Table S1 in the supplemental material). Communities between natural and restored sites differed only moderately (Fig. 2A) (analysis of similarity [ANOSIM] based on relative abundance of mcrA terminal restriction fragments [T-RFs], R = 0.20; presence/absence of T-RFs, R = 0.14; P < 0.001 in both) (see the supplemental material for details on statistical analyses). However, the communities of the restored sites showed significantly higher within-site heterogeneity (ANOVA of multivariate dispersion [3], P = 0.034) (Fig. 1). Community heterogeneity correlated negatively with CH4 emission rate (r = −0.967, P = 0.002) (Fig. 1C). For identification of T-RFs, we constructed and screened one clone library per site and sequenced a total of 52 clones (40). The main components of communities in both restored and natural sites were Methanoregulaceae (T-RFs 293, 470, 278, and 254 bp), Methanobacteriaceae (T-RFs 468, 275, and 251 bp), and Methanosarcinaceae (T-RFs 489 and 272 bp) (see Fig. S2 in the supplemental material). Several sequences were highly similar (>97% DNA identity) to those from peatlands and wetland soils (13–15, 23), but we also detected novel sequence types (<92% DNA identity to database sequences) associated with Methanobacteriales, Methanoregulaceae, and Methanocellales.
Fig 1.

Abundance (A and B) and within-site community heterogeneity (C and D) of methanogens (mcrA) and methanotrophs (pmoA) in six peatlands in relation to CH4 emission rate at the time of sampling. Black symbols represent natural sites, and gray symbols represent restored sites. Each data point represents the average of 7 or 8 field replicates ± standard error (SE). Microbial abundance was measured by quantitative PCR (gene copies cm−3 peat). Community heterogeneity was determined as multivariate dispersion based on T-RFLP or DGGE fingerprints. Note the logarithmic scale on the y axis in panels A, C, and D. The regression line is shown for data sets with significant relationships.
Fig 2.
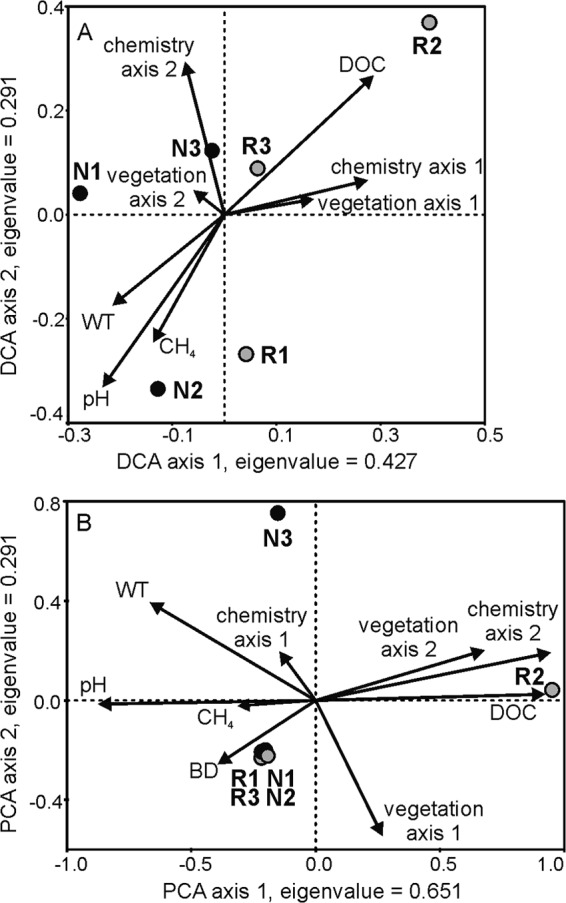
(A) Detrended correspondence analysis (DCA) of methanogen communities and (B) principal component analysis (PCA) of methanotroph communities in natural (N1 to N3) and restored (R1 to R3) peatlands (n = 7 to 8; only the site centroid is shown). The first two axes accounted for 42% (methanogens) and 87% (methanotrophs) of community variation. Communities were analyzed by mcrA T-RFLP and pmoA DGGE of peat samples. Arrows represent environmental variables. WT, water table level; DOC, dissolved organic carbon; CH4, mean emission rate for the growing season. Vegetation axis 1 refers to DCA axis 1 (wet-dry gradient) and vegetation axis 2 to DCA axis 2 (variation between restored sites related to plant species benefitting from disturbance) in Fig. S1 in the supplemental material. Chemistry axes 1 and 2 refer to PCA axes in Fig. S1C for methanogens and Fig. S1B for methanotrophs.
