

Abstract
Immunomodulatory properties are attributed to apoptotic cells. These properties have been used to modulate allogeneic immune responses in experimental transplantation settings. In independent studies, apoptotic cell infusion has been shown to favor hematopoietic cell engraftment, to increase heart graft survival and to delay the lethal onset of graft-versus-host disease (GVHD). The goal of this review was to discuss how apoptotic cell infusion interferes with graft rejection or host rejection (i.e., GVHD) and to focus on the potential mediators or “perpetuators” involved in apoptotic cell-induced immunomodulation. Particular emphasis on apoptotic cell phagocytosis, TGF-β secretion and regulatory T cell induction was performed. Stimulating “naturally” immunosuppressive molecules (i.e., TGF-β) or immunomodulatory cells (“alternatively-activated” macrophages, certain DC subsets or regulatory T cells) in a physiological manner by using apoptotic cell infusion can be a promising way to induce tolerance.
Keywords: Animals; Apoptosis; immunology; Cell Transplantation; Immune Tolerance; immunology; Transcription, Genetic; genetics; Transforming Growth Factor beta; genetics; immunology
Keywords: apoptotic cells, dendritic cells, macrophages, TGF-β, regulatory T cells
Although the use of non specific immunosuppressive drugs has significantly reduced the incidence of acute graft rejection, the benefits of such therapies on chronic rejection and overall long-term graft survival are uncertain. Persistent excessive immunosuppression after therapeutic approaches of this type is associated with considerable long-term toxicity including increased incidence of cancers, infectious complications as well as metabolic diseases (e.g., diabetes, atherosclerosis, etc). Immunomodulatory approaches allowing for increased specificity are therefore needed. Cell-based therapy is one way of achieving tolerance to donor antigens. The use of donor apoptotic cell infusion has been proposed as an “alternative” cell-based therapy approach to induce tolerance. The mediators or “perpetuators” involved in the immunomodulatory properties of apoptotic cells are discussed in this review.
Immunomodulatory properties of apoptotic cells
Immunomodulatory properties are attributed to apoptotic cells. This is due to apoptotic cells themselves (1, 2) or to cells phagocyting apoptotic cells (3, 4). Professional phagocytes, such as macrophages (3) or certain dendritic cells (DC) subsets (4), quickly capture apoptotic bodies, thereby preventing both the release of harmful products from dying cells turning to necrosis and the associated inflammatory responses. Several mechanisms have been described to explain the immunomodulatory effects of apoptotic cells (5). Engulfment of apoptotic cells inhibits proinflammatory cytokine production by blocking NF-κB activation in phagocyting cells (6). Apoptotic cell uptake by phagocytes blocks also their production of the type 1 cytokine IL-12 (7) as well as the Th17-related cytokines, IL-17 and IL-23 (8). Moreover, apoptotic neutrophils and T cells may prevent inflammatory cell recruitment by sequestering chemokines (2). In addition, apoptotic cell death is frequently associated with an immunomodulatory milieu that may neutralize immune cells present in the environment of dying cells. Such milieu consists mainly of anti-inflammatory factors like TGF-β, IL-10, IL-1 receptor antagonist, prostaglandin-E2 and hepatocyte growth factor (3, 9–11). These factors can be released by phagocytes (3) and/or by apoptotic cells themselves (1). However, the immunomodulatory effects induced by apoptotic cells can be modulated or influenced by several factors (5, 12) [Figure 1] such as cell death-inducing agents (13), soluble factors that bridge apoptotic cells to phagocytes (5, 14), or ligand receptor interactions involved in apoptotic cell uptake (5, 15). Rapidly after the initiation of the apoptotic process, changes including alterations in glycosylation, charge and surface lipids occur at the cell surface of dying cells. The best-characterized modification is the loss of phospholipid asymmetry and the exposure of phosphatidylserine on the outer surface. Recently, exposure of calcireticulin after tumor cell death has been shown to trigger an efficient immune response against dying tumors (16). The route of administration and the site where apoptotic cells die may trigger inflammation and vigorous immune responses. This can lead to immune responses directed against apoptotic cell-derived antigens (Fig. 1), as mentioned above (13, 16). Thus, in order to use apoptotic cells as a cell therapy product, all of the latter factors, including the route of administration, have to be defined. Intended to induce immunomodulation, the tolerogenic intravenous (i.v.) route will be requisite.
FIGURE 1. Factors influencing the immune response to apoptotic cells.
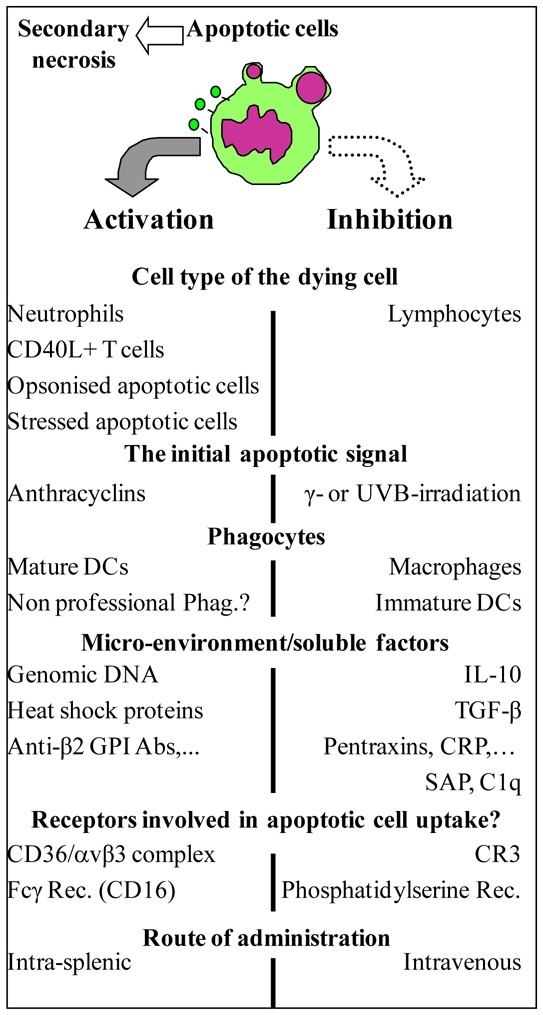
Several factors may influence the immune response to apoptotic cells. Factors involved in the activation of the immune response are listed on the left hand side, while those implicated in its down-regulation are mentioned on the right hand side. This figure is modified from Ref. 12 and updated with recent references (see text). Abbreviations: DC: dendritic cell; Phag: phagocytes; Rec: receptor; CR3: Complement receptor 3; CRP: C reactive protein; SAP: serum amyloid P component; C1q: one of the first fractions of the complement.
Use of apoptotic cell infusion in transplantation settings
Physiological mechanisms that maintain immune homeostasis and self-tolerance have been exploited for a long time to induce tolerance to allografts. This is why the inhibitory effects of apoptotic cell infusion have been used to modulate alloreactive responses in experimental transplantation models (17–21) [Table 1]. In hematopoietic cell (HC) transplantation, i.v. apoptotic cell infusion favors allogeneic HC engraftment (17), delays the onset of lethal acute GVHD (18) and prevents the anti-donor humoral response (19). In solid organ transplantation models, donor apoptotic cell infusion increases heart graft survival (20, 21) whatever the timing of infusion: 7 days before heart transplantation (19) (as previously done with donor specific blood transfusion) or 7 days after (20). The timing of apoptotic cell infusion does not seem to be a critical factor in the interference of alloreactive immune responses. The next questions to address are how i.v. apoptotic cell infusion may interfere with alloreactive responses and by which mediators (and mechanisms) the immunomodulatory effects are propagated?
TABLE 1.
Immunomodulatory effects of intravenous apoptotic cell infusion in transplantation settings: an overview.
| Experimental transplantation models: | Hematopoietic cell engraftment | Rat heart transplantation | Graft-versus-Host Disease | Mouse heart transplantation |
|---|---|---|---|---|
| Dependent on the origin of infused apoptotic cells: | no (17,19) | yes (only donor apoptotic cells prolong survival, [20]) | yesa | yes (only donor apoptotic cells prolong survival, [21]) |
| Timing of apoptotic cell infusion: | with the bone marrow graft (17–19) | 7 days before heart transplant (20) | with the bone marrow graft (18) | 7 days after transplant (21) |
| Required phagocytes: | splenic host macrophages (18) | host macrophages (20) | ND | splenic host CD8α+ DC (21) |
| Required cytokines: | TGF-β (18, 19) | ND | ND | intragraft TGF-β and IL-10 (21) |
| Treg induction: | yes, but not critical in the graft facilitating effects (18) | ND | yes, Treg participate to delay the onset of lethal GVHD (18) | suspected, Foxp3+ Treg present in the graft (21) |
| Inhibition of anti-donor immunization: | yes (19) in a TGF-β-dependent manner | ND | ND | yes (suppress systemic anti-donor response; IFN-γ inhibition, [21]) |
Kleinclauss F, de Carvalho Bittencourt M, Saas P, unpublished results
ND, not determined.