The small and heterogeneous methanogen communities at the restored sites could be an effect of drainage that restoration has not yet reversed, indicating incomplete restoration succession. Drainage restricts methanogenesis to anoxic microenvironments, and after restoration, proliferation of species selected by differing microenvironments could create large heterogeneity. The initially dominant species would shape the course of succession, similarly to models of vegetation succession (8, 39). Community heterogeneity could be further promoted by patchy vegetation formed with drainage (24). Studies on mined peatlands indicate that restored hydrology (1), but also recently formed peat or herbaceous vegetation, providing a good supply of labile carbon compounds, appears to be required to support pristine-like rates of methanogenesis (2, 4, 10, 21, 37). Since our restored sites did not have consistently low water table levels, but their vegetation still contained species typical of drained peatlands (see Fig. S1 in the supplemental material), the factor limiting the abundance and activity of CH4-cycling microbes could be peat quality (19, 32, 33). Resolution of this issue requires assessing the availability of labile carbon compounds for methanogenesis in restored peatlands.
Methanotroph communities were analyzed by denaturing gradient gel electrophoresis (DGGE) and band sequencing of pmoA coding for particulate methane monooxygenase using primers A189f and A621r with GC clamp (12, 20, 35). The reverse primer has been developed for amplification of type II methanotrophs when primer A682r yields no products (35). The restored and natural sites were not differentiated based on community composition (Fig. 2B) (ANOSIM R = 0.095, P = 0.027) or multivariate dispersion (Fig. 1D) (P = 0.730). Four out of six sites showed identical DGGE patterns of three bands, which were identified as type II methanotrophs grouping with Methylocystis spp. (see Fig. S3 and Table S2 in the supplemental material). Two bands unique to site R2 yielded sequences of the same sequence cluster, which contains several nearly identical pmoA sequences from wetlands and forest soils (5, 7, 20, 22, 27, 35, 40). No overall difference in methanotroph abundance was observed between natural and restored sites (nested ANOVA, P = 0.118), but the highest numbers occurred at natural sites (Fig. 1B). The pmoA copy numbers correlated with CH4 emission rate (r = 0.980, P = 0.001). Since we focused on pmoA-containing proteobacterial methanotrophs, our study cannot provide insight into methanotrophs lacking pmoA such as Methylocella. However, our results support the wide occurrence of Methylocystis in Sphagnum peat (5, 6, 17, 20). Drainage has been shown to affect distribution of Methylocystis (12, 40), but our results suggest that the community composition of methanotrophs may either respond to restoration in 10 years or may not always be affected by drainage. Methane oxidation has been observed to be less sensitive to peatland drainage than CH4 production (28), and similarly to our results, CH4 oxidation was suggested to be restricted by CH4 production in restored peat-mining sites (4).
In conclusion, our results suggest that unresponsiveness of methanogens to restoration rather than enhanced methane oxidation is behind the low CH4 emission of restored peatlands. Methanotroph abundance appeared to depend on CH4 supply. Methanogens typical of natural sites were present in restored peatlands, but clarification of the factors that limit their abundance and activity requires further studies. Nevertheless, our results indicate that time scales of over 10 years or additional restoration measures may be required for pristine-like microbial processes to return to forestry-drained peatlands.
Nucleotide sequence accession numbers.
The mcrA and pmoA sequences have been submitted to the EMBL database under accession numbers FN565431 to FN565482 (mcrA) and GQ468276 to GQ468281 (pmoA).
Supplementary Material
ACKNOWLEDGMENTS
This project was supported by the Academy of Finland (project 109816; H.J. was supported by project 133743 and E.-S.T. by project 218101) and the Maj and Tor Nessling Foundation.
We thank Minna Sinkkonen for technical assistance with microbial community analyses.
Footnotes
Published ahead of print 29 June 2012
Supplemental material for this article may be found at http://aem.asm.org/.