Mediators involved in the immunomodulatory effects of apoptotic cells
All three studies performed in transplantation settings (18, 20, 21) shared the requirement for apoptotic cell phagocytosis in a limited area (the spleen), but the nature of phagocytes differed. Two studies (18, 20) identified recipient macrophages as critical for the apoptotic cell infusion-induced immunomodulatory effects. This was attested by macrophage depletion before apoptotic cell infusion using gadolinium chloride (20) or clodronate-loaded liposomes (18). Conversely, host CD8α+ DC allowed prolonged graft survival when apoptotic cells were infused after heart transplantation, as shown by adoptive transfer experiments (21). However, a direct role of DC was excluded in Ref.18 where despite efficient depletion of CD11chigh DC in a transgenic mouse model, apoptotic cell infusion still favored HC engraftment. The read out used (depletion (18, 20) versus adoptive transfer, Ref.21) may influence the result. Nevertheless, it makes sense to identify different phagocytes depending on the transplantation model used: heart (21) versus HC (18). Macrophages and DC diverge in their antigen presentation capacity due to their lysosomal protease content (22). Macrophages rapidly degrade internalized antigens without presenting them, while DC exhibit limited lysosomal degradation capacity (22) favoring donor alloantigen presentation. A restricted donor-specific effect related to donor apoptotic cell phagocytosis by DC and subsequent donor alloantigen presentation may account for prolonged heart graft survival (21). Donor apoptotic cell-loaded DC may migrate to the thymus to perform alloantigen-specific T cell deletion. In HC transplantation, apoptotic cell phagocytosis by macrophages (18) may lead to a non-specific anti-inflammatory effect associated with TGF-β production (19, see below), independent of donor alloantigen presentation. Indeed, HC engraftment is favored whatever the origin (donor/recipient/other) of apoptotic cells (17, 19). The subsequent anti-inflammatory effect related to TGF-β secretion may neutralize host immune cells to allow HC engraftment and reduce the cytokine storm responsible for GVHD initiation.
To reconcile data obtained from different transplantation models (17, 19), an alternative explanation would be that both macrophages and DC are necessary for apoptotic cell-induced engraftment. In this scenario, anti-inflammatory cytokines produced by macrophages after phagocytosis (i.e., TGF-β) may induce DC to present alloantigens from apoptotic cells in an inefficient manner. This may explain why donor apoptotic cell-loaded DC infusion after heart transplantation induces an initial proliferation of alloreactive T cells followed by their deletion (21). This hypothesis is currently under investigation.
Another mediator involved in the immunomodulatory effects of apoptotic cell infusion in transplantation settings is TGF-β. Increased TGF-β transcripts were detected on day 3 post-transplantation in the spleen of mice that had received apoptotic spleen cells simultaneously with bone marrow (BM) grafts (Figure 2). Neutralization of this cytokine abrogates the effects of i.v. apoptotic cell infusion on BM engraftment (19) and the prevention of anti-donor alloimmunization (19). Indeed, i.v. apoptotic cell infusion given simultaneously with hematopoietic allografts prevents anti-BM donor immunization even in mice rejecting their graft (19). This alteration of allogeneic humoral responses was also observed when donor apoptotic cells were infused after heart transplantation (21). TGF-β is implicated in the conversion of naive CD4+ CD25− T cells into regulatory T cells (Treg) (23); analysis of Treg after apoptotic cell infusion was thus performed. Foxp3+ Treg were identified as propagating the effect of i.v. apoptotic cell infusion both in heart (21) and BM (18) transplantation models. In the BM transplantation model, the involvement of TGF-β secretion and splenic macrophages in the induction of Treg was demonstrated (18). Indeed, TGF-β neutralization or clodroante-loaded liposome infusion blocked Treg expansion induced by i.v. apoptotic cell infusion (18). In this model, a direct link between macrophage and TGF-β secretion has yet to be demonstrated. Interleukin-10 dependent-Treg induction after infusion of syngeneic splenocytes rendered apoptotic by photopheresis has also been reported (24). Interestingly, Treg induction after random blood product transfusion has been reported previously (25) and nonspecific immunomodulatory effects of preoperative blood product transfusion has been proposed to be related to the presence of apoptotic leukocytes in stored blood products (26). Treg induction after blood cell apoptosis –a frequent phenomenon, mimicked by i.v. infusion of apoptotic cells– may constitute an additional mechanism to prevent autoimmunity that could be used in transplantation.
FIGURE 2. Early increase of TGF-β mRNA transcript after intravenous donor apoptotic cell infusion simultaneously with bone marrow graft.
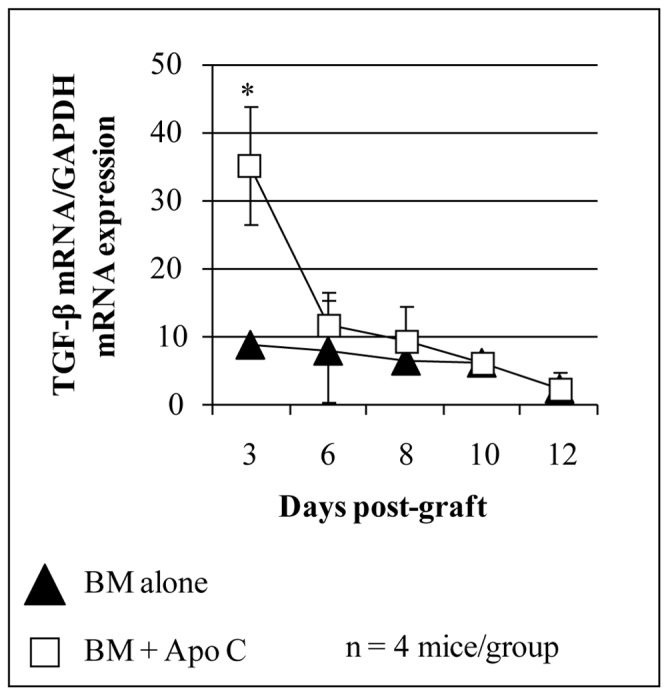
Total RNA was extracted from spleen of BALB/c mice grafted with 106 FVB BM (black triangles) or with 5x106 FVB apoptotic splenocytes (Apo C) plus 106 FVB BM (open square) using RNA extraction kit (Qiagen, Valencia, CA) and reverse transcribed using random hexamers and M-MLV reverse transcriptase (Life Technologies, Rockville, MD) to use as template for quantitative real time RT-PCR (QRT-PCR). QRT-PCR reactions were performed as described (18). Primers and dual labeled fluorescent probes were designed using Primer Express® software (Applied Biosystems, Forster City, CA) to be specific for RNA but not for genomic DNA. QRT-PCR primer pairs and related probes were as follow (sense, antisense and probe respectively). TGF-β : 5′-GCTCTTGTGACAGCAAAGATAACAA-3′, 5′-GGTCGCCCCGACGTTT-3′ and 5′-FAM-CACGTGGAAATCAACGGGATCAGCC-TAMRA-3′; Primer pairs for GAPDH (used as endogenous reference)were already described (18). The relative quantity of each unknown sample was determined automatically with the iCycler iQ® software (Bio-Rad Laboratories, Marnes-la-Coquette, France), using the threshold cycle (Ct). Data were expressed as normalized TGF-β1 expression, which was obtained by dividing the relative quantity of TGF-β1 mRNA for each sample by the relative quantity of GAPDH mRNA of the same sample. *P<0.05.
Conclusion
In conclusion, we propose infusing apoptotic cells to interfere with graft rejection or host rejection (GVHD). This approach permits to stimulate “naturally” immunosuppressive molecule production (i.e., TGF-β) or to activate immunomodulatory cells (such as Treg and particular DC or macrophage subsets) in a physiological manner. Exploiting such mechanisms may facilitate the induction of immunological tolerance to transplants. However, before the possible application of such therapy to clinical transplantation, several obstacles need to be overcome, including the interactions of such an approach with immunosuppressive regimens. In the future, i.v. donor apoptotic cells may become a well-defined cell therapy product and a substitute for the blood products formerly used in transfusion practice to prolong graft survival. Indeed, similitude exists between donor-specific blood transfusion and i.v. apoptotic leukocyte infusion including induction of Treg (25) or of TGF-β production (27). Some biological effects of transfusion may be related to the presence of apoptotic leukocytes in blood products (26). This idea of using the immunomodulatory properties of apoptotic cells in cell-based therapy is moving forward, since such an approach has also been proposed to increase cardiac muscle repair (28).
Acknowledgments
We thank Pierre Tiberghien, Estelle Seilles and Christophe Borg for their support Chantal Ferniot, Dominique Paris, Aliette Marandin-Decock for their expertise in animal care and management, and Moyra Barbier for excellent editorial assistance.