REFERENCES
- 1. Andersen R, Francez AJ, Rochefort L. 2006. The physicochemical and microbiological status of a restored bog in Québec: identification of relevant criteria to monitor success. Soil Biol. Biochem. 38: 1375–1387 [Google Scholar]
- 2. Andersen R, Grasset L, Thormann MN, Rochefort L, Francez A. 2010. Changes in microbial community structure and function following Sphagnum peatland restoration. Soil Biol. Biochem. 42: 291–301 [Google Scholar]
- 3. Anderson MJ. 2006. Distance-based tests for homogeneity of multivariate dispersions. Biometrics 62: 245–253 [DOI] [PubMed] [Google Scholar]
- 4. Basiliko N, Blodau C, Roehm C, Bengtson P, Moore TR. 2007. Regulation of decomposition and methane dynamics across natural, commercially mined, and restored northern peatlands. Ecosystems 10: 1148–1165 [Google Scholar]
- 5. Chen Y, et al. 2008. Diversity of the active methanotrophic community in acidic peatlands as assessed by mRNA and SIP-PLFA analyses. Environ. Microbiol. 10: 446–459 [DOI] [PubMed] [Google Scholar]
- 6. Dedysh SN, Derakshani M, Liesack W. 2001. Detection and enumeration of methanotrophs in acidic Sphagnum peat by 16S rRNA fluorescence in situ hybridization, including the use of newly developed oligonucleotide probes for Methylocella palustris. Appl. Environ. Microbiol. 67: 4850–4857 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Dumont MG, Radajewski SM, Miguez CB, McDonald IR, Murrell JC. 2006. Identification of a complete methane monooxygenase operon from soil by combining stable isotope probing and metagenomic analysis. Environ. Microbiol. 8: 1240–1250 [DOI] [PubMed] [Google Scholar]
- 8. Egler FE. 1954. Vegetation science concepts. I. Initial floristic composition, a factor in old-field vegetation development. Plant Ecol. 4: 412–417 [Google Scholar]
- 9. Freeman C, et al. 2002. Contrasted effects of simulated drought on the production and oxidation of methane in a mid-Wales wetland. Soil Biol. Biochem. 34: 61–67 [Google Scholar]
- 10. Glatzel S, Basiliko N, Moore T. 2004. Carbon dioxide and methane production potentials of peats from natural, harvested, and restored sites, eastern Québec, Canada. Wetlands 24: 261–267 [Google Scholar]
- 11. Hynninen A, et al. 2011. N2O fluxes from peatland buffer areas after high N loadings in five forested catchments in Finland. Wetlands 31: 1067–1077 [Google Scholar]
- 12. Jaatinen K, Tuittila ES, Laine J, Yrjälä K, Fritze H. 2005. Methane-oxidizing bacteria in a Finnish raised mire complex: effects of site fertility and drainage. Microb. Ecol. 50: 429–439 [DOI] [PubMed] [Google Scholar]
- 13. Juottonen H, et al. 2005. Methanogen communities and Bacteria along an ecohydrological gradient in a northern raised bog complex. Environ. Microbiol. 7: 1547–1557 [DOI] [PubMed] [Google Scholar]
- 14. Juottonen H, Galand PE, Yrjälä K. 2006. Detection of methanogenic archaea in peat: comparison of PCR primers targeting the mcrA gene. Res. Microbiol. 157: 914–921 [DOI] [PubMed] [Google Scholar]
- 15. Kemnitz D, Chin K, Bodelier P, Conrad R. 2004. Community analysis of methanogenic archaea within a riparian flooding gradient. Environ. Microbiol. 6: 449–461 [DOI] [PubMed] [Google Scholar]
- 16. Kettunen A, et al. 1999. Methane production and oxidation potentials in relation to water table fluctuations in two boreal mires. Soil Biol. Biochem. 31: 1741–1749 [Google Scholar]
- 17. Kip N, et al. 2010. Global prevalence of methane oxidation by symbiotic bacteria in peat-moss ecosystems. Nat. Geosci. 3: 617–621 [Google Scholar]
- 18. Komulainen VM, Nykänen H, Martikainen PJ, Laine J. 1998. Short-term effect of restoration on vegetation change and methane emissions from peatlands drained for forestry in southern Finland. Can. J. For. Res. 28: 402–411 [Google Scholar]
- 19. Laiho R. 2006. Decomposition in peatlands: reconciling seemingly contrasting results on the impacts of lowered water levels. Soil Biol. Biochem. 38: 2011–2024 [Google Scholar]
- 20. Larmola T, et al. 2010. The role of Sphagnum mosses in the methane cycling of a boreal mire. Ecology 91: 2356–2365 [DOI] [PubMed] [Google Scholar]