ABBREVIATIONS
- BM
Bone marrow
- DC
Dendritic cell
- Foxp3
Forkhead-box transcription factor p3
- GVHD
Graft-versus-host disease
- HC
Hematopoietic cell
- i.v
Intravenous
- TGF-β
Transforming growth factor-β
- Treg
Regulatory T cells
Footnotes
This work was supported by grants from the Association pour la Recherche sur le Cancer (#3851), the Association pour la Recherche en Transfusion (ART) and the Etablissement Français du Sang (#2004.10). FB received financial support from the Conseil Régional de Franche-Comté and the INSERM. SKP received financial support from the Fondation Transplantation (#ET-051226).
References
- 1.Chen W, Frank ME, Jin W, Wahl SM. TGF-beta released by apoptotic T cells contributes to an immunosuppressive milieu. Immunity. 2001;14:715. doi: 10.1016/s1074-7613(01)00147-9. [DOI] [PubMed] [Google Scholar]
- 2.Ariel A, Fredman G, Sun YP, et al. Apoptotic neutrophils and T cells sequester chemokines during immune response resolution through modulation of CCR5 expression. Nat Immunol. 2006;7:1209. doi: 10.1038/ni1392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Fadok VA, Bratton DL, Konowal A, Freed PW, Westcott JY, Henson PM. Related Macrophages that have ingested apoptotic cells in vitro inhibit proinflammatory cytokine production through autocrine/paracrine mechanisms involving TGF-beta, PGE2, and PAF. J Clin Invest. 1998;101:890. doi: 10.1172/JCI1112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Steinman RM, Turley S, Mellman I, Inaba K. The induction of tolerance by dendritic cells that have captured apoptotic cells. J Exp Med. 2000;191:411. doi: 10.1084/jem.191.3.411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Savill J, Dransfield I, Gregory C, Haslett C. A blast from the past: clearance of apoptotic cells regulates immune responses. Nat Rev Immunol. 2002;2:965. doi: 10.1038/nri957. [DOI] [PubMed] [Google Scholar]
- 6.Sen P, Wallet MA, Yi Z, et al. Apoptotic cells induce Mer tyrosine kinase-dependent blockade of NF-{kappa}B activation in dendritic cells. Blood. 2007;109:653. doi: 10.1182/blood-2006-04-017368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kim S, Elkon KB, Ma X. Transcriptional suppression of interleukin-12 gene expression following phagocytosis of apoptotic cells. Immunity. 2004;21:643. doi: 10.1016/j.immuni.2004.09.009. [DOI] [PubMed] [Google Scholar]
- 8.Stark MA, Huo Y, Burcin TL, Morris MA, Olson TS, Ley K. Phagocytosis of apoptotic neutrophils regulates granulopoiesis via IL-23 and IL-17. Immunity. 2005;22:285. doi: 10.1016/j.immuni.2005.01.011. [DOI] [PubMed] [Google Scholar]
- 9.Voll RE, Herrmann M, Roth EA, Stach C, Kalden JR, Girkontaite I. Immunosuppressive effects of apoptotic cells. Nature. 1997;390:350–1. doi: 10.1038/37022. [DOI] [PubMed] [Google Scholar]
- 10.Craciun LI, DiGiambattista M, Schandene L, Laub R, Goldman M, Dupont E. Anti-inflammatory effects of UV-irradiated lymphocytes: induction of IL-1Ra upon phagocytosis by monocyte/macrophages. Clin Immunol. 2005;114:320. doi: 10.1016/j.clim.2004.11.006. [DOI] [PubMed] [Google Scholar]
- 11.Morimoto K, Amano H, Sonoda F, et al. Alveolar macrophages that phagocytose apoptotic neutrophils produce hepatocyte growth factor during bacterial pneumonia in mice. Am J Respir Cell Mol Biol. 2001;24:608. doi: 10.1165/ajrcmb.24.5.4292. [DOI] [PubMed] [Google Scholar]
- 12.Kleinclauss F, Perruche S, Cahn JY, Tiberghien P, Saas P. Administration of donor apoptotic cells: an alternative cell-based therapy to induce tolerance? Transplantation. 2003;75(9 Suppl):43S. doi: 10.1097/01.TP.0000067951.90241.54. [DOI] [PubMed] [Google Scholar]
- 13.Casares N, Pequignot MO, Tesniere A, et al. Caspase-dependent immunogenicity of doxorubicin-induced tumor cell death. J Exp Med. 2005;202:1691. doi: 10.1084/jem.20050915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Krispin A, Bledi Y, Atallah M, et al. Apoptotic cell thrombospondin-1 and heparin binding domain lead to dendritic cell phagocytic and tolerizing states. Blood. 2006 doi: 10.1182/blood-2006-03-013334. In press. [DOI] [PubMed] [Google Scholar]
- 15.Skoberne M, Somersan S, Almodovar W, et al. The apoptotic-cell receptor CR3, but not alphavbeta5, is a regulator of human dendritic-cell immunostimulatory function. Blood. 2006;108:947. doi: 10.1182/blood-2005-12-4812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Obeid M, Tesniere A, Ghiringhelli F, et al. Calreticulin exposure dictates the immunogenicity of cancer cell death. Nat Med. 2007;13:54. doi: 10.1038/nm1523. [DOI] [PubMed] [Google Scholar]
- 17.Bittencourt MC, Perruche S, Contassot E, et al. Intravenous injection of apoptotic leukocytes enhances bone marrow engraftment across major histocompatibility barriers. Blood. 2001;98:224. doi: 10.1182/blood.v98.1.224. [DOI] [PubMed] [Google Scholar]
- 18.Kleinclauss F, Perruche S, Masson E, et al. Intravenous apoptotic spleen cell infusion induces a TGF-beta-dependent regulatory T-cell expansion. Cell Death Differ. 2006;13:41. doi: 10.1038/sj.cdd.4401699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Perruche S, Kleinclauss F, de Carvalho Bittencourt M, Paris D, Tiberghien P, Saas P. Intravenous infusion of apoptotic cells simultaneously with allogeneic hematopoietic grafts alters anti-donor humoral immune responses. Am J Transplant. 2004;4:1361. doi: 10.1111/j.1600-6143.2004.00509.x. [DOI] [PubMed] [Google Scholar]
- 20.Sun E, Gao Y, Chen J, et al. Allograft tolerance induced by donor apoptotic lymphocytes requires phagocytosis in the recipient. Cell Death Differ. 2004;11:1258. doi: 10.1038/sj.cdd.4401500. [DOI] [PubMed] [Google Scholar]
- 21.Wang Z, Larregina AT, Shufesky WJ, et al. Use of the inhibitory effect of apoptotic cells on dendritic cells for graft survival via T-cell deletion and regulatory T cells. Am J Transplant. 2006;6:1297. doi: 10.1111/j.1600-6143.2006.01308.x. [DOI] [PubMed] [Google Scholar]
- 22.Delamarre L, Pack M, Chang H, Mellman I, Trombetta ES. Differential lysosomal proteolysis in antigen-presenting cells determines antigen fate. Science. 2005;307:1630. doi: 10.1126/science.1108003. [DOI] [PubMed] [Google Scholar]
- 23.Chen W, Jin W, Hardegen N, et al. Conversion of peripheral CD4+CD25− naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J Exp Med. 2003;198:1875. doi: 10.1084/jem.20030152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Maeda A, Schwarz A, Kernebeck K, et al. Intravenous infusion of syngeneic apoptotic cells by photopheresis induces antigen-specific regulatory T cells. J Immunol. 2005;174:5968. doi: 10.4049/jimmunol.174.10.5968. [DOI] [PubMed] [Google Scholar]
- 25.Bushell A, Karim M, Kingsley CI, Wood KJ. Pretransplant blood transfusion without additional immunotherapy generates CD25+CD4+ regulatory T cells: a potential explanation for the blood-transfusion effect. Transplantation. 2003;76:449. doi: 10.1097/01.TP.0000083043.84630.99. [DOI] [PubMed] [Google Scholar]
- 26.Snyder EL, Kuter DJ. Apoptosis in transfusion medicine: of death and dying--is that all there is? Transfusion. 2000;40:135. doi: 10.1046/j.1537-2995.2000.40020135.x. [DOI] [PubMed] [Google Scholar]
- 27.Josien R, Douillard P, Guillot C, et al. A critical role for transforming growth factor-beta in donor transfusion-induced allograft tolerance. J Clin Invest. 1998;102:1920. doi: 10.1172/JCI4221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Thum T, Bauersachs J, Poole-Wilson PA, Volk HD, Anker SD. The dying stem cell hypothesis: immune modulation as a novel mechanism for progenitor cell therapy in cardiac muscle. J Am Coll Cardiol. 2005;46:1799. doi: 10.1016/j.jacc.2005.07.053. [DOI] [PubMed] [Google Scholar]