- 21. Marinier M, Glatzel S, Moore TR. 2004. The role of cotton-grass (Eriophorum vaginatum) in the exchange of CO2 and CH4 at two restored peatlands, eastern Canada. Ecoscience 11: 141–149 [Google Scholar]
- 22. McDonald IR, Murrell JC. 1997. The particulate methane monooxygenase gene pmoA and its use as a functional gene probe for methanotrophs. FEMS Microbiol. Lett. 156: 205–210 [DOI] [PubMed] [Google Scholar]
- 23. Metje M, Frenzel P. 2005. Effect of temperature on anaerobic ethanol oxidation and methanogenesis in acidic peat from a northern wetland. Appl. Environ. Microbiol. 71: 8191–8200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Minkkinen K, Laine J. 2006. Vegetation heterogeneity and ditches create spatial variability in methane fluxes from peatlands drained for forestry. Plant Soil 285: 289–304 [Google Scholar]
- 25. Nykänen H, Alm J, Silvola J, Tolonen K, Martikainen PJ. 1998. Methane fluxes on boreal peatlands of different fertility and the effect of long term experimental lowering of the water table on flux rates. Global Biogeochem. Cycles 12: 53–69 [Google Scholar]
- 26. Paavilainen E, Päivänen J. 1995. Peatland forestry—ecology and principles. Ecological studies: analysis and synthesis, vol 111 Springer-Verlag, Berlin, Germany [Google Scholar]
- 27. Radajewski S, et al. 2002. Identification of active methylotroph populations in an acidic forest soil by stable-isotope probing. Microbiology 148: 2331–2342 [DOI] [PubMed] [Google Scholar]
- 28. Roulet NT, Ash R, Quinton W, Moore T. 1993. Methane flux from drained northern peatlands: effect of a persistent water table lowering on flux. Global Biogeochem. Cycles 7: 749–769 [Google Scholar]
- 29. Sallantaus T, Vasander H, Laine J. 1998. Metsätalouden vesistöhaittojen torjuminen ojitetuista soista muodostettujen puskurivyöhykkeiden avulla. Suo 49: 125–133 [Google Scholar]
- 30. Steinberg LM, Regan JM. 2008. Phylogenetic comparison of the methanogenic communities from an acidic, oligotrophic fen and an anaerobic digester treating municipal wastewater sludge. Appl. Environ. Microbiol. 74: 6663–6671 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Strack M, Waddington JM, Tuittila ES. 2004. Effect of water table drawdown on northern peatland methane dynamics: implications for climate change. Global Biogeochem. Cycles 18: GB4003 doi:10.1029/2003GB002209 [Google Scholar]
- 32. Straková P, et al. 2010. Litter quality and its response to water level drawdown in boreal peatlands at plant species and community level. Plant Soil 335: 501–520 [Google Scholar]
- 33. Svensson BH, Sundh I. 1992. Factors affecting methane production in peat soils. Suo 43: 183–190 [Google Scholar]
- 34. Tuittila ES, et al. 2000. Methane dynamics of a restored cut-away peatland. Global Change Biol. 6: 569–581 [Google Scholar]
- 35. Tuomivirta TT, Yrjälä K, Fritze H. 2009. Quantitative PCR of pmoA using a novel reverse primer correlates with potential methane oxidation in Finnish fen. Res. Microbiol. 160: 751–756 [DOI] [PubMed] [Google Scholar]
- 36. Vikman A, et al. 2010. Nitrogen retention by peatland buffer areas at six forested catchments in southern and central Finland. Hydrobiologia 641: 171–183 [Google Scholar]
- 37. Waddington JM, Day SM. 2007. Methane emissions from a peatland following restoration. J. Geophys. Res. 112: G03018 doi:10.1029/2007JG000400 [Google Scholar]
- 38. Whalen SC. 2005. Biogeochemistry of methane exchange between natural wetlands and the atmosphere. Environ. Eng. Sci. 22: 73–94 [Google Scholar]
- 39. Wilson JB, Gitay H, Roxburgh SH, King WM, Tangney RS. 1992. Egler's concept of ‘initial floristic composition’ in succession: ecologists citing it don't agree what it means. Oikos 64: 591–593 [Google Scholar]
- 40. Yrjälä K, et al. 2011. CH4 production and oxidation processes in a boreal fen ecosystem after long-term water table drawdown. Global Change Biol. 17: 1311–1320 [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.